
Picrotoxin Delineates Different Transport Configurations for Malate and γ Aminobutyric Acid through TaALMT1

 ,
, 
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
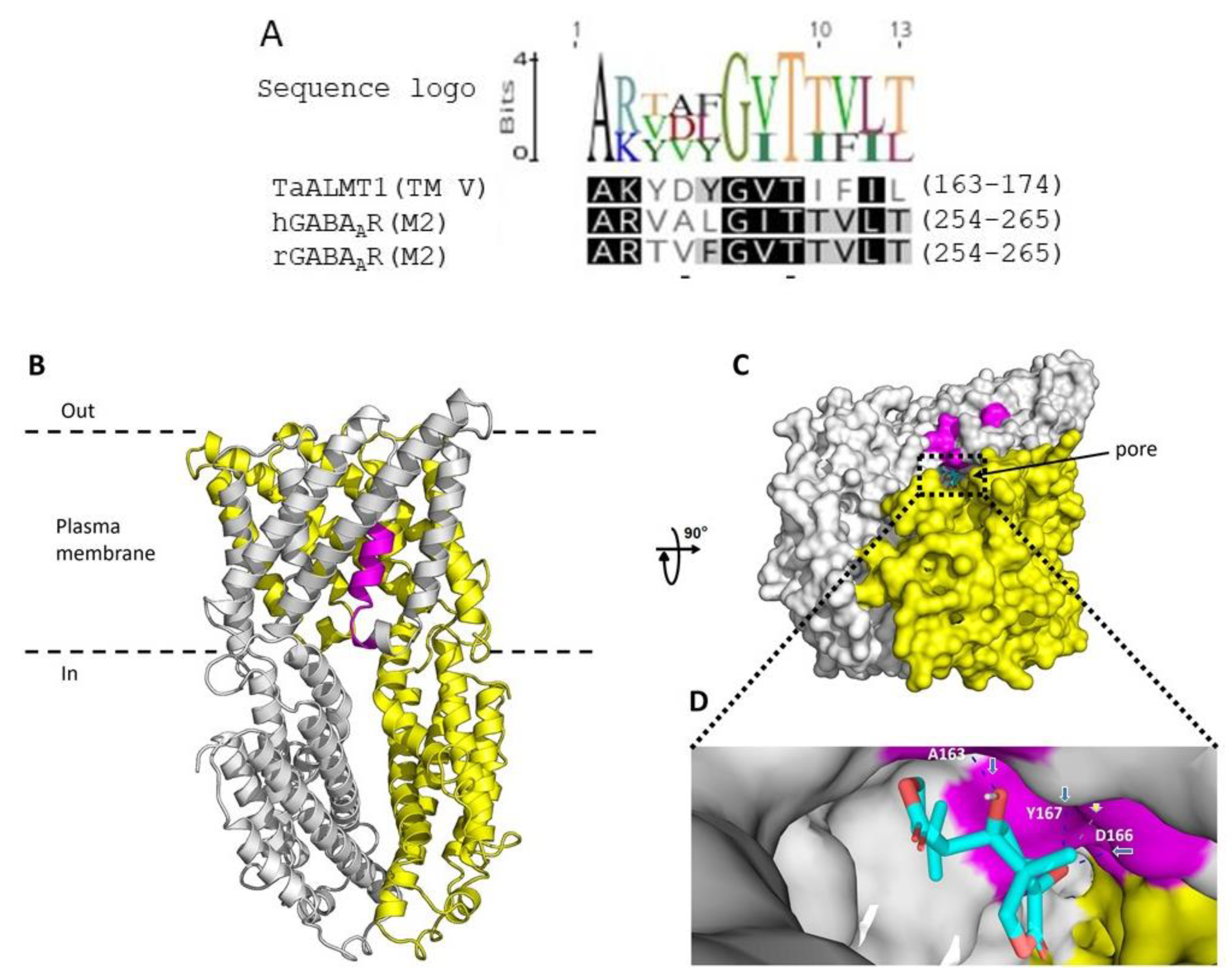{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. cRNA Synthesis
2.3. Voltage-Clamp Electrophysiology
2.4. Patch Clamping
2.5. Malate Efflux Measurements
2.6. GABA Efflux Meaurements
2.7. Tobacco BY2 Malate Efflux and GABA Efflux
2.8. Logo and Homology Modelling
2.9. Statistics
3. Results
3.1. Wheat Root Malate and GABA Efflux
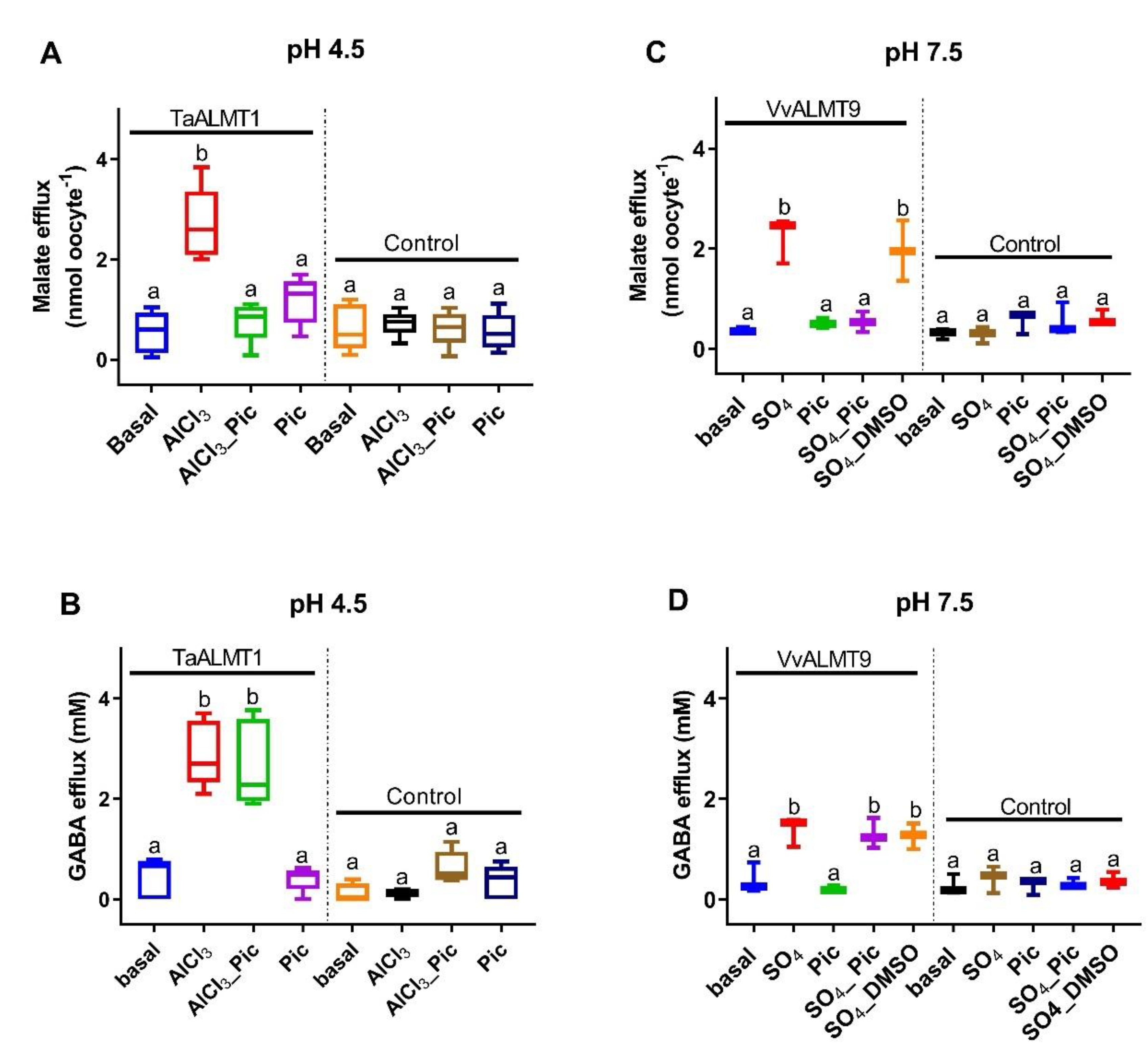
3.2. Malate and GABA Efflux in Xenopus Laevis Oocytes Expressing ALMT
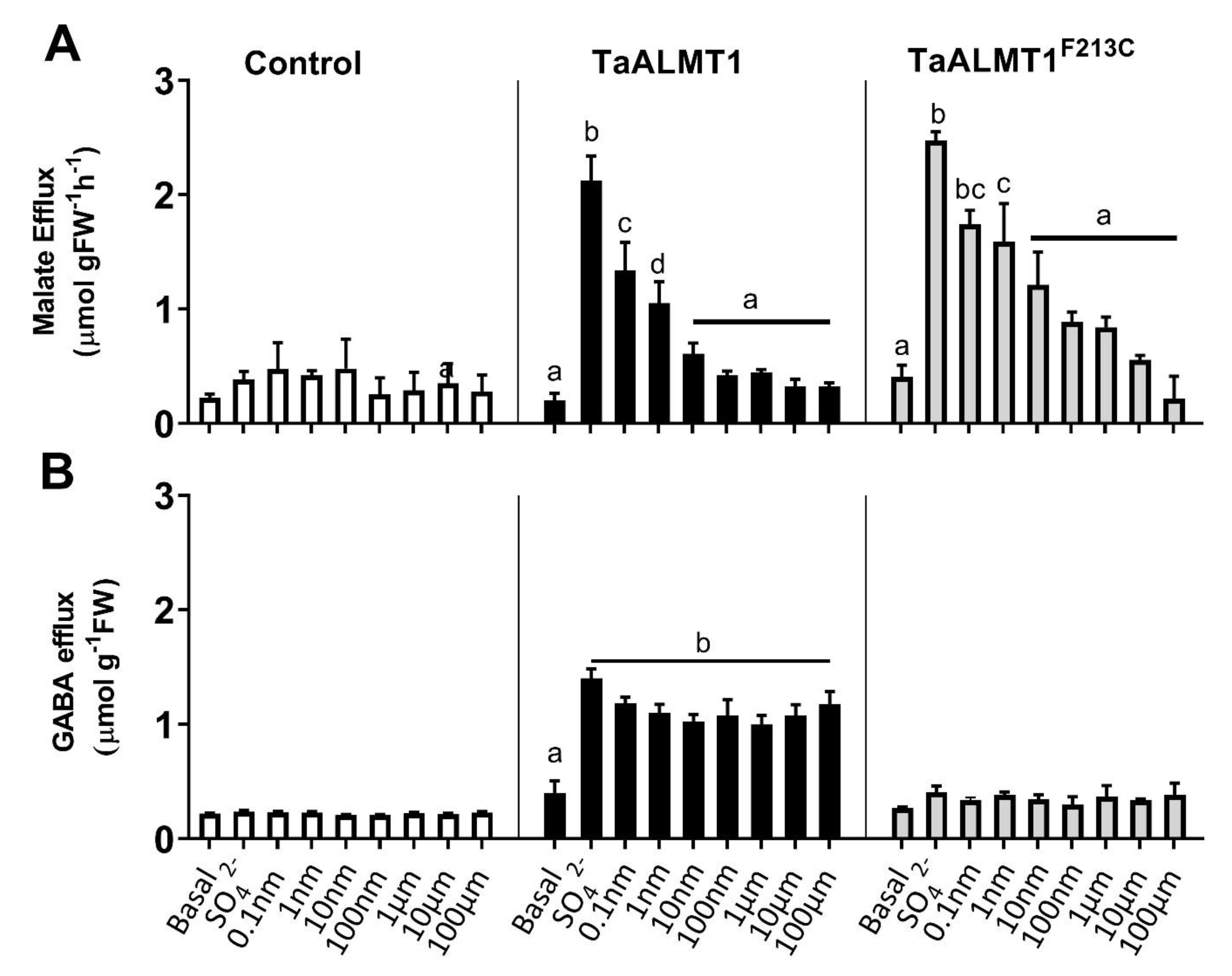
3.3. Malate and GABA Efflux in Tobacco BY2 Expressing TaALMT1 and TaALMT1F213C
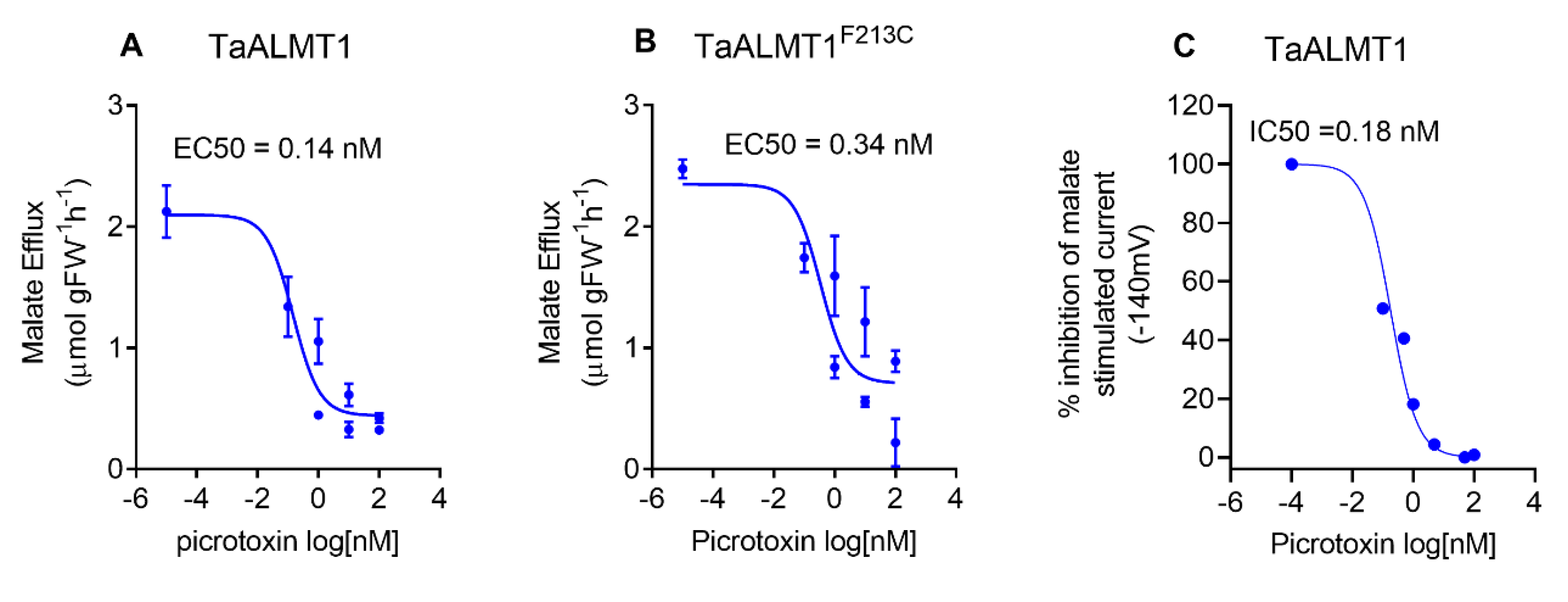
3.4. Picrotoxin Inhibits Malate Efflux with High Affinity
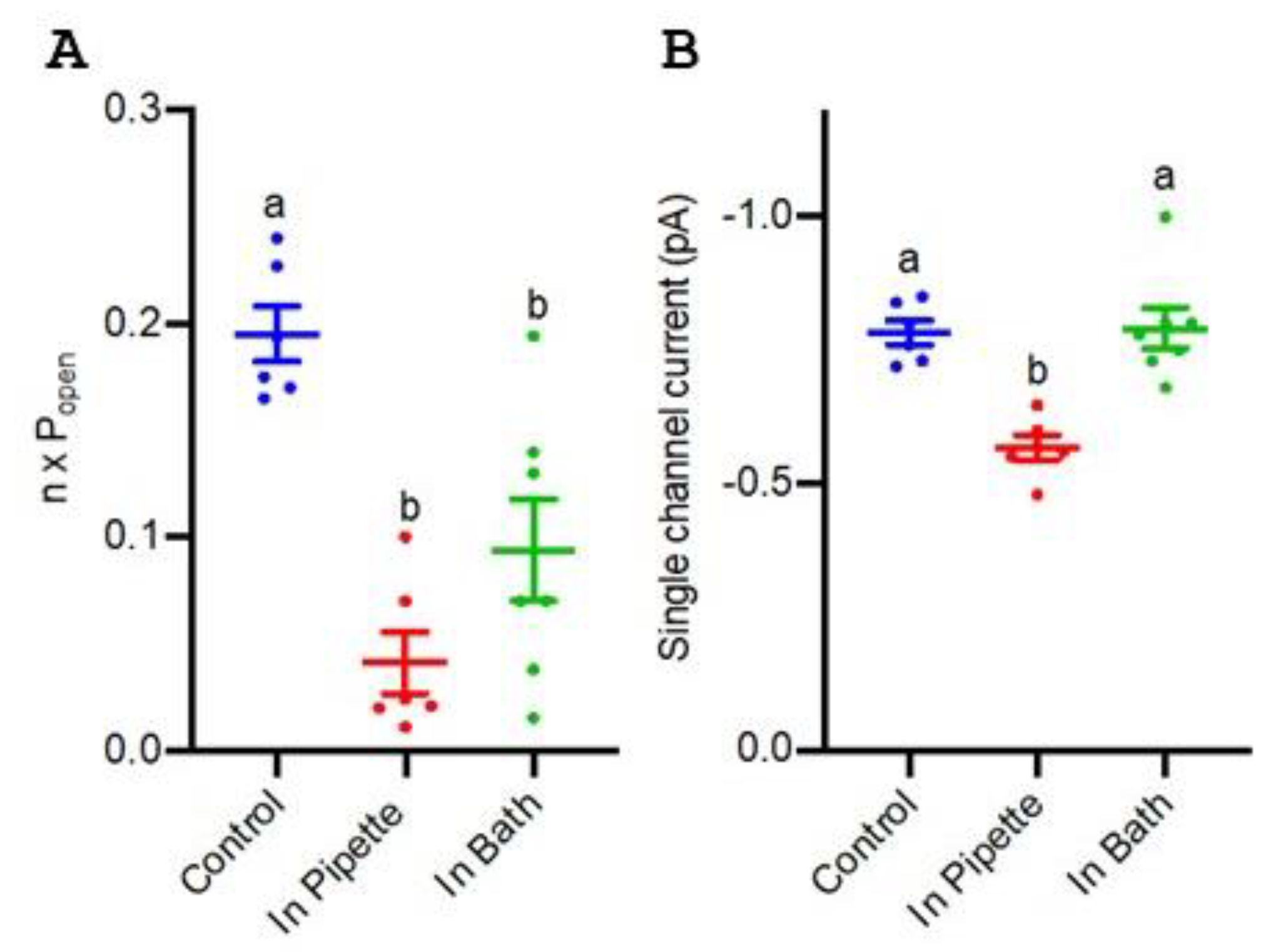
3.5. Blockade of TaALMT1 Single Channels by Picrotoxin
3.6. Probable Binding Site for Picrotoxin on ALMT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kinnersley, A.M.; Turano, F.J. Gamma aminobutyric acid (GABA) and plant responses to stress. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Palanivelu, R.; Brass, L.; Edlund, A.F.; Preuss, D. Pollen tube growth and guidance is regulated by POP2, an Arabidopsis gene that controls GABA levels. Cell 2003, 114, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.R. Does GABA Act as a Signal in Plants? Hints from Molecular Studies: Hints from Molecular Studies. Plant Signal. Behav. 2007, 2, 408–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, S.A.; Tyerman, S.D.; Xu, B.; Bose, J.; Kaur, S.; Conn, V.; Domingos, P.; Ullah, S.; Wege, S.; Shabala, S.; et al. GABA signalling modulates plant growth by directly regulating the activity of plant-specific anion transporters. Nat. Commun. 