
Neurofeedback and Affect Regulation Circuitry in Depressed and Healthy Adolescents
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
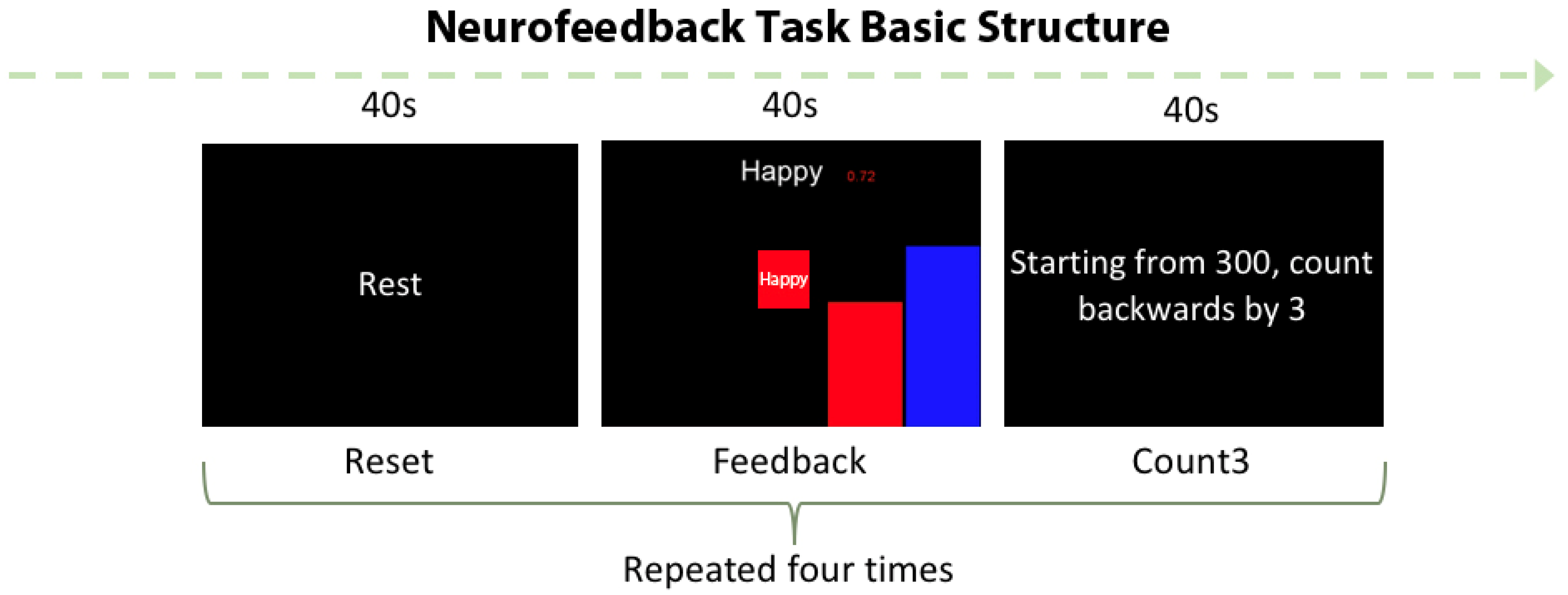
2.1. “Happy” Word Neurofeedback Task
2.2. Online Analysis: Real-Time Feedback from the Brain
