
Morphological Continua Make Poor Species: Genus-Wide Morphometric Survey of the European Bee Orchids (Ophrys L.)


Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Bee Orchids as an Evolutionary Case-Study
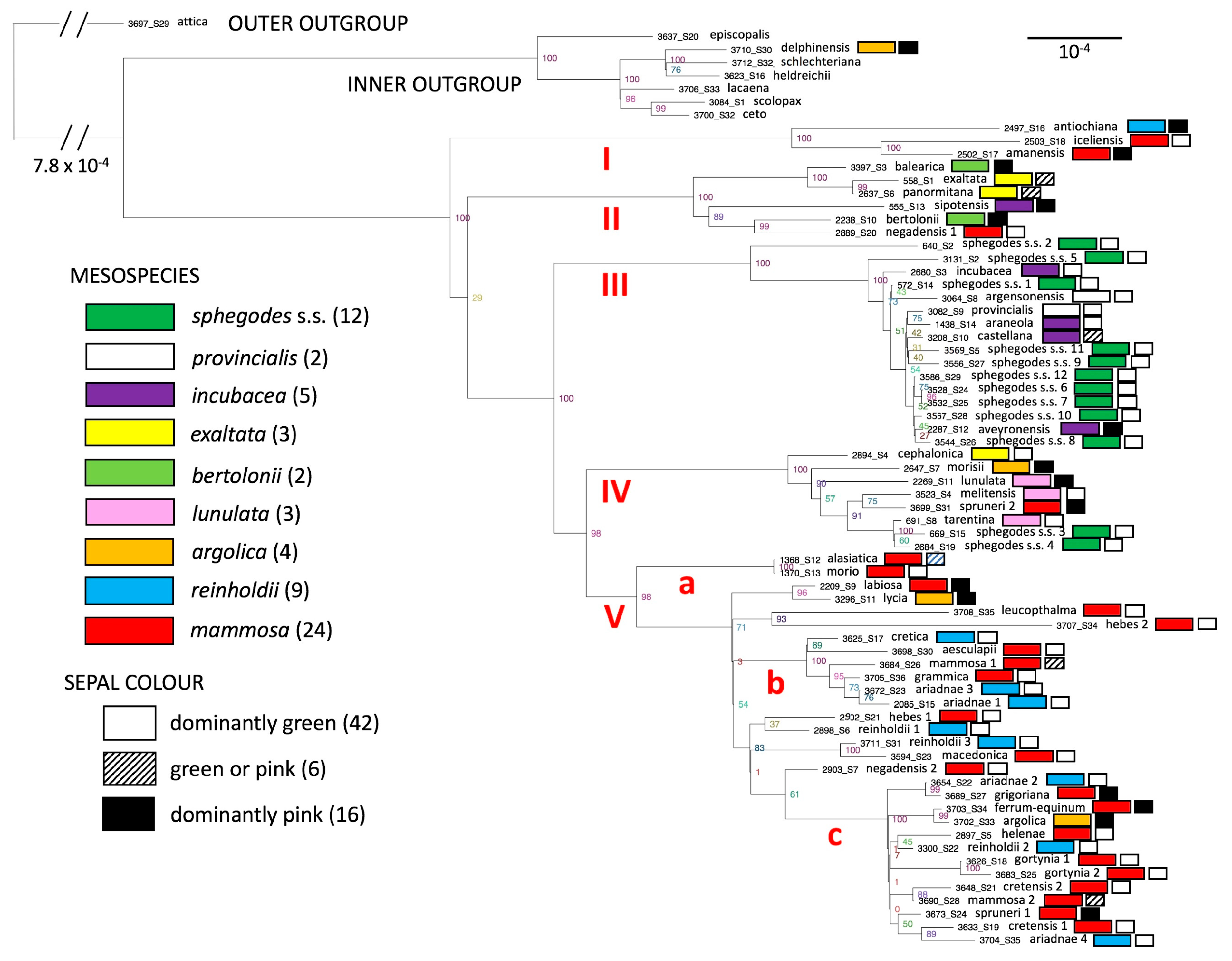
1.2. The Explicit Taxonomic Controversy: Microspecies, Mesospecies and Macrospecies
1.3. The General Taxonomic Controversy: Natural Versus Artificial Species
1.4. Aims of the Present Study
2. Materials and Methods
2.1. Plant Materials
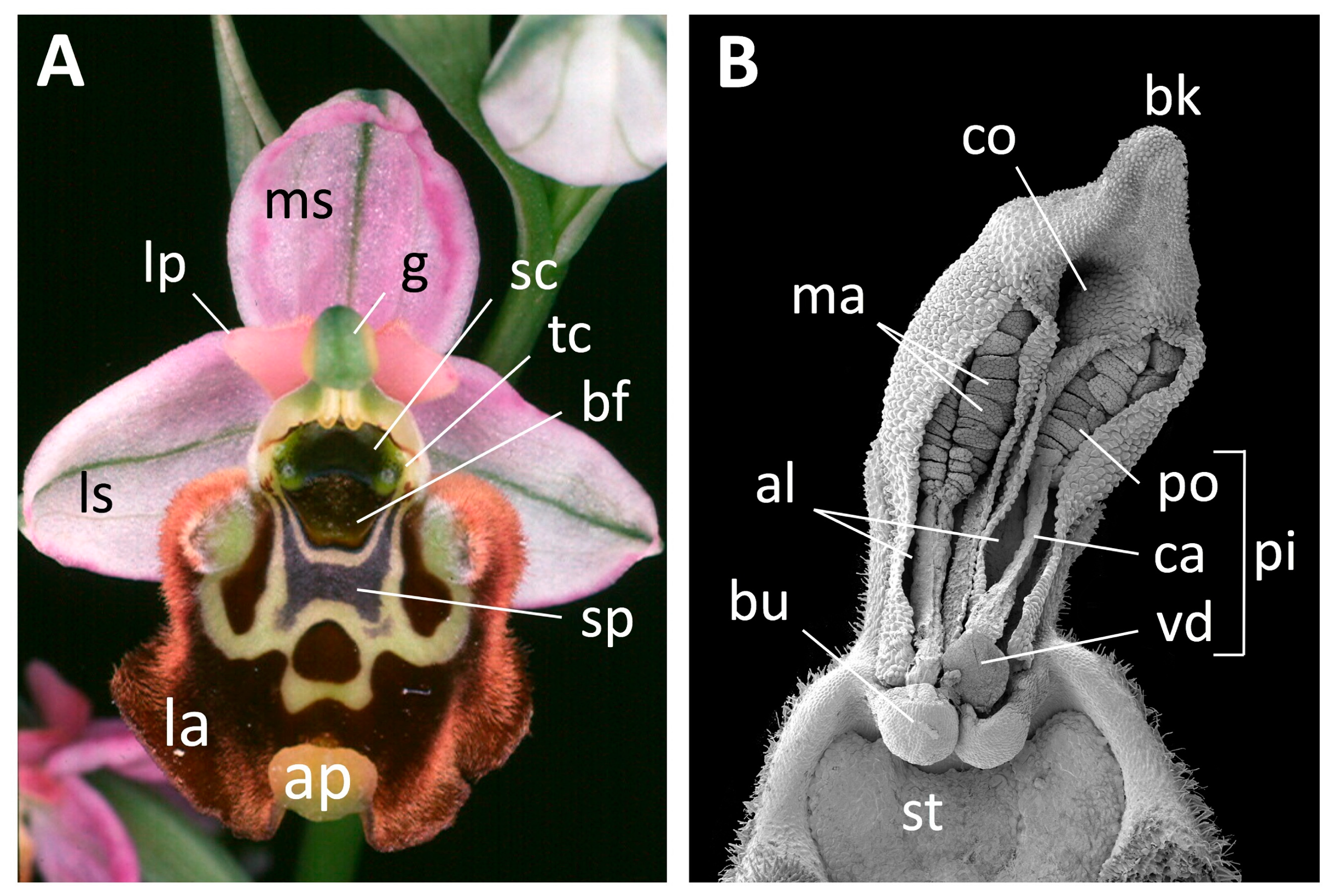
2.2. Morphometric Data Collection
2.3. Morphometric Data Analysis
2.4. Scanning Electron Microscopy and Anatomy
3. Results
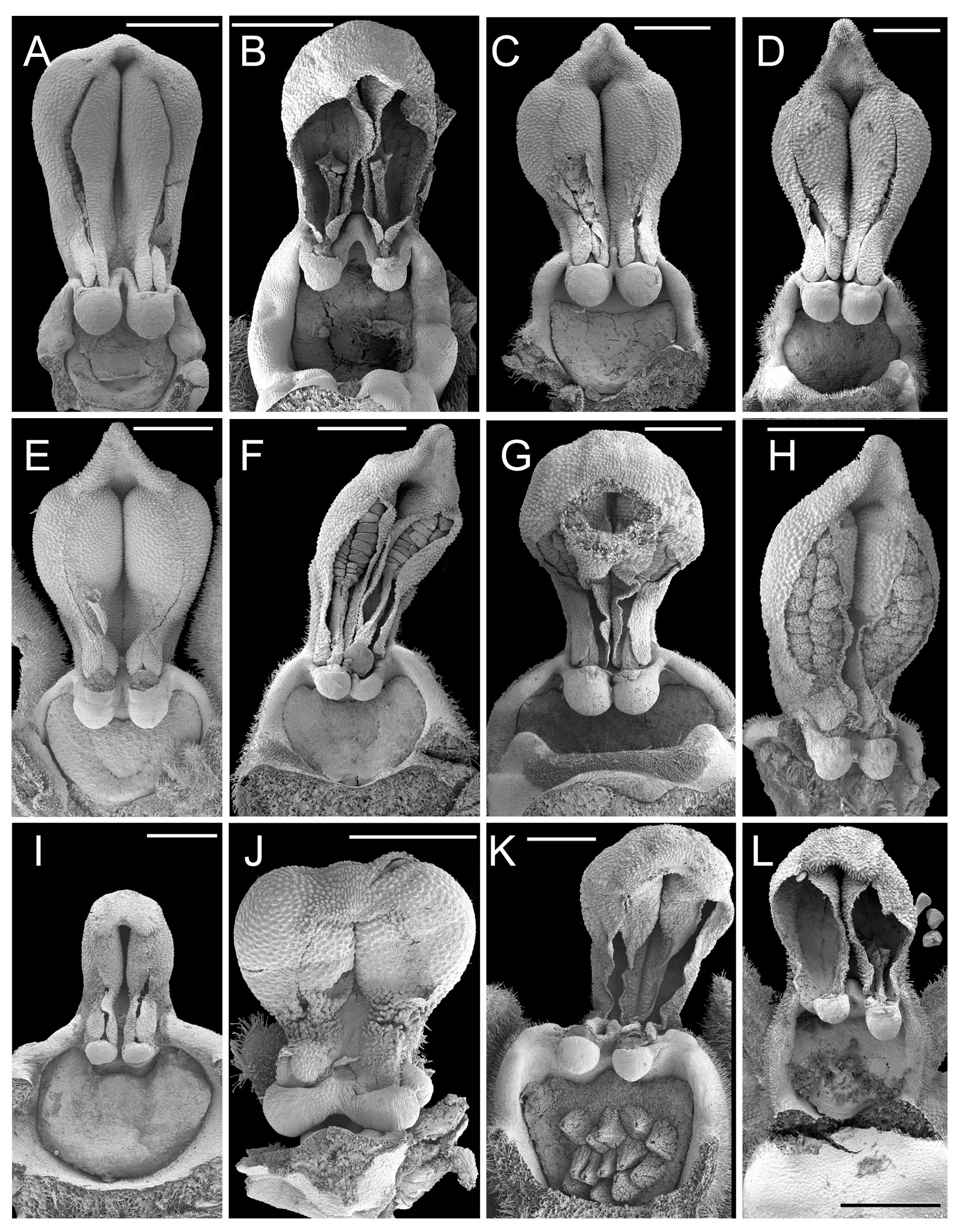
