
Climate Change Helps Polar Invasives Establish and Flourish: Evidence from Long-Term Monitoring of the Blowfly Calliphora vicina
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Thermal Requirements of Calliphora vicina Individuals from the Kerguelen Islands
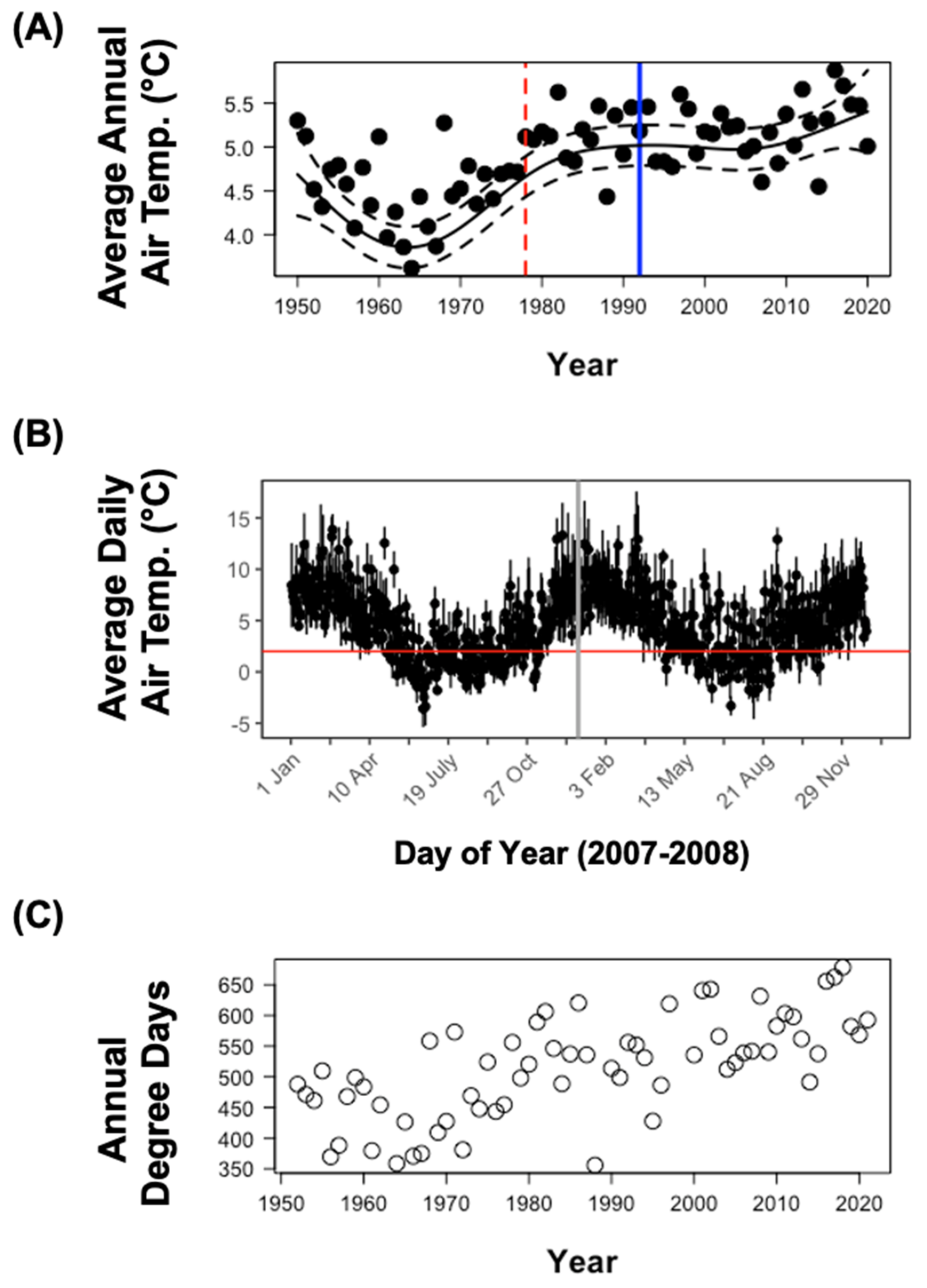
3.2. Thermal Trends on the Kerguelen Islands over Previous Seven Decades
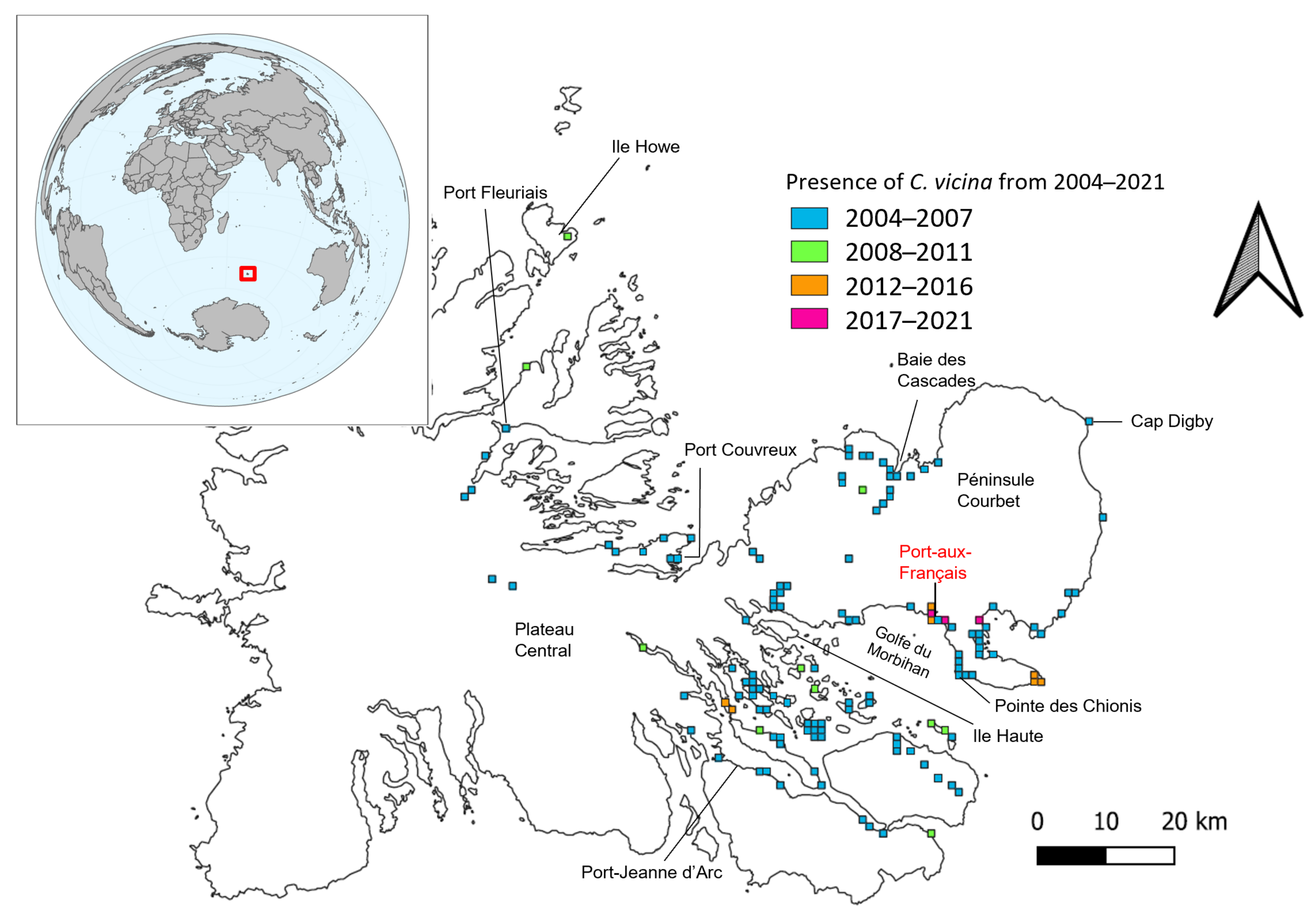
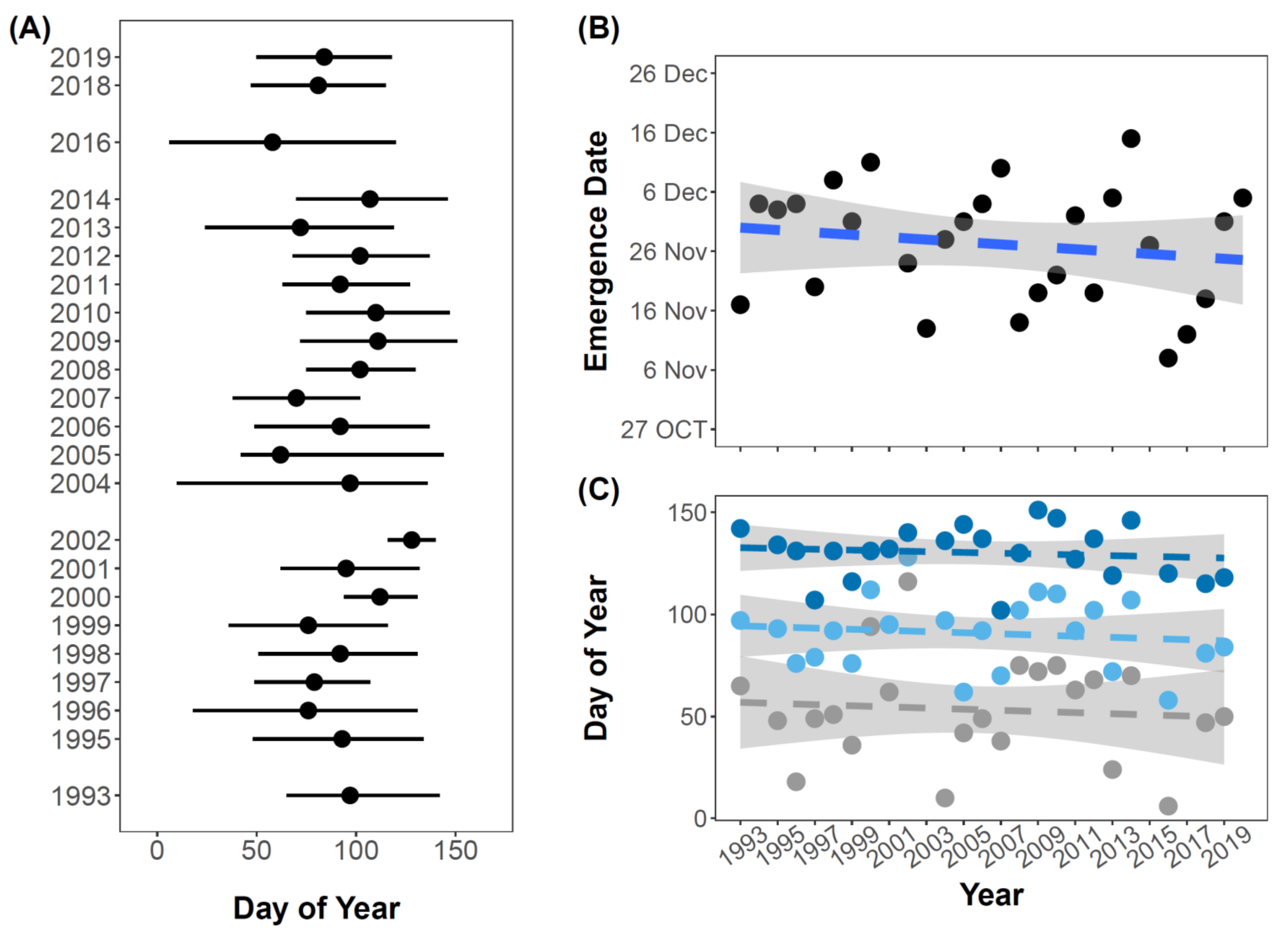
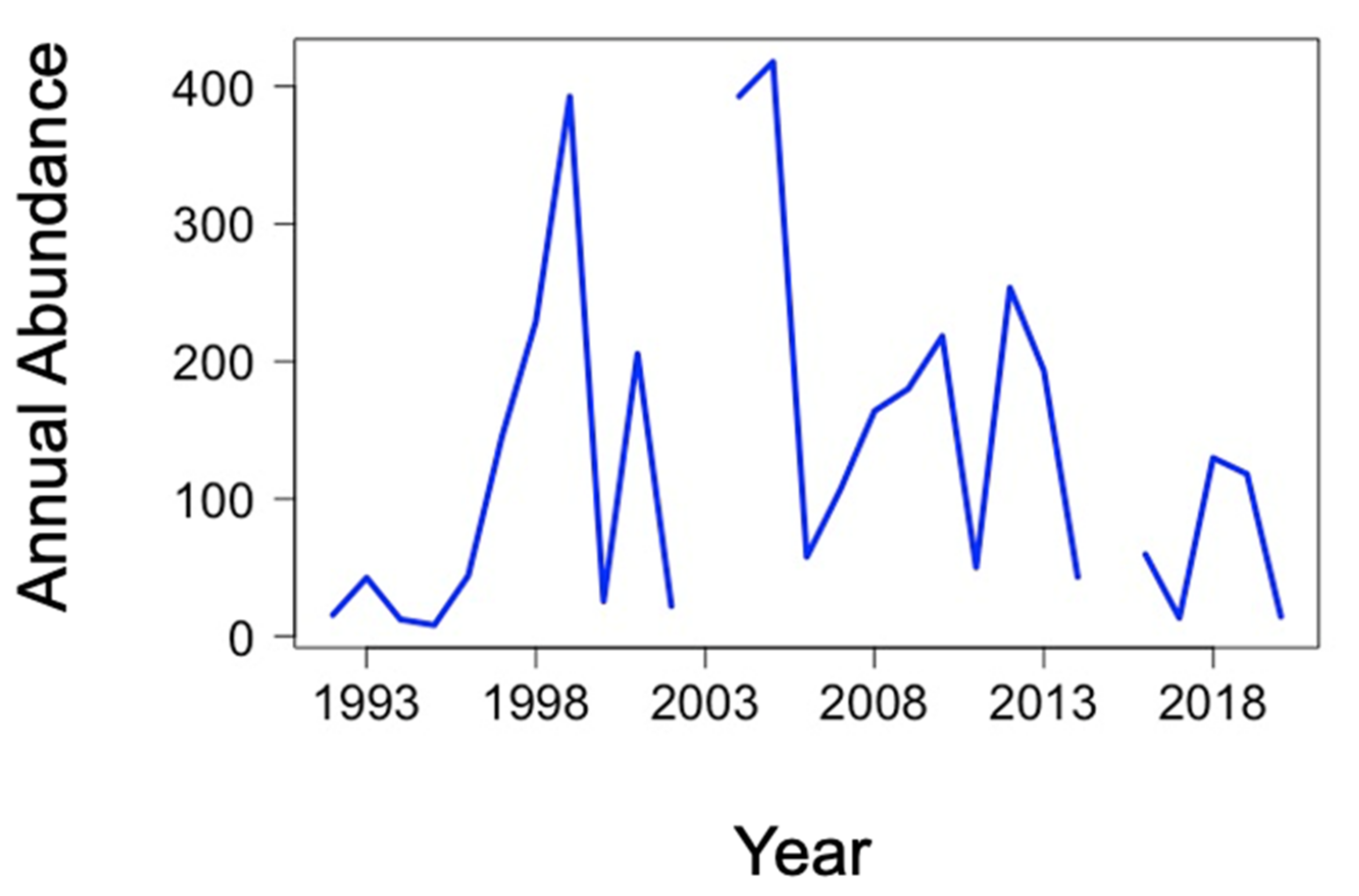
3.3. Long-Term Spatial and Temporal Monitoring of Calliphora vicina in the Kerguelen Islands
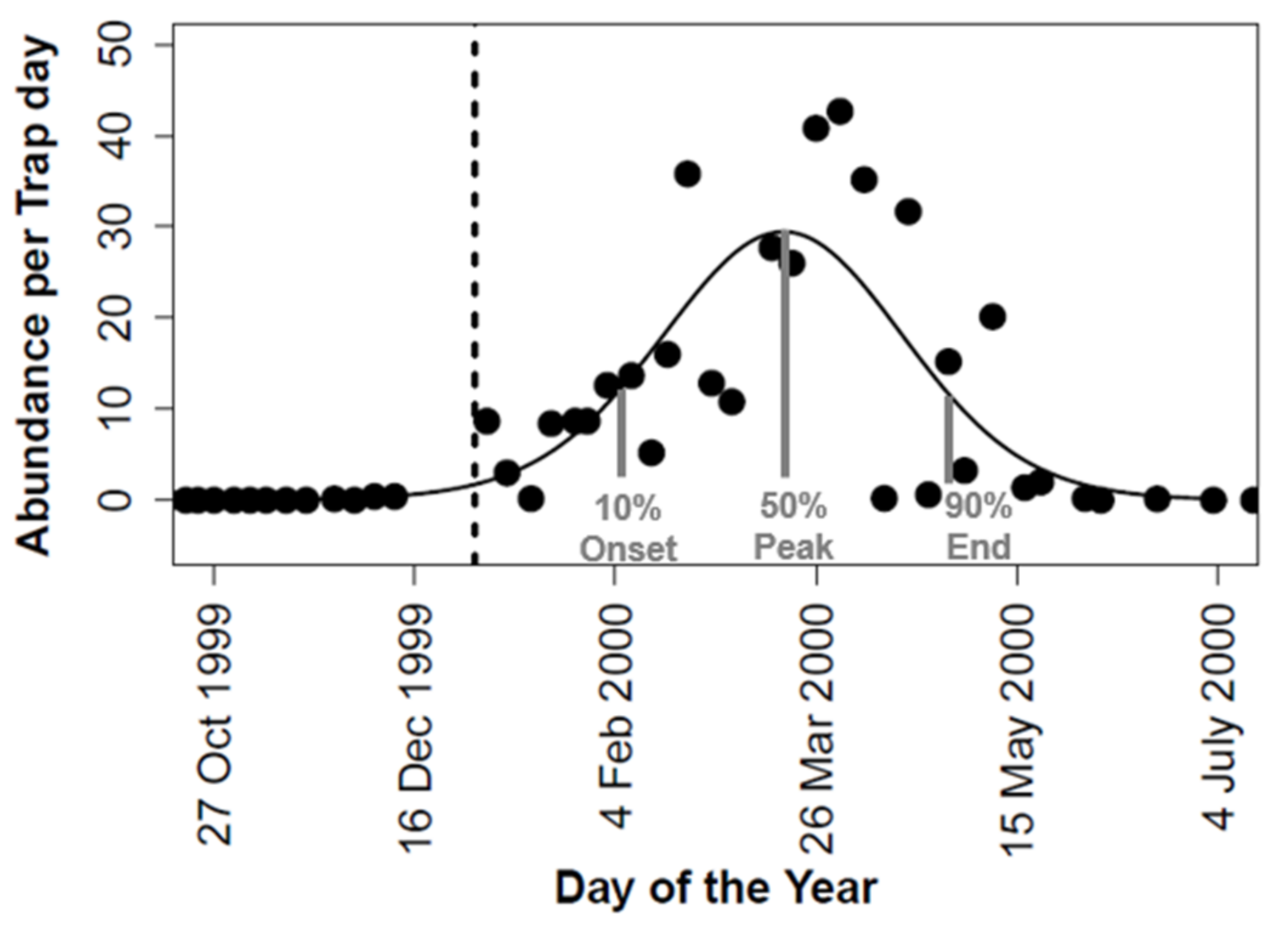
3.4. Phenological Niche of Calliphora vicina
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Emergence Onset | Emergence Peak | Emergence End | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Model | Coef. | SE | R2 | p | Coef. | SE | R2 | p | Coef. | SE | R2 | p |
| Temp. | 0.13 | 5.78 | 2.54 × 10−5 | 0.98 | −11.00 | 2.00 | 0.57 | <0.0001 | −9.54 | 1.81 | 0.55 | <0.0001 |
| Precip. | −1.77 | 6.90 | 0.003 | 0.80 | 3.77 | 4.52 | 0.03 | 0.41 | −2.65 | 2.20 | 0.02 | 0.24 |
| Temp. + Precip. | −1.45 | 8.11 | 0.005 | 0.86 | 1.60 | 2.43 | 0.54 | 0.52 | −2.74 | 2.00 | 0.57 | 0.19 |

References
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging Threats and Persistent Conservation Challenges for Freshwater Biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ Warning on Invasive Alien Species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef]
- Dukes, J.S.; Mooney, H.A. Does Global Change Increase the Success of Biological Invaders? Trends Ecol. Evol. 1999, 14, 135–139. [Google Scholar] [CrossRef]
