

Fossil Biomarkers and Biosignatures Preserved in Coprolites Reveal Carnivorous Diets in the Carboniferous Mazon Creek Ecosystem

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
Palaeoenvironmental Setting
2. Materials and Methods
2.1. Sample Preparation and Extraction
2.2. Gas Chromatography-Mass Spectrometry (GC-MS)
2.3. Gas Chromatography-Isotope Ratio-Mass Spectrometry (GC-irMS)
2.4. Bulk Stable Carbon Isotopes
2.5. Polar Compound Analysis
2.6. In Situ Raman Microspectroscopy and ChemoSpace Analysis of Spectral Data
2.7. X-ray Diffraction
2.8. Elemental Analysis
2.9. Total Organic Carbon
3. Results and Discussion
3.1. Inorganic Composition
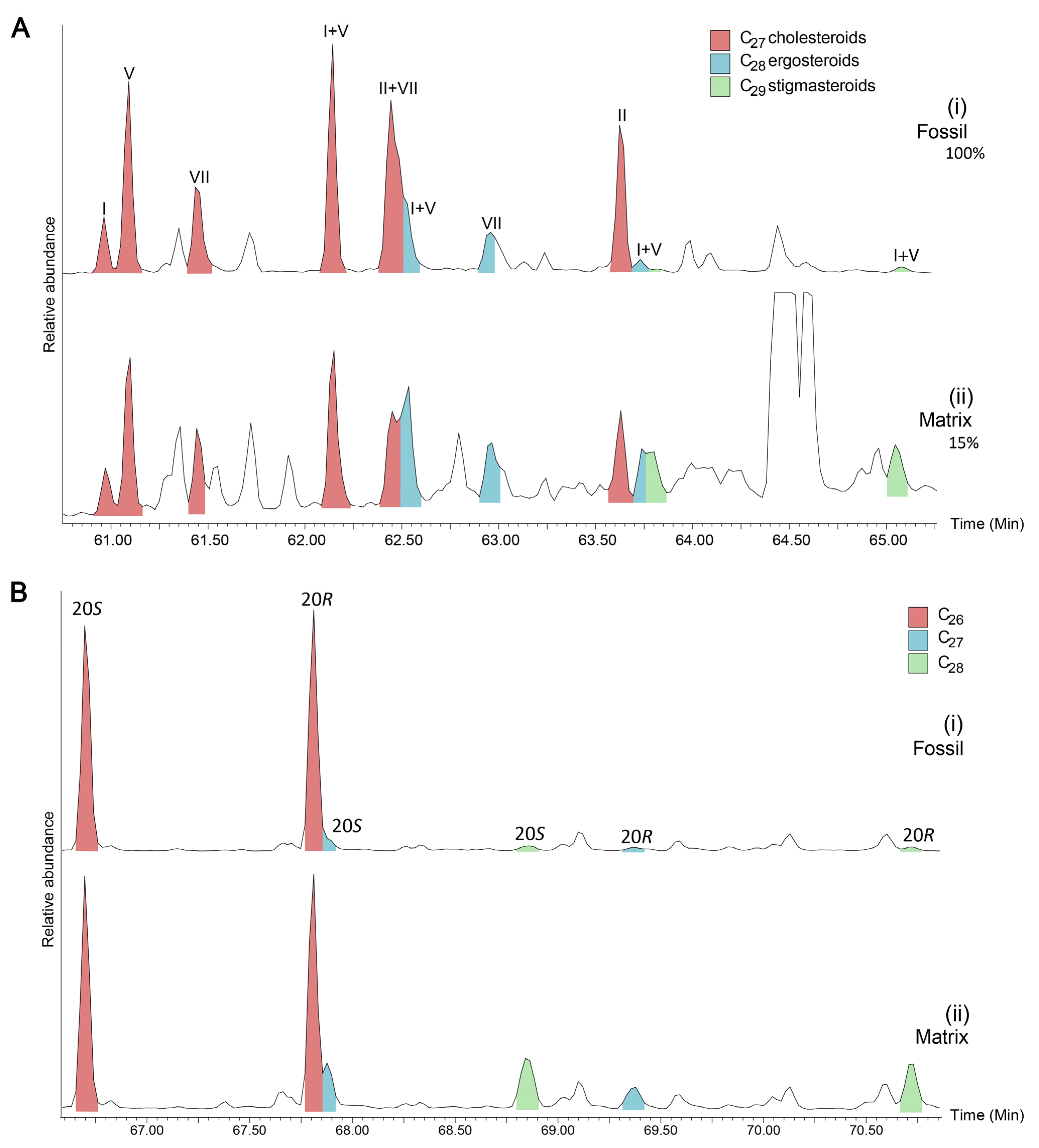
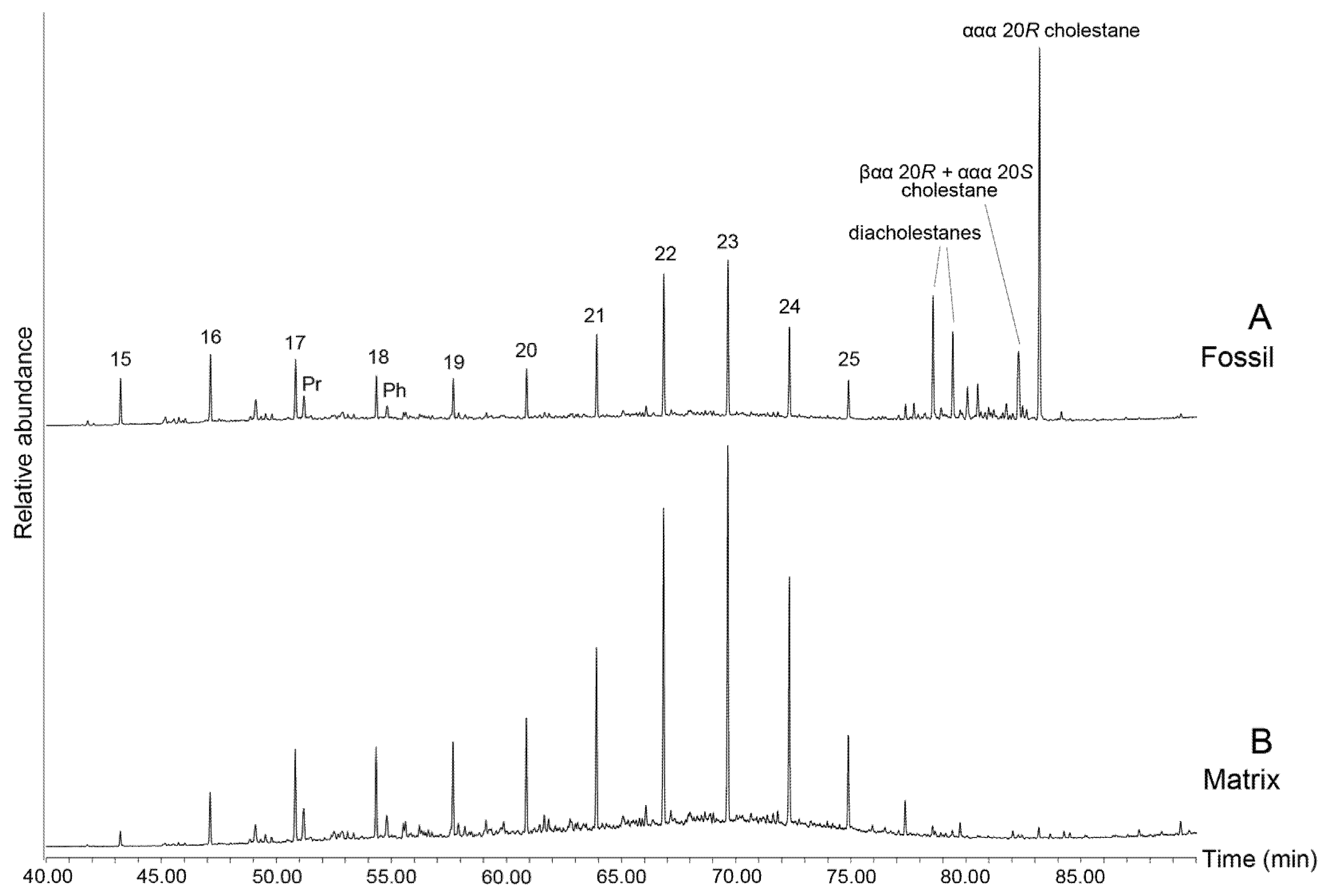
3.2. Lipid Biomarkers of Coprolites
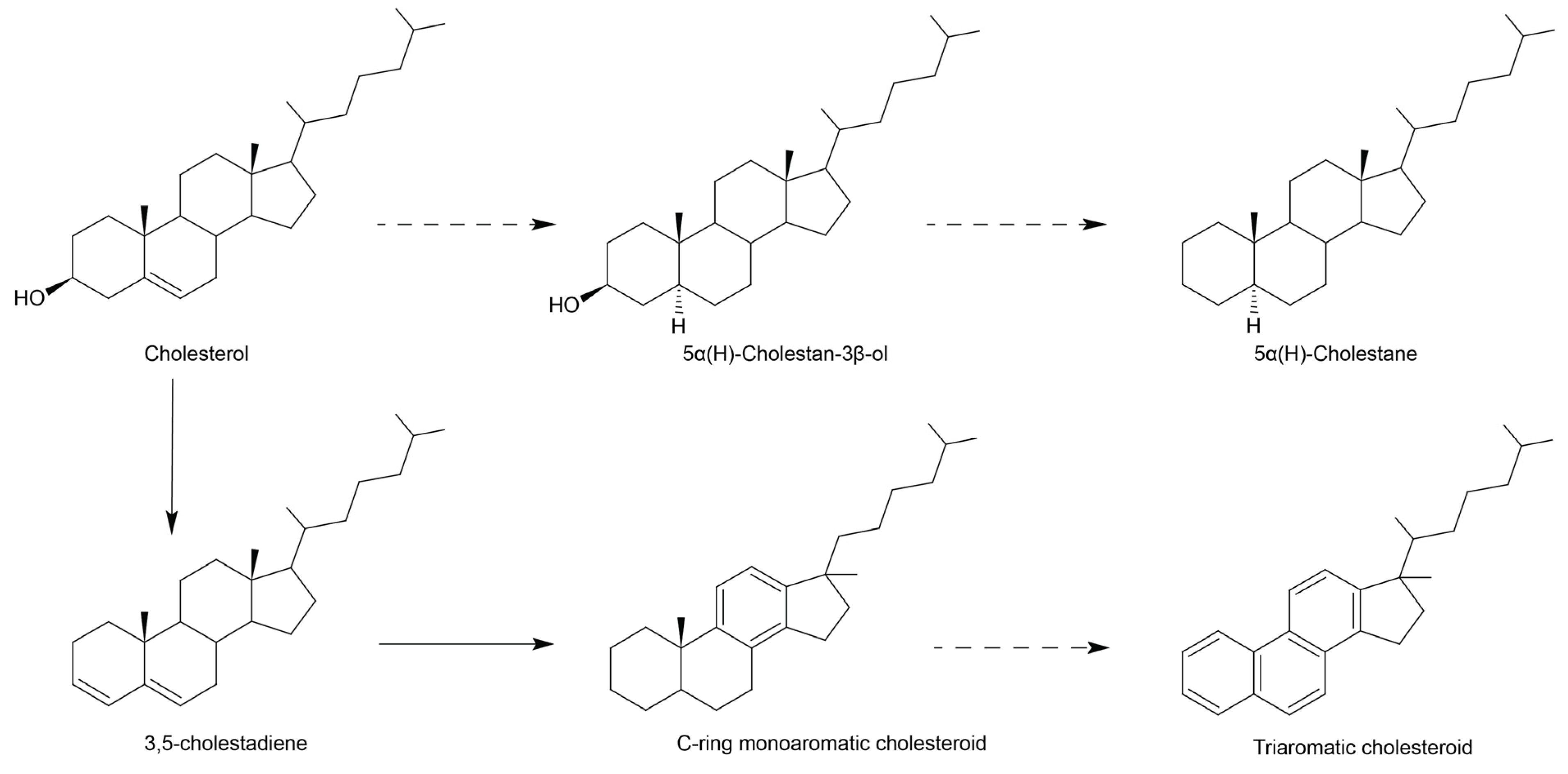
