
Effect of Ethanol Consumption on the Placenta and Liver of Partially IGF-1-Deficient Mice: The Role of Metabolism via CYP2E1 and the Antioxidant Enzyme System

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
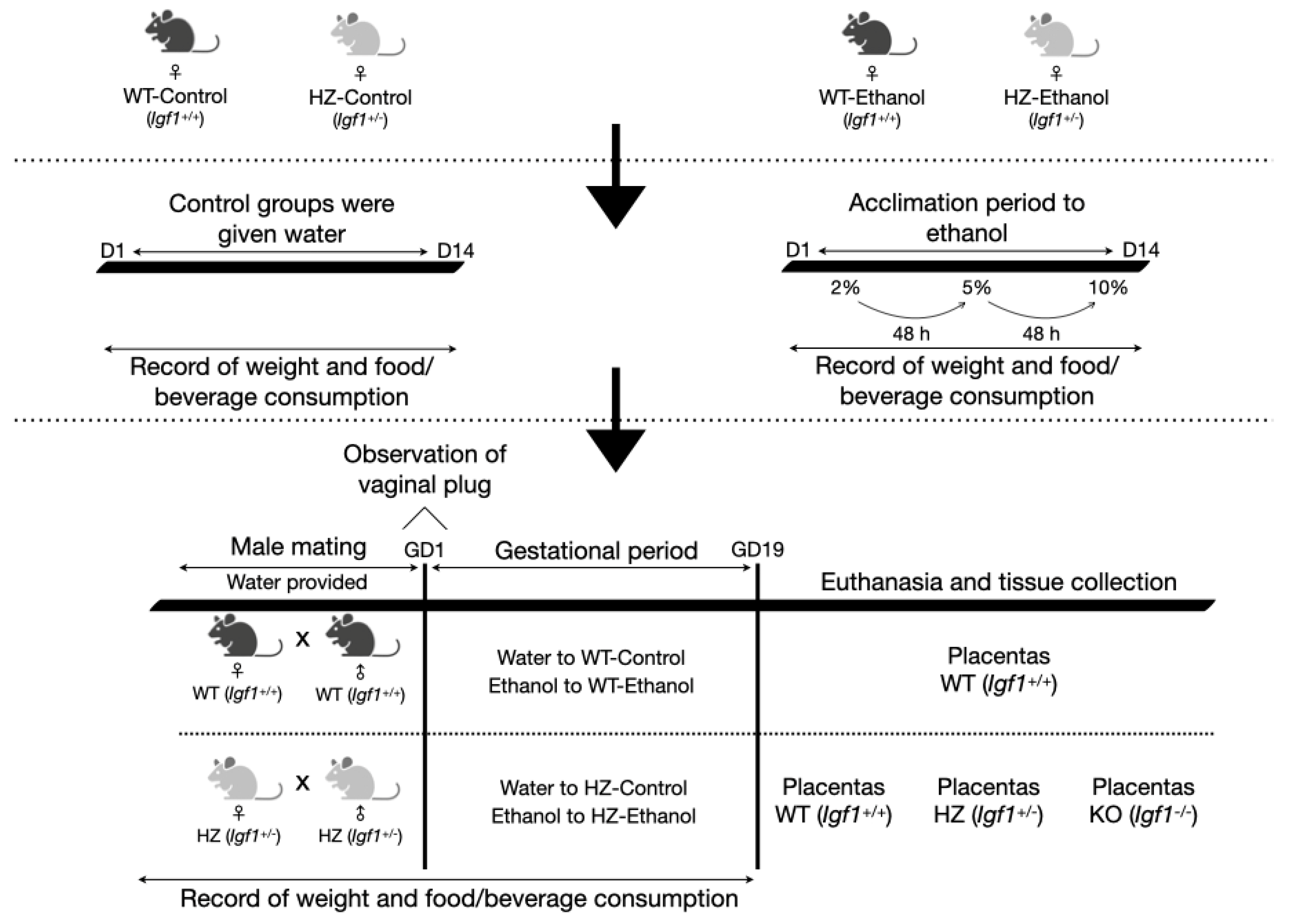
2.1. Animals and Experimental Design
2.2. Sexing and Genotyping of Animals
2.3. Gene Expression Studies via RT-qPCR
2.4. Immunoblotting Analysis
2.5. Immunohistochemical Analysis
2.6. Antioxidant Enzymatic Status
2.7. Statistical Analysis
3. Results
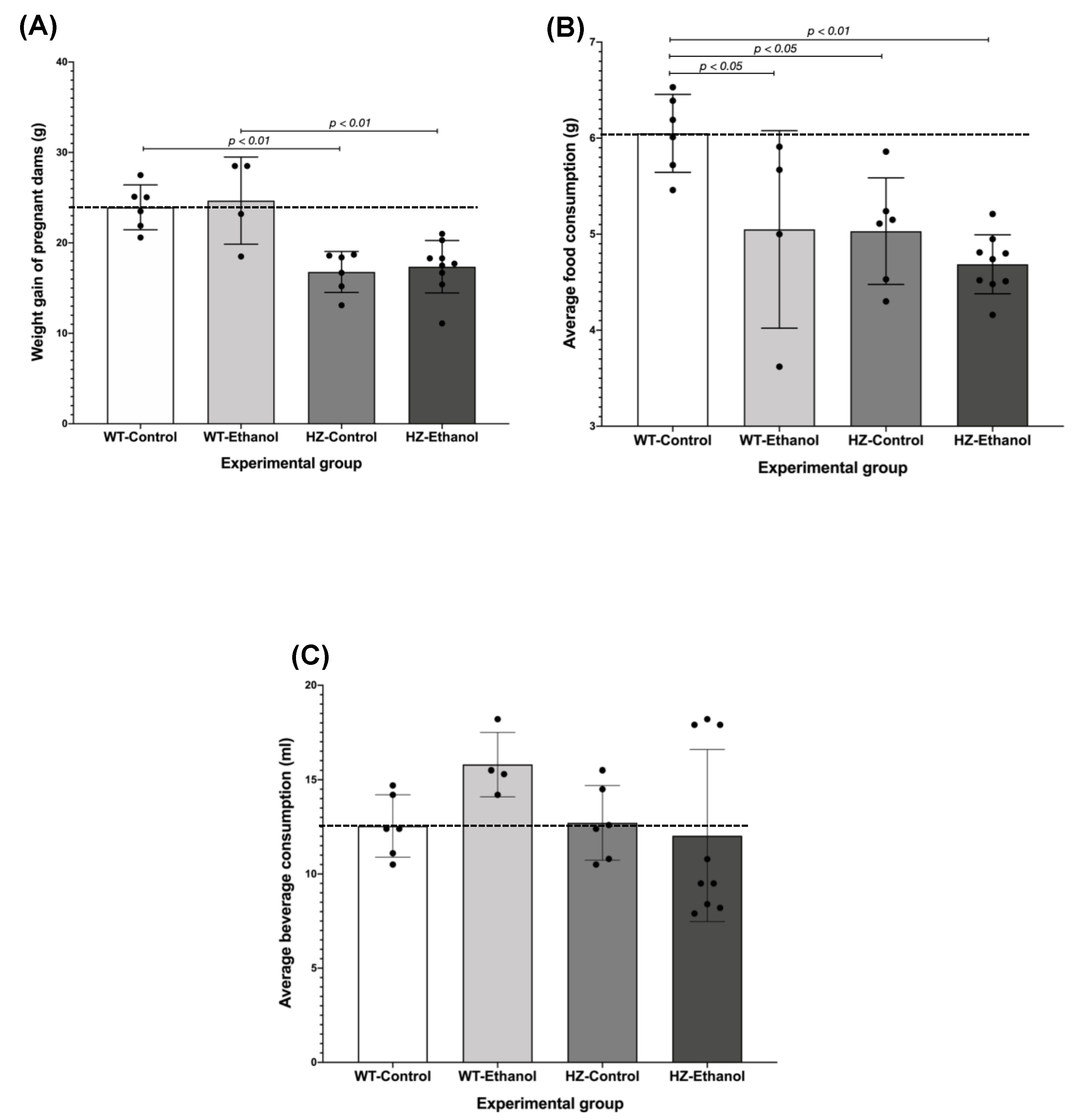
3.1. The Effect of Ethanol Consumption in Maternal Parameters before and after Gestation
3.2. The Effect of Ethanol Consumption in Fetal and Placental Parameters
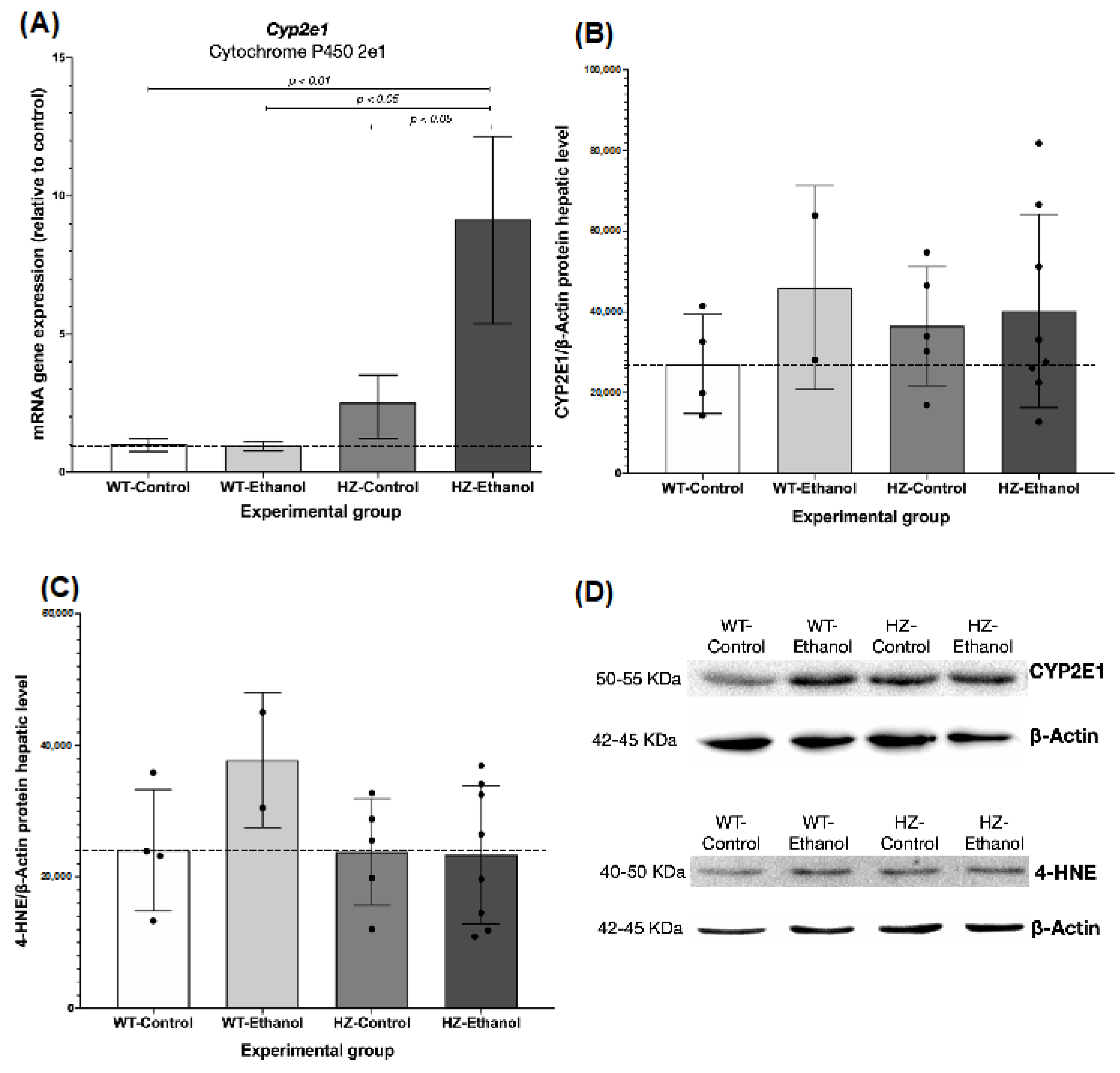
3.3. The Effect of Ethanol Consumption in Placental and Hepatic Expression of CYP2E1
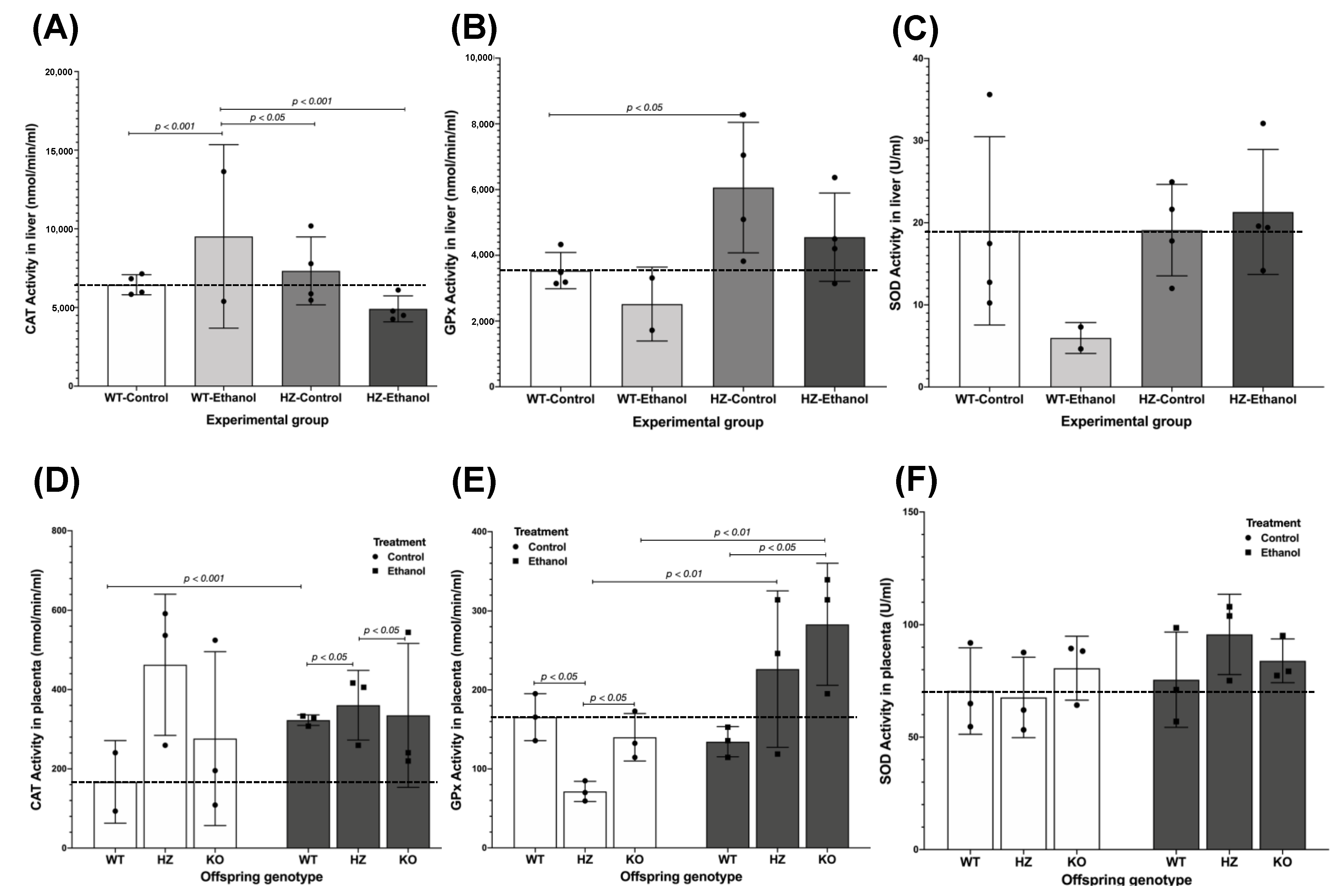
3.4. The Effect of Ethanol Consumption on the Oxidative Environment in Liver from Pregnant Dams
3.5. Antioxidant Enzyme Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martín-Estal, I.; Rodriguez-Zambrano, M.A.; Castilla-Cortázar, I. Biochemical Assessment of Placental Function. In Fetal Growth Restriction; Springer International Publishing: Cham, Switzerland, 2019; pp. 83–116. [Google Scholar]
- Creeth, H.D.J.; John, R.M. The Placental Programming Hypothesis: Placental Endocrine Insufficiency and the Co-Occurrence of Low Birth Weight and Maternal Mood Disorders. Placenta 2020, 98, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Netchine, I.; Azzi, S.; Le Bouc, Y.; Savage, M.O. IGF1 Molecular Anomalies Demonstrate Its Critical Role in Fetal, Postnatal Growth and Brain Development. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Guerri, C.; Bazinet, A.; Riley, E.P. Foetal Alcohol Spectrum Disorders and Alterations in Brain and Behaviour. Alcohol. Alcohol. 2009, 44, 108–114. [Google Scholar] [CrossRef]
