
Immunogenetic Predisposition to SARS-CoV-2 Infection

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cohort
2.2. DNA Preparation
2.3. Next-Generation Sequencing HLA Typing
2.4. Next-Generation Sequencing Blood Group Typing (BG) and Platelet Antigen Genes (HPA)
2.4.1. Regions of Interest
2.4.2. Primer Design
2.4.3. Next-Generation Library Preparation
2.4.4. Evaluation Criteria
2.5. Analysis of the NGS Data
2.5.1. Allele Assignment and Phenotype Determination (BG and HPA)
Referenced Alleles
Non-Referenced Alleles
2.6. Statistical Analysis
3. Results
3.1. Review of Current Results on COVID-19 Infections
3.2. Analysis of the HLA Alleles
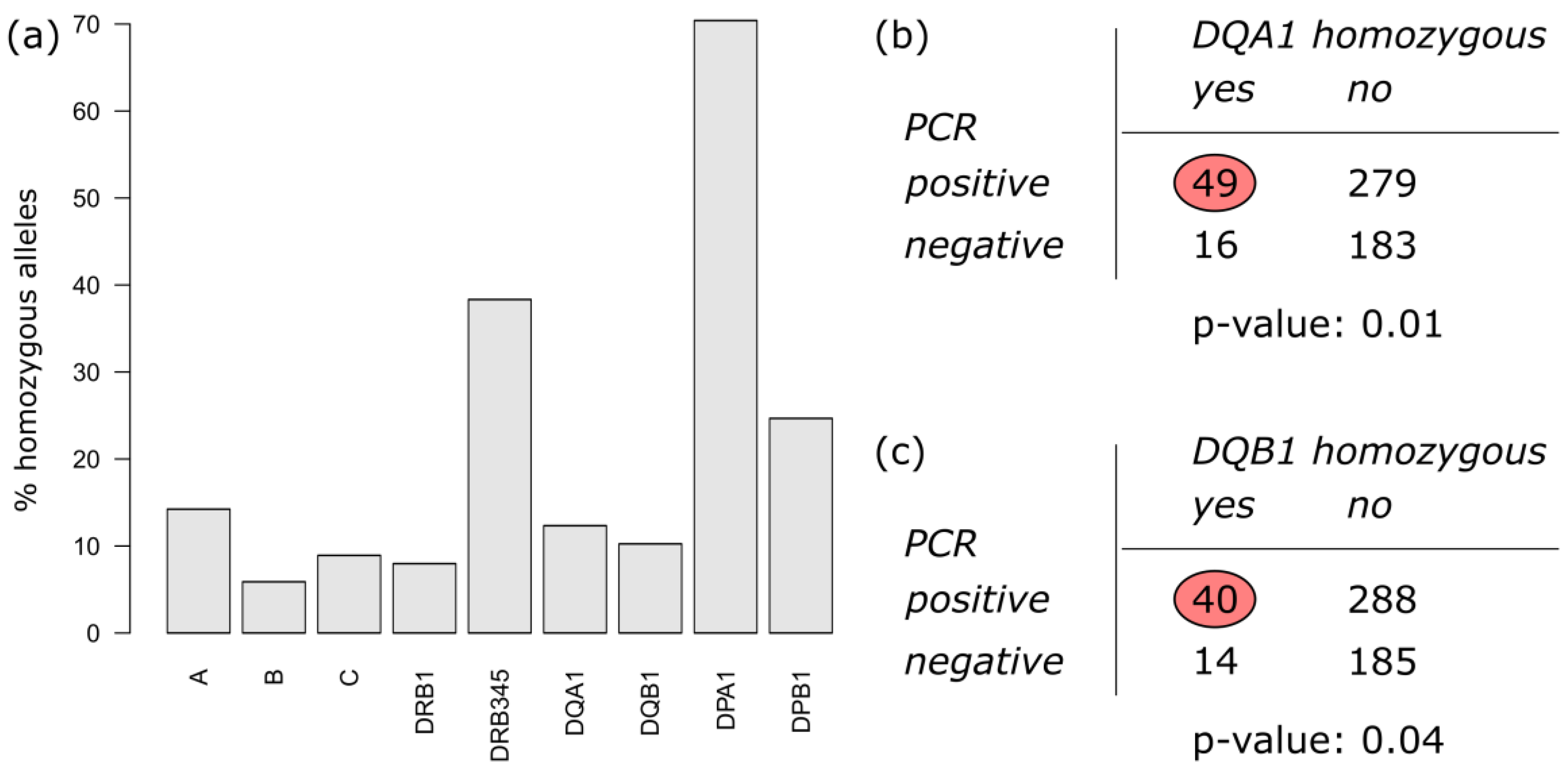
3.2.1. Homozygosity of HLA Alleles