3.1. Scanning Electron Microscopy and Anatomy
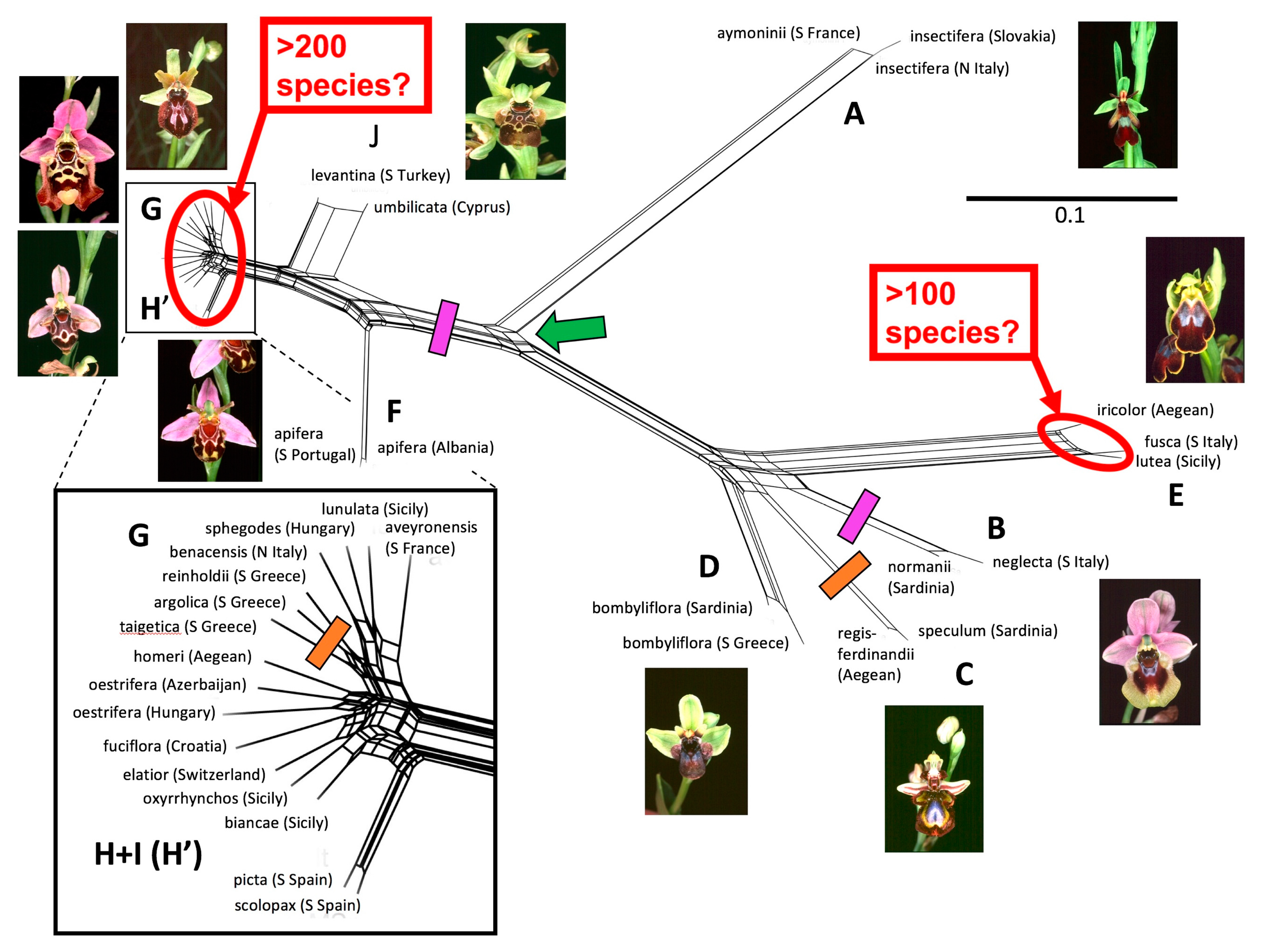
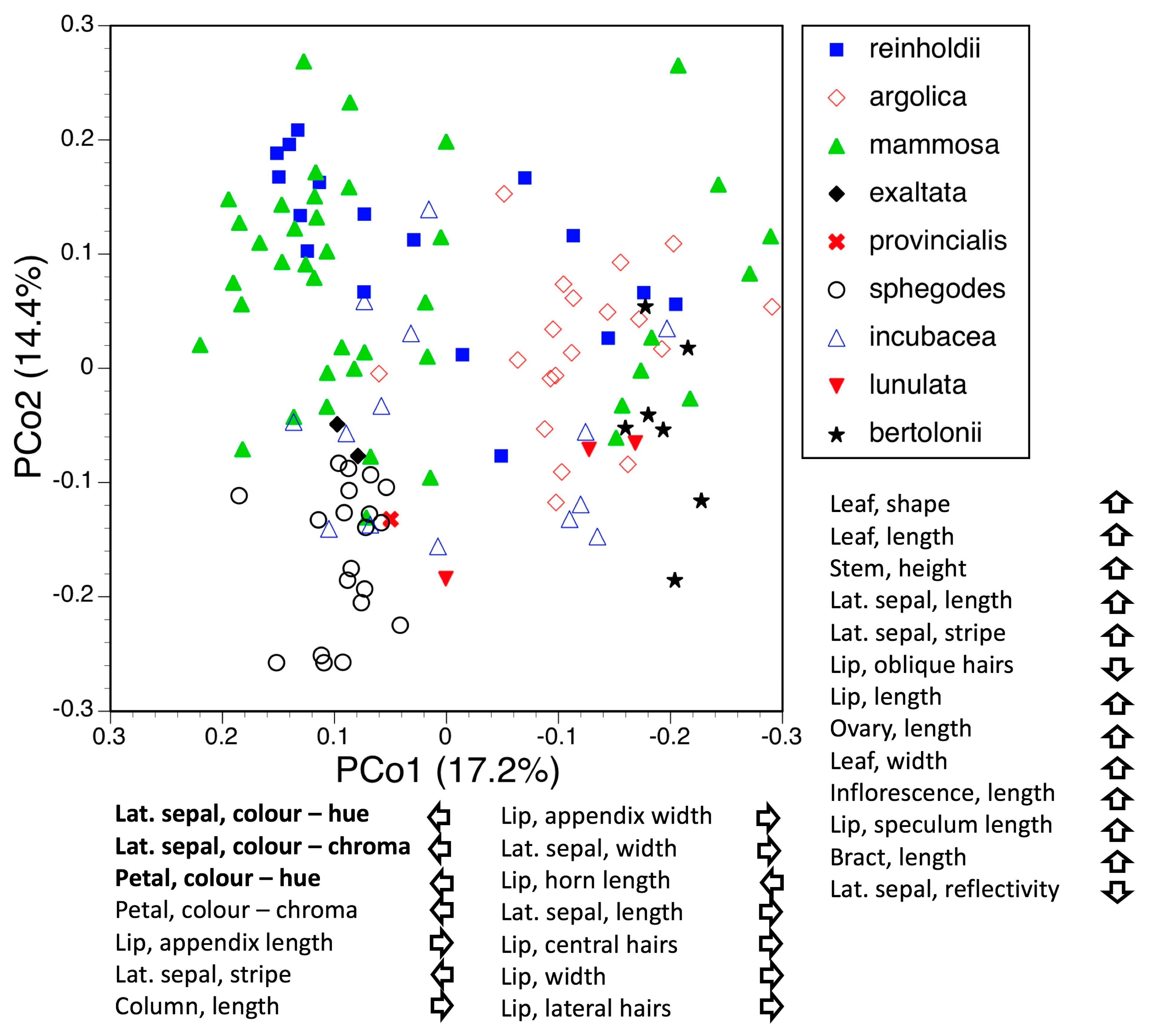
3.2. Morphometrics: Analyses Involving Multiple Macrospecies
3.2.1. All Nine Macrospecies
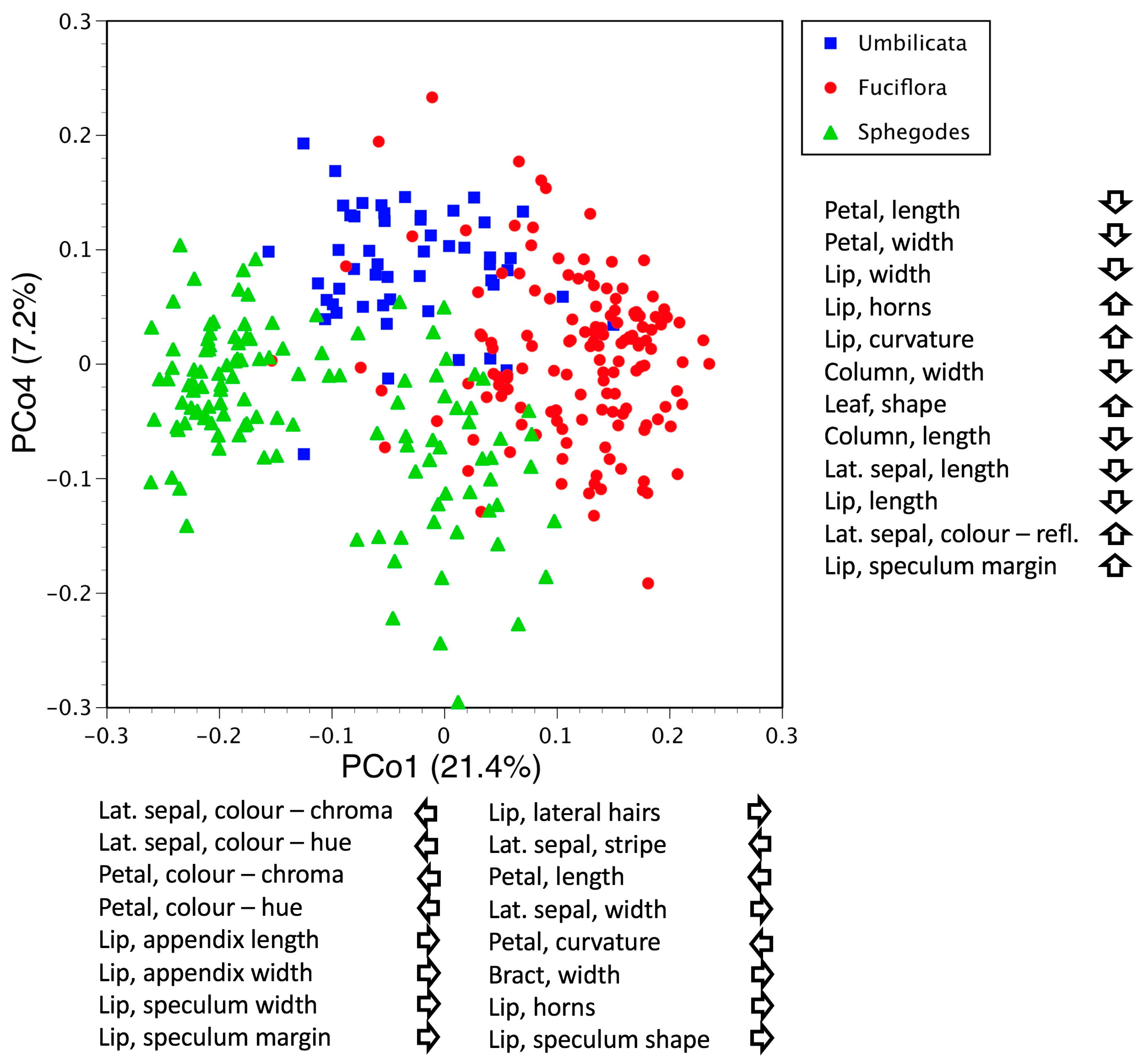
3.2.2. The Umbilicata-Fuciflora-Sphegodes Clade
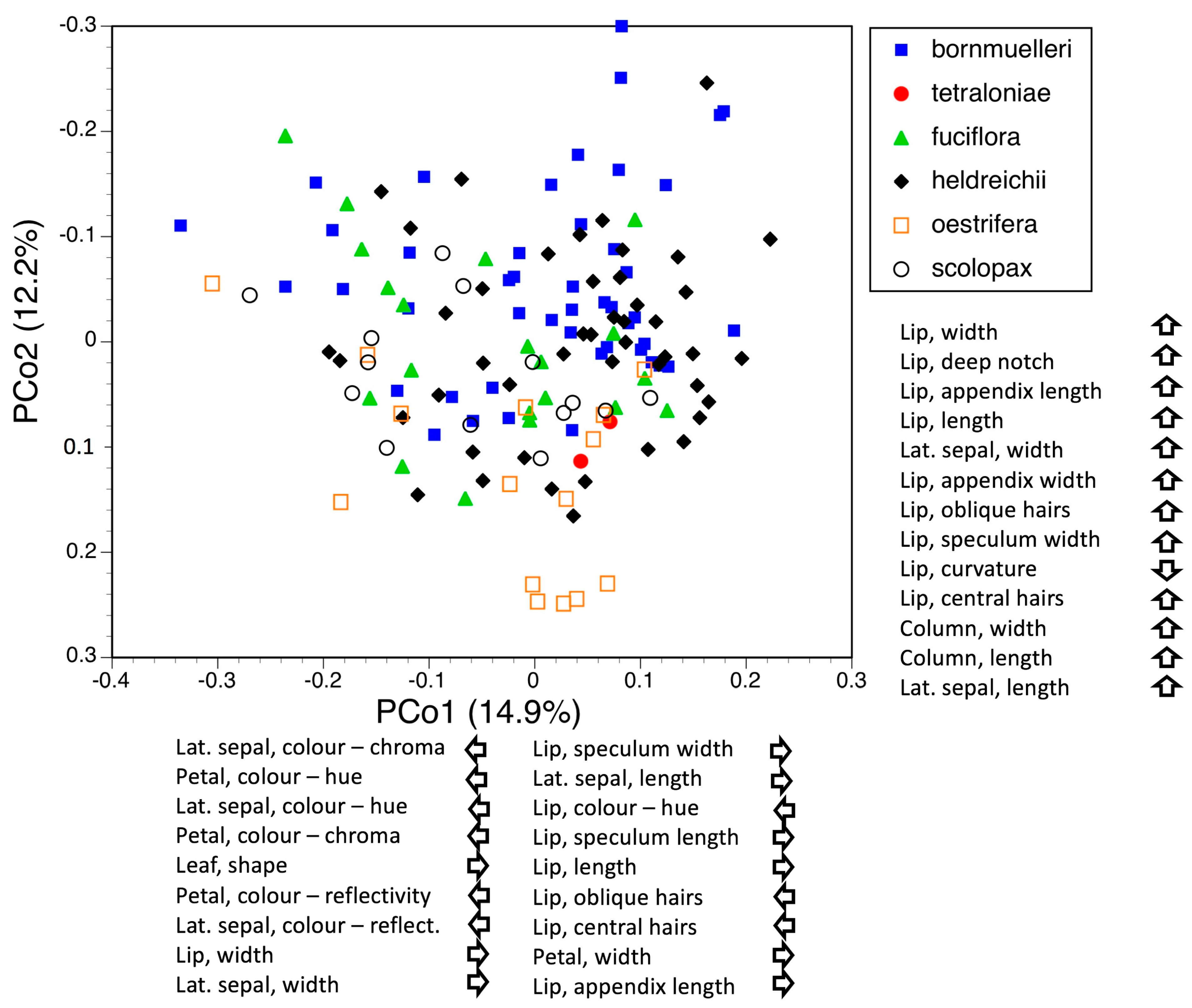
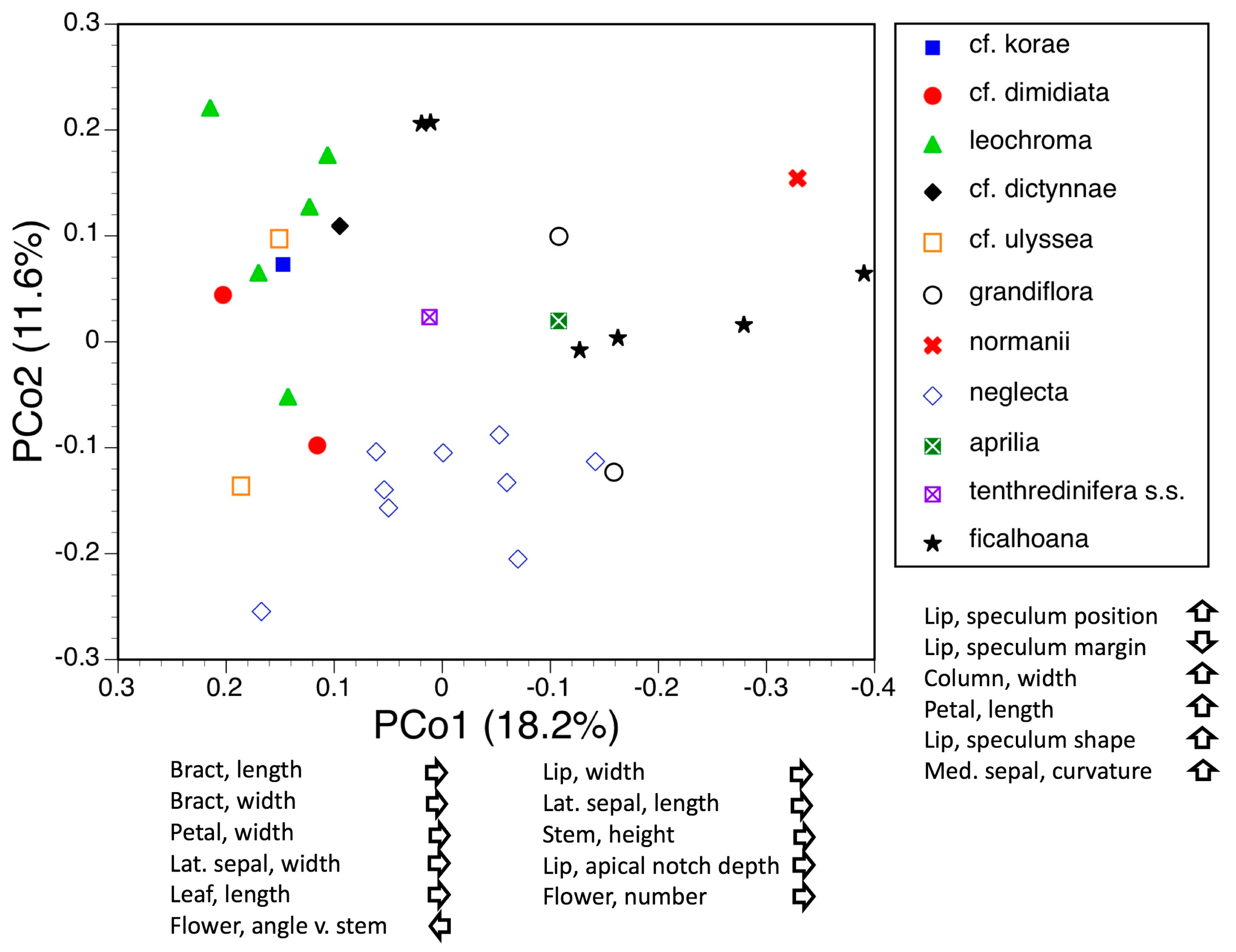
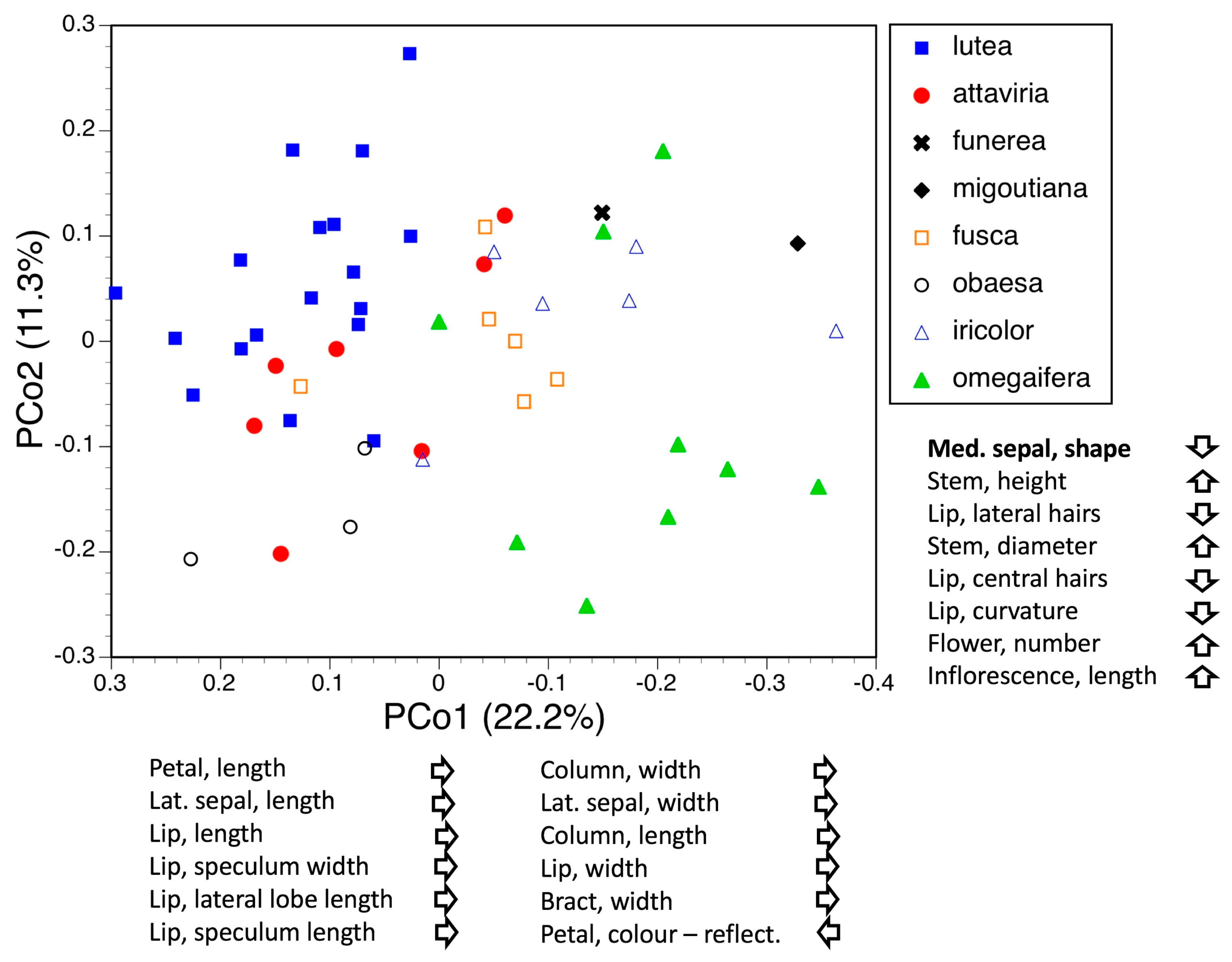
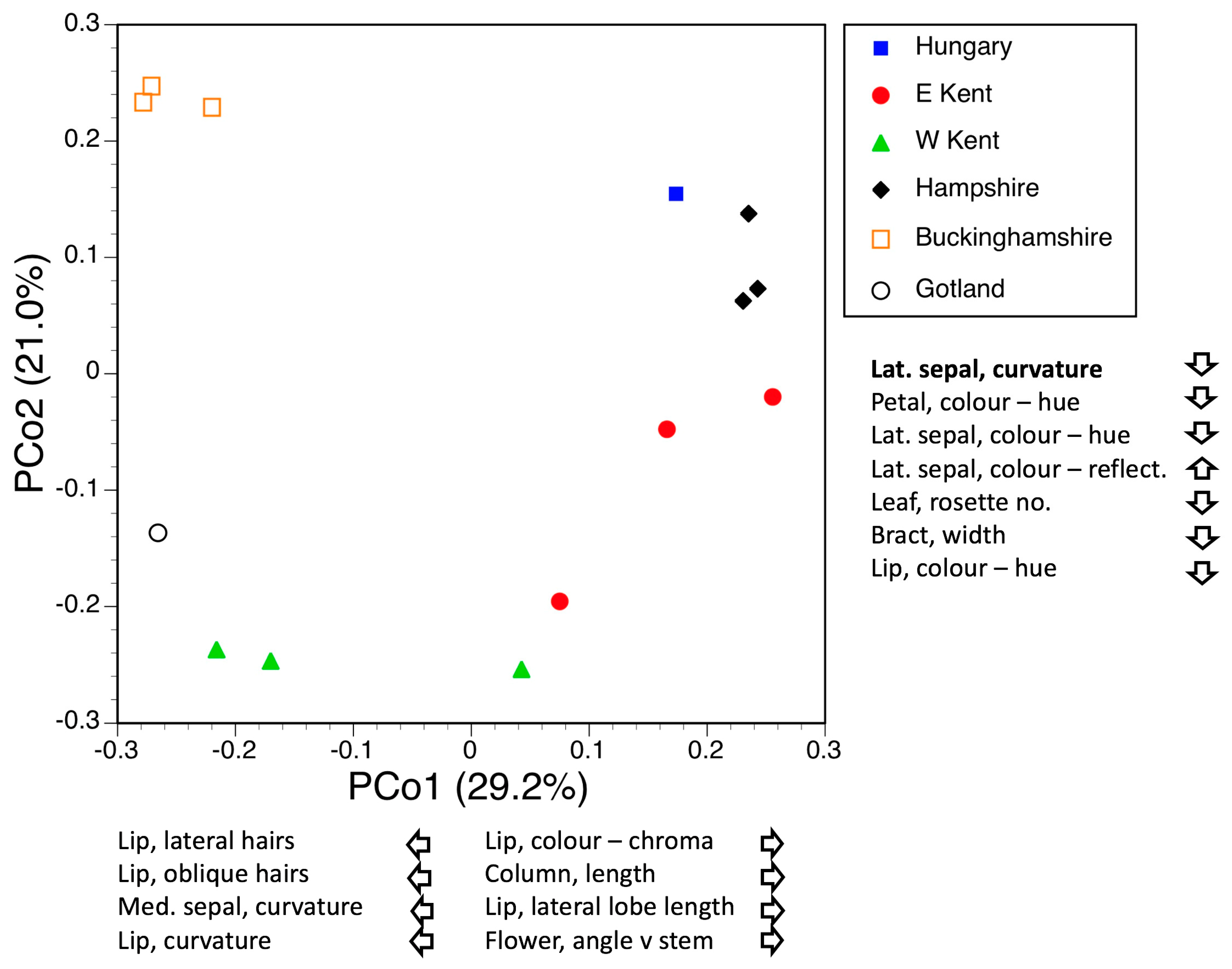
3.3. Morphometrics: Analyses Involving Single Macrospecies but Multiple Mesospecies
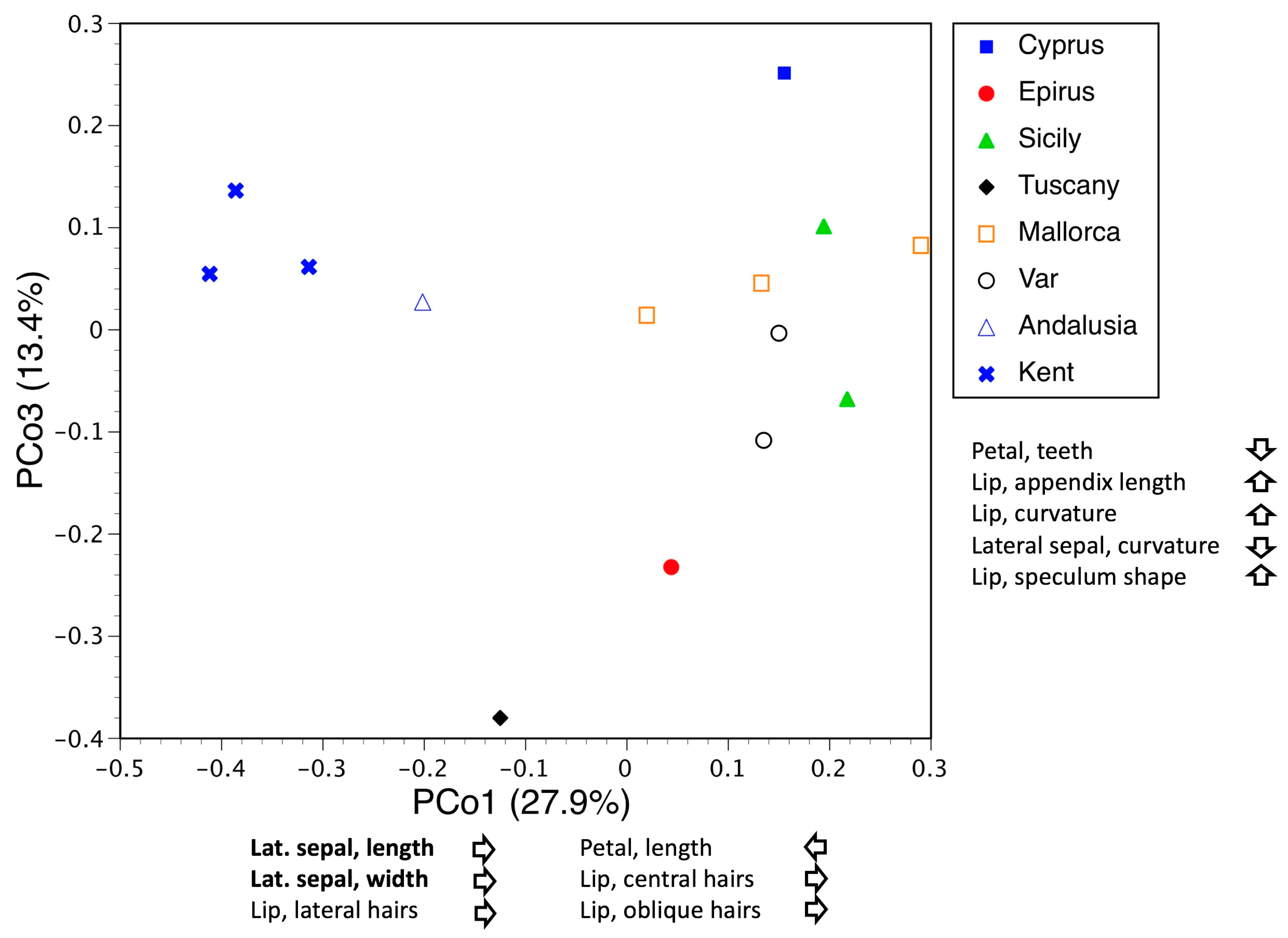
3.3.1. Macrospecies Sphegodes
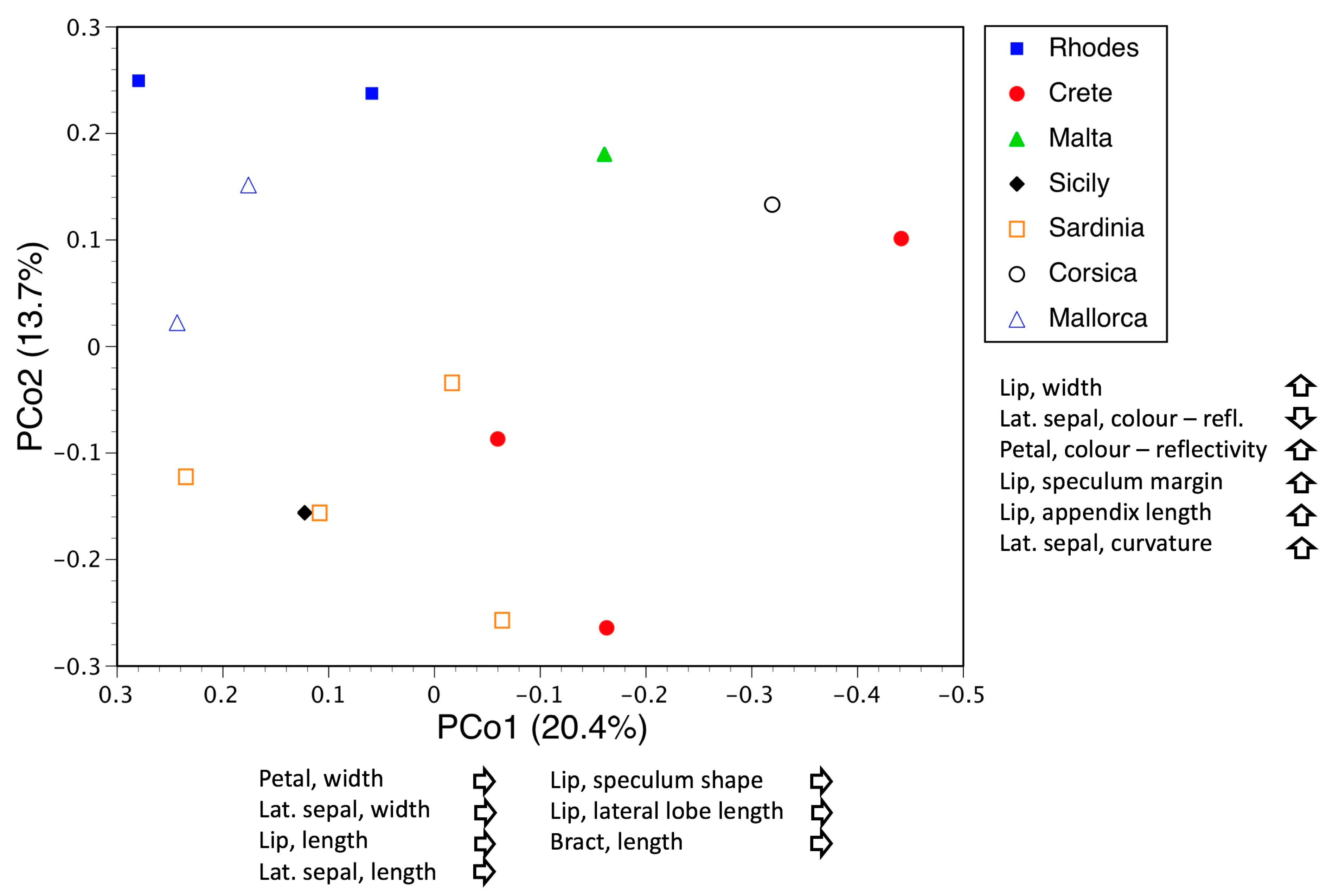
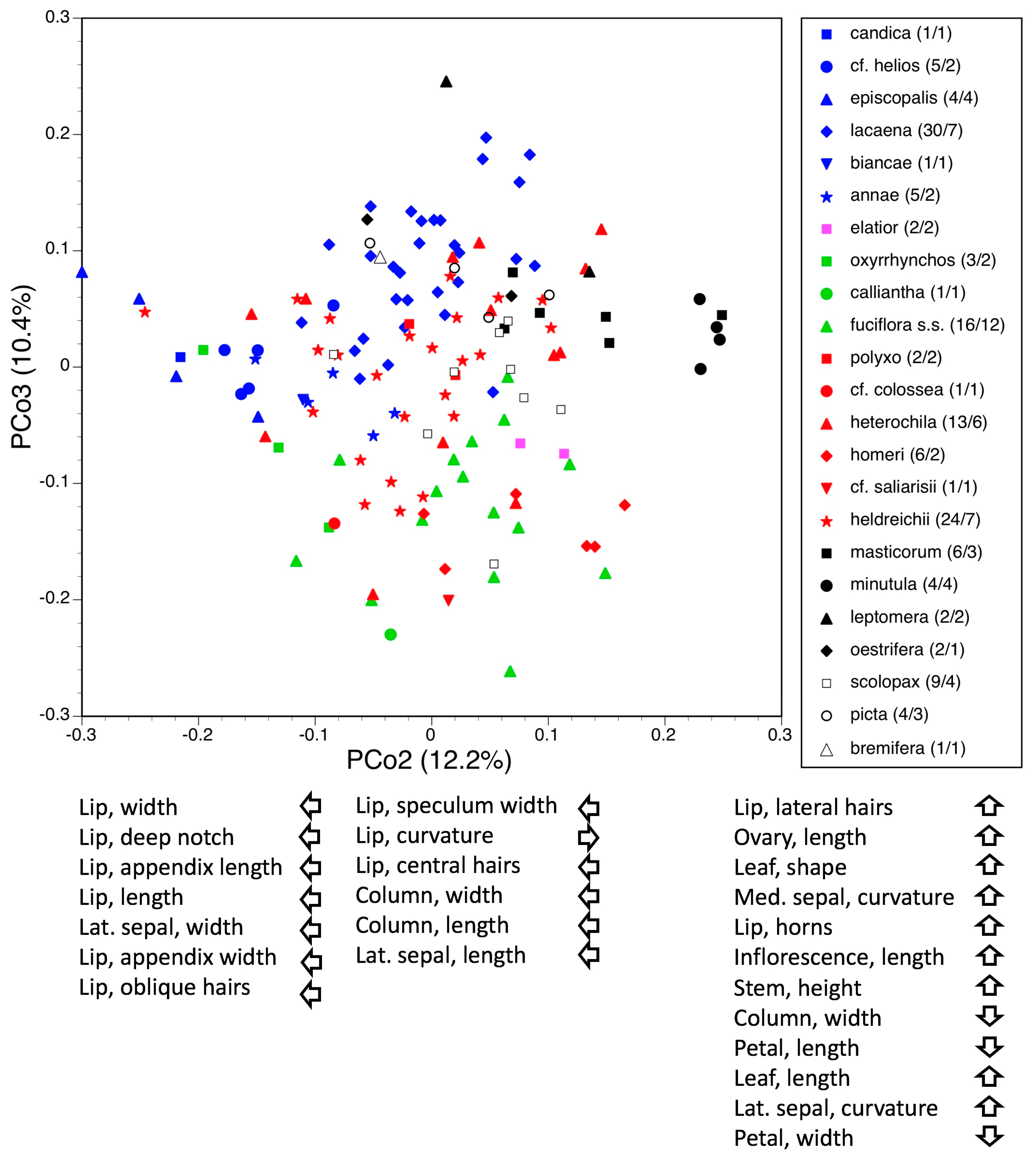
3.3.2. Macrospecies Fuciflora