- Wilson, D.F.; Matschinsky, F.M. Ethanol Metabolism: The Good, the Bad, and the Ugly. Med. Hypotheses 2020, 140, 109638. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.M.; Huuskonen, P.; Juvonen, R.; Sahlman, H.; Repo, J.; Myöhänen, K.; Myllynen, P.; Woo, C.-S.J.; Karttunen, V.; Vähäkangas, K. Activities of Metabolizing Enzymes in Human Placenta. Toxicol. Lett. 2020, 326, 70–77. [Google Scholar] [CrossRef]
- Nguyen, T.M.T.; Steane, S.E.; Moritz, K.M.; Akison, L.K. Prenatal Alcohol Exposure Programmes Offspring Disease: Insulin Resistance in Adult Males in a Rat Model of Acute Exposure. J. Physiol. 2019, 597, 5619–5637. [Google Scholar] [CrossRef]
- Koop, D.R. Alcohol Metabolism’s Damaging Effects on the Cell: A Focus on Reactive Oxygen Generation by the Enzyme Cytochrome P450 2E1. Alcohol Res. Health 2006, 29, 274–280. [Google Scholar]
- Ramachandran, V.; Perez, A.; Chen, J.; Senthil, D.; Schenker, S.; Henderson, G.I. In Utero Ethanol Exposure Causes Mitochondrial Dysfunction, Which Can Result in Apoptotic Cell Death in Fetal Brain: A Potential Role for 4-Hydroxynonenal. Alcohol. Clin. Exp. Res. 2001, 25, 862–871. [Google Scholar] [CrossRef]
- Gårdebjer, E.M.; Cuffe, J.S.M.; Pantaleon, M.; Wlodek, M.E.; Moritz, K.M. Periconceptional Alcohol Consumption Causes Fetal Growth Restriction and Increases Glycogen Accumulation in the Late Gestation Rat Placenta. Placenta 2014, 35, 50–57. [Google Scholar] [CrossRef]