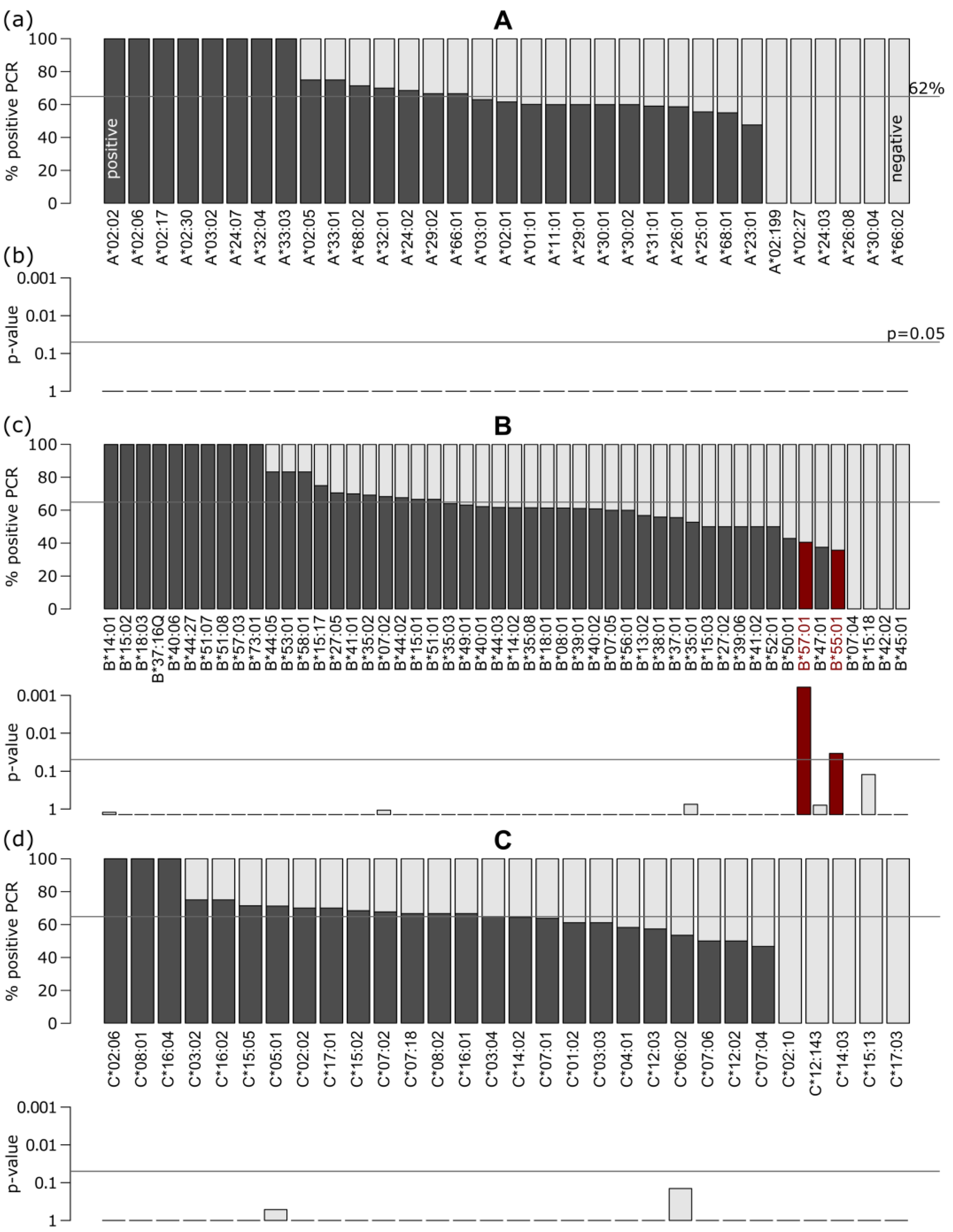
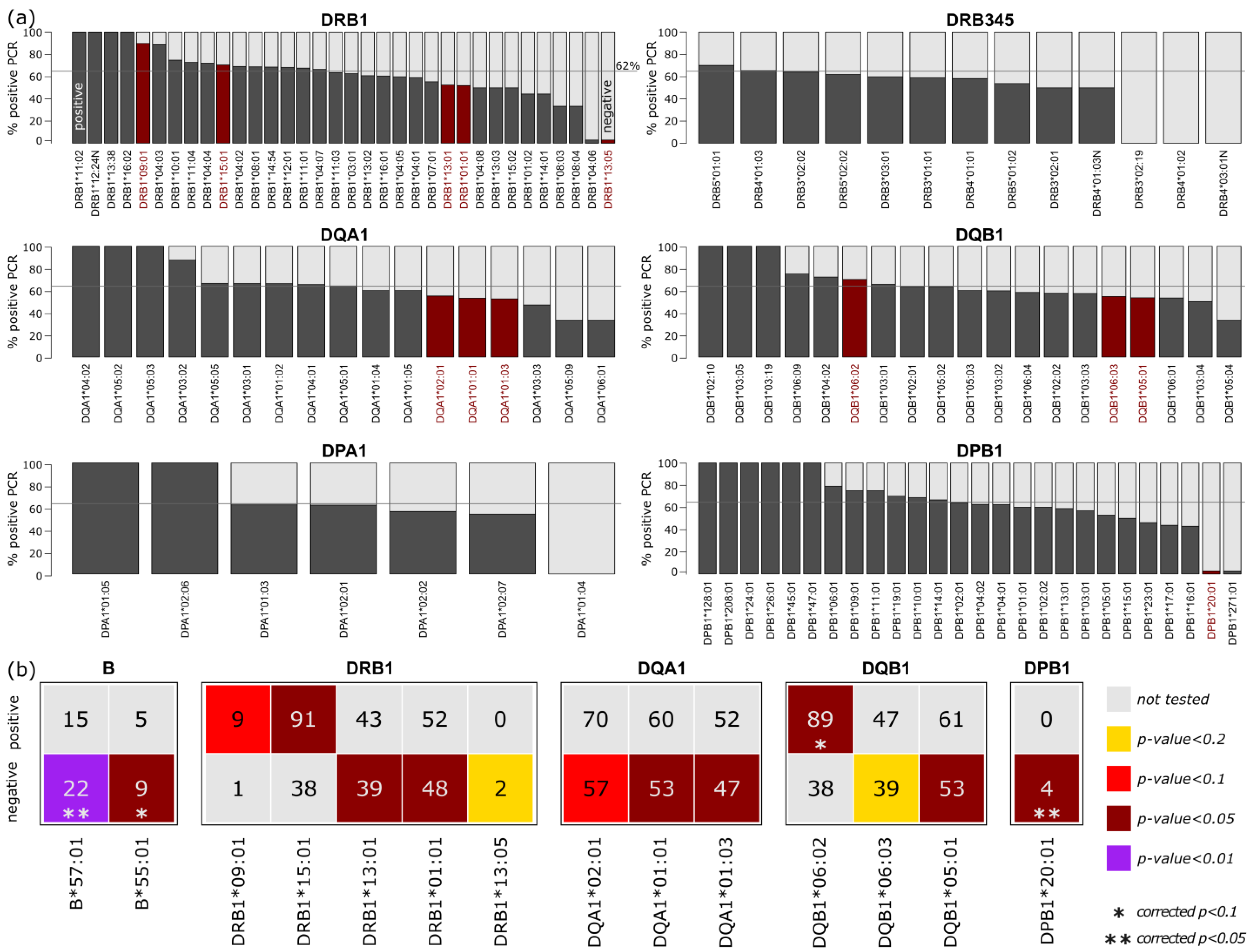
3.2.2. Association of HLA Alleles in the Study
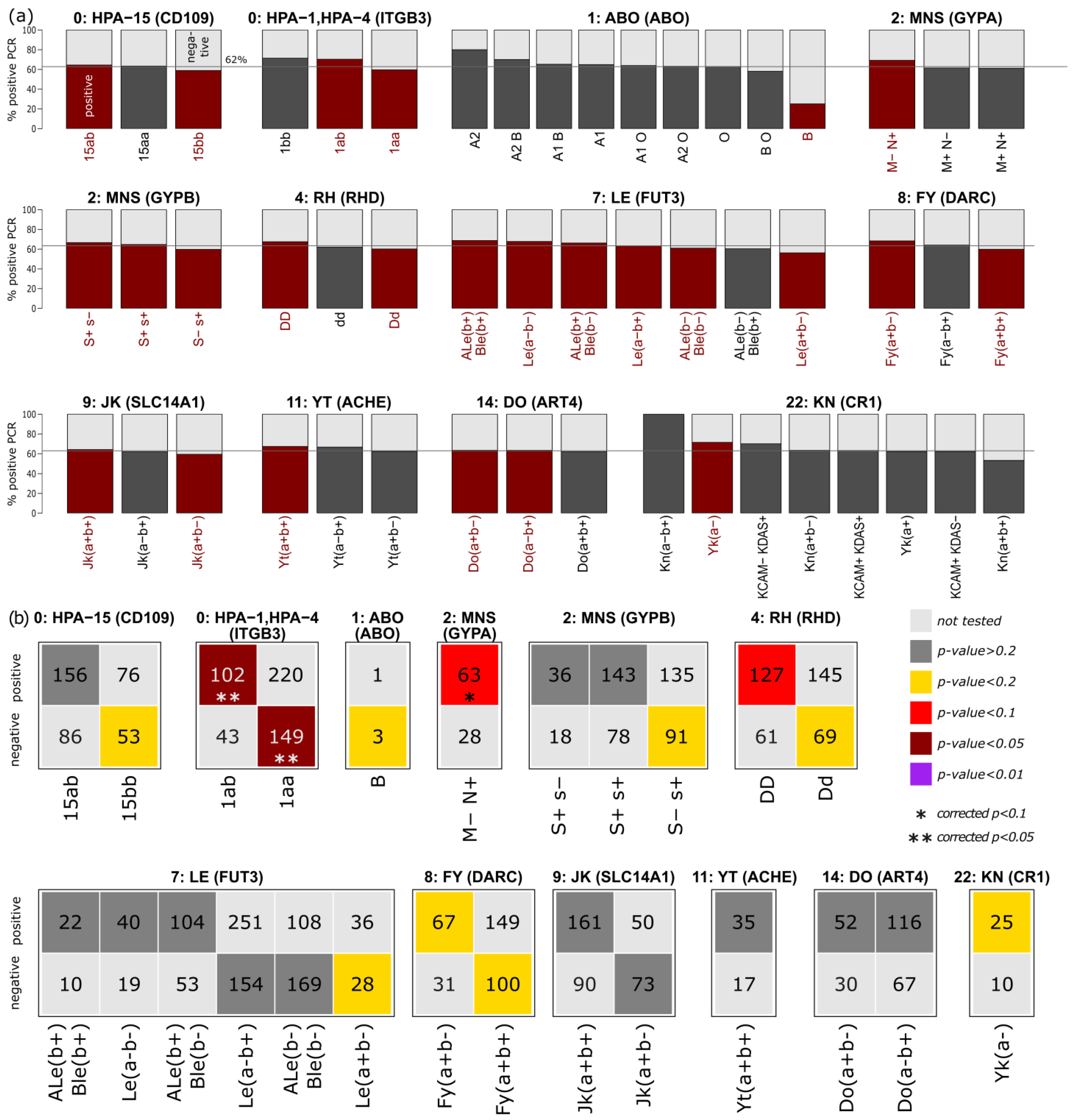
3.3. Enrichment of Blood Group and Platelet Antigens in the Two Subgroups
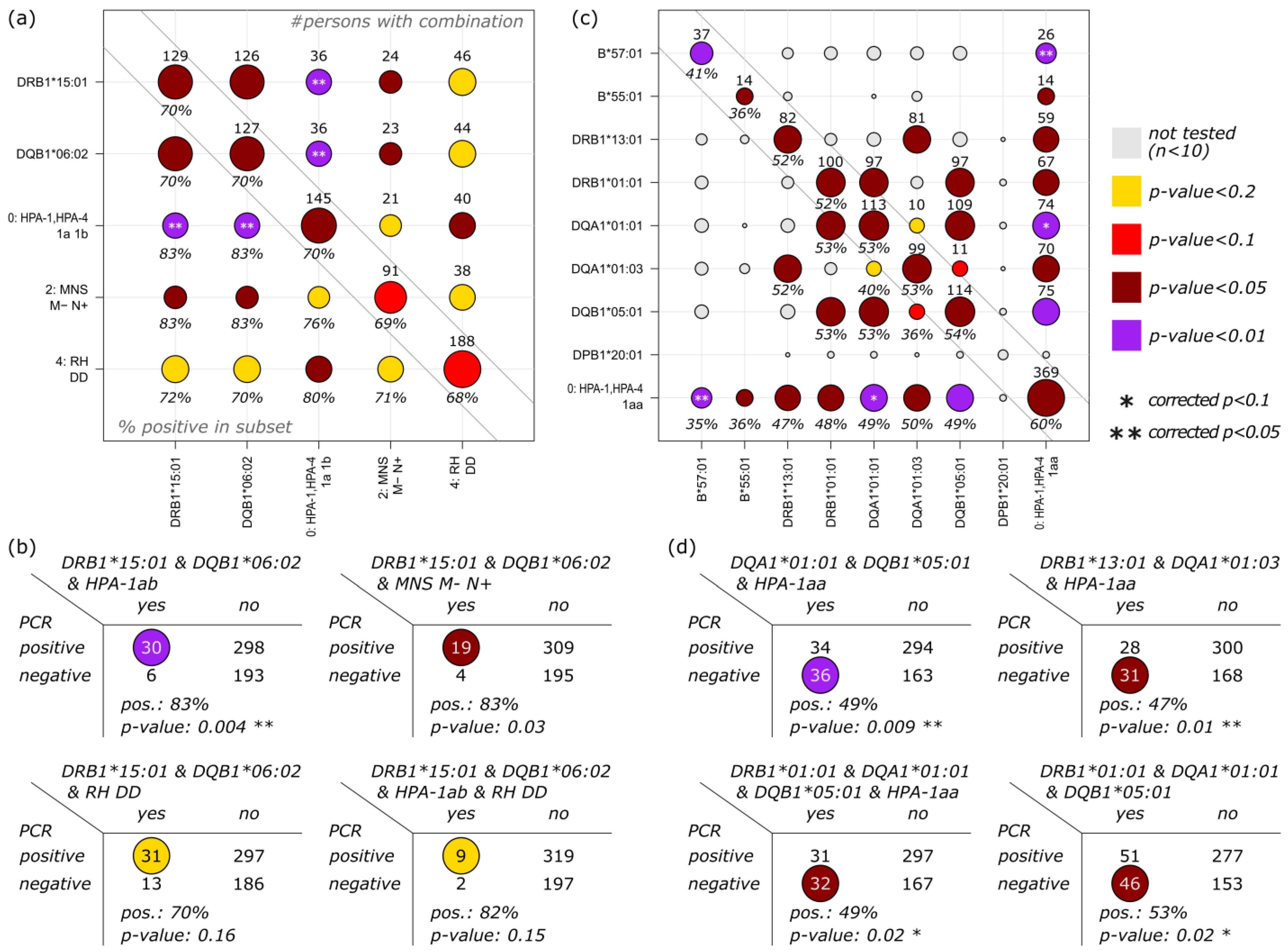
3.4. Correlation of HLA, Blood Group and Platelet Antigens
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Available online: www.corona-in-zahlen.deBundeslaender (accessed on 15 November 2022).
- Finstad, J.; Good, R.A. The evolution of the immune response. 3. Immunologic responses in the lamprey. J. Exp. Med. 1964, 120, 1151–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: www.who.int/emergencies/disease-outbreak-news (accessed on 15 November 2022).
