

A Fuzzy-Based Model to Predict the Spatio-Temporal Performance of the Dolichogenidea gelechiidivoris Natural Enemy against Tuta absoluta under Climate Change

 ,
,  ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Assumption and Modelling Procedure
2.3. Data
2.4. Land Use/Land Cover Characterization
2.5. Presence of the Parasitoid Host, T. absoluta
2.6. Predicting the Spatio-Temporal Performance of D. gelechiidivoris
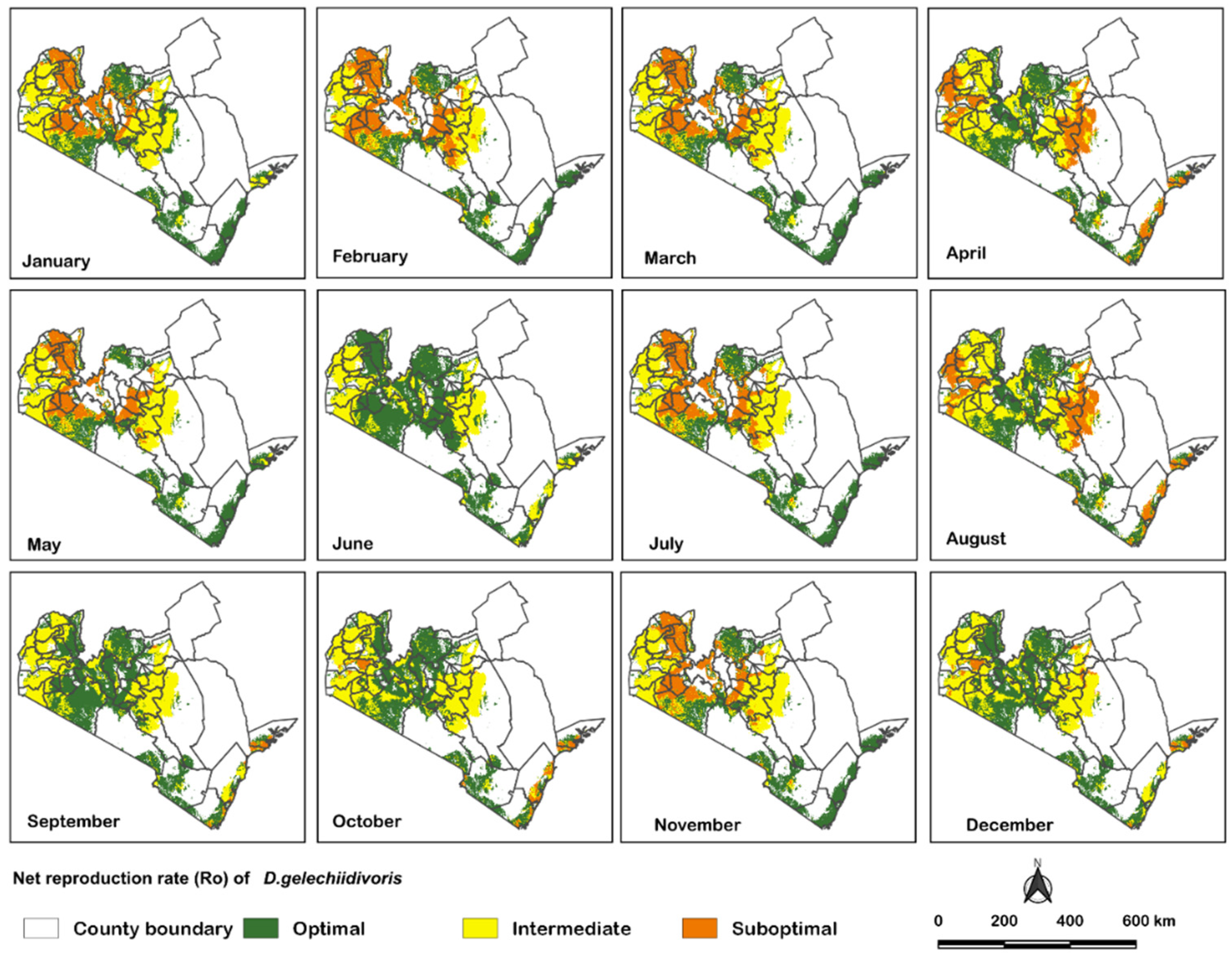
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lux, S.A.; Copeland, R.S.; White, I.M.; Manrakhan, A.; Billah, M.K. A new invasive fruit fly species from the Bactrocera dorsalis (Hendel) group detected in East Africa. Int. J. Trop. Insect Sci. 2003, 23, 355–361. [Google Scholar] [CrossRef]
- Mohamed, E.S.I.; Mohamed, M.E.; Gamiel, S.A. First record of the tomato leafminer, Tuta absoluta (Meyrick)(L epidoptera: Gelechiidae) in Sudan. EPPO Bull. 2012, 42, 325–327. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall Armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed]
- Kwadha, C.A.; Okwaro, L.A.; Kleman, I.; Rehermann, G.; Revadi, S.; Ndela, S.; Khamis, F.M.; Nderitu, P.W.; Kasina, M.; George, M.K.; et al. Detection of the spotted wing drosophila, Drosophila suzukii, in continental sub-Saharan Africa. J. Pest Sci. 2021, 94, 251–259. [Google Scholar] [CrossRef]
- Giorgini, M.; Guerrieri, E.; Cascone, P.; Gontijo, L. Current strategies and future outlook for managing the neotropical tomato pest Tuta absoluta (Meyrick) in the Mediterranean Basin. Neotrop. Entomol. 2019, 48, 1–17. [Google Scholar] [CrossRef]
- Illakwahhi, D.T.; Srivastava, P.B.B.L. Control and management of tomato leafminer -Tuta Absoluta (Meyrick) (Lepidotera, Gelechiidae).A Review. IOSR J. Appl. Chem. 2017, 10, 14–22. [Google Scholar] [CrossRef]
- Machekano, H.; Mutamiswa, R.; Nyamukondiwa, C. Evidence of rapid spread and establishment of Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae) in semi-arid Botswana. Agric. Food Secur. 2018, 7, 1–12. [Google Scholar] [CrossRef]
- Mohamed, S.A.; Wamalwa, M.; Obala, F.; Tonnang, H.E.Z.; Tefera, T.; Calatayud, P.-A.; Subramanian, S.; Ekesi, S. A deadly encounter: Alien invasive Spodoptera frugiperda in Africa and indigenous natural enemy, Cotesia icipe (Hymenoptera, Braconidae). PLoS ONE 2021, 16, e0253122. [Google Scholar] [CrossRef]