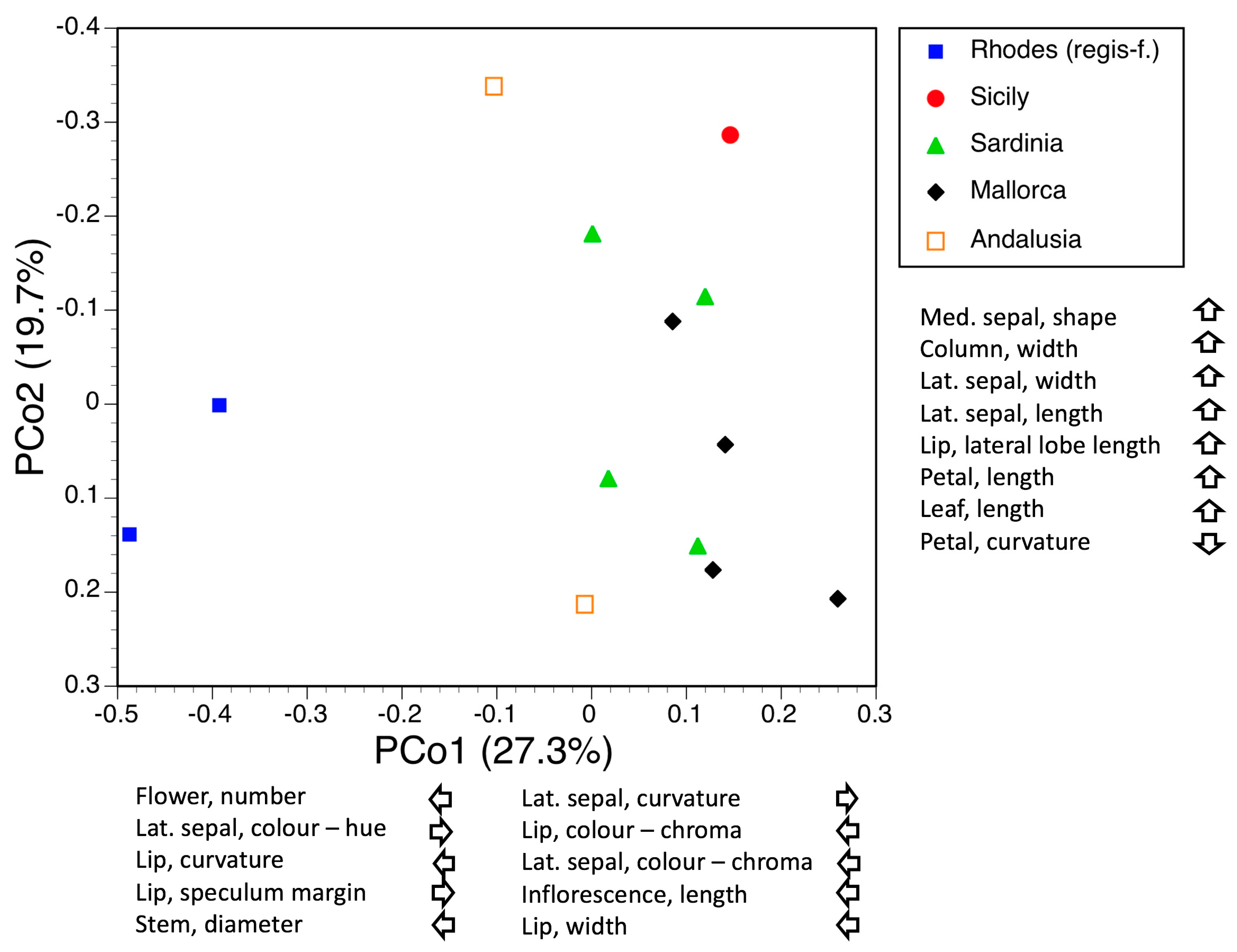
3.3.3. Macrospecies Umbilicata
3.3.4. Macrospecies Tenthredinifera
3.3.5. Macrospecies Fusca
3.4. Morphometrics: Analyses Involving Single Macrospecies and Single Mesospecies
3.4.1. Macrospecies Speculum
3.4.2. Macrospecies Bombyliflora
3.4.3. Macrospecies Apifera
3.4.4. Macrospecies Insectifera
3.5. Deeper Analysis of Macrospecies Fuciflora
4. Discussion
4.1. Strengths and Weaknesses of Our Chosen Morphometric Approach: General Principles
4.2. Strengths and Weaknesses of Our Chosen Morphometric Approach as Applied to Ophrys
4.3. Overview of Morphological Variation within Ophrys
4.3.1. The Search for Discontinuities among Taxa
4.3.2. The Search for Trends in Character Correlation
4.3.3. Residual Incongruence between Macrospecies and Mesospecies
4.4. Comparison with Morphological Variation Observed in Other European Orchid Genera
4.5. Review of Features Encouraging the Three Phases of Pseudo-Copulatory Pollination
4.5.1. Tactile Stimuli
4.5.2. Visual Stimuli: Significance (or Otherwise) of Sepal Colour
4.5.3. Olfactory Stimuli
4.5.4. Overview
4.6. Microspecies versus Macrospecies
4.6.1. Basis of the Ethological Species Concept
- (1)
- Male pollinators (most commonly solitary bees) emerge before females, are more likely than females to disperse beyond their foraging range, and are hard-wired to recognise and avoid both kin and non-receptive females.