- Hulme, P.E. Climate Change and Biological Invasions: Evidence, Expectations, and Response Options. Biol. Rev. Camb. Philos. Soc. 2017, 92, 1297–1313. [Google Scholar] [CrossRef]
- Vilà, M.; Hulme, P.E. Non-Native Species, Ecosystem Services, and Human Well-Being. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P.E., Eds.; Invading Nature—Springer Series in Invasion Ecology; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–14. ISBN 978-3-319-45121-3. [Google Scholar]
- Flood, P.J.; Duran, A.; Barton, M.; Mercado-Molina, A.E.; Trexler, J.C. Invasion Impacts on Functions and Services of Aquatic Ecosystems. Hydrobiologia 2020, 847, 1571–1586. [Google Scholar] [CrossRef]
- Frost, C.M.; Allen, W.J.; Courchamp, F.; Jeschke, J.M.; Saul, W.-C.; Wardle, D.A. Using Network Theory to Understand and Predict Biological Invasions. Trends Ecol. Evol. 2019, 34, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Daly, E.Z.; Chabrerie, O.; Massol, F.; Facon, B.; Hess, M.C.M.; Tasiemski, A.; Grandjean, F.; Chauvat, M.; Viard, F.; Forey, E.; et al. A Synthesis of Biological Invasion Hypotheses Associated with the Introduction-Naturalisation-Invasion Continuum. Oikos 2023. [Google Scholar]
- Mahoney, P.J.; Beard, K.H.; Durso, A.M.; Tallian, A.G.; Long, A.L.; Kindermann, R.J.; Nolan, N.E.; Kinka, D.; Mohn, H.E. Introduction Effort, Climate Matching and Species Traits as Predictors of Global Establishment Success in Non-Native Reptiles. Divers. Distrib. 2015, 21, 64–74. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.-L.; Du, D.; van Kleunen, M. Global Change and Biological Invasions. J. Plant Ecol. 2022, 15, 425–428. [Google Scholar] [CrossRef]