3.3. Early Diagenetic Transformation of Dietary Sterols
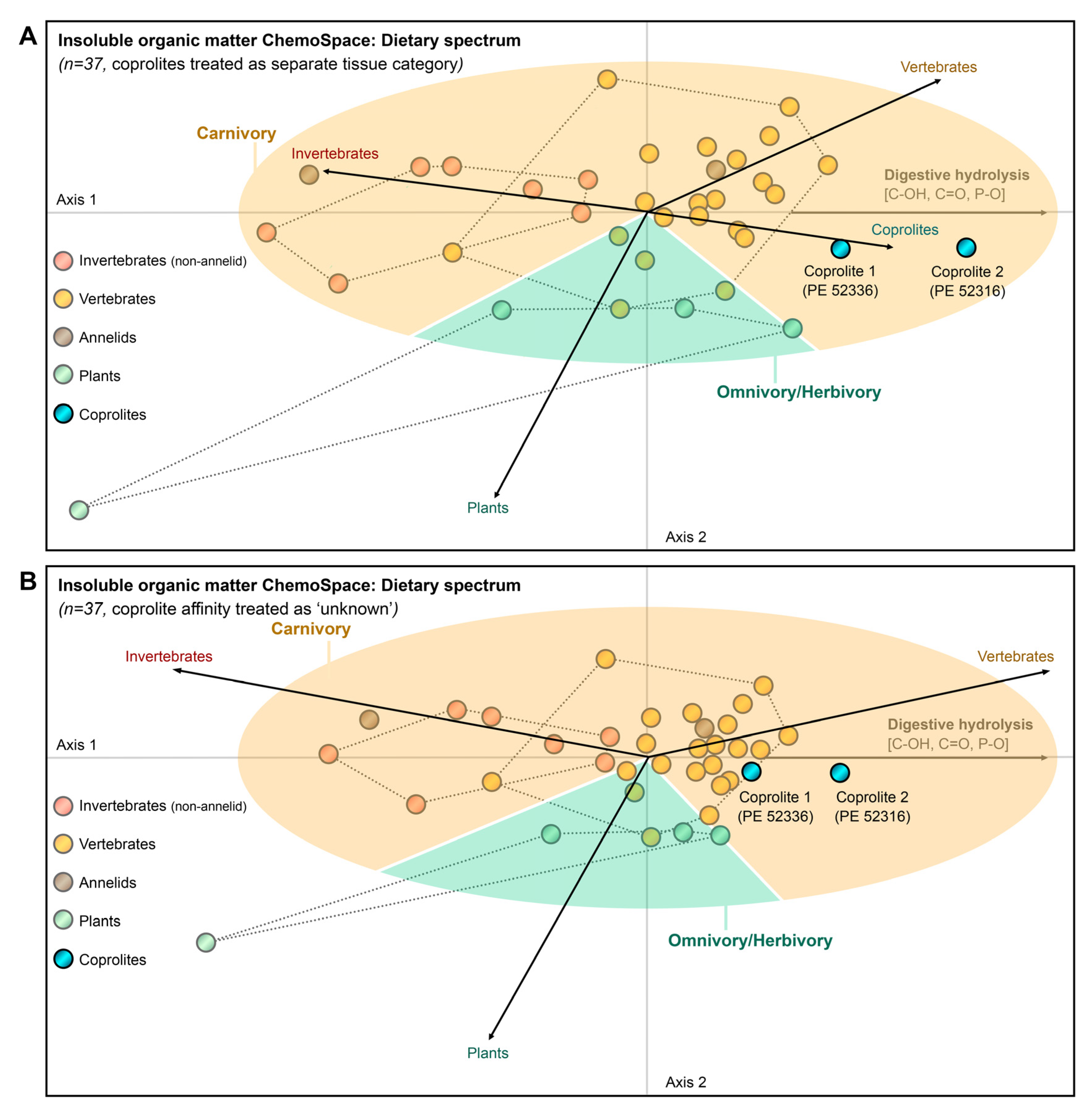
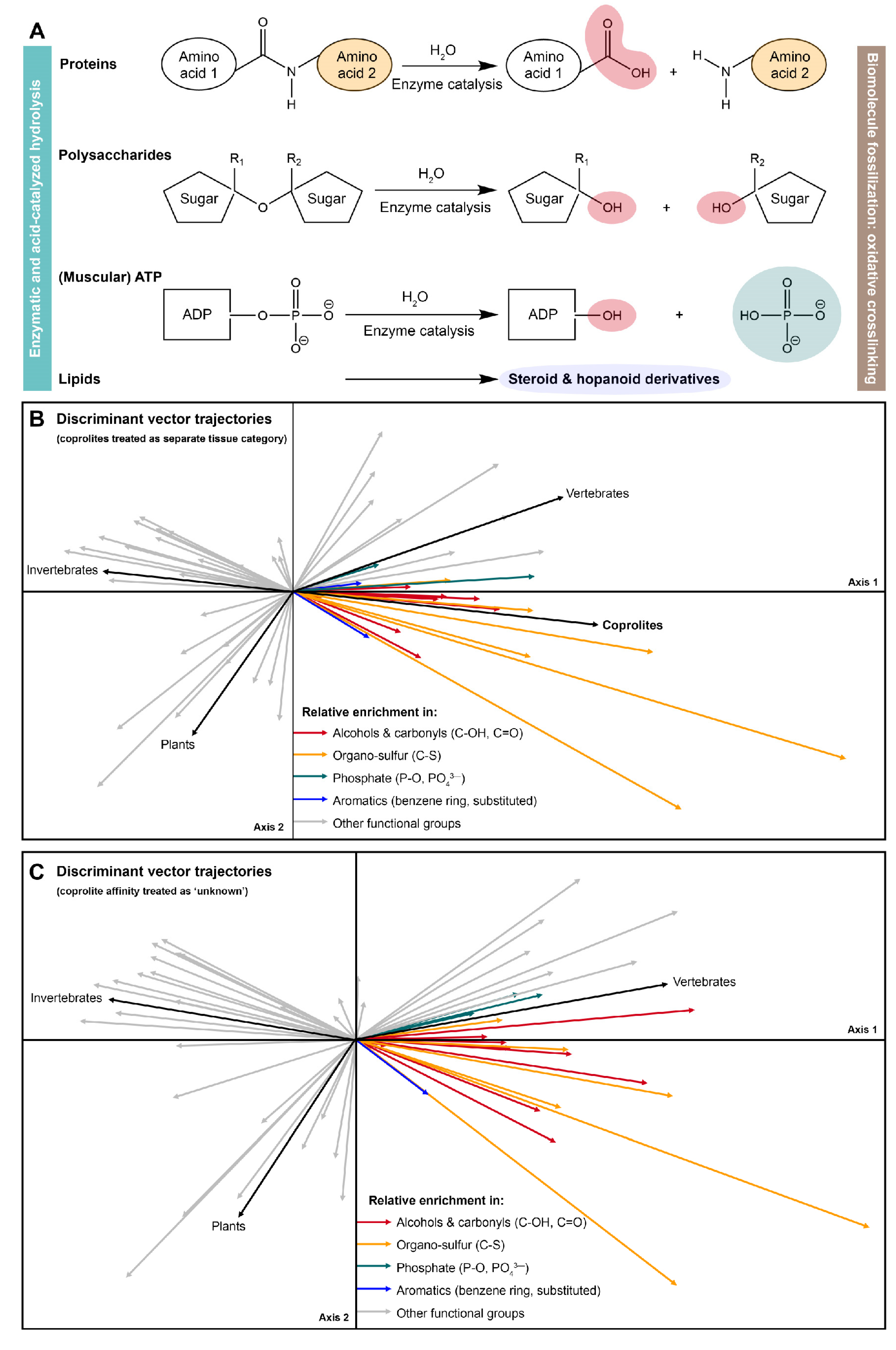
3.4. Raman ChemoSpace
3.5. Lipid Biomarkers of Matrix (Palaeoenvironmental Signal)
3.6. Stable Carbon Isotopes
4. Conclusions
- (1)
- Cholesteroids including intact 5α-cholestan-3β-ol and coprostanol have been preserved in siderite concretions hosting 306 million-year-old coprolites.