- Available online: www.nobelprize.org/prizes/medicine/1980/summary (accessed on 15 November 2022).
- Burian, A.; Wang, K.L.; Finton, K.A.K.; Lee, N.; Ishitani, A.; Strong, R.K.; Geraghty, D.E. HLA-F and MHC-I Open Conformers Bind Natural Killer Cell Ig-Like Receptor KIR3DS1. PLoS ONE 2016, 11, e0163297. [Google Scholar] [CrossRef] [PubMed]
- Available online: www.hla.alleles.org (accessed on 15 November 2022).
- Crux, N.B.; Elahi, S. Human Leukocyte Antigen (HLA) and Immune Regulation: How Do Classical and Non-Classical HLA Alleles Modulate Immune Response to Human Immunodeficiency Virus and Hepatitis C Virus Infections? Front. Immunol. 2017, 8, 832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, J.M.; Jamieson, S.E.; Burgner, D. HLA and Infectious Diseases. Clin. Microbiol. Rev. 2009, 22, 370–385. [Google Scholar] [CrossRef] [Green Version]
- Maira, D.; Vansan, A.; Maria, A.; Laguila, V.; Jeane, E.; de Souz, C.A. HLA and Infectious Diseases. In HLA and Associated Important Diseases; Xi, Y., Ed.; Intech Open: London, UK, 2014. [Google Scholar] [CrossRef]
- de Wit, J.; Borghans, J.A.M.; Kesmir, C.; van Baarle, D. Editorial: Role of HLA and KIR in Viral Infections. Front. Immunol. 2016, 7, 286. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Tseng, H.-K.; Trejaut, J.A.; Lee, H.-L.; Loo, J.-H.; Chun-Hsiung, H.; Chen, P.-J.; Su, Y.-W.; Lim, K.H.; Tsai, Z.-U.; et al. Association of HLA class I with severe acute respiratory syndrome coronavirus infection. BMC Med. Genet. 2003, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.; David, J.K.; Maden, S.K.; Wood, M.A.; Weeder, B.R.; Nellore, A.; Thompson, R.F. Human Leukocyte Antigen Susceptibility Map for Severe Acute Respiratory Syndrome Coronavirus 2. J. Virol. 2020, 94, e00510-20. [Google Scholar] [CrossRef] [Green Version]
- Cooling, L. Blood Groups in Infection and Host Susceptibility. Clin. Microbiol. Rev. 2015, 28, 801–870. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Yang, Y.; Huang, H.; Li, D.; Gu, D.; Lu, X.; Zhang, Z.; Liu, L.; Liu, T.; Liu, Y.; et al. Relationship between the ABO Blood Group and the COVID-19 Susceptibility. Clin. Infect. Dis. 2021, 73, 328–331. [Google Scholar] [CrossRef]
- Asselta, R.; Paraboschi, E.M.; Mantovani, A.; Duga, S. ACE2 and TMPRSS2 variants and expression as candidates to sex and country differences in COVID-19 severity in Italy. Aging 2020, 12, 10087–10098. [Google Scholar] [CrossRef]
- Alnaqbi, H.; Tay, G.K.; Jelinek, H.F.; Francis, A.; Alefishat, E.; Chehadeh, S.E.H.; Saeed, A.T.; Hussein, M.; Salameh, L.; Mahboub, B.H.; et al. HLA repertoire of 115 UAE nationals infected with SARS-CoV-2. Hum. Immunol. 2021, 83, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Severe Covid-19 GWAS Group. Genomewide Association Study of Severe Covid-19 with Respiratory Failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M.; Cruciani, M.; Mengoli, C.; Marano, G.; Candura, F.; Lopez, N.; Pati, I.; Pupella, S.; De Angelis, V. ABO blood group and COVID-19: An updated systematic literature review and meta-analysis. Blood Transfus. = Trasfus. del sangue 2021, 19, 317–326. [Google Scholar] [CrossRef]
- Iturrieta-Zuazo, I.; Rita, C.G.; García-Soidán, A.; Pintos-Fonseca, A.D.M.; Alonso-Alarcón, N.; Pariente-Rodríguez, R.; Tejeda-Velarde, A.; Serrano-Villar, S.; Castañer-Alabau, J.L.; Nieto-Gañán, I. Possible role of HLA class-I genotype in SARS-CoV-2 infection and progression: A pilot study in a cohort of Covid-19 Spanish patients. Clin. Immunol. 2020, 219, 108572. [Google Scholar] [CrossRef] [PubMed]
- Langton, D.J.; Bourke, S.C.; Lie, B.A.; Reiff, G.; Natu, S.; Darlay, R.; Burn, J.; Echevarria, C. The influence of HLA genotype on the severity of COVID-19 infection. HLA 2021, 98, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Littera, R.; Campagna, M.; Deidda, S.; Angioni, G.; Cipri, S.; Melis, M.; Firinu, D.; Santus, S.; Lai, A.; Porcella, R.; et al. Human Leukocyte Antigen Complex and Other Immunogenetic and Clinical Factors Influence Susceptibility or Protection to SARS-CoV-2 Infection and Severity of the Disease Course. The Sardinian Experience. Front. Immunol. 2020, 11, 605688. [Google Scholar] [CrossRef]