- Porter, E.M.; Bowman, W.D.; Clark, C.M.; Compton, J.E.; Pardo, L.H.; Soong, J.L. Interactive Effects of Anthropogenic Nitrogen Enrichment and Climate Change on Terrestrial and Aquatic Biodiversity. Biogeochemistry 2013, 114, 93–120. [Google Scholar] [CrossRef] [Green Version]
- Essl, F.; Lenzner, B.; Bacher, S.; Bailey, S.; Capinha, C.; Daehler, C.; Dullinger, S.; Genovesi, P.; Hui, C.; Hulme, P.E.; et al. Drivers of Future Alien Species Impacts: An Expert-Based Assessment. Glob. Chang. Biol. 2020, 26, 4880–4893. [Google Scholar] [CrossRef] [PubMed]
- Gvoždík, L. Just What Is the Thermal Niche? Oikos 2018, 127, 1701–1710. [Google Scholar] [CrossRef] [Green Version]
- Renault, D.; Leclerc, C.; Colleu, M.-A.; Boutet, A.; Hotte, H.; Colinet, H.; Chown, S.L.; Convey, P. The Rising Threat of Climate Change for Arthropods from Earth’s Cold Regions: Taxonomic Rather than Native Status Drives Species Sensitivity. Glob. Chang. Biol. 2022, 28, 5914–5927. [Google Scholar] [CrossRef] [PubMed]
- Stevens, G.C. The Latitudinal Gradient in Geographical Range: How so Many Species Coexist in the Tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Lebouvier, M.; Laparie, M.; Hullé, M.; Marais, A.; Cozic, Y.; Lalouette, L.; Vernon, P.; Candresse, T.; Frenot, Y.; Renault, D. The Significance of the Sub-Antarctic Kerguelen Islands for the Assessment of the Vulnerability of Native Communities to Climate Change, Alien Insect Invasions and Plant Viruses. Biol. Invasions 2011, 13, 1195–1208. [Google Scholar] [CrossRef]