2015, 6, 7879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. Gamma-Aminobutyric acid (GABA) signalling in plants. Cell. Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, A.; Dennis, P.G.; Forstner, C.; Raghavendra, A.K.; Mathesius, U.; Richardson, A.E.; Delhaize, E.; Gilliham, M.; Watt, M.; Ryan, P.R. Manipulating exudate composition from root apices shapes the microbiome throughout the root system. Plant Physiol. 2021, 187, 2279–2295. [Google Scholar] [CrossRef]
- Shelp, B.J.; Allan, W.L.; Faure, D. Role of g-aminobutyrate and g-hydroxybutyrate in plant communication. In Plant-Environment Interactions; Springer: Berlin/Heidelberg, Germany, 2009; pp. 73–84. [Google Scholar]
- Shelp, B.J.; Bown, A.W.; Zarei, A. 4-Aminobutyrate (GABA): A metabolite and signal with practical significance. Botany 2017, 95, 1015–1032. [Google Scholar] [CrossRef] [Green Version]
- Bouche, N.; Lacombe, B.; Fromm, H. GABA signaling: A conserved and ubiquitous mechanism. Trends Cell Biol. 2003, 13, 607–610. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bown, A.W.; Faure, D. Extracellular γ-aminobutyrate mediates communication between plants and other organisms. Plant Physiol. 2006, 142, 1350–1352. [Google Scholar] [CrossRef] [Green Version]
- Shelp, B.J.; Bown, A.W.; McLean, M.D. Metabolism and functions of gamma-aminobutyric acid. Trends Plant Sci. 1999, 4, 446–452. [Google Scholar] [CrossRef]
- Serraj, R.; Shelp, B.J.; Sinclair, T.R. Accumulation of γ-aminobutyric acid in nodulated soybean in response to drought stress. Physiol. Plant. 1998, 102, 79–86. [Google Scholar] [CrossRef]