- Gatford, K.L.; Dalitz, P.A.; Cock, M.L.; Harding, R.; Owens, J.A. Acute Ethanol Exposure in Pregnancy Alters the Insulin-like Growth Factor Axis of Fetal and Maternal Sheep. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E494-500. [Google Scholar] [CrossRef]
- Seiler, A.E.; Ross, B.N.; Green, J.S.; Rubin, R. Differential Effects of Ethanol on Insulin-like Growth Factor-I Receptor Signaling. Alcohol. Clin. Exp. Res. 2000, 24, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Castilla-Cortazar, I.; Guerra, L.; Puche, J.E.; Munoz, U.; Barhoum, R.; Escudero, E.; Lavandera, J.L. An Experimental Model of Partial Insulin-like Growth Factor-1 Deficiency in Mice. J. Physiol. Biochem. 2014, 71, 129–139. [Google Scholar] [CrossRef]
- Kleiber, M.L.; Wright, E.; Singh, S.M. Maternal Voluntary Drinking in C57BL/6J Mice: Advancing a Model for Fetal Alcohol Spectrum Disorders. Behav. Brain Res. 2011, 223, 376–387. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.C.; Senekal, M.; Dodge, N.C.; Bechard, L.J.; Meintjes, E.M.; Molteno, C.D.; Duggan, C.P.; Jacobson, J.L.; Jacobson, S.W. Maternal Alcohol Use and Nutrition During Pregnancy: Diet and Anthropometry. Alcohol. Clin. Exp. Res. 2017, 41, 2114–2127. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.C.; Jacobson, J.L.; Sokol, R.J.; Avison, M.J.; Jacobson, S.W. Fetal Alcohol-Related Growth Restriction from Birth through Young Adulthood and Moderating Effects of Maternal Prepregnancy Weight. Alcohol. Clin. Exp. Res. 2013, 37, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.P.; Grimberg, A. Insulin-Like Growth Factor-I Is a Marker for the Nutritional State. Pediatr. Endocrinol. Rev. 2015, 13, 499–511. [Google Scholar]