- Aigbedion-Atalor, P.O.; Mohamed, S.A.; Hill, M.P.; Zalucki, M.P.; Azrag, A.G.A.; Srinivasan, R.; Ekesi, S. Host stage preference and performance of Dolichogenidea gelechiidivoris (Hymenoptera: Braconidae), a candidate for classical biological control of Tuta absoluta in Africa. Biol. Control 2020, 144, 104215. [Google Scholar] [CrossRef]
- Guimapi, R.Y.A.; Mohamed, S.A.; Okeyo, G.O.; Ndjomatchoua, F.T.; Ekesi, S.; Tonnang, H.E.Z. Modeling the risk of invasion and spread of Tuta absoluta in Africa. Ecol. Complex. 2016, 28, 77–93. [Google Scholar] [CrossRef]
- Fiaboe, K.R.; Agboka, K.; Agboyi, L.K.; Koffi, D.; Ofoe, R.; Kpadonou, G.E.; Agnamba, A.O.; Assogba, K.; Adjevi, M.K.A.; Zanou, K.T.; et al. First report and distribution of the South American tomato pinworm, Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae) in Togo. Phytoparasitica 2021, 49, 167–177. [Google Scholar] [CrossRef]
- Mansour, R.; Brévault, T.; Chailleux, A.; Cherif, A.; Grissa-Lebdi, K.; Haddi, K.; Mohamed, S.A.; Nofemela, R.S.; Oke, A.; Sylla, S.; et al. Occurrence, biology, natural enemies and management of Tuta absoluta in Africa. Entomol. Gen. 2018, 38, 83–112. [Google Scholar] [CrossRef]
- Yalcin, M.; Mermer, S.; Kozaci, L.D.; Turgut, C. Insecticide resistance in two populations of Tuta absoluta (Meyrick, 1917)(Lepidoptera: Gelechiidae) from Turkey. Türkiye Entomol. Derg. 2015, 39, 137–145. [Google Scholar]
- Sridhar, V.; Onkara naik, S.; Nitin, K.S.; Ashokan, R.; Swathi, P.; Gadad, H. Efficacy of integrated pest management tools evaluated against Tuta absoluta (Meyrick) on tomato in India. J. Biol. Control 2019, 33, 264–270. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.A.; Tsagkarakou, A.; Vontas, J.; Nauen, R. Insecticide resistance in the tomato pinworm Tuta absoluta: Patterns, spread, mechanisms, management and outlook. J. Pest Sci. 2019, 92, 1329–1342. [Google Scholar] [CrossRef]
- Santana, P.A.; Kumar, L.; Da Silva, R.S.; Picanço, M.C. Global geographic distribution of Tuta absoluta as affected by climate change. J. Pest Sci. 2019, 92, 1373–1385. [Google Scholar] [CrossRef]
- Silva, G.A.; Picanço, M.C.; Bacci, L.; Crespo, A.L.B.; Rosado, J.F.; Guedes, R.N.C. Control failure likelihood and spatial dependence of insecticide resistance in the tomato pinworm, Tuta absoluta. Pest Manag. Sci. 2011, 67, 913–920. [Google Scholar] [CrossRef]