- Janion-Scheepers, C.; Phillips, L.; Sgrò, C.M.; Duffy, G.A.; Hallas, R.; Chown, S.L. Basal Resistance Enhances Warming Tolerance of Alien over Indigenous Species across Latitude. Proc. Natl. Acad. Sci. USA 2018, 115, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European Phenological Response to Climate Change Matches the Warming Pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A Globally Coherent Fingerprint of Climate Change Impacts across Natural Systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cleland, E.E. The Phenology of Plant Invasions: A Community Ecology Perspective. Front. Ecol. Environ. 2011, 9, 287–294. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; Cleland, E.E. Phenological Niches and the Future of Invaded Ecosystems with Climate Change. AoB PLANTS 2014, 6, plu013. [Google Scholar] [CrossRef] [Green Version]
- Arias, P.; Bellouin, N.; Coppola, E.; Jones, R.; Krinner, G.; Marotzke, J.; Naik, V.; Palmer, M.; Plattner, G.-K.; Rogelj, J.; et al. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Technical Summary; Masson-Delmotte, V., Zhai, P., Pirani, A., Conners, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Pauchard, A.; Milbau, A.; Albihn, A.; Alexander, J.; Burgess, T.; Daehler, C.; Englund, G.; Essl, F.; Evengård, B.; Greenwood, G.B.; et al. Non-Native and Native Organisms Moving into High Elevation and High Latitude Ecosystems in an Era of Climate Change: New Challenges for Ecology and Conservation. Biol. Invasions 2016, 18, 345–353. [Google Scholar] [CrossRef]
- Bauer, A.; Bauer, A.M.; Tomberlin, J.K. Impact of Diet Moisture on the Development of the Forensically Important Blow Fly Cochliomyia macellaria (Fabricius) (Diptera: Calliphoridae). Forensic Sci. Int. 2020, 312, 110333. [Google Scholar] [CrossRef]
- Marchenko, M.I. Medicolegal Relevance of Cadaver Entomofauna for the Determination of the Time of Death. Forensic Sci. Int. 2001, 120, 89–109. [Google Scholar] [CrossRef] [PubMed]