- Koutsaki, M.; Sifakis, S.; Zaravinos, A.; Koutroulakis, D.; Koukoura, O.; Spandidos, D.A. Decreased Placental Expression of HPGH, IGF-I and IGFBP-1 in Pregnancies Complicated by Fetal Growth Restriction. Growth Horm. IGF Res. 2011, 21, 31–36. [Google Scholar] [CrossRef]
- Gundogan, F.; Elwood, G.; Mark, P.; Feijoo, A.; Longato, L.; Tong, M.; de la Monte, S.M. Ethanol-Induced Oxidative Stress and Mitochondrial Dysfunction in Rat Placenta: Relevance to Pregnancy Loss. Alcohol. Clin. Exp. Res. 2010, 34, 415–423. [Google Scholar] [CrossRef]
- Martín-Estal, I.; Fajardo-Ramírez, O.R.; Bermúdez De León, M.; Zertuche-Mery, C.; Rodríguez-Mendoza, D.; Gómez-Álvarez, P.; Galindo-Rangel, M.; Leal López, A.; Castilla-Cortázar, I.; Castorena Torres, F. Ethanol Consumption during Gestation Promotes Placental Alterations in IGF-1 Deficient Mice. F1000Research 2021, 10, 1284. [Google Scholar] [CrossRef]
- Dey, A.; Cederbaum, A.I. Alcohol and Oxidative Liver Injury. Hepatology 2006, 43, S63–S74. [Google Scholar] [CrossRef] [PubMed]
- Probyn, M.E.; Zanini, S.; Ward, L.C.; Bertram, J.F.; Moritz, K.M. A Rodent Model of Low- to Moderate-Dose Ethanol Consumption during Pregnancy: Patterns of Ethanol Consumption and Effects on Fetal and Offspring Growth. Reprod. Fertil. Dev. 2012, 24, 859. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, A.J.; Walker, K.R.; Kobus, S.M.; Poklewska-Koziell, M.; Reynolds, J.N.; Brien, J.F. Differential Effects of Chronic Ethanol Exposure on Cytochrome P450 2E1 and the Hypothalamic–Pituitary–Adrenal Axis in the Maternal–Fetal Unit of the Guinea Pig. Neurotoxicol. Teratol. 2010, 32, 164–170. [Google Scholar] [CrossRef]