- Shkurnikov, M.; Nersisyan, S.; Jankevic, T.; Galatenko, A.; Gordeev, I.; Vechorko, V.; Tonevitsky, A. Association of HLA Class I Genotypes With Severity of Coronavirus Disease-19. Front. Immunol. 2021, 12, 641900. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, W.; Zhang, J.; He, J.; Zhu, F. Distribution of HLA allele frequencies in 82 Chinese individuals with coronavirus disease-2019 (COVID-19). HLA 2020, 96, 194–196. [Google Scholar] [CrossRef]
- Franchini, M.; Liumbruno, G.M.; Pezzo, M. COVID-19 vaccine-associated immune thrombosis and thrombocytopenia (VITT): Diagnostic and therapeutic recommendations for a new syndrome. Eur. J. Haematol. 2021, 107, 173–180. [Google Scholar] [CrossRef]
- Matzhold, E.M.; Berghold, A.; Bemelmans, M.K.B.; Banfi, C.; Stelzl, E.; Kessler, H.H.; Steinmetz, I.; Krause, R.; Wurzer, H.; Schlenke, P.; et al. Lewis and ABO histo-blood types and the secretor status of patients hospitalized with COVID-19 implicate a role for ABO antibodies in susceptibility to infection with SARS-CoV-2. Transfusion 2021, 61, 2736–2745. [Google Scholar] [CrossRef]
- McCullough, J. RBCs as targets of infection. In Hematology; American Society of Hematology, Education Program: Washington, DC, USA, 2014; Volume 2014, pp. 404–409. [Google Scholar] [CrossRef] [Green Version]
- Rios, M.; Bianca, C. The role of blood group antigens in infectious diseases. Semin. Hematol. 2000, 37, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguiar, V.R.C.; Augusto, D.G.; Castelli, E.C.; Hollenbach, J.A.; Meyer, D.; Nunes, K.; Petzl-Erler, M.L. An immunogenetic view of COVID-19. Genet. Mol. Biol. 2021, 44 (Suppl. S1), e20210036. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A. A Sequence Homology and Bioinformatic Approach Can Predict Candidate Targets for Immune Responses to SARS-CoV-2. Cell Host Microbe 2020, 27, 671–680.e2. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Joshi, S.; Joshi, M.; Degani, M.S. Tackling SARS-CoV-2: Proposed targets and repurposed drugs. Future Med. Chem. 2020, 12, 1579–1601. [Google Scholar] [CrossRef]
- Kiyotani, K.; Toyoshima, Y.; Nemoto, K.; Nakamura, Y. Bioinformatic prediction of potential T cell epitopes for SARS-Cov-2. J. Hum. Genet. 2020, 65, 569–575. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mazumder, P.; Joshi, M.; Joshi, C.; Dalvi, S.V.; Kumar, M. Biomedical application, drug delivery and metabolic pathway of antiviral nanotherapeutics for combating viral pandemic: A review. Environ. Res. 2020, 191, 110119. [Google Scholar] [CrossRef]
- Vashi, A.P.; Coiado, O.C. The future of COVID-19: A vaccine review. J. Infect. Public Heal. 2021, 14, 1461–1465. [Google Scholar] [CrossRef]
- Yarmarkovich, M.; Warrington, J.M.; Farrel, A.; Maris, J.M. Identification of SARS-CoV-2 Vaccine Epitopes Predicted to Induce Long-Term Population-Scale Immunity. Cell Rep. Med. 2020, 1, 100036. [Google Scholar] [CrossRef]
- De Marco, R.; Faria, T.C.; Mine, K.L.; Cristelli, M.; Medina-Pestana, J.O.; Tedesco-Silva, H.; Gerbase-DeLima, M. HLA-A homozygosis is associated with susceptibility to COVID-19. HLA 2021, 98, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Aguiar, V.R.C.; Bitarello, B.D.; Brandt, D.Y.C.; Nunes, K. A genomic perspective on HLA evolution. Immunogenetics 2018, 70, 5–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, J.; McLaren, P.J.; Chaturvedi, N.; Carrington, M.; Fellay, J.; Lenz, T.L. HIV peptidome-wide association study reveals patient-specific epitope repertoires associated with HIV control. Proc. Natl. Acad. Sci. USA 2019, 116, 944–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrington, M.; Nelson, G.W.; Martin, M.P.; Kissner, T.; Vlahov, D.; Goedert, J.J.; Kaslow, R.; Buchbinder, S.; Hoots, K.; O’Brien, S.J. HLA and HIV-1: Heterozygote Advantage and B*35-Cw*04 Disadvantage. Science 1999, 283, 1748–1752. [Google Scholar] [CrossRef]