- Bown, A.W.; Shelp, B.J. The metabolism and functions of -aminobutyric acid. Plant Physiol. Biochem. 1997, 115, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bown, A.; Shelp, B. The metabolism and physiological roles of 4-aminobutyric acid. Biochem. Life Sci. Adv. 1989, 8, 21–25. [Google Scholar]
- Bown, A.W.; Hall, D.E.; MacGregor, K.B. Insect footsteps on leaves stimulate the accumulation of 4-aminobutyrate and can be visualized through increased chlorophyll fluorescence and superoxide production. Plant Physiol. 2002, 129, 1430–1434. [Google Scholar] [CrossRef] [Green Version]
- Bown, A.W.; MacGregor, K.B.; Shelp, B.J. Gamma-aminobutyrate: Defense against invertebrate pests? Trends Plant Sci. 2006, 11, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Mekonnen, D.W.; Flügge, U.-I.; Ludewig, F. Gamma-aminobutyric acid depletion affects stomata closure and drought tolerance of Arabidopsis thaliana. Plant Sci. 2016, 245, 25–34. [Google Scholar] [CrossRef]
- Crawford, L.A.; Bown, A.W.; Breitkreuz, K.E.; Guinel, F.C. The synthesis of [gamma]-aminobutyric acid in response to treatments reducing cytosolic pH. Plant Physiol. 1994, 104, 865–871. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Long, Y.; Feng, X.; Zhu, X.; Sai, N.; Chirkova, L.; Betts, A.; Herrmann, J.; Edwards, E.J.; Okamoto, M. GABA signalling modulates stomatal opening to enhance plant water use efficiency and drought resilience. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Tyerman, S.D.; Gilliham, M. Cytosolic GABA inhibits anion transport by wheat ALMT1. New Phytol. 2020, 225, 671–678. [Google Scholar] [CrossRef]