- Defilippo, F.; Bonilauri, P.; Dottori, M. Effect of Temperature on Six Different Developmental Landmarks within the Pupal Stage of the Forensically Important Blowfly Calliphora vicina (Robineau-Desvoidy) (Diptera: Calliphoridae). J. Forensic Sci. 2013, 58, 1554–1557. [Google Scholar] [CrossRef]
- Chevrier, M.; Vernon, P.; Frenot, Y. Potential Effects of Two Alien Insects on a Sub-Antarctic Wingless Fly in the Kerguelen Islands. In Antarctic Communities: Species, Structure and Survival; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Wells, J.D.; Greenberg, B. Interaction between Chrysomya rufifacies and Cochliomyia macellaria (Diptera: Calliphoridae): The Possible Consequences of an Invasion. Bull. Entomol. Res. 1992, 82, 133–137. [Google Scholar] [CrossRef]
- Carmo, R.F.R.; Vasconcelos, S.D.; Brundage, A.L.; Tomberlin, J.K. How Do Invasive Species Affect Native Species? Experimental Evidence from a Carrion Blowfly (Diptera: Calliphoridae) System. Ecol. Entomol. 2018, 43, 483–493. [Google Scholar] [CrossRef]
- Frenot, Y.; Chown, S.L.; Whinam, J.; Selkirk, P.M.; Convey, P.; Skotnicki, M.; Bergstrom, D.M. Biological Invasions in the Antarctic: Extent, Impacts and Implications. Biol. Rev. Camb. Philos. Soc. 2005, 80, 45–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellard, C.; Rysman, J.-F.; Leroy, B.; Claud, C.; Mace, G.M. A Global Picture of Biological Invasion Threat on Islands. Nat. Ecol. Evol. 2017, 1, 1862–1869. [Google Scholar] [CrossRef]
- Hullé, M.; Vernon, P. Terrestrial Macro-Arthropods of the Sub-Antarctic Islands of Possession (Crozet Archipelago) and Kerguelen: Inventory of Native and Non-Native Species. Zoosystema 2021, 43, 549–561. [Google Scholar] [CrossRef]