- Jin, M.; Ande, A.; Kumar, A.; Kumar, S. Regulation of Cytochrome P450 2e1 Expression by Ethanol: Role of Oxidative Stress-Mediated Pkc/Jnk/Sp1 Pathway. Cell Death Dis. 2013, 4, e554. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, J.B. Transcriptional Regulation Is Only Half the Story. Mol. Syst. Biol. 2010, 6, 406. [Google Scholar] [CrossRef]
- Li, Z.; Chen, X.; Liu, Z.; Ye, W.; Li, L.; Qian, L.; Ding, H.; Li, P.; Aung, L.H.H. Recent Advances: Molecular Mechanism of RNA Oxidation and Its Role in Various Diseases. Front. Mol. Biosci. 2020, 7, 184. [Google Scholar] [CrossRef]
- Myllynen, P.; Immonen, E.; Kummu, M.; Vähäkangas, K. Developmental Expression of Drug Metabolizing Enzymes and Transporter Proteins in Human Placenta and Fetal Tissues. Expert Opin. Drug Metab. Toxicol. 2009, 5, 1483–1499. [Google Scholar] [CrossRef] [PubMed]
- Rojanathammanee, L.; Rakoczy, S.; Kopchick, J.; Brown-Borg, H.M. Effects of Insulin-like Growth Factor 1 on Glutathione S-Transferases and Thioredoxin in Growth Hormone Receptor Knockout Mice. Age 2014, 36, 9687. [Google Scholar] [CrossRef]
- Marianian, A.; Atalyan, A.; Bohora, S.; Darenskaya, M.; Grebenkina, L.; Kolesnikova, L.; Kolesnikov, S.; Mikhaylevich, I.; Protopopova, N.; Stockett, M.; et al. The Effect of Low Alcohol Consumption during Pregnancy on the Lipid Peroxidation-antioxidant Defense System of Women, Their Alcohol-exposed Infants, and Growth, Health, and Developmental Outcomes. Birth Defects Res. 2020, 112, 40–53. [Google Scholar] [CrossRef]
- Bailey, S.M.; Patel, V.B.; Young, T.A.; Asayama, K.; Cunningham, C.C. Chronic Ethanol Consumption Alters the Glutathione/Glutathione Peroxidase-1 System and Protein Oxidation Status in Rat Liver. Alcohol. Clin. Exp. Res. 2001, 25, 726–733. [Google Scholar] [CrossRef]