- John, R.A.; Charteris, A. The reaction of amino-oxyacetate with pyridoxal phosphate-dependent enzymes. Biochem. J. 1978, 171, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.; McRae, D.; Joy, K. Glutamate and y-Aminobutyrate Metabolism. Mol. Plant-Microbe Interact. 1991, 4, 37–45. [Google Scholar] [CrossRef]
- Snedden, W.A.; Chung, I.; Pauls, R.H.; Bown, A.W. Proton/L-glutamate symport and the regulation of intracellular pH in isolated mesophyll cells. Plant Physiol. 1992, 99, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Livingston, J.; Beaumont, D.; Arzimanoglou, A.; Aicardi, J. Vigabatrin in the treatment of epilepsy in children. Br. J. Clin. Pharmacol. 1989, 27, 109S–112S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanavati, S.M.; Silverman, R.B. Mechanisms of inactivation of. gamma-aminobutyric acid aminotransferase by the antiepilepsy drug. gamma.-vinyl GABA (vigabatrin). J. Am. Chem. Soc. 1991, 113, 9341–9349. [Google Scholar] [CrossRef]
- Condie, B.G.; Bain, G.; Gottlieb, D.I.; Capecchi, M.R. Cleft palate in mice with a targeted mutation in the γ-aminobutyric acid-producing enzyme glutamic acid decarboxylase 67. Proc. Natl. Acad. Sci. USA 1997, 94, 11451–11455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magri, C.; Giacopuzzi, E.; La Via, L.; Bonini, D.; Ravasio, V.; Elhussiny, M.E.; Orizio, F.; Gangemi, F.; Valsecchi, P.; Bresciani, R. A novel homozygous mutation in GAD1 gene described in a schizophrenic patient impairs activity and dimerization of GAD67 enzyme. Sci. Rep. 2018, 8, 15470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, S.A.; Kamran, M.; Sullivan, W.; Chirkova, L.; Okamoto, M.; Degryse, F.; McLaughlin, M.; Gilliham, M.; Tyerman, S.D. Aluminum-Activated Malate Transporters Can Facilitate GABA Transport. Plant Cell 2018, 30, 1147–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delhaize, E.; Ryan, P.R. Aluminum toxicity and tolerance in plants. Plant Physiol. 