- (2)
- The molecular data obtained by GC-MS, GC-MRM, GC-irMS and Raman microspectroscopy supports a primarily carnivorous diet and suggest an elevated trophic position for the coprolite producer.
- (3)
- The preservation of intact dietary sterols and macromolecular biosignatures is attributed to rapid encapsulation of the coprolites within days to months after egestion.
- (4)
- Siderite (FeCO3) concretions (of Carboniferous age) seem to preserve intact and modified, but not unrecognisable, biomolecules much like calcium carbonate concretions of Jurassic and Devonian ages.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hollocher, K.T.; Alcober, O.A.; Colombi, C.E.; Hollocher, T.C. Carnivore Coprolites from the Upper Triassic Ischigualasto Formation, Argentina: Chemistry, Mineralogy, and Evidence for Rapid Initial Mineralization. Palaios 2005, 20, 51–63. [Google Scholar] [CrossRef]
- Chin, K. The Paleobiological Implications of Herbivorous Dinosaur Coprolites from the Upper Cretaceous Two Medicine Formation of Montana: Why Eat Wood? Palaios 2007, 22, 554–566. [Google Scholar] [CrossRef]
- Gill, F.L.; Crump, M.P.; Schouten, R.; Bull, I.D. Lipid analysis of a ground sloth coprolite. Quat. Res. 2009, 72, 284–288. [Google Scholar] [CrossRef]
- Hollocher, K.T.; Hollocher, T.C.; Rigby, J.K. A Phosphatic Coprolite Lacking Diagenetic Permineralization from the Upper Cretaceous Hell Creek Formation, Northeastern Montana: Importance of Dietary Calcium Phosphate in Preservation. Palaios 2010, 25, 132–140. [Google Scholar] [CrossRef]
- Khosla, A.; Chin, K.; Alimohammadin, H.; Dutta, D. Ostracods, plant tissues, and other inclusions in coprolites from the Late Cretaceous Lameta Formation at Pisdura, India: Taphonomical and palaeoecological implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 418, 90–100. [Google Scholar] [CrossRef]
- Northwood, C. Early Triassic coprolites from Australia and their palaeobiological significance. Palaeontology 2005, 48, 49–68. [Google Scholar] [CrossRef]
- Bajdek, P.; Owocki, K.; Niedźwiedzki, G. Putative dicynodont coprolites from the Upper Triassic of Poland. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 411, 1–17. [Google Scholar] [CrossRef]
- Boast, A.P.; Weyrich, L.S.; Wood, J.R.; Metcalf, J.L.; Knight, R.; Cooper, A. Coprolites reveal ecological interactions lost with the extinction of New Zealand birds. Proc. Natl. Acad. Sci. USA 2018, 115, 1546–1551. [Google Scholar] [CrossRef]
- Witt, K.E.; Yarlagadda, K.; Allen, J.M.; Bader, A.C.; Simon, M.L.; Kuehn, S.R.; Swanson, K.S.; Cross, T.-W.L.; Hedman, K.M.; Ambrose, S.H.; et al. Integrative analysis of DNA, macroscopic remains and stable isotopes of dog coprolites to reconstruct community diet. Sci. Rep. 2021, 11, 3113. [Google Scholar] [CrossRef]
- Lengger, S.K.; Melendez, I.M.; Summons, R.E.; Grice, K. Mudstones and embedded concretions show differences in lithology-related, but not source-related biomarker distributions. Org. Geochem. 2017, 113, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Melendez, I.; Grice, K.; Trinajstic, K.; Ladjavardi, M.; Greenwood, P.; Thompson, K. Biomarkers reveal the role of photic zone euxinia in exceptional fossil preservation: An organic geochemical perspective. Geology 2013, 41, 123–126. [Google Scholar] [CrossRef]
- Plet, C.; Grice, K.; Pagès, A.; Verrall, M.; Coolen, M.J.L.; Ruebsam, W.; Rickard, W.D.A.; Schwark, L. Palaeobiology of red and white blood cell-like structures, collagen and cholesterol in an ichthyosaur bone. Sci. Rep. 2017, 7, 13776. [Google Scholar] [CrossRef] [PubMed]
- Allison, P.A. Konservat-Lagerstatten: Cause and Classification. Paleobiology 1988, 14, 331–344. [Google Scholar] [CrossRef]
- Briggs, D.E.G. The role of decay and mineralization in the preservation of soft-bodied fossils. Annu. Rev. Earth Planet. Sci. 2003, 31, 275–301. [Google Scholar] [CrossRef]
- Parry, L.A.; Smithwick, F.; Nordén, K.K.; Saitta, E.T.; Lozano-Fernandez, J.; Tanner, A.R.; Caron, J.B.; Edgecombe, G.D.; Briggs, D.E.; Vinther, J. Soft-Bodied Fossils Are Not Simply Rotten Carcasses—Toward a Holistic Understanding of Exceptional Fossil Preservation. Bioessays 2018, 40, 1700167. [Google Scholar] [CrossRef]
- Seilacher, A. Begriff und bedeutung der Fossil-Lagerstätten: Neues Jarhbuch für Geologie und Paläontologie. Neues Jahrb. Geol. Paläontologie Mon. 1970, 34–39. [Google Scholar]
- Grice, K.; Holman, A.I.; Plet, C.; Tripp, M. Fossilised Biomolecules and Biomarkers in Carbonate Concretions from Konservat-Lagerstätten. Minerals 2019, 9, 158. [Google Scholar] [CrossRef]
- Mccoy, V.E. Concretions as agents of soft-tissue preservation: A review. Paleontol. Soc. Pap. 2014, 20, 147–162. [Google Scholar] [CrossRef]
- Sagemann, J.; Bale, S.J.; Briggs, D.E.; Parkes, R.J. Controls on the formation of authigenic minerals in association with decaying organic matter: An experimental approach. Geochim. Cosmochim. Acta 1999, 63, 1083–1095. [Google Scholar]