- Krikken, J.; Huijbregts, J. Insects as Forensic Informants: The Dutch Experience and Procedure. Proc. Exp. Appl. Entomol. 2001, 12, 159–164. [Google Scholar]
- Vinogradova, E.B.; Zinovjeva, K.B. Maternal Induction of Larval Diapause in the Blowfly, Calliphora vicina. J. Insect Physiol. 1972, 18, 2401–2409. [Google Scholar] [CrossRef]
- Graham-Smith, G.S. Observations on the Habits and Parasites of Common Flies. Parasitology 1916, 8, 440–544. [Google Scholar] [CrossRef]
- Vinogradova, E.B. Geographical Variation and Ecological Control of Diapause in Flies. In The Evolution of Insect Life Cycles; Taylor, F., Karban, R., Eds.; Springer: New York, NY, USA, 1986; pp. 35–47. [Google Scholar]
- Aak, A.; Birkemoe, T.; Leinaas, H.P. Phenology and Life History of the Blowfly Calliphora vicina in Stockfish Production Areas. Entomol. Exp. Appl. 2011, 139, 35–46. [Google Scholar] [CrossRef]
- Allen, J.C. A Modified Sine Wave Method for Calculating Degree Days 1. Environ. Entomol. 1976, 5, 388–396. [Google Scholar] [CrossRef]
- Donovan, S.E.; Hall, M.J.R.; Turner, B.D.; Moncrieff, C.B. Larval Growth Rates of the Blowfly, Calliphora vicina, over a Range of Temperatures. Med. Vet. Entomol. 2006, 20, 106–114. [Google Scholar] [CrossRef]
- Putman, R.J. Dynamics of the Blowfly, Calliphora erythrocephala, Within Carrion. J. Anim. Ecol. 1977, 46, 853–866. [Google Scholar] [CrossRef]
- Simpson, G.L. Modelling Palaeoecological Time Series Using Generalised Additive Models. Front. Ecol. Evol. 2018, 6, 149. [Google Scholar] [CrossRef] [Green Version]
- Welcome to the QGIS Project! Available online: https://qgis.org/en/site/ (accessed on 10 December 2022).