- (2)
- There exists strong infra-specific competition among Ophrys individuals for the attraction of species-specific hymenopteran pollinators. The remarkable learning and memory abilities shown by bees in particular induce strong selection pressure for increased variation, especially in pseudo-pheromone bouquets. The novel orchid bouquets are best regarded as evolutionary darts, thrown randomly at a dartboard of many novel potential pollinators co-occurring with the orchid, in the hope of proving adaptive by securing a reasonable match with a female insect’s bouquet and thus initiating a pollinator shift into a supposed pollinator-free space.
- (3)
- Once the orchid has fortuitously generated an effective bouquet, directional selection then reinforces the attraction of the newly acquired pollinator by adaptively refining the morphology and/or phenology of the flower to better suit the pollinator.
- (4)
- Male members of the pollinating species then increasingly rely on their co-evolving Ophrys species to locate, from a distance, suitable habitats within ecologically complex landscapes, thereby increasing the probability that they will encounter a receptive female. Once males have successfully memorised the local diversity of pseudo-pheromone bouquets (and been repulsed by post-pollination alterations in the bouquets), they are more likely to disperse to other suitable habitats, thereby reducing the frequency of self-fertilisation in both the orchid species and its pollinator.
4.6.2. Reassessing the Evidence: Do Prior Assumptions Cloud Objectivity?
4.6.3. Reproductive Isolation and Lineage Separation Are Only Assumptions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) Labellum (20 characters) |
| 1. Maximum width |
| 2. Maximum length (excluding appendix) |
| 3. Depth of indentation [if present], from the maximum extent of the lateral portion of the central lobe inward to the base of the notch containing the appendix |
| 4. Maximum length of speculum |
| 5. Maximum width of speculum |
| 6. Speculum position relative to stigma (scale 1–3) (grades into stigma: connected to stigma: not connected to stigma) |
| 7. Pale zone along lower half to entire margin of speculum (scale 0–2) (absent: subdued: prominent) |
| 8. Speculum shape (scale 1–4) (entire + U + W: I I + o o: H: single ring with radiating projections + three rings) |
| X. Base colour immediately below speculum |
| 9. Colour (x) |
| 10. Colour (y) |
| 11. Colour (Y, %) |
| 12. Width of pale-coloured marginal zone [if present] of labellum |
| 13. Pilosity of central lobe margin of labellum 1 mm inside the margin and immediately above the appendix (scale 0–2) (none/negligible: short: long) |
| 14. Pilosity of central lobe margin of labellum 1 mm inside the margin and at 45° to the vertical (scale 0–2) (none/negligible: short: long) |
| 15. Pilosity of “shoulders”/lateral lobes of labellum 1 mm inside the margin (scale 0–2(none/negligible: short: long) |
| 16. Appendix length [if present] |
| 17. Appendix width [if present] |
| 18. Length of “horns” [if present] |
| 19. Maximum length of lateral lobes [if present] following mounting |
| 20. Degree of curvature of labellum viewed transversely from base (scale 1–3) (±flat: gently convex: lateral lobes strongly recurved) |
| (B) Lateral petals and sepals (16 characters) |
| 21. Length of lateral petals |
| 22. Maximum width of lateral petals |
| 23. Basal lateral teeth on lateral petals (scale 0–2) (absent: subdued: prominent) |
| X. Base colour of lateral petals |
| 24. Colour (x) |
| 25. Colour (y) |
| 26. Colour (Y, %) |
| 27. Degree of curvature of lateral petals (scale 1–5) (strongly deflexed: deflexed: ±flat: recurved: strongly recurved) |
| 28. Length of lateral sepals |
| 29. Maximum width of lateral sepals |
| X. Base colour of upper half of lateral sepals |
| 30. Colour (x) |
| 31. Colour (y) |
| 32. Colour (Y, %) |
| 33. Degree of curvature of lateral sepals (scale 1–5) (strongly deflexed: deflexed: ±flat: recurved: strongly recurved) |
| 34. Degree of curvature of median sepal (scale 1–5) (strongly deflexed: deflexed: ±flat: recurved: strongly recurved) |
| 35. Outline shape of median sepal (scale 1–3) (basally expanded obovate: ovate: apically expanded obovate) |
| 36. Suffusion of dark pigment in lower half of lateral sepal (0: 1 = absent/present) |
| (C) Column and ovary (3 characters) |
| 37. Length of ovary |
| 38. Length of column |
| 39. Maximum width of column |
| (D) Stem and inflorescence (5 characters) |
| 40. Stem height |
| 41. Stem diameter immediately above leaves |
| 42. Inflorescence length |
| 43. Number of flowers/buds |
| 44. Angle subtended by labellum relative to stem (scale 1–3) (0–30° = parallel: 31–60°: 61–90° = perpendicular) |
| (E) Leaves and bracts (7 characters) |
| 45. Number of basal (spreading) leaves |
| 46. Number of sheathing (± upright) leaves |
| 47. Length of longest basal leaf |
| 48. Maximum width of longest basal leaf |
| 49. Position of maximum with relative to position along length from base (scale 1–2) (<50%: >50%, = ovate-lanceolate: obovate leaf shapes) |
| 50. Length of basal bract |
| 51. Maximum width of basal bract |
References
- Rudall, P.J.; Bateman, R.M. Roles of synorganisation, zygomorphy and heterotopy in floral evolution: The gynostemium and labellum of orchids and other lilioid monocots. Biol. Rev. 2002, 77, 403–441. [Google Scholar] [CrossRef] [PubMed]
- Mondragon-Palomino, M. Perspectives on MADS-box expression during orchid flower evolution and development. Front. Plant Sci. 2013, 4, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, R.M.; Rudall, P.J. Hyper-epigyny is the ultimate constraint on orchid floral morphology and an ideal model for testing the Extended Synthesis. In Perspectives in Evolutionary and Developmental Biology; Fusco, G., Ed.; Padova University Press: Padova, Italy, 2019; pp. 203–233. [Google Scholar]
- Cozzolino, S.; Scopece, G. Specificity in pollination and consequences for postmating reproductive isolation in deceptive Mediterranean orchids. Phil. Trans. R. Soc. B 2008, 363, 3037–3046. [Google Scholar] [CrossRef] [Green Version]
- Claessens, J.; Kleynen, J. The Flower of the European Orchid: Form and Function; Jean Claessens & Jacques Kleynen: Voerendaal, The Netherlands, 2011. [Google Scholar]
- Bateman, R.M. Systematics and conservation of British and Irish orchids: A “state of the union” assessment to accompany Atlas 2020. Kew Bull. 2022, 77, 355–402. [Google Scholar] [CrossRef]
- Glover, B. Understanding Flowers and Flowering, 2nd ed.; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Johnson, S.D.; Schiestl, F.D. Floral Mimicry; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Kullenberg, B. Studies in Ophrys pollination. Zool. Bidr. Uppsala 1961, 34, 1–340. [Google Scholar]
- Agren, L.; Kullenberg, B.; Sensenbaugh, T. Congruences in pilosity between three species of Ophrys (Orchidaceae) and their hymenopteran pollinators. Nova Acta Reg. Soc. Sci. Uppsaliensis V 1984, 3, 5–25. [Google Scholar]
- Borg-Karlson, A.-K.; Groth, I.; Agren, L.; Kullenberg, B. Form-specific fragrances from Ophrys insectifera L. (Orchidaceae) attract species of different pollinator genera: Evidence of sympatric speciation? Chemoecology 1993, 4, 39–45. [Google Scholar] [CrossRef]
- Bradshaw, E.; Rudall, P.J.; Devey, D.S.; Thomas, M.M.; Glover, B.J.; Bateman, R.M. Comparative labellum micromorphology in the sexually deceptive temperate orchid genus Ophrys: Diverse epidermal cell types and multiple origins of structural colour. Bot. J. Linn. Soc. 2010, 162, 502–540. [Google Scholar] [CrossRef] [Green Version]
- Francisco, A.; Porto, M.; Ascensao, L. Morphological phylogenetic analysis of Ophrys (Orchidaceae): Insights from morpho-anatomical floral traits into the interspecific relationships in an unresolved clade. Bot. J. Linn. Soc. 2015, 179, 454–476. [Google Scholar] [CrossRef] [Green Version]
- Bateman, R.M.; Guy, J.J.; Rudall, P.J.; Leitch, I.J.; Pellicer, J.; Leitch, A.R. Evolutionary and functional potential of ploidy increase occurring within individual plants: Somatic ploidy mapping of the complex labellum of sexually deceptive bee orchids. Ann. Bot. 2018, 122, 133–150. [Google Scholar] [CrossRef]
- Ayasse, M.; Schiestl, F.P.; Paulus, H.F.; Löfstedt, C.; Hansson, B.; Ibarra, F.; Francke, W. Evolution of reproductive strategies in the sexually deceptive orchid Ophrys sphegodes: How does flower-specific variation of odor signals influence reproductive success? Evolution 2000, 54, 1999–2006. [Google Scholar]
- Ayasse, M.; Schiestl, F.P.; Paulus, H.F.; Ibarra, F.; Francke, W. Pollinator attraction in a sexually deceptve orchid by means on unconventional chemicals. Proc. R. Soc. Lond. B 2003, 270, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayasse, M.; Stökl, J.; Francke, W. Chemical ecology and pollinator-driven speciation in sexually deceptive orchids. Phytochemistry 2011, 72, 1667–1677. [Google Scholar] [CrossRef]
- Sedeek, K.E.M.; Whittle, E.; Guthörl, D.; Grossniklaus, U.; Shanklin, J.; Schlüter, P.M. Amino acid change in an orchid desaturase enables mimicry of the pollinator’s sex pheromone. Curr. Biol. 2016, 28, 1505–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliva, M.; Kocyan, A.; Widmer, A. Molecular phylogenetics of the sexually deceptive orchid genus Ophrys (Orchidaceae) based on nuclear and chloroplast DNA sequences. Mol. Phylog. Evol. 2001, 20, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.M.; Hollingsworth, P.M.; Preston, J.; Luo, Y.-B.; Pridgeon, A.M.; Chase, M.W. Molecular phylogenetics and evolution of Orchidinae and selected Habenariinae (Orchidaceae). Bot. J. Linn. Soc. 2003, 142, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Devey, D.S.; Bateman, R.M.; Fay, M.F.; Hawkins, J.A. Friends or relatives? Phylogenetics and species delimitation in the controversial European orchid genus Ophrys. Ann. Bot. 2008, 101, 385–402. [Google Scholar]