- Peters, K.E.; Walters, C.C.; Moldowan, J.M. Volume I: Biomarkers and Isotopes in the Environment and Human History. In The Biomarker Guide; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Volkman, J.K. A review of sterol markers for marine and terrigenous organic matter. Org. Geochem. 1986, 9, 83–99. [Google Scholar] [CrossRef]
- Huang, W.Y.; Meinschein, W.G. Sterols as source indicators of organic materials in sediments. Geochim. Cosmochim. Acta 1976, 40, 323–330. [Google Scholar]
- Mackenzie, A.S.; Brassel, S.C.; Eglinton, G.; Maxwell, J.R. Chemical fossils: The geological fate of steroids. Science 1982, 217, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Brassell, S.C.; McEvoy, J.; Hoffmann, C.F.; Lamb, N.A.; Peakman, T.M.; Maxwell, J.R. Isomerisation, rearrangement and aromatisation of steroids in distinguishing early stages of diagenesis. Org. Geochem. 1984, 6, 11–23. [Google Scholar] [CrossRef]
- Mackenzie, A.S.; Lamb, N.A.; Maxwell, J.R. Steroid hydrocarbons and the thermal history of sediments. Nature 1982, 295, 223–226. [Google Scholar] [CrossRef]
- Melendez, I.; Grice, K.; Schwark, L. Exceptional preservation of Palaeozoic steroids in a diagenetic continuum. Sci. Rep. 2013, 3, 2768. [Google Scholar] [CrossRef]
- Bull, I.D.; Lockheart, M.J.; Elhmmali, M.M.; Roberts, D.J.; Evershed, R.P. The origin of faeces by means of biomarker detection. Environ. Int. 2002, 27, 647–654. [Google Scholar] [CrossRef]
- Gill, F.L.; Bull, I.D. Lipid analysis of vertebrate coprolites. N. M. Mus. Nat. Hist. Sci. Bull. 2012, 57, 93–98. [Google Scholar]
- Umamaheswaran, R.; Prasad, G.V.R.; Rudra, A.; Dutta, S. Biomarker Signatures in Triassic Coprolites. Palaios 2019, 34, 458–467. [Google Scholar] [CrossRef]
- Weber, D.J.; Lawler, G.C. Lipid Components of the Coprolites. Geology, Paleontology and Paleoecology of a Late Triassic Lake, Western New Mexico. Brigh. Young Univ. Geol. Stud. 1978, 25, 75–87. [Google Scholar]
- Zatoń, M.; Niedźwiedzki, G.; Marynowski, L.; Benzerara, K.; Pott, C.; Cosmidis, J.; Krzykawski, T.; Filipiak, P. Coprolites of Late Triassic carnivorous vertebrates from Poland: An integrative approach. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 430, 21–46. [Google Scholar] [CrossRef]
- Harrault, L.; Milek, K.; Jardé, E.; Jeanneau, L.; Derrien, M.; Anderson, D.G. Faecal biomarkers can distinguish specific mammalian species in modern and past environments. PLoS ONE 2019, 14, e0211119. [Google Scholar] [CrossRef] [PubMed]
- Leeming, R.; Ball, A.; Ashbolt, N.; Nichols, P. Using faecal sterols from humans and animals to distinguish faecal pollution in receiving waters. Water Res. 1996, 30, 2893–2900. [Google Scholar] [CrossRef]
- McCoy, V.E.; Wiemann, J.; Lamsdell, J.C.; Whalen, C.D.; Lidgard, S.; Mayer, P.; Petermann, H.; Briggs, D.E. Chemical signatures of soft tissues distinguish between vertebrates and invertebrates from the Carboniferous Mazon Creek Lagerstatte of Illinois. Geobiology 2020, 18, 560–565. [Google Scholar] [CrossRef]
- Sistiaga, A.; Berna, F.; Laursen, R.; Goldberg, P. Steroidal biomarker analysis of a 14,000 years old putative human coprolite from Paisley Cave, Oregon. J. Archaeol. Sci. 2014, 41, 813–817. [Google Scholar] [CrossRef]
- Clements, T.; Purnell, M.; Gabbott, S. The Mazon Creek Lagerstätte: A diverse late Paleozoic ecosystem entombed within siderite concretions. J. Geol. Soc. 2018, 176, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wittry, J. The Mazon Creek Fossil Fauna; ESCONI: Downers Grove, IL, USA, 2012. [Google Scholar]
- Peppers, R.A. Palynological Correlation of Major Pennsylvanian (Middle and Upper Carboniferous) Chronostratigraphic Boundaries in the Illinois and Other Coal Basins; The Geological Society of America, Inc.: Boulder, CO, USA, 1996. [Google Scholar]
- Pfefferkorn, H.W. High diversity and stratigraphic age of the Mazon Creek flora. In Mazon Creek Fossils; Nitecki, M.H., Ed.; Academic Press, Inc.: New York, NY, USA, 1979; pp. 129–142. [Google Scholar]
- Baird, G.C. Lithology and fossil distribution, Francis Creek Shale in northeastern Illinois. In Mazon Creek Fossils; Nitecki, M.H., Ed.; Academic Press, Inc.: New York, NY, USA, 1979; pp. 41–67. [Google Scholar]
- Baird, G.C.; Shabica, C.W.; Anderson, J.L.; Richardson, E.S. Biota of a Pennsylvanian muddy coast: Habitats within the Mazonian delta complex, northeast Illinois. J. Paleontol. 1985, 59, 253–281. [Google Scholar]
- Baird, G.C.; Sroka, S.D.; Shabica, C.W.; Keucher, G.J. Taphonomy of Middle Pennsylvanian Mazon Creek area fossil localities, Northeast Illinois; significance of exceptional fossil preservation in syngenetic concretions. Palaios 1986, 1, 271–285. [Google Scholar] [CrossRef]