- Cocco, A.; Deliperi, S.; Delrio, G. Control of Tuta absoluta (Meyrick)(Lepidoptera: Gelechiidae) in greenhouse tomato crops using the mating disruption technique. J. Appl. Entomol. 2013, 137, 16–28. [Google Scholar] [CrossRef]
- Bawin, T.; Dujeu, D.; De Backer, L.; Francis, F.; Verheggen, F.J. Ability of Tuta absoluta (Lepidoptera: Gelechiidae) to develop on alternative host plant species. Can. Entomol. 2016, 148, 434–442. [Google Scholar] [CrossRef]
- Palacios, M.; Cisneros, F. Management of the potato tuber moth. Program 4. In Integrated Pest Management; Program Report; International Potato Center: Lima, Peru, 1995; pp. 87–91. [Google Scholar]
- Vallejo Cabrera, F.A. Mejoramiento Genético y Producción de Tomate en Colombia; Universidad Nacional de Colombia: Bogotá, Colombia, 1999. [Google Scholar]
- Moore, M.E.; Hill, C.A.; Kingsolver, J.G. Differing thermal sensitivities in a host–parasitoid interaction: High, fluctuating developmental temperatures produce dead wasps and giant caterpillars. Funct. Ecol. 2021, 35, 675–685. [Google Scholar] [CrossRef]
- Bai, B.; Smith, S.M. Effect of host availability on reproduction and survival of the parasitoid wasp Trichogramma minutum. J. Ecol. Antomol. 1993, 18, 279–286. [Google Scholar] [CrossRef]
- Hohmann, C.L.; Luck, R.F. Effect of host availability and egg load in Trichogramma platneri Nagarkatti (Hymenoptera: Trichogrammatidae) and its consequences on progeny quality. Braz. Arch. Biol. Technol. 2004, 47, 413–422. [Google Scholar] [CrossRef]
- Iverson, A.L.; Gonthier, D.J.; Pak, D.; Ennis, K.K.; Burnham, R.J.; Perfecto, I.; Rodriguez, M.R.; Vandermeer, J.H. A multifunctional approach for achieving simultaneous biodiversity conservation and farmer livelihood in coffee agroecosystems. Biol. Conserv. 2019, 238, 108179. [Google Scholar] [CrossRef]
- Agboka, K.M.; Tonnang, H.; Abdel-Rahman, E.; Kimathi, E.; Mutanga, O.; Odindi, J.; Niassy, S.; Mohamed, S.; Ekesi, S. A systematic methodological approach to estimate the impacts of a classical biological control agents dispersal at landscape: Application to fruit fly Bactrocera dorsalis and its endoparasitoid Fopius arisanus. Authorea, 2022; Preprint. Available online: https://europepmc.org/article/ppr/ppr475970(accessed on 10 April 2022).