- Wood, S.N. An Introduction with R, Second Edition, 2nd ed.; Chapman and Hall/CRC: New York, NY, USA, 2017; ISBN 978-1-315-37027-9. [Google Scholar]
- Rouget, M.; Robertson, M.P.; Wilson, J.R.U.; Hui, C.; Essl, F.; Renteria, J.L.; Richardson, D.M. Invasion Debt—Quantifying Future Biological Invasions. Divers. Distrib. 2016, 22, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Coutts, S.R.; Helmstedt, K.J.; Bennett, J.R. Invasion Lags: The Stories We Tell Ourselves and Our Inability to Infer Process from Pattern. Divers. Distrib. 2018, 24, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Davies, L.; Ratcliffe, G.G. Development Rates of Some Pre-Adult Stages in Blowflies with Reference to Low Temperatures. Med. Vet. Entomol. 1994, 8, 245–254. [Google Scholar] [CrossRef]
- Salimi, M.; Rassi, Y.; Oshaghi, M.; Chatrabgoun, O.; Limoee, M.; Rafizadeh, S. Temperature Requirements for the Growth of Immature Stages of Blowflies Species, Chrysomya albiceps and Calliphora vicina, (Diptera:Calliphoridae) under Laboratory Conditions. Egypt. J. Forensic Sci. 2018, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Chown, S.L.; Marais, E.; Terblanche, J.S.; Klok, C.J.; Lighton, J.R.B.; Blackburn, T.M. Scaling of Insect Metabolic Rate Is Inconsistent with the Nutrient Supply Network Model. Funct. Ecol. 2007, 21, 282–290. [Google Scholar] [CrossRef]
- Strayer, D.L.; D’Antonio, C.M.; Essl, F.; Fowler, M.S.; Geist, J.; Hilt, S.; Jarić, I.; Jöhnk, K.; Jones, C.G.; Lambin, X.; et al. Boom-Bust Dynamics in Biological Invasions: Towards an Improved Application of the Concept. Ecol. Lett. 2017, 20, 1337–1350. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-Scale Drivers of Natural Disturbances Prone to Anthropogenic Amplification: The Dynamics of Bark Beetle Eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Battisti, A.; Stastny, M.; Netherer, S.; Robinet, C.; Schopf, A.; Roques, A.; Larsson, S. Expansion of Geographic Range in the Pine Processionary Moth Caused by Increased Winter Temperatures. Ecol. Appl. 2005, 15, 2084–2096. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Appelberg, G.; Harding, S.; Bärring, L. Spatio-Temporal Impact of Climate Change on the Activity and Voltinism of the Spruce Bark Beetle, Ips typographus. Glob. Chang. Biol. 2009, 15, 486–499. [Google Scholar] [CrossRef]
- Mitton, J.B.; Ferrenberg, S.M.; Benkman, N.H.E.C.W. Mountain Pine Beetle Develops an Unprecedented Summer Generation in Response to Climate Warming. Am. Nat. 2012, 179, E163–E171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pöyry, J.; Leinonen, R.; Söderman, G.; Nieminen, M.; Heikkinen, R.K.; Carter, T.R. Climate-Induced Increase of Moth Multivoltinism in Boreal Regions. Glob. Ecol. Biogeogr. 2011, 20, 289–298. [Google Scholar] [CrossRef]
- Macgregor, C.J.; Thomas, C.D.; Roy, D.B.; Beaumont, M.A.; Bell, J.R.; Brereton, T.; Bridle, J.R.; Dytham, C.; Fox, R.; Gotthard, K.; et al. Climate-Induced Phenology Shifts Linked to Range Expansions in Species with Multiple Reproductive Cycles per Year. Nat. Commun. 2019, 10, 4455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosati, J.Y. Spatial and Temporal Variability in the Carrion Insect Community: Using Blow Flies (Family: Calliphoridae) as a Model System to Study Coexistence Mechanisms at Multiple Scales. Doctoral Thesis, University of Windsor, Windsor, ON, Canada, 2014. [Google Scholar]