- Carrington, M.; O’Brien, S.J. The Influence of HLA Genotype on AIDS. Annu. Rev. Med. 2003, 54, 535–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaakkola, E.; Herzberg, I.; Laiho, K.; Barnardo, M.C.N.M.; Pointon, J.J.; Kauppi, M.; Kaarela, K.; Tuomilehto-Wolf, E.; Tuomilehto, J.; Wordsworth, B.P.; et al. Finnish HLA studies confirm the increased risk conferred by HLA-B27 homozygosity in ankylosing spondylitis. Ann. Rheum. Dis. 2006, 65, 775–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marmor, M.; Sheppard, H.W.; Donnell, D.; Bozeman, S.; Celum, C.; Buchbinder, S.; Koblin, B.; Seage, G.R. Homozygous and heterozygous CCR5-Delta32 genotypes are associated with resistance to HIV infection. Am. J. Ther. (1999) 2001, 27, 472–481. [Google Scholar] [CrossRef]
- Rohlfing, A.-K.; Rath, D.; Geisler, T.; Gawaz, M. Platelets and COVID-19. Hamostaseologie 2021, 41, 379–385. [Google Scholar] [CrossRef]
- Ruiz-Ortiz, E.; Montraveta, M.; Cabré, E.; Herrero-Mata, M.J.; Pujol-Borrell, R.; Palou, E.; Faner, R. HLA-DQ2/DQ8 and HLA-DQB1*02 homozygosity typing by real-time polymerase chain reaction for the assessment of celiac disease genetic risk: Evaluation of a Spanish celiac population. Tissue Antigens 2014, 84, 545–553. [Google Scholar] [CrossRef]
- Sauermann, U.; Stahl-Hennig, C.; Stolte, N.; Mühl, T.; Krawczak, M.; Spring, M.; Fuchs, D.; Kaup, F.-J.; Hunsmann, G.; Sopper, S. Homozygosity for a conserved Mhc class II DQ-DRB haplotype is associated with rapid disease progression in simian immunodeficiency virus-infected macaques: Results from a prospective study. J. Infect. Dis. 2000, 182, 716–724. [Google Scholar] [CrossRef]
- Fischer, J.C.; Balz, V.; Jazmati, D.; Bölke, E.; Freise, N.F.; Keitel, V.; Feldt, T.; Jensen, B.-E.O.; Bode, J.; Lüdde, T.; et al. Prognostic markers for the clinical course in the blood of patients with SARS-CoV-2 infection. Eur. J. Med. Res. 2022, 27, 255. [Google Scholar] [CrossRef] [PubMed]
- Weiss, L.J.; Drayss, M.; Manukjan, G.; Zeitlhöefler, M.J.; Kleiss, J.; Weigel, M.L.; Herrmann, J.; Mott, K.; Beck, S.; Burkard, P.; et al. Uncoupling of platelet granule release and integrin activation suggests GPIIb/IIIa as therapeutic target in COVID-19. Blood Adv. 2022; in press. [Google Scholar] [CrossRef]
- Metcalfe, P.; Watkins, N.A.; Ouwehand, W.H.; Kaplan, C.; Newman, P.; Kekomaki, R.; De Haas, M.; Aster, R.; Shibata, Y.; Smith, J.; et al. Nomenclature of human platelet antigens. Vox Sang. 2003, 85, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, K.V.; Bray, P.F. Molecular mechanisms of prothrombotic risk due to genetic variations in platelet genes: Enhanced outside-in signaling through the Pro33 variant of integrin β3. Exp. Biol. Med. 2006, 231, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The cytokine storm in COVID-19: An overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.; Chitty, D.; Lee, M.; Hakim, N.; Lakhani, S.; Rajdev, L.; Zhu, X.; Saif, M.W. COVID-19 Clinical Research. J. Cell. Signal. 2020, 1, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wu, X.; Zheng, X.; Luo, S.; Xu, S.; Weng, J. Targeting inflammation and cytokine storm in COVID-19. Pharmacol. Res. 2020, 159, 105051. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Martin, J.C. Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 355–362. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ISBT System No. | System Name | System Symbol | Gene(s) | Reference Sequence (Transcript) | Reference Sequence (Genomic) |
|---|---|---|---|---|---|
| 001 | ABO | ABO | ABO | NM_020469 | NG_006669.2 |
| 002 | MNS | MNS | GYPA | NM_002099.6 | NG_007470.3 |
| GYPB | NM_002100.4 | NG_007483.2 | |||
| GYPE | NM_002102.3 | NG_009173.1 | |||
| 003 | P1PK | P1PK | A4GALT | NM_017436.4 | NG_007495.1 |
| 004 | Rhesus | RH | RHCE | NM_020485.4 | NG_009208.3 |
| RHD | NM_016124.3 | NG_007494.1 | |||
| 005 | Lutheran | LU | B-CAM | NM_005581.4 | NG_007480.1 |
| 006 | Kell | KEL | KEL | NM_000420.2 | NG_007492.3 |
| 007 | Lewis | LE | FUT3 | NM_000149.4 | NG_007482.2 |
| 008 | Duffy | FY | DARC | NM_002036.3 | NG_011626.2 |
| 009 | Kidd | JK | SLC14A1 | NM_015865.6 | NG_011775.4 |