1995, 107, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineros, M.A.; Cançado, G.M.; Kochian, L.V. Novel properties of the wheat aluminum tolerance organic acid transporter (TaALMT1) revealed by electrophysiological characterization in Xenopus oocytes: Functional and structural implications. Plant Physiol. 2008, 147, 2131–2146. [Google Scholar] [CrossRef] [Green Version]
- Kamran, M.; Ramesh, S.A.; Gilliham, M.; Tyerman, S.D.; Bose, J. Role of TaALMT1 malate-GABA transporter in alkaline pH tolerance of wheat. Plant Cell Environ. 2020, 43, 2443–2459. [Google Scholar] [CrossRef]
- Wang, J.; Yu, X.; Ding, Z.J.; Zhang, X.; Luo, Y.; Xu, X.; Xie, Y.; Li, X.; Yuan, T.; Zheng, S.J. Structural basis of ALMT1-mediated aluminum resistance in Arabidopsis. Cell Res. 2022, 32, 89–98. [Google Scholar] [CrossRef]
- Rho, J.M.; Donevan, S.D.; Rogawski, M.A. Direct activation of GABAA receptors by barbiturates in cultured rat hippocampal neurons. J. Physiol. 1996, 497 Pt 2, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Newland, C.F.; Cull-Candy, S.G. On the mechanism of action of picrotoxin on GABA receptor channels in dissociated sympathetic neurones of the rat. J. Physiol. 1992, 447, 191. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Covey, D.F.; Akabas, M.H. Interaction of picrotoxin with GABAA receptor channel-lining residues probed in cysteine mutants. Biophys. J. 1995, 69, 1858–1867. [Google Scholar] [CrossRef] [Green Version]