- Galindo, L.A.; Moral, R.A.; Moretti, T.C.; Godoy, W.A.C.; Demétrio, C.G.B. Intraguild Predation Influences Oviposition Behavior of Blow Flies (Diptera: Calliphoridae). Parasitol. Res. 2016, 115, 2097–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebouvier, M.; Lambret, P.; Garnier, A.; Convey, P.; Frenot, Y.; Vernon, P.; Renault, D. Spotlight on the Invasion of a Carabid Beetle on an Oceanic Island over a 105-Year Period. Sci. Rep. 2020, 10, 17103. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; VerCauteren, K. The Role of Scavenging in Disease Dynamics. In Carrion Ecology and Management; Olea, P.P., Mateo-Tomás, P., Sánchez-Zapata, J.A., Eds.; Wildlife Research Monographs; Springer International Publishing: Cham, Switzerland, 2019; pp. 161–182. ISBN 978-3-030-16501-7. [Google Scholar]
- Basson, L.; Hassim, A.; Dekker, A.; Gilbert, A.; Beyer, W.; Rossouw, J.; van Heerden, H. Blowflies as Vectors of Bacillus anthracis in the Kruger National Park. Koedoe 2018, 60, 6. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging Infectious Diseases of Wildlife—Threats to Biodiversity and Human Health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Eppinga, M.B.; Rietkerk, M.; Dekker, S.C.; De Ruiter, P.C.; Van der Putten, W.H.; Van der Putten, W.H. Accumulation of Local Pathogens: A New Hypothesis to Explain Exotic Plant Invasions. Oikos 2006, 114, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Catts, E.P. Problems in Estimating the Postmortem Interval in Death Investigations. J. Agric. Entomol. 1992, 9, 245–255. [Google Scholar]
- Richards, C.S.; Price, B.W.; Villet, M.H. Thermal Ecophysiology of Seven Carrion-Feeding Blowflies in Southern Africa. Entomol. Exp. Appl. 2009, 131, 11–19. [Google Scholar] [CrossRef]
- Convey, P.; Key, R.S.; Key, R.J.D. The Establishment of a New Ecological Guild of Pollinating Insects on Sub-Antarctic South Georgia. Antarct. Sci. 2010, 22, 508–512. [Google Scholar] [CrossRef] [Green Version]
- Simberloff, D.; Von Holle, B. Positive Interactions of Nonindigenous Species: Invasional Meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]




| Days to Stage | |||||||
|---|---|---|---|---|---|---|---|
| Temp. (°C) | Hatch | 1st Larval Instar | 2nd Larval Instar | 3rd Larval Instar | Pupa | Adult | Total DD |
| 4 | 31 | - | - | - | - | - | - |
| 8 | 4.5 | 6 | 9 | 18.5 | 23 | 92.8 | 556.8 |
| 12 | 2 | 4 | 5 | 7.5 | 12 | 41.5 | 415 |
| 16 | 1 | 3 | 3 | 5 | 7 | 29.9 | 418.6 |
| 20 | 1 | 2 | 2.5 | 4 | 5 | 22.6 | 406.8 |
| AC | 4 | 3 | 4.5 | 10 | 16 | 73 | 397.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daly, E.Z.; Gerlich, H.S.; Frenot, Y.; Høye, T.T.; Holmstrup, M.; Renault, D. Climate Change Helps Polar Invasives Establish and Flourish: Evidence from Long-Term Monitoring of the Blowfly Calliphora vicina. Biology 2023, 12, 111. https://doi.org/10.3390/biology12010111
Daly EZ, Gerlich HS, Frenot Y, Høye TT, Holmstrup M, Renault D. Climate Change Helps Polar Invasives Establish and Flourish: Evidence from Long-Term Monitoring of the Blowfly Calliphora vicina. Biology. 2023; 12(1):111. https://doi.org/10.3390/biology12010111
Chicago/Turabian StyleDaly, Ella Z., Hannah Sørine Gerlich, Yves Frenot, Toke T. Høye, Martin Holmstrup, and David Renault. 2023. "Climate Change Helps Polar Invasives Establish and Flourish: Evidence from Long-Term Monitoring of the Blowfly Calliphora vicina" Biology 12, no. 1: 111. https://doi.org/10.3390/biology12010111
APA StyleDaly, E. Z., Gerlich, H. S., Frenot, Y., Høye, T. T., Holmstrup, M., & Renault, D. (2023). Climate Change Helps Polar Invasives Establish and Flourish: Evidence from Long-Term Monitoring of the Blowfly Calliphora vicina. Biology, 12(1), 111. https://doi.org/10.3390/biology12010111