- Jamielniak, J.A. A mathematical approach to study stress-related behaviors in captive golden-bellied capuchins (Sapajus xanthosthernos). Comput. Ecol. Softw. 2016, 6, 83. [Google Scholar]
- Zadeh, L.A. Fuzzy sets. Inf. Control 1965, 8, 338–353. [Google Scholar] [CrossRef]
- Bone, C.; Dragicevic, S.; Roberts, A. Integrating high resolution remote sensing, GIS and fuzzy set theory for identifying susceptibility areas of forest insect infestations. Int. J. Remote Sens. 2005, 26, 4809–4828. [Google Scholar] [CrossRef]
- Garcia, A.G.; Diniz, A.J.F.; Parra, J.R.P. A fuzzy-based index to identify suitable areas for host-parasitoid interactions: Case study of the Asian citrus psyllid Diaphorina citri and its natural enemy Tamarixia radiata. Biol. Control 2019, 135, 135–140. [Google Scholar] [CrossRef]
- Louis, G.N.; Aloo, F.; Were, K.; Kebeney, J.K.; Kibwage, J.; Sikei, G.; Wokabi, S.; Paul, M.N. Land, agriculture and livestock. no. GoK 2007. 2011; pp. 108–123. Available online: https://www.nema.go.ke/images/Docs/Regulations/KenyaSoECh6.pdf (accessed on 10 January 2022).
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Bajonero, J.; Córdoba, N.; Cantor, F.; Rodríguez, D.; Cure, J.R. Biología y ciclo reproductivo de Apanteles gelechiidivoris (Hymenoptera: Braconidae), parasitoide de Tuta absoluta (Lepidoptera: Gelechiidae). Agron. Colomb. 2008, 26, 417–426. [Google Scholar]
- Cely Pardo, N.L. Determinación de Niveles de daño Ocasionados por Diferentes Densidades de Población de Tuta Absoluta (Lepidoptera: Gelechiidae) en Tomate Bajo Invernadero; CO-BAC: Bogotá, Colombia, 2006. [Google Scholar]
- Kinyanjui, G.; Khamis, F.M.; Ombura, F.L.O.; Kenya, E.U.; Ekesi, S.; Mohamed, S.A. Distribution, abundance and natural enemies of the invasive tomato leafminer, Tuta absoluta (Meyrick) in Kenya. Bull. Entomol. Res. 2021, 111, 658–673. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Booth, T.H. Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important. Austral Ecol. 2018, 43, 852–860. [Google Scholar] [CrossRef]
- Aigbedion-Atalor, P.; Hill, M.P.; Azrag, A.G.A.; Zalucki, M.P.; Mohamed, S.A. Disentangling thermal effects using life cycle simulation modelling on the biology and demographic parameters of Dolichogenidea gelechiidivoris, a Parasitoid of Tuta Absoluta. J. Therm. Biol. 2022, 107, 103260. [Google Scholar] [CrossRef] [PubMed]
- Salehie, O.; Hamed, M.M.; bin Ismail, T.; Shahid, S. Projection of droughts in Amu Darya River Basin for shared socioeconomic pathways. Res. Sq. 2021; preprint. Available online: https://www.researchsquare.com/article/rs-1088081/v1(accessed on 10 April 2022).
- Hemati, M.; Hasanlou, M.; Mahdianpari, M.; Mohammadimanesh, F. A systematic review of landsat data for change detection applications: 50 years of monitoring the earth. Remote Sens. 2021, 13, 2869. [Google Scholar] [CrossRef]
- Young, N.E.; Anderson, R.S.; Chignell, S.M.; Vorster, A.G.; Lawrence, R.; Evangelista, P.H. A survival guide to Landsat preprocessing. Ecology 2017, 98, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.3–7. 2020. Available online: https://cran.r-project.org/package=raster (accessed on 10 January 2022).