- Sedeek, K.E.M.; Scopece, G.; Staedler, Y.M.; Schönenberger, J.; Cozzolino, S.; Schiestl, F.P.; Schlüter, P.M. Genic rather than genomewide differences between sexually deceptive Ophrys orchids with different pollinators. Mol. Ecol. 2014, 23, 6192–6205. [Google Scholar] [CrossRef]
- Pineiro-Fernandez, L.; Byers, K.J.R.P.; Cai, J.; Sedeek, K.E.M.; Kellenberger, R.T.; Russo, A.; Qi, W.; Aquino-Fournier, C.; Schlüter, P.M. Phylogenomic analysis of the floral transcriptomes of sexually deceptive and rewarding European orchids, Ophrys and Gymnadenia. Front. Plant Sci. 2019, 10, 1553. [Google Scholar] [CrossRef]
- Breitkopf, H.; Onstein, R.E.; Cafasso, D.; Schlüter, P.M.; Cozzolino, S. Multiple shifts to different pollinators fuelled rapid diversification in sexually deceptive Ophrys orchids. New Phytol. 2015, 207, 377–386. [Google Scholar] [CrossRef]
- Baguette, M.; Betrand, J.; Stevens, V.M.; Schatz, B. Why are there so many bee-orchid species? Adaptive radiation by intraspecific competition for mnemonic pollinators. Biol. Rev. 2020, 95, 1630–1663. [Google Scholar] [CrossRef]
- Bertrand, J.; Baguette, M.N.; Schatz, B. Challenges inherent in the systematics and taxonomy of genera that have recently experienced explosive radiation: The case of orchids of the genus Ophrys. In Systematics and the Exploration of Life; Grandcolas, P., Maurel, M.-C., Eds.; Wiley/Iste: New York, NY, USA, 2021; pp. 113–134. [Google Scholar]
- Bateman, R.M.; Bradshaw, E.; Devey, D.S.; Glover, B.J.; Malmgren, S.; Sramkó, G.; Thomas, M.M.; Rudall, P.J. Species arguments: Clarifying concepts of species delimitation in the pseudo-copulatory orchid genus Ophrys. Bot. J. Linn. Soc. 2011, 165, 336–347. [Google Scholar] [CrossRef]
- Bateman, R.M. Two bees or not two bees? An overview of Ophrys systematics. Ber. Arbeitskr. Heim. Orchideen 2018, 35, 5–46. [Google Scholar]
- Bateman, R.M. Species circumscription in ‘cryptic’ clades: A nihilist’s view. In Cryptic Species: Morphological Stasis, Circumscription and Hidden Diversity; Monro, A.K., Mayo, S.J., Eds.; Cambridge University Press: Cambridge, UK, 2022; pp. 36–77. [Google Scholar]
- Vereecken, N.J.; Streinzer, M.; Ayasse, M.; Spaethe, J.; Paulus, H.F.; Stökl, J.; Cortis, P.; Schiestl, F.P. Integrating past and present studies on Ophrys pollination—A comment on Bradshaw et al. Bot. J. Linn. Soc. 2011, 165, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Cuypers, V.; Reydon, T.A.C.; Artois, T. Deceiving insects, deceiving taxonomists? Making theoretical sense of taxonomic disagreement in the European orchid genus Ophrys. Persp. Plant Ecol. Evol. Syst. 2022, 56, 125686. [Google Scholar] [CrossRef]
- Nelson, E. Monographie und Ikonographie der Gattung Ophrys; E. Nelson: Chernex-Montreux, Switzerland, 1962. [Google Scholar]
- De Soó, R. Dactylorhiza Necker ex Nevski. In Flora Europaea 5; Tutin, T.G., Heywood, V.H., Burgess, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1980; pp. 333–337. [Google Scholar]
- Sundermann, H. Europäische und Mediterrane Orchideen—Ein Bestimmungsflora, 3rd ed.; Schmersow: Hildesheim, Germany, 1980. [Google Scholar]
- Pedersen, H.; Faurholdt, N. Ophrys: The Bee Orchids of Europe; Kew Publishing: Kew, UK, 2007. [Google Scholar]
- Kühn, R.; Pedersen, H.A.; Cribb, P. Field Guide to the Orchids of Europe and the Mediterranean; Royal Botanic Gardens Kew: Kew, UK, 2019. [Google Scholar]
- Devillers, P.; Devillers-Terschuren, J. Essai d’analyse systématique du genre Ophrys. Nat. Belges 1994, 75, 273–400. [Google Scholar]
- Delforge, P. Orchidées d’Europe, d’Afrique du Nord et du Proche-Orient, 1st ed.; Delachaux et Niéstle: Paris, France, 1994. [Google Scholar]
- Delforge, P. Orchidées d’Europe, d’Afrique du Nord et du Proche-Orient, 4th ed.; Delachaux et Niéstle: Paris, France, 2016. [Google Scholar]
- Paulus, H.F. Deceived males—Pollination biology of the Mediterranean orchid genus Ophrys (Orchidaceae). J. Eur. Orch. 2006, 38, 303–353. [Google Scholar]
- Paulus, H.F. Bestäuber als Isolationsmechanismen: Freilandbeobachtungen und Experimente zur Spezifität der Bestäuberanlockung in der Gattung Ophrys (Orchidaceae und Insecta, Hymenoptera, Apoidea). Ber. Arbeitskr. Heim. Orchid. 2015, 32, 142–199. [Google Scholar]
- Paulus, H.F. Pollinators as isolation mechanisms: Field observations and field experiments regarding specificity of pollinator attraction in the genus Ophrys (Orchidaceae und Insecta, Hymenoptera, Apoidea). Entom. Gener. 2018, 37, 261–316. [Google Scholar] [CrossRef]
- Bateman, R.M.; Sramkó, G.; Paun, O. Integrating restriction site-associated DNA sequencing (RAD-seq) with morphological cladistic analysis clarifies evolutionary relationships among major species groups of bee orchids. Ann. Bot. 2018, 121, 85–105. [Google Scholar] [CrossRef] [Green Version]
- Tyteca, D.; Baguette, M. Ophrys (Orchidaceae) systematics: When molecular phylogenetics, morphology and biology reconcile. Ber. Arbeitskr. Heim. Orchideen 2017, 34, 37–103. [Google Scholar]
- Bateman, R.M.; Rudall, P.J.; Murphy, A.R.M.; Cowan, R.S.; Devey, D.S.; Perez-Escobar, O.A. Even whole plastomes are not enough: Phylogenomic and morphometric exploration at multiple demographic levels of the bee orchid clade Ophrys sect. Sphegodes. J. Exp. Bot. 2021, 72, 654–681. [Google Scholar] [CrossRef] [PubMed]
- Kreutz, C.A.J. Kompendium der Europäischen Orchideen; Kreutz Publishers: Landgraaf, The Netherlands, 2004. [Google Scholar]
- Baumann, H.; Künkele, S.; Lorenz, R. Orchideen Europas; Ulmer: Stuttgart, Germany, 2006. [Google Scholar]
- Mayr, E. Systematics and the Origin of Species; Columbia University Press: New York, NY, USA, 1942. [Google Scholar]
- Mayr, E. Speciation and macroevolution. Evolution 1982, 36, 1119–1132. [Google Scholar] [CrossRef] [PubMed]
- Mayden, R.L. A hierarchy of species concepts: The denouement in the saga of the species problem. In Species: The Units of Biodiversity; Claridge, M.F., Dawah, H.A., Wilson, M.R., Eds.; Chapman & Hall: London, UK, 1997; pp. 381–421. [Google Scholar]
- Wheeler, Q.D.; Meier, R. (Eds.) Species Concepts and Phylogenetic Theory: A Debate; Columbia University Press: New York, NY, USA, 2000. [Google Scholar]
- Rieseberg, L.H.; Wood, T.E.; Baack, E.J. The nature of plant species. Nature 2006, 440, 524–527. [Google Scholar] [CrossRef]
- de Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Hart, M.W. The species concept as an emergent property of population biology. Evolution 2011, 65, 613–616. [Google Scholar] [CrossRef]
- Hausdorf, B. Progress toward a general species concept. Evolution 2011, 65, 923–931. [Google Scholar] [CrossRef]
- Mishler, B.D. What, If Anything, Are Species? CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Monro, A.K.; Mayo, S.J. (Eds.) Cryptic Species: Morphological Stasis, Circumscription and Hidden Diversity; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Stökl, J.; Schlüter, P.M.; Stuessy, T.F.; Paulus, H.F.; Assum, G.; Ayasse, M. Scent variation and hybridization cause the displacement of a sexually deceptive orchid species. Am. J. Bot. 2008, 95, 472–481. [Google Scholar] [CrossRef]
- Triponez, Y.; Arrigo, N.; Pelissier, L.; Schatz, B.; Alvarez, N. Morphological, ecological and genetic aspects associated with endemism in the Fly Orchid group. Mol. Ecol. 2013, 22, 1431–1446. [Google Scholar] [CrossRef]
- Gögler, J.; Stökl, J.; Cortis, P.; Beyrle, H.; Barone Lumaga, M.R.; Cozzolino, S.; Ayasse, M. Increased divergence in floral morphology strongly reduces gene flow in sympatric sexually deceptive orchids with the same pollinator. Evol. Ecol. 2015, 29, 703–717. [Google Scholar] [CrossRef]
- Gögler, J.; Zitari, A.; Paulus, H.; Cozzolino, S.; Ayasse, M. Species boundaries in the Ophrys iricolor group in Tunisia: Do local endemics always matter? Plant Syst. Evol. 2016, 302, 481–489. [Google Scholar] [CrossRef]
- Véla, E.; Rebbas, K.; Martin, R.; Premorel, G.; de Tison, J.-M. Waiting for integrative taxonomy: Morphospecies as an operational proxy for the radiative and reticulate genus Ophrys L. (Orchidaceae)? Eur. J. Environ. Sci. 2015, 5, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Lussu, M.; De Agostini, A.; Marignani, M.; Cogoni, A.; Cortis, P. Ophrys annae and Ophrys chestermanniae: An impossible love between two orchid sister species. Nordic J. Bot. 2018, 36, e01798. [Google Scholar] [CrossRef]
- Cozzolino, S.; Scopece, G.; Cortis, P.; Barone Lumaga, M.R.; Cafasso, D.; Ayasse, M. Reproductive character displacement allows two sexually deceptive orchids to coexist and attract the same specific pollinator. Evol. Ecol. 2022, 36, 217–232. [Google Scholar] [CrossRef]
- Turco, A.; Medagli, P.; Wagensommer, R.P.; D’Emerico, S.; Gennaio, R.; Albano, A. A morphometric study on Ophrys sect. Pseudophrys in Apulia (Italy) and discovery of Ophrys japigiae. Plant Biosyst. 2022, 156, 560–571. [Google Scholar]
- Bateman, R.M.; Denholm, I. A reappraisal of the British and Irish dactylorchids, 1. The tetraploid marsh-orchids. Watsonia 1983, 14, 347–376. [Google Scholar]
- Bateman, R.M. Evolution and classification of European orchids: Insights from molecular and morphological characters. J. Eur. Orch. 2001, 33, 33–119. [Google Scholar]
- Bateman, R.M. The perils of addressing long-term challenges in a short-term world: Making descriptive taxonomy predictive. In Climate Change, Ecology and Systematics; Hodkinson, T.R., Jones, M.B., Waldren, S., Parnell, J.A.N., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 67–95. [Google Scholar]