- Das, S.K.; Vasudevan, D.M. Effect of Ethanol on Liver Antioxidant Defense Systems: Adose Dependent Study. Indian J. Clin. Biochem. 2005, 20, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Aksu, I.; Baykara, B.; Kiray, M.; Gurpinar, T.; Sisman, A.R.; Ekerbicer, N.; Tas, A.; Gokdemir-Yazar, O.; Uysal, N. Serum IGF-1 Levels Correlate Negatively to Liver Damage in Diabetic Rats. Biotech. Histochem. 2013, 88, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Brown-Borg, H.M.; Rakoczy, S.G.; Romanick, M.A.; Kennedy, M.A. Effects of Growth Hormone and Insulin-Like Growth Factor-1 on Hepatocyte Antioxidative Enzymes. Exp. Biol. Med. 2002, 227, 94–104. [Google Scholar] [CrossRef]
- Nogales, F.; Ojeda, M.L.; Jotty, K.; Murillo, M.L.; Carreras, O. Maternal Ethanol Consumption Reduces Se Antioxidant Function in Placenta and Liver of Embryos and Breastfeeding Pups. Life Sci. 2017, 190, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Roland, L.; Beauchemin, D.; Acteau, G.; Fradette, C.; St-Pierre, I.; Bilodeau, J.-F. Effects of Labor on Placental Expression of Superoxide Dismutases in Preeclampsia. Placenta 2010, 31, 392–400. [Google Scholar] [CrossRef]
- Puche, J.E.; Castilla-Cortázar, I. Human Conditions of Insulin-like Growth Factor-I (IGF-I) Deficiency. J. Transl. Med. 2012, 10, 224. [Google Scholar] [CrossRef]
- Castilla-Cortazar, I.; Femat-Roldán, G.; Rodríguez-Rivera, J.; Aguirre, G.A.; García-Magariño, M.; Martín-Estal, I.; Espinosa, L.; Díaz-Olachea, C. Mexican Case Report of a Never-Treated Laron Syndrome Patient Evolving to Metabolic Syndrome, Type 2 Diabetes, and Stroke. Clin. Case Rep. 2017, 5, 1852–1855. [Google Scholar] [CrossRef]