- Sedelnikova, A.; Erkkila, B.E.; Harris, H.; Zakharkin, S.O.; Weiss, D.S. Stoichiometry of a pore mutation that abolishes picrotoxin-mediated antagonism of the GABAA receptor. J. Physiol. 2006, 577, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Erkkila, B.E.; Sedelnikova, A.V.; Weiss, D.S. Stoichiometric pore mutations of the GABA(A)R reveal a pattern of hydrogen bonding with picrotoxin. Biophys. J. 2008, 94, 4299–4306. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.W.; Rajendra, S.; Barry, P.H.; Schofield, P.R. Mutations affecting the glycine receptor agonist transduction mechanism convert the competitive antagonist, picrotoxin, into an allosteric potentiator. J. Biol. Chem. 1995, 270, 13799–13806. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.W. Picrotoxin-like channel blockers of GABAA receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 6081–6082. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Yamamoto, Y.; Ezaki, B.; Katsuhara, M.; Ahn, S.J.; Ryan, P.R.; Delhaize, E.; Matsumoto, H. A wheat gene encoding an aluminium-activated malate transporter. Plant J. 2004, 37, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Preuss, C.P.; Huang, C.Y.; Gilliham, M.; Tyerman, S.D. Channel-Like Characteristics of the Low-Affinity Barley Phosphate Transporter PHT1;6 When Expressed in Xenopus Oocytes. Plant Physiol. 2010, 152, 1431–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoekenga, O.A.; Maron, L.G.; Piñeros, M.A.; Cançado, G.M.; Shaff, J.; Kobayashi, Y.; Ryan, P.R.; Dong, B.; Delhaize, E.; Sasaki, T. AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 9738–9743. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Chung, Y.-W.; Wong, P. Potassium channel activity recorded from the apical membrane of freshly isolated epithelial cells in rat caudal epididymis. Biol. Reprod. 