- Bateman, R.M.; Rudall, P.J.; Denholm, I. In situ morphometric survey elucidates the evolutionary systematics of the orchid genus Gymnadenia in the British Isles. Syst. Biodiv. 2021, 19, 571–600. [Google Scholar] [CrossRef]
- Payne, R.W.; Harding, S.A.; Murray, D.A.; Souter, D.M.; Baird, D.B.; Glaser, A.I.; Welham, S.J.; Gilmour, A.R.; Thompson, R.; Webster, R. (Eds.) Genstat v14; VSN International: Hemel Hempstead, UK, 2011. [Google Scholar]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–872. [Google Scholar] [CrossRef]
- Gower, J.C.; Ross, G.J.S. Minimum spanning trees and single linkage cluster analysis. J. R. Stat. Soc. C 1969, 18, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Gower, J.C. Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika 1966, 52, 325–338. [Google Scholar] [CrossRef]
- Gower, J.C. Measures of similarity, dissimilarity and distance. In Encyclopedia of Statistical Sciences 5; Wiley: New York, NY, USA, 1985; pp. 397–405. [Google Scholar]
- Devey, D.S.; Bateman, R.M.; Fay, M.F.; Hawkins, J.A. Genetic structure and systematic relationships within the Ophrys fuciflora aggregate (Orchidinae: Orchidaceae): High diversity in Kent and a wind-induced discontinuity bisecting the Adriatic. Ann. Bot. 2009, 104, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.M.; Rudall, P.J. The life and death of a mythical British endemic, Orchis militaris L. var. tenuifrons P.D. Sell: Why infraspecific taxonomy requires a field-based morphometric approach. N. J. Bot. 2011, 1, 98–110. [Google Scholar]
- Bateman, R.M.; Denholm, I.; McLeod, L.; Craig, W.; Ennos, R.A. Systematic reappraisal of Marsh-orchids native to Scotland. Kew Bull. 2023, 79, 21. [Google Scholar]
- Bateman, R.M. Phenotypic versus genotypic disparity in the Eurasian orchid genus Gymnadenia: Exploring the limits of phylogeny reconstruction. Syst. Biodiv. 2021, 19, 400–422. [Google Scholar] [CrossRef]
- Bateman, R.M.; James, K.E.; Rudall, P.J. Contrast in morphological versus molecular divergence between two closely related Eurasian species of Platanthera (Orchidaceae) suggests recent evolution with a strong allometric component. New J. Bot. 2012, 2, 110–148. [Google Scholar] [CrossRef]
- Bateman, R.M.; Rudall, P.J.; Moura, M. Systematic revision of Platanthera in the Azorean archipelago: Not one but three species, including arguably Europe’s rarest orchid. PeerJ 2013, 1, e218. [Google Scholar] [CrossRef] [Green Version]
- Bateman, R.M.; Smith, R.J.; Fay, M.F. Morphometric and population-genetic analyses elucidate the origin, evolutionary significance and conservation implications of Orchis ×angusticruris (O. purpurea × O. simia), a hybrid orchid new to Britain. Bot. J. Linn. Soc. 2008, 157, 687–711. [Google Scholar] [CrossRef] [Green Version]
- Bateman, R.M.; Molnár, A.V.; Sramkó, G. In situ morphometric survey elucidates the evolutionary systematics of the Eurasian Himantoglossum clade (Orchidaceae: Orchidinae). PeerJ 2017, 5, e2893. [Google Scholar] [CrossRef] [Green Version]
- Bateman, R.M.; Rudall, P.J.; Bidartondo, M.I.; Cozzolino, S.; Tranchida-Lombardo, V.; Carine, M.A.; Moura, M. Speciation via floral heterochrony and presumed mycorrhizal host-switching of endemic butterfly orchids on the Azorean archipelago. Am. J. Bot. 2014, 101, 979–1001. [Google Scholar] [CrossRef]
- Jin, W.-T.; Schuiteman, A.; Chase, M.W.; Li, J.-W.; Chung, S.-W.; Hsu, T.-C.; Jin, X.-H. Phylogenetics of subtribe Orchidinae s.l. (Orchidaceae; Orchidoideae) based on seven markers (plastid matK, psaB, rbcL, trnL-F, trnH-psba, and nuclear nrITS, Xdh): Implications for generic delimitation. BMC Plant Biol. 2017, 17, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, A.K.; Roberts, D.L.; Hawkins, J.A.; Rudall, P.J.; Box, M.S.; Bateman, R.M. Comparative morphology of nectariferous and nectarless labellar spurs in selected clades of subtribe Orchidinae (Orchidaceae). Bot. J. Linn. Soc. 2009, 160, 369–387. [Google Scholar] [CrossRef] [Green Version]
- Shipunov, A.B.; Bateman, R.M. Geometric morphometrics as a tool for understanding Dactylorhiza (Orchidaceae) diversity in European Russia. Biol. J. Linn. Soc. 2005, 85, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Breitkopf, H.; Schlüter, P.M.; Xu, S.; Schiestl, F.P.; Cozzolino, S.; Scopece, G. Pollinator shifts between Ophrys sphegodes populations: Might adaptation to different pollinators drive population divergence? J. Evol. Biol. 2013, 26, 2197–2208. [Google Scholar] [CrossRef] [PubMed]
- Bersweden, L.; Viruel, J.; Schatz, B.; Harland, J.; Gargiulo, R.; Cowan, R.S.; Calevo, J.; Juan, A.; Clarkson, J.J.; Leitch, A.R.; et al. Microsatellites and petal morphology reveal new patterns of admixture in Orchis hybrid zones. Am. J. Bot. 2021, 108, 1388–1404. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.M.; Farrington, O.S. A morphometric study of XOrchiaceras bergonii (Nanteuil) Camus and its parents (Aceras anthropophorum (L.) Aiton f. and Orchis simia Lamarck) in Kent. Watsonia 1987, 16, 397–407. [Google Scholar]
- Bateman, R.M.; Farrington, O.S. Morphometric comparison of populations of Orchis simia Lam. (Orchidaceae) from Oxfordshire and Kent. Bot. J. Linn. Soc. 1989, 100, 205–218. [Google Scholar] [CrossRef]
- Bateman, R.M.; Denholm, I. Taxonomic reassessment of the British and Irish tetraploid marsh-orchids. New J. Bot. 2012, 2, 37–55. [Google Scholar] [CrossRef]
- Paun, O.; Bateman, R.M.; Luna, J.A.; Moat, J.; Fay, M.F.; Chase, M.W. Altered gene expression and ecological divergence in sibling allopolyploids of Dactylorhiza (Orchidaceae). BMC Evol. Biol. 2011, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Balao, F.; Trucchi, E.; Wolfe, T.M.; Hao, B.-H.; Lorenzo, M.T.; Baar, J.; Sedman, L.; Kosiol, C.; Amman, F.; Chase, M.W.; et al. Adaptive sequence evolution is driven by biotic stress in a pair of orchid species (Dactylorhiza) with distinct ecological optima. Mol. Ecol. 2017, 26, 3649–3662. [Google Scholar] [CrossRef] [Green Version]
- Brandrud, M.K.; Baar, J.; Lorenzo, M.T.; Athanasiadis, A.; Bateman, R.M.; Chase, M.W.; Hedrén, M.; Paun, O. Phylogenomic relationships of diploids and the origins of allotetraploids in Dactylorhiza (Orchidaceae): RADseq data track reticulate evolution. Syst. Biol. 2020, 61, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Chapurlat, E.; Le Ronce, I.; Agren, J.; Sletvold, N. Divergent selection on flowering phenology but not on floral morphology between two closely related orchids. Ecol. Evol. 2020, 10, 5547–5737. [Google Scholar] [CrossRef] [PubMed]
- Hedrén, M.; Lorenz, R.; Teppner, H.; Dolinar, B.; Giotta, C.; Griebl, N.; Hansson, S.; Heidtke, U.; Klein, E.; Perazza, G.; et al. Evolution and systematics of polyploid Nigritella (Orchidaceae). Nordic J. Bot. 2018, 36, 01539. [Google Scholar] [CrossRef]
- Stejskal, K.; Streinzer, M.; Dyer, A.; Paulus, H.F.; Spaethe, J. Functional significance of labellum pattern variation in a sexually deceptive orchid (Ophrys heldreichii): Evidence of individual signature learning effects. PLoS ONE 2015, 10, e0142971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francisco, A.; Ascensao, L. Structure of the osmophore and labellum micromorphology in the sexually deceptive orchids Ophrys bombyliflora and Ophrys tenthredinifera (Orchidaceae). Int. J. Plant Sci. 2013, 174, 619–636. [Google Scholar] [CrossRef]
- Cortis, P.; Vereecken, N.J.; Schiestl, F.P.; Barone Lumaga, M.R.; Scrugli, A.; Cozzolino, S. Pollinator convergence and the nature of species’ boundaries in sympatric Sardinian Ophrys (Orchidaceae). Ann. Bot. 2009, 104, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakosy, D.; Cuervo, M.; Paulus, H.F.; Ayasse, M. Looks matter: Changes in floral form affect pollination effectiveness in a sexually deceptive orchid. J. Evol. Biol. 2017, 30, 1978–1993. [Google Scholar] [CrossRef] [Green Version]
- Spaethe, J.; Moser, W.H.; Paulus, H.F. Increase of pollinator attraction by means of a visual signal in the sexually deceptive orchid Ophrys heldreichii (Orchidaceae). Plant Syst. Evol. 2007, 264, 31–40. [Google Scholar] [CrossRef]
- Streinzer, M.; Paulus, H.F.; Spaethe, J. Floral colour signal increases short-range detectability of a sexually deceptive orchid to its bee pollinator. J. Exp. Biol. 2009, 212, 1365–1370. [Google Scholar] [CrossRef] [Green Version]
- Rakosy, D.; Streinzer, M.; Paulus, H.F.; Spathe, J. Floral visual signal increases reproductive success in a sexually deceptive orchid. Arthropod-Plant Interac. 2012, 6, 671–681. [Google Scholar] [CrossRef] [Green Version]
- Streinzer, M.; Ellis, T.; Paulus, H.F.; Spathe, J. Visual discrimination between two sexually deceptive Ophrys species by a bee pollinator. Arthropod-Plant Interac. 2010, 4, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vereecken, N.J.; Schiestl, F.P. On the roles of colour and scent in a specialized floral mimicry system. Ann. Bot. 2009, 104, 1077–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kevan, P.; Giurfa, M.; Chittka, L. Why are there so many and so few white flowers? Trends Plant Sci. 1996, 1, 280–284. [Google Scholar] [CrossRef]
- Vignolini, S.; Davey, M.P.; Bateman, R.M.; Rudall, P.J.; Moyroud, E.; Tratt, J.; Malmgren, S.; Steiner, U.; Glover, B.J. The mirror crack’d: Both structure and pigment contribute to the metallic blue appearance of the Mirror Orchid, Ophrys speculum. New Phyt. 2012, 196, 1038–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmgren, S. Are there 25 or 250 Ophrys species? J. Hardy Orchid Soc. 2008, 5, 95–100. [Google Scholar]
- Spaethe, J.; Streinzer, M.; Paulus, H.F. Why sexually deceptive orchids have colored flowers. Commun. Integr. Biol. 2010, 3, 139–141. [Google Scholar] [CrossRef]