- Castilla-Cortazar, I.; De Ita, J.R.; García-Magariño, M.; Aguirre, G.A.; Castorena-Torres, F.; Valdez-Garcia, J.E.; Ortiz-Urbina, J.; de la Garza, R.G.; Fraustro-Avila, E.; Rodríguez-Zambrano, M.A.; et al. A 42-Year-Old Woman with Untreated Growth Hormone Insensitivity, Diabetic Retinopathy, and Gene Sequencing Identifies a Variant of Laron Syndrome. Am. J. Case Rep. 2019, 20, 689–696. [Google Scholar] [CrossRef]
- Castilla-Cortazar, I.; de Ita, J.R.; Aguirre, G.A.; Castorena-Torres, F.; Ortiz-Urbina, J.; García-Magariño, M.; de la Garza, R.G.; Diaz Olachea, C.; Elizondo Leal, M.I. Fanconi Anemia and Laron Syndrome. Am. J. Med. Sci. 2017, 353, 425–432. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Maternal Body Weight (G) | Maternal Liver Weight (G) | Ratio Maternal Liver/Body Weight | Food Consumption (G)/BODY Weight (G) |
|---|---|---|---|---|
| WT-Control (n = 6) | 52.05 ± 1.21 | 2.73 ± 0.08 | 5.24% | 0.175 ± 0.005 |
| WT-Ethanol (n = 4) | 51.73 ± 5.82 | 2.88 ± 0.10 | 5.57% * | 0.140 ± 0.011 a,* |
| HZ-Control (n = 6) | 40.73 ± 1.28 b,c | 2.24 ± 0.07 c | 5.50% | 0.188 ± 0.021 c |
| HZ-Ethanol (n = 9) | 42.76 ± 1.32 d | 2.49 ± 0.16 | 5.82% * | 0.157 ± 0.004 d,* |
| Group | Offspring Genotype | Fetal Height (CM) | Fetal Weight (G) | Placental Weight (G) |
|---|---|---|---|---|
| Controls | WT (n = 9) | 3.33 ± 0.066 | 1.08 ± 0.039 | 0.10 ± 0.007 |
| HZ (n = 32) | 3.22 ± 0.035 | 1.03 ± 0.021 | 0.09 ± 0.004 | |
| KO (n = 19) | 2.75 ± 0.045 b,c | 0.72 ± 0.028 b,c | 0.08 ± 0.005 | |
| Ethanol exposure | WT (n = 23) | 3.17 ± 0.041 a | 1.03 ± 0.024 | 0.10 ± 0.004 |
| HZ (n = 44) | 3.18 ± 0.030 | 0.98 ± 0.018 a | 0.09 ± 0.003 | |