1999, 60, 1509–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maksaev, G.; Haswell, E.S. Expressing and characterizing mechanosensitive channels in Xenopus oocytes. In Plant Gravitropism; Springer: Berlin/Heidelberg, Germany, 2015; pp. 151–169. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, W.; Gilbert, D.; Hawthorne, R.; Hryciw, D.H.; Frings, S.; Poronnik, P.; Lynch, J.W. A yellow fluorescent protein-based assay for high-throughput screening of glycine and GABAA receptor chloride channels. Neurosci. Lett. 2005, 380, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Etter, A.; Cully, D.; Liu, K.; Reiss, B.; Vassilatis, D.; Schaeffer, J.; Arena, J. Picrotoxin blockade of invertebrate glutamate-gated chloride channels: Subunit dependence and evidence for binding within the pore. J. Neurochem. 1999, 72, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Hawthorne, R.; Lynch, J.W. A picrotoxin-specific conformational change in the glycine receptor M2–M3 loop. J. Biol. Chem. 2005, 280, 35836–35843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kash, T.L.; Jenkins, A.; Kelley, J.C.; Trudell, J.R.; Harrison, N.L. Coupling of agonist binding to channel gating in the GABAA receptor. Nature 2003, 421, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P. Pharmacological evidence for two types of postsynaptic glycinergic receptors on the Mauthner cell of 52-h-old zebrafish larvae. J. Neurophysiol. 1997, 77, 2400–2415. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.; Covey, D.; Rothman, S. Multiple mechanisms of picrotoxin block of GABA-induced currents in rat hippocampal neurons. J. Physiol. 1993, 464, 423–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dibas, M.I.; Gonzales, E.B.; Das, P.; Bell-Horner, C.L.; Dillon, G.H. Identification of a novel residue within the second transmembrane domain that confers use-facilitated block by picrotoxin in glycine α1 receptors. J. Biol. Chem. 2002, 277, 9112–9117. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Weiss, D.S. Site-specific fluorescence reveals distinct structural changes with GABA receptor activation and antagonism. Nat. Neurosci. 2002, 5, 1163. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Eskandari, S.; Grallath, S.; Rentsch, D. AtGAT1, a high affinity transporter for gamma-aminobutyric acid in Arabidopsis thaliana. J. Biol. Chem. 2006, 281, 7197–7204. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramesh, S.A.; Long, Y.; Dashtbani-Roozbehani, A.; Gilliham, M.; Brown, M.H.; Tyerman, S.D. Picrotoxin Delineates Different Transport Configurations for Malate and γ Aminobutyric Acid through TaALMT1. Biology 2022, 11, 1162. https://doi.org/10.3390/biology11081162
Ramesh SA, Long Y, Dashtbani-Roozbehani A, Gilliham M, Brown MH, Tyerman SD. Picrotoxin Delineates Different Transport Configurations for Malate and γ Aminobutyric Acid through TaALMT1. Biology. 2022; 11(8):1162. https://doi.org/10.3390/biology11081162
Chicago/Turabian StyleRamesh, Sunita A., Yu Long, Abolfazl Dashtbani-Roozbehani, Matthew Gilliham, Melissa H. Brown, and Stephen D. Tyerman. 2022. "Picrotoxin Delineates Different Transport Configurations for Malate and γ Aminobutyric Acid through TaALMT1" Biology 11, no. 8: 1162. https://doi.org/10.3390/biology11081162
APA StyleRamesh, S. A., Long, Y., Dashtbani-Roozbehani, A., Gilliham, M., Brown, M. H., & Tyerman, S. D. (2022). Picrotoxin Delineates Different Transport Configurations for Malate and γ Aminobutyric Acid through TaALMT1. Biology, 11(8), 1162. https://doi.org/10.3390/biology11081162