| 010 | Diego | DI | SLC4A1 | NM_000342.3 | NG_007498.1 |
| 011 | Cartwright | YT | ACHE | NM_015831.2 | NG_007474.2 |
| 014 | Dombrock | DO | ART4 | NM_021071.2 | NG_007477.2 |
| 015 | Colton | CO | AQP1 | NM_198098.2 | NG_007475.2 |
| 018 | H | H | FUT1 | NM_000148.3 | NG_007510.1 |
| FUT2 | NM_000511.5 | NG_007511.1 | |||
| 020 | Gerbich | GE | GYPC | NM_002101.4 | NG_007479.1 |
| 021 | Cromer | CROM | CD55 | NM_000574.3 | NG_007465.1 |
| 022 | Knops | KN | CR1 | NM_000573.3 | NG_007481.1 |
| 023 | Indian | IN | CD44 | NM_0010011391.1 | NG_008937.1 |
| 032 | Junior | JR | ABCG2 | NM_004827.2 | NG_032067.2 |
| 033 | Langereis | LAN | ABCB6 | NM_005689.2 | NG_032110.1 |
| 034 | Vel | VEL | SMIM1 | NM_001163724.3 | NG_033869.1 |
| 036 | Augustine | AUG | SLC29A1 | NM_004955.2 | NG_042893.1 |
| HPA-1/HPA-4 | ITGB3 | NM_000212.2 | NG_008332.2 | ||
| HPA-2 | GP1BA | NM_000173.7 | NG_008767.2 | ||
| HPA-3 | ITGA2B | NM_000419.5 | NG_008331.1 | ||
| HPA-5 | ITGA2 | NM_002203.4 | NG_008330.2 | ||
| HPA-15 | CD109 | NM_133493.3 | NG_033971.1 |
| Population | Methods | Cohort | HLA Association Results | Reference |
|---|---|---|---|---|
| Russian | NGS, BAP | 111 deceased patients with COVID-19 | A*01:01 homozygosity high risk A*02:01, A*03:01 protective | [22] |
| British | NGS | 147 European COVID-19 infected patients with variable outcomes (49 severe, 69 asymptomatic hospital staff positive for COVID-19) | DRB1*04:01 protective DQA1*01:01-DQB1*05:01-DRB1*01:01 higher frequency in asymptomatic group | [20] |
| Italian/Spanish | GWAS | 835 (Italy) and 775 (Spain) COVID-19 infected patients from 7 hospitals | no significant HLA association chromosome 3p21.31, the peak association signal covered a cluster of six genes (SLC6A20, LZTFL1, CCR9, FYCO1, CXCR6, and XCR1) higher risk blood group A protective blood group O | [17] |
| Sardinian | SSP-PCR | 182 COVID-19 infected patients asymptomatic, pauci-symptomatic, hospitalized | A*02:05, B*58:01, C*07:01, DRB1*03:01 protective A*30:02, B*14:02, C*08:02, DRB1*08:01 higher risk | [21] |
| Spanish | SSO, BAP | 45 COVID-19 patients with mild, moderate, and severe infection (only HLA class I) | SARS-CoV-2 binding capacity of different HLA alleles A2, C1 subtypes; homozygosity of HLA alleles | [19] |
| Chinese | NGS | 82 COVID-19 infected patients | C*08:01G, B*15:27, B*40:06, DRB1*04:06, and DPB1*36:01 higher in COVID-19 patients DRB1*12:02, DPB1*04:01 lower in COVID-19 patients | [23] |
| Multi ethnic | BAP | A*02:02, B*15:03, C*12:03 highest predicted capacity for SARS-CoV-2 epitope presentation A*25:01, B*46:01, C*01:02 lowest predicted capacity for SARS-CoV-2 epitope presentation | [12] | |
| United Arab Emirates Abu Dhabi | NGS | 115 patients with mild, moderate, and severe SARS-CoV-2 infection age mean mild: 34 ± 14 age mean moderate/severe: 58 ± 15 | A*03:01, B44, DRB1*15:01 heterozygosity significant association with severity A*26:01, B*51:01 negative association (protectiv) | [16] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehmann, C.; Loeffler-Wirth, H.; Balz, V.; Enczmann, J.; Landgraf, R.; Lakowa, N.; Gruenewald, T.; Fischer, J.C.; Doxiadis, I. Immunogenetic Predisposition to SARS-CoV-2 Infection. Biology 2023, 12, 37. https://doi.org/10.3390/biology12010037
Lehmann C, Loeffler-Wirth H, Balz V, Enczmann J, Landgraf R, Lakowa N, Gruenewald T, Fischer JC, Doxiadis I. Immunogenetic Predisposition to SARS-CoV-2 Infection. Biology. 2023; 12(1):37. https://doi.org/10.3390/biology12010037
Chicago/Turabian StyleLehmann, Claudia, Henry Loeffler-Wirth, Vera Balz, Juergen Enczmann, Ramona Landgraf, Nicole Lakowa, Thomas Gruenewald, Johannes C. Fischer, and Ilias Doxiadis. 2023. "Immunogenetic Predisposition to SARS-CoV-2 Infection" Biology 12, no. 1: 37. https://doi.org/10.3390/biology12010037
APA StyleLehmann, C., Loeffler-Wirth, H., Balz, V., Enczmann, J., Landgraf, R., Lakowa, N., Gruenewald, T., Fischer, J. C., & Doxiadis, I. (2023). Immunogenetic Predisposition to SARS-CoV-2 Infection. Biology, 12(1), 37. https://doi.org/10.3390/biology12010037