| KO (n = 16) | 2.74 ± 0.050 b,c | 0.66 ± 0.030 b,c | 0.10 ± 0.005 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Estal, I.; Fajardo-Ramírez, Ó.R.; Bermúdez de León, M.; Zertuche-Mery, C.; Benavides-Guajardo, R.; García-Cruz, M.I.; Rodríguez De Ita, J.; Castilla-Cortázar, I.; Castorena-Torres, F. Effect of Ethanol Consumption on the Placenta and Liver of Partially IGF-1-Deficient Mice: The Role of Metabolism via CYP2E1 and the Antioxidant Enzyme System. Biology 2022, 11, 1264. https://doi.org/10.3390/biology11091264
Martín-Estal I, Fajardo-Ramírez ÓR, Bermúdez de León M, Zertuche-Mery C, Benavides-Guajardo R, García-Cruz MI, Rodríguez De Ita J, Castilla-Cortázar I, Castorena-Torres F. Effect of Ethanol Consumption on the Placenta and Liver of Partially IGF-1-Deficient Mice: The Role of Metabolism via CYP2E1 and the Antioxidant Enzyme System. Biology. 2022; 11(9):1264. https://doi.org/10.3390/biology11091264
Chicago/Turabian StyleMartín-Estal, Irene, Óscar R. Fajardo-Ramírez, Mario Bermúdez de León, Carolina Zertuche-Mery, Rodolfo Benavides-Guajardo, María Isabel García-Cruz, Julieta Rodríguez De Ita, Inma Castilla-Cortázar, and Fabiola Castorena-Torres. 2022. "Effect of Ethanol Consumption on the Placenta and Liver of Partially IGF-1-Deficient Mice: The Role of Metabolism via CYP2E1 and the Antioxidant Enzyme System" Biology 11, no. 9: 1264. https://doi.org/10.3390/biology11091264
APA StyleMartín-Estal, I., Fajardo-Ramírez, Ó. R., Bermúdez de León, M., Zertuche-Mery, C., Benavides-Guajardo, R., García-Cruz, M. I., Rodríguez De Ita, J., Castilla-Cortázar, I., & Castorena-Torres, F. (2022). Effect of Ethanol Consumption on the Placenta and Liver of Partially IGF-1-Deficient Mice: The Role of Metabolism via CYP2E1 and the Antioxidant Enzyme System. Biology, 11(9), 1264. https://doi.org/10.3390/biology11091264