- Joffard, N.; Arnal, V.; Buatois, B.; Schatz, B.; Montgelard, C. Floral scent evolution in the section Pseudophrys: Pollinator-mediated selection or phylogenetic constraints? Plant Biol. 2020, 20, 881–889. [Google Scholar] [CrossRef]
- Schiestl, F.P.; Cozzolino, S. Evolution of sexual mimicry in the orchid subtribe Orchidinae: The role of preadaptations in the attraction of male bees as pollinators. BMC Evol. Biol. 2008, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Mant, J.; Peakall, R.; Schiestl, F.P. Does selection on floral odor promote differentiation among populations and species of the sexually deceptive orchid genus Ophrys? Evolution 2005, 59, 1449–1463. [Google Scholar]
- Vereeckren, N.J.; Mant, J.; Schiestl, F.P. Population differentiation in female sex pheromone and male preferences in a solitary bee. Behav. Ecol. Sociobiol. 2007, 61, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Vereecken, N.J.; Schiestl, F.P. The evolution of imperfect floral mimicry. Proc. Natl. Acad. Sci. USA 2008, 105, 7484–7488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlüter, P.M.; Ruas, P.M.; Kohl, G.; Ruas, C.F.; Stuessy, T.F.; Paulus, H.F. Evidence for progenitor-driven speciation in sexually deceptive orchids. Ann. Bot. 2011, 108, 895–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Schlüter, P.M.; Scopece, G.; Breitkopf, H.; Gross, K.; Cozzolino, S.; Schiestl, F.P. Floral isolation is the main reproductive barrier among closely related sexually deceptive orchids. Evolution 2011, 65, 2606–2620. [Google Scholar] [CrossRef] [PubMed]
- Paulus, H.F. Speciation, pattern recognition and the maximization of pollination: General questions and answers given by the reproductive biology of the orchid genus Ophrys. J. Comp. Physiol. A 2019, 205, 285–300. [Google Scholar] [CrossRef] [Green Version]
- Vereecken, N.J. Deceptive behaviour in plants. I. Pollination by sexual deception in orchids: A host–parasite perspective. In Plant–Environment Interactions; Baluska, F., Ed.; Springer: Berlin, Germany, 2009; pp. 203–222. [Google Scholar]
- Ayasse, M.; Gögler, J.; Stökl, J. Pollinator-driven speciation in sexually deceptive orchids of the genus Ophrys. In Evolution in Action; Glaubrecht, M., Ed.; Springer: Berlin, Germany, 2010; pp. 101–118. [Google Scholar]
- Xu, S.; Schlüter, P.M.; Schiestl, F.P. Pollinator-driven speciation in sexually deceptive orchids. Int. J. Ecol. 2012, 2012, 285081. [Google Scholar] [CrossRef] [Green Version]
- Schiestl, F.P.; Johnson, S.D. Pollinator-mediated evolution of floral signals. Trends Ecol. Evol. 2013, 28, 307–315. [Google Scholar] [CrossRef]
- Vereecken, N.J.; Francisco, A. Ophrys pollination: From Darwin to the present day. In Darwin’s Orchids: Then and Now; Edens-Meier, R., Bernhardt, P., Eds.; University of Chicago Press: Chicago, IL, USA, 2014; pp. 47–69. [Google Scholar]
- Schatz, B.; Genoud, D.; Esudié, P.; Geniez, P.; Wünch, K.G.; Joffard, N. Is Ophrys pollination more opportunistic than previously thought? Insights from different field methods of pollinator observation. Bot. Lett. 2021, 168, 333–347. [Google Scholar] [CrossRef]
- Wong, B.B.M.; Schiestl, F.P. How an orchid harms its pollinator. Proc. R. Soc. Lond. B 2002, 269, 1529–1532. [Google Scholar] [CrossRef]
- Stökl, J.; Schlüter, P.M.; Steussy, T.F.; Paulus, H.F.; Fraberger, R.; Erdmann, D.; Ayasse, M. Speciation in sexually deceptive orchids: Pollinator-driven selection maintains discrete odour phenotypes in hybridizing species. Biol. J. Linn. Soc. 2009, 98, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Devillers-Terschuren, J.; Devillers, P. Nomenclatural and taxonomic contributions to European orchids (Orchidaceae). Nat. Belges 1991, 72, 99–101. [Google Scholar]
- Vereecken, N.J.; Cozzolino, S.; Schiestl, F.P. Hybrid floral scent novelty drives pollinator shift in sexually deceptive orchids. BMC Evol. Biol. 2010, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermosilla, C.E.; Amardeilh, J.P.; Soca, R. Sterictiphora furcata Villers, pollinisateur d’Ophrys subinsectifera Hermosilla & Sabando. L’Orchidophile 1999, 139, 247–254. [Google Scholar]
- Joffard, N.; Buatois, B.; Schatz, B. Integrative taxonomy of the fly orchid group: Insights from chemical ecology. Sci. Nat. 2016, 103, 77. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, D.L.; Selosse, M.A.; Sauve, M.; Francke, W.; Vereecken, N.J.; Cozzolino, S.; Schiestl, F.P. Floral scent and species divergence in a pair of sexually deceptive orchids. Ecol. Evol. 2017, 7, 6023–6034. [Google Scholar] [CrossRef]
- Pausic, I.; Lipovsek, M.; Jakely, D.; Pavlec, N.; Ivajnsic, D.; Kaligaric, M. Local climate and latitude affect flower form of Ophrys fuciflora (Orchidaceae): Evidence for clinal variation. Bot. Lett. 2019, 166, 499–512. [Google Scholar] [CrossRef]
- Hennecke, M.; Galanos, C. Ophrys subgen. Bombyliflorae sect. Tenthrediniferae: A synopsis. GIROS 2020, 63, 358–419. [Google Scholar]
- Cozzolino, S.; Scopece, G.; Roma, L.; Schlüter, P.M. Different filtering strategies of genotyping-by-sequencing data provide complementary resolutions of species boundaries and relationships in a clade of sexually deceptive orchids. J. Syst. Evol. 2020, 58, 133–144. [Google Scholar] [CrossRef]
- Joffard, N.; Buatois, B.; Arnal, V.; Véla, E.; Montgelard, C.; Schatz, B. Delimiting species in the taxonomically challenging orchid section Pseudophrys: Bayesian analyses of genetic and phenotypic data. Front. Ecol. Evol. 2022, 10, 1058550. [Google Scholar] [CrossRef]
- Ruiz-Perez, D.; Guan, H.; Madhivanan, P.; Mathee, K.; Narasimhan, G. So you think you can PLS-DA? BMC Bioinf. 2020, 21, 2. [Google Scholar] [CrossRef]
- Gölz, P.; Reinhard, H. Bemerkungen zu “Baumann, H. & S. Künkele (1986): Die Gattung Ophrys L.—Eine taxonomische Übersicht”. Mitt. Bl. Arbeitskrs. Heim. Orch. Baden-Württ. 1987, 19, 866–894. [Google Scholar]
- Devillers, P.; Devillers-Terschuren, J.; Tyteca, D. Notes on some of the taxa comprising the group of Ophrys tenthredinifera Willdenow. J. Eur. Orch. 2003, 35, 109–161. [Google Scholar]
- Gögler, J.; Stökl, J.; Sramkova, A.; Twele, R.; Francke, W.; Cozzolino, S.; Cortis, P.; Scrugli, A.; Ayasse, M. Ménage a trois—Two endemic species of deceptive orchids and one pollinator species. Evolution 2009, 63, 2222–2234. [Google Scholar] [CrossRef] [PubMed]
- Paulus, H.F.; Hirth, M. Bestäubungsbiologie und Systematik der Ophrys tenthredinifera-Artengruppe in der Ostägäis (Orchidaceae, Insecta). J. Eur. Orch. 2012, 44, 625–686. [Google Scholar]
- Wucherpfennig, W. Die westmediterranen Arten der Ophrys tenthredinifera-Gruppe ein Vergleich. Ber. Arbeitskrs. Heim. Orch. 2016, 33, 110–150. [Google Scholar]
- Sedeek, K.E.M.; Qi, W.; Schauer, M.A.; Gupta, A.K.; Poveda, L.; Xu, S.; Liu, Z.-J.; Grossniklaus, U.; Schiestl, F.P.; Schlüter, P.M. Transcriptome and proteome data reveal candidate genes for pollinator attraction in sexually deceptive orchids. PLoS ONE 2013, 8, e64621. [Google Scholar] [CrossRef]
- Paun, O.; Bateman, R.M.; Fay, M.F.; Hedrén, M.; Civeyrel, L.; Chase, M.W. Stable epigenetic effects impact evolution and adaptation in allopolyploid orchids. Mol. Biol. Evol. 2010, 27, 2465–2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, T.M.; Balao, F.; Trucchi, E.; Bachmann, G.; Gu, W.; Baar, J.; Hedrén, M.; Weckwerth, W.; Leitch, A.R.; Paun, O. Recurrent allopolyploidization events diversify eco-physiological traits in marsh orchids. bioRxiv 2021. [Google Scholar] [CrossRef]


















| RAD-Seq (and ITS) Groups | Number of Sampled: | ||||
|---|---|---|---|---|---|
| Plants | Microspecies | Mesospecies | Localities | Countries | |
| Insectifera (A) | 14 | 1 | 1 | 6 | 3 |
| Speculum (C) | 13 | 2 | 1 | 12 | 3 |
| Bombyliflora (D) | 14 | 1 | 1 | 14 | 5 |
| Tenthredinifera (B) | 31 | 11 | 1 | 27 | 5 |
| Fusca (E) | 51 | 33 | 8 | 46 | 7 |
| Apifera (F) | 15 | 1 | 1 | 15 | 6 |
| Umbilicata (J) | 52 | 8 * | 1 * | 44 | 3 |
| Fuciflora (H + I) | 143 | 23 | 6 | 68 | 8 |
| Sphegodes (G) | 124 | 33 | 9 | 104 | 7 |
| TOTAL | 457 | 113 | 29 |
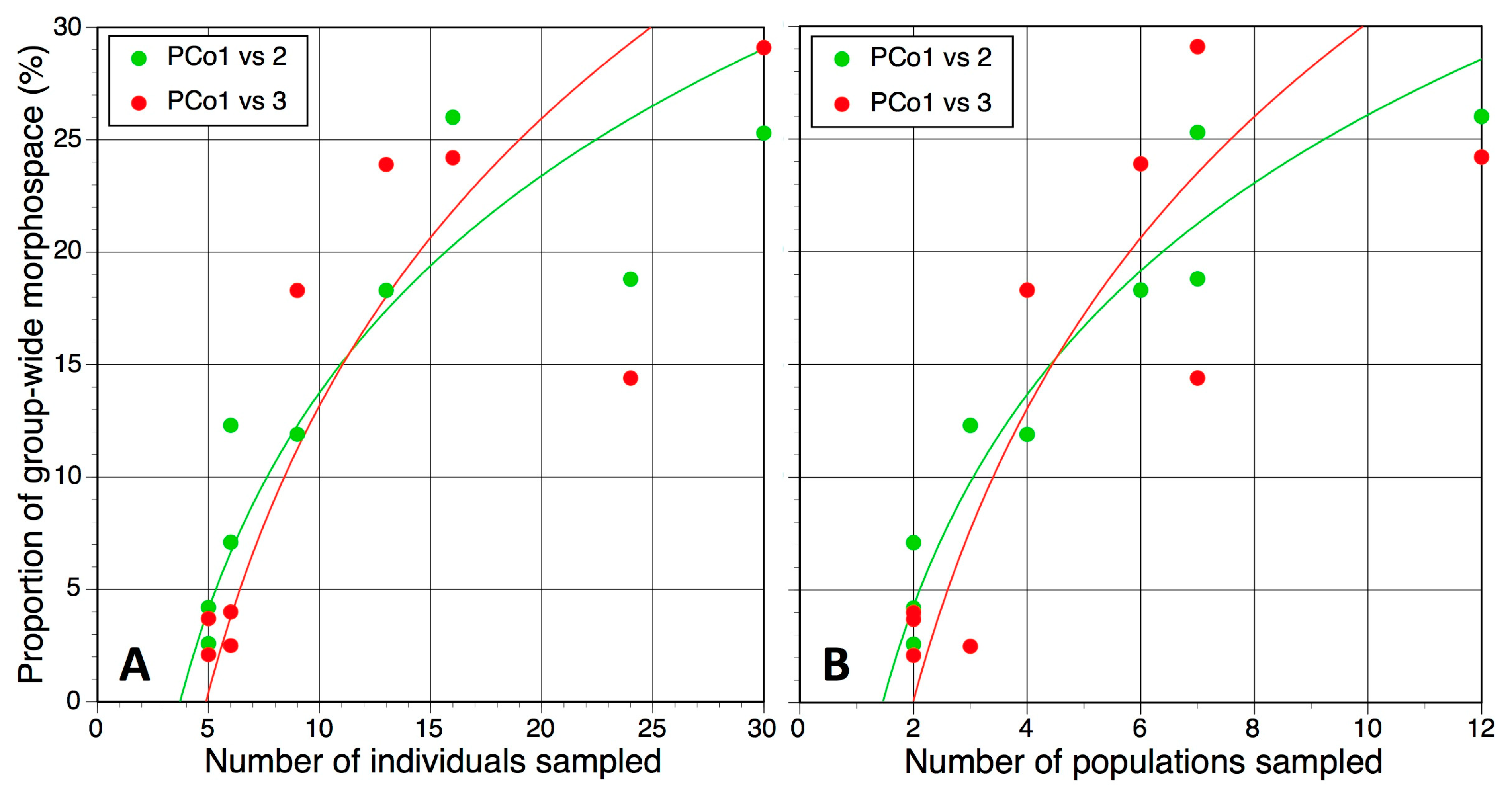
| Coordinates Plotted (and % of Total Variance Represented by Plot) | Mean ± SD (and Range) of Proportion of Group-Wide Variation (PGWV: %) | PGWV vs. Sampling Effort, Microspecies (–ve expl. r2) | ||
|---|---|---|---|---|
| Microspecies | Mesospecies | Individuals | Populations | |
| PCo1 vs. 2 (27.1) | 12.8 ± 14.1 (2.6–25.3) | 40.7 ± 21.4 (20.9–66.6) | 0.82 | 0.92 |
| PCo1 vs. 3 (25.3) | 13.6 ± 10.8 (2.1–29.1) | 38.5 ± 14.3 (24.3–54.2) | 0.73 | 0.76 |
| PCo2 vs. 3 (22.6) | 13.4 ± 11.8 (1.2–35.8) | 32.7 ± 13.6 (18.6–52.3) | 0.53 | 0.67 |
| PCo3 vs. 4 (18.5) | 12.9 ± 11.2 (1.0–27.5) | 33.4 ± 12.8 (19.4–44.9) | 0.72 | 0.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bateman, R.M.; Rudall, P.J. Morphological Continua Make Poor Species: Genus-Wide Morphometric Survey of the European Bee Orchids (Ophrys L.). Biology 2023, 12, 136. https://doi.org/10.3390/biology12010136
Bateman RM, Rudall PJ. Morphological Continua Make Poor Species: Genus-Wide Morphometric Survey of the European Bee Orchids (Ophrys L.). Biology. 2023; 12(1):136. https://doi.org/10.3390/biology12010136
Chicago/Turabian StyleBateman, Richard M., and Paula J. Rudall. 2023. "Morphological Continua Make Poor Species: Genus-Wide Morphometric Survey of the European Bee Orchids (Ophrys L.)" Biology 12, no. 1: 136. https://doi.org/10.3390/biology12010136
APA StyleBateman, R. M., & Rudall, P. J. (2023). Morphological Continua Make Poor Species: Genus-Wide Morphometric Survey of the European Bee Orchids (Ophrys L.). Biology, 12(1), 136. https://doi.org/10.3390/biology12010136