- Cotroneo, S.; Schiffbauer, J.D.; McCoy, V.E.; Wortmann, U.G.; Darroch, S.A.F.; Peng, Y.; Laflamme, M. A new model of the formation of Pennsylvanian iron carbonate concretions hosting exceptional soft-bodied fossils in Mazon Creek, Illinois. Geobiology 2016, 14, 543–555. [Google Scholar] [CrossRef]
- Shabica, C.W. Pennsylvanian sedimentation in Northern Illinois: Examination of delta models. In Mazon Creek Fossils; Nitecki, M.H., Ed.; Academic Press, Inc.: New York, NY, USA, 1979. [Google Scholar]
- Baird, G.C.; Sroka, S.D.; Shabica, C.W.; Beard, T.L. Mazon Creek-type fossil assemblages in the U.S. midcontinent Pennsylvanian: Their recurrent character and palaeoenvironmental significance. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1985, 311, 87–99. [Google Scholar]
- Berner, R.A. A new geochemical classification of sedimentary environments. J. Sediment. Petrol. 1981, 51, 359–365. [Google Scholar]
- Berner, R.A. Sulphate reduction, organic matter decomposition and pyrite formation. Philos. Trans. R. Soc. Lond. Ser. A Math. Phys. Sci. 1985, 315, 25–38. [Google Scholar]
- Claypool, G.E.; Kaplan, I.R. The Origin and Distribution of Methane in Marine Sediments. In Natural Gases in Marine Sediments; Maine Science; Kaplan, I.R., Ed.; Springer: Boston, MA, USA, 1974; Volume 3, pp. 99–139. [Google Scholar]
- Allison, P.A. The role of anoxia in the decay and mineralization of proteinaceous macro-fossils. Paleobiology 1988, 14, 139–154. [Google Scholar] [CrossRef]
- Berner, R.A. Rate of concretion growth. Geochim. Cosmochim. Acta 1968, 32, 477–483. [Google Scholar] [CrossRef]
- Briggs, D.E.; Kear, A.J. Decay and preservation of polychaetes: Taphonomic thresholds in soft-bodied organisms. Paleobiology 1993, 19, 107–135. [Google Scholar] [CrossRef]
- McCoy, V.E.; Young, R.T.; Briggs, D.E.G. Sediment Permeability and the Preservation of Soft-Tissues in Concretions: An Experimental Study. Palaios 2015, 30, 608–612. [Google Scholar] [CrossRef]
- Yoshida, H.; Ujihara, A.; Minami, M.; Asahara, Y.; Katsuta, N.; Yamamoto, K.; Sirono, S.; Maruyama, I.; Nishimoto, S.; Metcalfe, R. Early post-mortem formation of carbonate concretions around tusk-shells over week-month timescales. Sci. Rep. 2015, 5, 14123. [Google Scholar] [CrossRef]
- Yoshida, H.; Yamamoto, K.; Minami, M.; Katsuta, N.; Sin-ichi, S.; Metcalfe, R. Generalized conditions of spherical carbonate formation around decaying organic matter in early diagenesis. Sci. Rep. 2018, 8, 6308. [Google Scholar] [CrossRef]
- Brocks, J.J.; Hope, J.M. Tailing of chromatographic peaks in GC-MS caused by interaction of halogenated solvents with the ion source. J. Chromatogr. Sci. 2014, 52, 471–475. [Google Scholar] [CrossRef]
- Coplen, T.B.; Brand, W.A.; Gehre, M.; Gröning, M.; Meijer, H.A.J.; Toman, B.; Verkouteren, R.M. New Guidelines for δ13C Measurements. Anal. Chem. 2006, 78, 2439–2441. [Google Scholar] [CrossRef] [Green Version]
- Wiemann, J.; Crawford, J.M.; Briggs, D.E.G. Phylogenetic and physiological signals in metazoan fossil biomolecules. Sci. Adv. 2020, 6, eaba6883. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.B.; Shurvell, H.F.; Lightner, D.A.; Cooks, R.G.; Stout, G.H. Introduction to Organic Spectroscopy; Macmillan: New York, NY, USA, 1987. [Google Scholar]
- Chin, K.; Tokaryk, T.T.; Erickson, G.M.; Calk, L.C. A king-sized theropod coprolite. Nature 1998, 393, 680–682. [Google Scholar] [CrossRef]
- Briggs, D.E.; Wilby, P.R. The role of the calcium carbonate-calcium phosphate switch in the mineralization of soft-bodied fossils. J. Geol. Soc. 1996, 153, 665–668. [Google Scholar] [CrossRef]
- Briggs, D.E.; Kear, A.J.; Martill, D.M.; Wilby, P.R. Phosphatization of soft-tissue in experiments and fossils. J. Geol. Soc. 1993, 150, 1035–1038. [Google Scholar] [CrossRef]
- Trueman, C.N. Rare Earth Element Geochemistry and Taphonomy of Terrestrial Vertebrate Assemblages. Palaios 1999, 14, 555–568. [Google Scholar] [CrossRef]
- Fleet, M.E.; Pan, Y. Site preference of rare earth elements in fluorapatite. Am. Mineral. 1995, 80, 329–335. [Google Scholar] [CrossRef]
- Suarez, C.A.; Macpherson, G.L.; González, L.A.; Grandstaff, D.E. Heterogeneous rare earth element (REE) patterns and concentrations in a fossil bone: Implications for the use of REE in vertebrate taphonomy and fossilization history. Geochim. Cosmochim. Acta 2010, 74, 2970–2988. [Google Scholar] [CrossRef]
- Trueman, C.N.; Benton, M.J.; Palmer, M.R. Geochemical taphonomy of shallow marine vertebrate assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 197, 151–169. [Google Scholar] [CrossRef]
- Trueman, C.N.; Behrensmeyer, A.K.; Potts, R.; Tuross, N. High-resolution records of location and stratigraphic provenance from the rare earth element composition of fossil bones. Geochim. Cosmochim. Acta 2006, 70, 4343–4355. [Google Scholar] [CrossRef]