- Leroy, B.; Meynard, C.N.; Bellard, C.; Courchamp, F. virtualspecies, an R package to generate virtual species distributions. Ecography 2016, 39, 599–607. [Google Scholar] [CrossRef]
- Perfilieva, I. Fuzzy IF-THEN rules from logical point of view. In Computational Intelligence, Theory and Applications; Springer: Berlin/Heidelberg, Germany, 2006; pp. 691–697. [Google Scholar]
- Keshwani, D.R.; Jones, D.D.; Meyer, G.E.; Brand, R.M. Rule-based Mamdani-type fuzzy modeling of skin permeability. Appl. Soft Comput. 2008, 8, 285–294. [Google Scholar] [CrossRef]
- Zadeh, L.A. Outline of a new approach to the analysis of complex systems and decision processes. IEEE Trans. Syst. Man. Cybern. 1973, 1, 28–44. [Google Scholar] [CrossRef]
- Perfilieva, I. Analytical theory of fuzzy if-then rules with compositional rule of inference. In Fuzzy Logic; Springer: Berlin/Heidelberg, Germany, 2007; pp. 174–191. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System; Open Source Geospatial Foundation: Chicago, IL, USA, 2009. [Google Scholar]
- Mama Sambo, S.; Ndlela, S.; du Plessis, H.; Obala, F.; Mohamed, S.A. Ratio dependence effects of the parasitoid Dolichogenidea gelechiidivoris on its associated host Tuta absoluta. Biocontrol Sci. Technol. 2022, 32, 497–510. [Google Scholar] [CrossRef]
- Groth, M.Z.; Loeck, A.E.; Nornberg, S.D.; Bernardi, D.; Nava, D.E. Biology and thermal requirements of Fopius arisanus (Sonan, 1932) (Hymenoptera: Braconidae) reared on Ceratitis capitata eggs (Wiedemann) (Diptera: Tephritidae). Neotrop. Entomol. 2017, 46, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Silva-Torres, C.S.A.; Matthews, R.W. Development of Melittobia australica Girault and M. digitata Dahms (Parker) (Hymenoptera: Eulophidae) parasitizing Neobellieria bullata (Parker) (Diptera: Sarcophagidae) puparia. Neotrop. Entomol. 2003, 32, 645–651. [Google Scholar] [CrossRef]
- Sow, G.; Arvanitakis, L.; Niassy, S.; Diarra, K.; Bordat, D. Performance of the parasitoid Oomyzus sokolowskii (Hymenoptera: Eulophidae) on its host Plutella xylostella (Lepidoptera: Plutellidae) under laboratory conditions. Int. J. Trop. Insect Sci. 2013, 33, 38–45. [Google Scholar] [CrossRef]
- Le Provost, G.; Thiele, J.; Westphal, C.; Penone, C.; Allan, E.; Neyret, M.; van der Plas, F.; Ayasse, M.; Bardgett, R.D.; Birkhofer, K.; et al. Contrasting responses of above-and belowground diversity to multiple components of land-use intensity. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Foba, C.N.; Akutse, K.S.; Fiaboe, K.K.M.; Lagat, Z.O.; Gitonga, L.M. Interaction between Phaedrotoma scabriventris Nixon and Opius dissitus Muesebeck (Hymenoptera: Braconidae): Endoparasitoids of Liriomyza leafminer. Afr. Entomol. 2015, 23, 120–131. [Google Scholar] [CrossRef]
- Nderitu, P.W.; Muturi, J.; Otieno, M.; Arunga, E.E.; Mattias, J. Tomato leafminer (Tuta absoluta) (Meyrick 1917) (Lepidoptera: Gelechiidae) prevalence and farmer management practices in Kirinyanga County, Kenya. J. Entomol. Nematol. 2018, 10, 43–49. [Google Scholar] [CrossRef]
- Usery, E.L.; Finn, M.P.; Scheidt, D.J.; Ruhl, S.; Beard, T.; Bearden, M. Geospatial data resampling and resolution effects on watershed modeling: A case study using the agricultural non-point source pollution model. J. Geogr. Syst. 2004, 6, 289–306. [Google Scholar] [CrossRef]