- Trueman, C.N.; Palmer, M.R.; Field, J.; Privat, K.; Ludgate, N.; Chavagnac, V.; Eberth, D.A.; Cifelli, R.; Rogers, R.R. Comparing rates of recrystallisation and the potential for preservation of biomolecules from the distribution of trace elements in fossil bones. Comptes Rendus Palevol 2008, 7, 145–158. [Google Scholar] [CrossRef]
- Trueman, C.N.; Tuross, N. Trace Elements in Recent and Fossil Bone Apatite. Rev. Mineral. Geochem. 2002, 48, 489–521. [Google Scholar] [CrossRef]
- Niedźwiedzki, G.; Bajdek, P.; Owocki, K.; Kear, B.P. An Early Triassic polar predator ecosystem revealed by vertebrate coprolites from the Bulgo Sandstone (Sydney Basin) of southeastern Australia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 464, 5–15. [Google Scholar] [CrossRef]
- Owocki, K.; Niedźwiedzki, G.; Sennikov, A.G.; Golubev, V.K.; Janiszewska, K.; Sulej, T. Upper Permian vertebrate coprolites from Vyazniki and Gorokhovets, Vyatkian Regional Stage, Russian Platform. Palaios 2012, 27, 867–877. [Google Scholar] [CrossRef]
- Brocks, J.J.; Grice, K. Biomarkers (Molecular Fossils). In Encyclopedia of Geobiology; Reitner, J., Thiel, V., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 147–167. [Google Scholar]
- Volkman, J.K. Sterols in microorganisms. Appl. Microbiol. Biotechnol. 2003, 60, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.E.; Walters, C.C.; Moldowan, J.M. Volume 2: Biomarkers and Isotopes in Petroleum Exploration and Earth History. In The Biomarker Guide; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Grice, K.; Klein Breteler, W.C.M.; Schouten, S.; Grossi, V.; de Leeuw, J.W.; Damsté, J.S.S. Effects of zooplankton herbivory on biomarker proxy records. Paleoceanography 1998, 13, 686–693. [Google Scholar] [CrossRef]
- Goad, L.J. Sterol biosynthesis and metabolism in marine invertebrates. Pure Appl. Chem. 1981, 53, 837–852. [Google Scholar] [CrossRef]
- Kanazawa, A. Sterols in marine invertabrates. Fish. Sci. 2001, 67, 997–1007. [Google Scholar] [CrossRef]
- Prost, K.; Birk, J.J.; Lehndorff, E.; Gerlach, R.; Amelung, W. Steroid Biomarkers Revisited—Improved Source Identification of Faecal Remains in Archaeological Soil Material. PLoS ONE 2017, 12, e0164882. [Google Scholar] [CrossRef]
- Nabbefeld, B.; Grice, K.; Schimmelmann, A.; Summons, R.E.; Troitzsch, U.; Twitchett, R.J. A comparison of thermal maturity parameters between freely extracted hydrocarbons (Bitumen I) and a second extract (Bitumen II) from within the kerogen matrix of Permian and Triassic sedimentary rocks. Org. Geochem. 2010, 41, 78–87. [Google Scholar] [CrossRef]
- Van Kaam-Peters, H.M.E.; Köster, J.; Van Der Gaast, S.J.; Dekker, M.; De Leeuw, J.W.; Sinninge Damsté, J.S. The effect of clay minerals on diasterane/sterane ratios. Geochim. Cosmochim. Acta 1998, 62, 2923–2929. [Google Scholar] [CrossRef]
- Peakman, T.M.; De Leeuw, J.W.; Rijpstra, C. Identification and origin of Δ8(14)5α- and Δ145α-sterenes and related hydrocarbons in an immature bitumen from the Monterey Formation, California. Geochim. Cosmochim. Acta 1992, 56, 1223–1230. [Google Scholar] [CrossRef]
- Gagosian, R.B.; Farrington, J.W. Sterenes in surface sediments from the southwest African shelf and slope. Geochim. Cosmochim. Acta 1978, 42, 1091–1101. [Google Scholar] [CrossRef]
- Brassell, S.C.; Murchison, D.G.; Mason, R.; Durand, B.; Eglinton, G.; Comet, P.A.; Curtis, C.D.; Bada, J.; de Leeuw, J.W. Molecular Changes in Sediment Lipids as Indicators of Systematic Early Diagenesis [and Discussion]. Philos. Trans. R. Soc. Lond. Ser. A Math. Phys. Sci. 1985, 315, 57–75. [Google Scholar]
- Rubinstein, I.; Sieskind, O.; Albrecht, P. Rearranged sterenes in a shale: Occurrence and simulated formation. J. Chem. Soc. Perkin Trans. 1 1975, 19, 1833–1836. [Google Scholar] [CrossRef]
- Hussler, G.; Albrecht, P. C27–C29 Monoaromatic anthrasteroid hydrocarbons in Cretaceous black shales. Nature 1983, 304, 262–263. [Google Scholar] [CrossRef]
- Hussler, G.; Chappe, B.; Wehrung, P.; Albrecht, P. C27–C29 ring A monoaromatic steroids in Cretaceous black shales. Nature 1981, 294, 556–558. [Google Scholar] [CrossRef]
- Schüpfer, P.; Finck, Y.; Houot, F.; Gülaçar, F.O. Acid catalysed backbone rearrangement of cholesta-2,4,6-triene: On the origin of ring A and ring B aromatic steroids in recent sediments. Org. Geochem. 2007, 38, 671–681. [Google Scholar] [CrossRef]
- Riolo, J.; Hussler, G.; Albrecht, P.; Connan, J. Distribution of aromatic steroids in geological samples: Their evaluation as geochemical parameters. Org. Geochem. 1986, 10, 981–990. [Google Scholar] [CrossRef]
- Mackenzie, A.S. Organic Reactions as Indicators of the Burial and Temperature Histories of Sedimentary Sequences. Clay Miner. 1984, 19, 271–286. [Google Scholar] [CrossRef]
- Ling, Y.C.; Moreau, J.; Berwick, L.; Tulipani, S.; Grice, K.; Bush, R. Distribution of iron- and sulfate-reducing bacteria across a coastal acid sulfate soil (CASS) environment: Implications for passive bioremediation by tidal inundation. Front. Microbiol. 2015, 6, 624. [Google Scholar] [CrossRef]
- Gaskell, S.J.; Eglinton, G. Sterols of a contemporary lacustrine sediment. Geochim. Cosmochim. Acta 1976, 40, 1221–1228. [Google Scholar] [CrossRef]
- Cranwell, P.A. Lipid geochemistry of sediments from Upton Broad, a small productive lake. Org. Geochem. 1984, 7, 25–37. [Google Scholar] [CrossRef]
- Ficken, K.J.; Li, B.; Swain, D.L.; Eglinton, G. An n-alkane proxy for the sedimentary input of submerged/floating freshwater aquatic macrophytes. Org. Geochem. 2000, 31, 745–749. [Google Scholar] [CrossRef]
- Schouten, S.; Klein Breteler, W.C.M.; Blokker, P.; Schogt, N.; Rijpstra, W.I.C.; Grice, K.; Baas, M.; Damsté, J.S.S. Biosynthetic effects on the stable carbon isotopic compositions of algal lipids: Implications for deciphering the carbon isotopic biomarker record. Geochim. Cosmochim. Acta 1998, 62, 1397–1406. [Google Scholar] [CrossRef]
- Summons, R.E.; Jahnke, L.L.; Roksandic, Z. Carbon isotopic fractionation in lipids from methanotrophic bacteria: Relevance for interpretation of the geochemical record of biomarkers. Geochim. Cosmochim. Acta 1994, 58, 2853–2863. [Google Scholar] [CrossRef]
- Bojanowski, M.J.; Clarkson, E.N.K. Origin of Siderite Concretions in Microenvironments of Methanogenesis Developed in a Sulfate Reduction Zone: An Exception or a Rule? J. Sediment. Res. 2012, 82, 585–598. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PE52315 | PE52316 | PE52336 | |||||
|---|---|---|---|---|---|---|---|
| Fossil | Matrix | Fossil | Matrix | Fossil | Matrix | ||
| 1 Steranes (%) | C27 | 86.2 | 69.1 | 92.8 | 50.1 | 99.0 | 78.0 |
| C28 | 7.3 | 16.3 | 3.5 | 25.0 | 0.6 | 9.8 | |
| C29 | 6.5 | 14.6 | 3.7 | 24.9 | 0.4 | 12.2 | |
| 2 Reg steranes/hopanes | 1.9 | 0.9 | 4.8 | 0.5 | 80.1 | 1.1 | |
| 3 Monoaromatic steroids (%) | C27 | 54.8 | 41.8 | 68.3 | 38.9 | 82.2 | 56.1 |
| C28 | 31.7 | 27.7 | 25.1 | 25.9 | 15.2 | 22.7 | |
| C29 | 13.5 | 30.5 | 6.6 | 35.1 | 2.5 | 21.2 | |
| 4 Triaromatic steroids (%) | C27 | 43.6 | 26.7 | 84.6 | 41.4 | 94.8 | 68.4 |
| C28 | 19.4 | 27.2 | 6.1 | 19.9 | 2.7 | 11.5 | |
| C29 | 37.0 | 46.1 | 9.3 | 38.7 | 2.5 | 20.1 | |
| PE 52315 | PE 52316 | PE 52336 | Procedural Blank | ||||
|---|---|---|---|---|---|---|---|
| Fossil | Matrix | Fossil | Matrix | Fossil | Matrix | ||
| 5α-Cholestan-3β-ol | - | - | 0.43 | - | 0.16 | - | - |
| Cholesterol | 0.07 | 0.06 | 0.05 | 0.17 | 0.07 | 0.14 | 0.07 |
| Coprostan-3-ol | 0.03 | - | - | - | - | - | - |
| PE 52315 | PE 52316 | PE 52336 | ||||
|---|---|---|---|---|---|---|
| Fossil | Matrix | Fossil | Matrix | Fossil | Matrix | |
| 1 Dia/reg C27 steranes | 0.64 | 1.03 | 0.30 | 1.50 | 0.32 | 0.76 |
| 2 Dia/reg C28 steranes | 1.61 | 2.35 | 0.74 | 1.87 | 0.44 | 2.09 |
| 3 Dia/reg C29 steranes | 1.25 | 1.52 | 0.89 | 1.47 | * | 1.46 |
| C29 20S/(20S + 20R) steranes | 0.28 | 0.30 | 0.18 | 0.27 | 0.21 | 0.25 |
| 4 Ts/(Ts + Tm) | 0.52 | 0.55 | 0.47 | 0.52 | 0.50 | 0.44 |
| βα/(βα + αβ) C30 hopane | 0.11 | 0.10 | 0.08 | 0.09 | 0.09 | 0.11 |
| 5 Pr/Ph | 1.00 | 0.99 | 0.85 | 1.03 | 1.65 | 1.42 |
| PE 52316 | PE 52336 | |||
|---|---|---|---|---|
| Fossil | Matrix | Fossil | Matrix | |
| δ13Corg | −23.6 (0.07)3 | −23.8 (0.03)2 | −23.9 (0.12)3 | −23.7 (0.03)3 |
| δ13C17 | −28.4 (0.65 *)3 | −30.0 (0.24)3 | −29.4 (0.54 *)3 | −29.0 (0.62 *)3 |
| δ13CPristane | −28.8 (0.82 *)2 | −29.2 (0.30)3 | −28.4 (0.53 *)3 | −29.3 (0.51 *)3 |
| δ13C18 | −30.4 (0.41 *)3 | −31.7 (0.31)3 | −31.5 (0.13)3 | −30.7 (0.40)3 |
| δ13CPhytane | −33.0 (0.11)2 | −29.5 (0.23)3 | −35.1 (0.39)3 | −33.1 (0.31)3 |
| 1δ13C20–25 | −30.5 | −31.0 | −30.8 | −30.7 |
| δ13Ccholestane | −32.9 (0.39)3 | - | −32.6 (0.29)3 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripp, M.; Wiemann, J.; Brocks, J.; Mayer, P.; Schwark, L.; Grice, K. Fossil Biomarkers and Biosignatures Preserved in Coprolites Reveal Carnivorous Diets in the Carboniferous Mazon Creek Ecosystem. Biology 2022, 11, 1289. https://doi.org/10.3390/biology11091289
Tripp M, Wiemann J, Brocks J, Mayer P, Schwark L, Grice K. Fossil Biomarkers and Biosignatures Preserved in Coprolites Reveal Carnivorous Diets in the Carboniferous Mazon Creek Ecosystem. Biology. 2022; 11(9):1289. https://doi.org/10.3390/biology11091289
Chicago/Turabian StyleTripp, Madison, Jasmina Wiemann, Jochen Brocks, Paul Mayer, Lorenz Schwark, and Kliti Grice. 2022. "Fossil Biomarkers and Biosignatures Preserved in Coprolites Reveal Carnivorous Diets in the Carboniferous Mazon Creek Ecosystem" Biology 11, no. 9: 1289. https://doi.org/10.3390/biology11091289
APA StyleTripp, M., Wiemann, J., Brocks, J., Mayer, P., Schwark, L., & Grice, K. (2022). Fossil Biomarkers and Biosignatures Preserved in Coprolites Reveal Carnivorous Diets in the Carboniferous Mazon Creek Ecosystem. Biology, 11(9), 1289. https://doi.org/10.3390/biology11091289