- Bell, J.R.; Aralimarad, P.; Lim, K.-S.; Chapman, J.W. Predicting insect migration density and speed in the daytime convective boundary layer. PLoS ONE 2013, 8, e54202. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Tobin, P.C. Population ecology of insect invasions and their management. Annu. Rev. Entomol. 2008, 53, 387–408. [Google Scholar] [CrossRef]
- Ibrahim, E.A.; Salifu, D.; Mwalili, S.; Dubois, T.; Collins, R.; Tonnang, H.E.Z. An expert system for insect pest population dynamics prediction. Comput. Electron. Agric. 2022, 198, 107124. [Google Scholar] [CrossRef]
- Center, B.; Verma, B.P. Fuzzy logic for biological and agricultural systems. In Artificial Intelligence for Biology and Agriculture; Springer: Berlin/Heidelberg, Germany, 1998; pp. 213–225. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Months 1 | Western | Nyanza | Rift Valley | Central | Eastern | Coast |
|---|---|---|---|---|---|---|
| January | 21.0 | 23.6 | 18.9 | 19.3 | 19.6 | 27.9 |
| February | 21.9 | 24.6 | 19.5 | 20.0 | 20.5 | 28.1 |
| March | 21.6 | 23.9 | 19.3 | 19.8 | 20.4 | 28.7 |
| April | 20.5 | 22.8 | 18.1 | 18.6 | 19.5 | 28.1 |
| May | 19.5 | 22.5 | 17.2 | 17.7 | 18.7 | 26.6 |
| June | 18.6 | 22.4 | 16.4 | 16.8 | 17.8 | 25.9 |
| July | 18.2 | 22.5 | 16.1 | 16.3 | 17.2 | 25.4 |
| August | 18.3 | 22.7 | 16.5 | 16.7 | 17.6 | 25.5 |
| September | 19.0 | 23.2 | 17.5 | 17.9 | 18.8 | 26.2 |
| October | 19.7 | 23.2 | 18.3 | 18.6 | 19.6 | 26.8 |
| November | 19.8 | 22.7 | 17.7 | 17.8 | 18.7 | 27.2 |
| December | 20.2 | 22.9 | 18.1 | 18.2 | 18.7 | 27.7 |
| Temperature Threshold | Net Reproduction Rate (R0) | Fuzzy Partition Variable Names |
|---|---|---|
| 10–15 °C | 0.13–1.55 | Suboptimal lower temperature threshold |
| 20 and 25 °C | 15–14 | Optimal |
| 30–35 °C | 2.18–0.06 | Suboptimal higher temperature threshold |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agboka, K.M.; Tonnang, H.E.Z.; Abdel-Rahman, E.M.; Odindi, J.; Mutanga, O.; Mohamed, S.A. A Fuzzy-Based Model to Predict the Spatio-Temporal Performance of the Dolichogenidea gelechiidivoris Natural Enemy against Tuta absoluta under Climate Change. Biology 2022, 11, 1280. https://doi.org/10.3390/biology11091280
Agboka KM, Tonnang HEZ, Abdel-Rahman EM, Odindi J, Mutanga O, Mohamed SA. A Fuzzy-Based Model to Predict the Spatio-Temporal Performance of the Dolichogenidea gelechiidivoris Natural Enemy against Tuta absoluta under Climate Change. Biology. 2022; 11(9):1280. https://doi.org/10.3390/biology11091280
Chicago/Turabian StyleAgboka, Komi Mensah, Henri E. Z. Tonnang, Elfatih M. Abdel-Rahman, John Odindi, Onisimo Mutanga, and Samira A. Mohamed. 2022. "A Fuzzy-Based Model to Predict the Spatio-Temporal Performance of the Dolichogenidea gelechiidivoris Natural Enemy against Tuta absoluta under Climate Change" Biology 11, no. 9: 1280. https://doi.org/10.3390/biology11091280
APA StyleAgboka, K. M., Tonnang, H. E. Z., Abdel-Rahman, E. M., Odindi, J., Mutanga, O., & Mohamed, S. A. (2022). A Fuzzy-Based Model to Predict the Spatio-Temporal Performance of the Dolichogenidea gelechiidivoris Natural Enemy against Tuta absoluta under Climate Change. Biology, 11(9), 1280. https://doi.org/10.3390/biology11091280