2.3. Off-Line Analysis: Whole Brain Analysis
2.4. Off-Line Analysis: AMYHIPPO and ACC Activity ROIs Analyses
3. Results
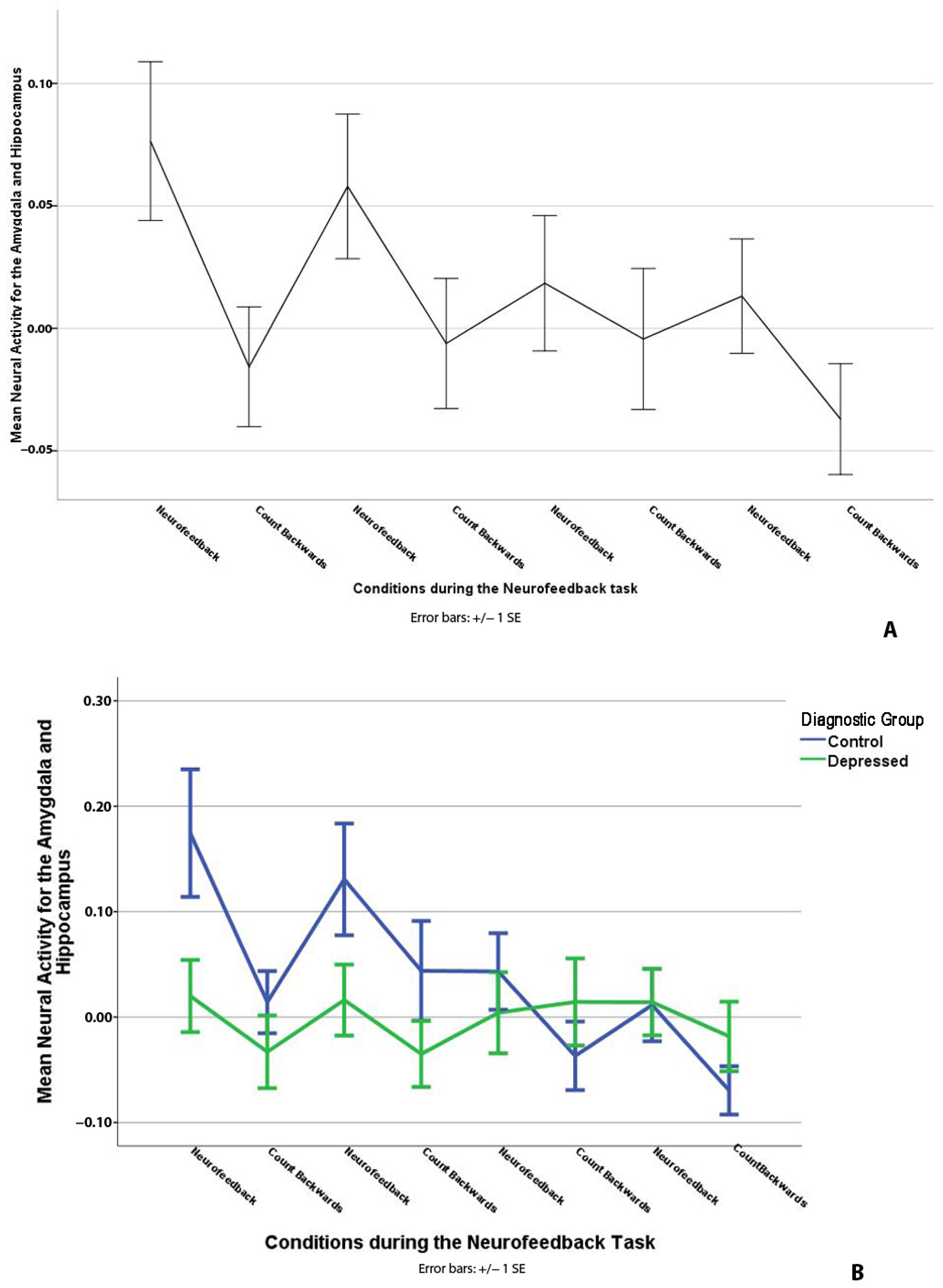
3.1. “Happy” Word Neurofeedback Task: AMYHIPPO ROI Analysis
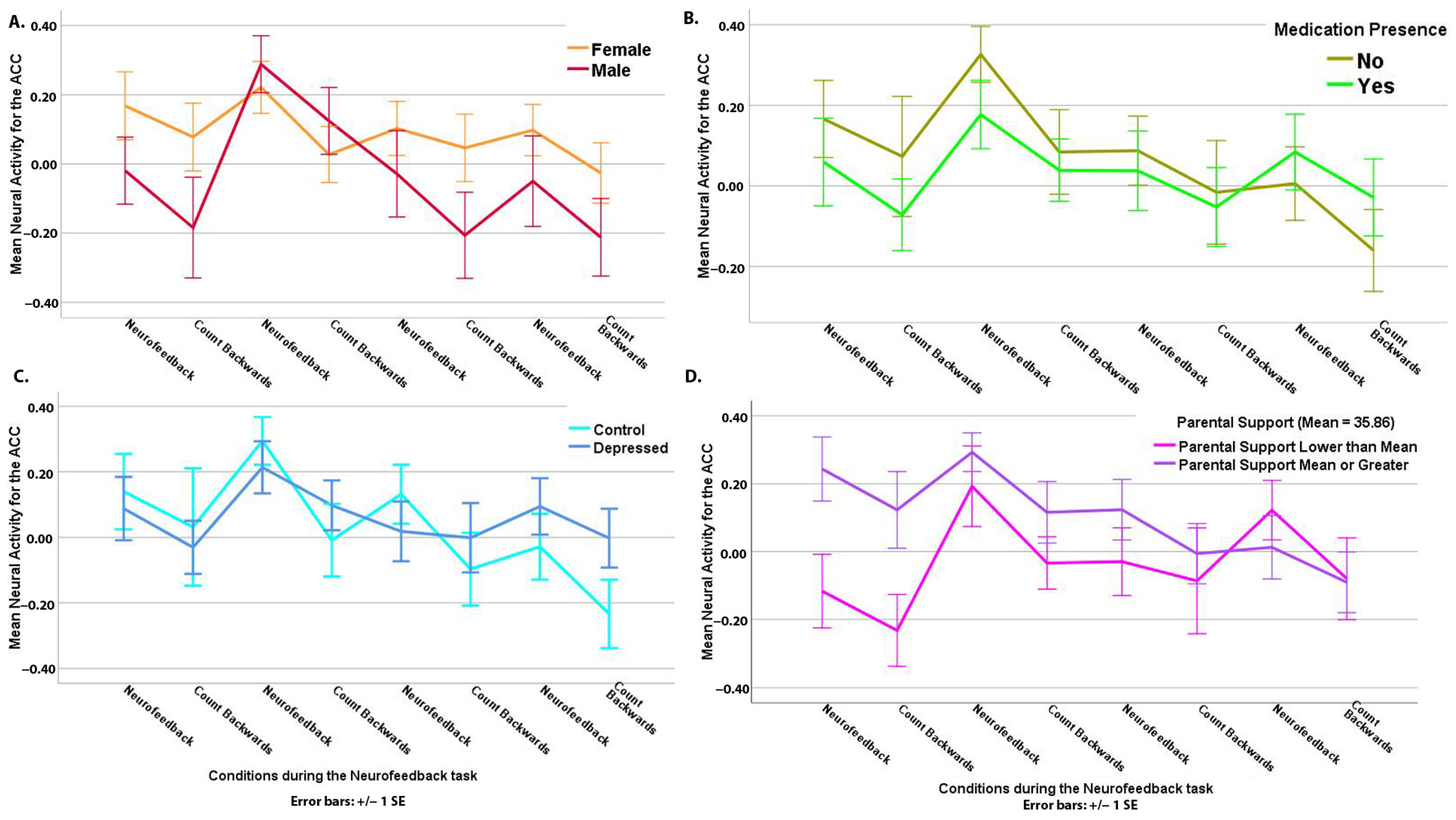
3.2. “Happy” Word Neurofeedback Task: ACC ROI Analysis
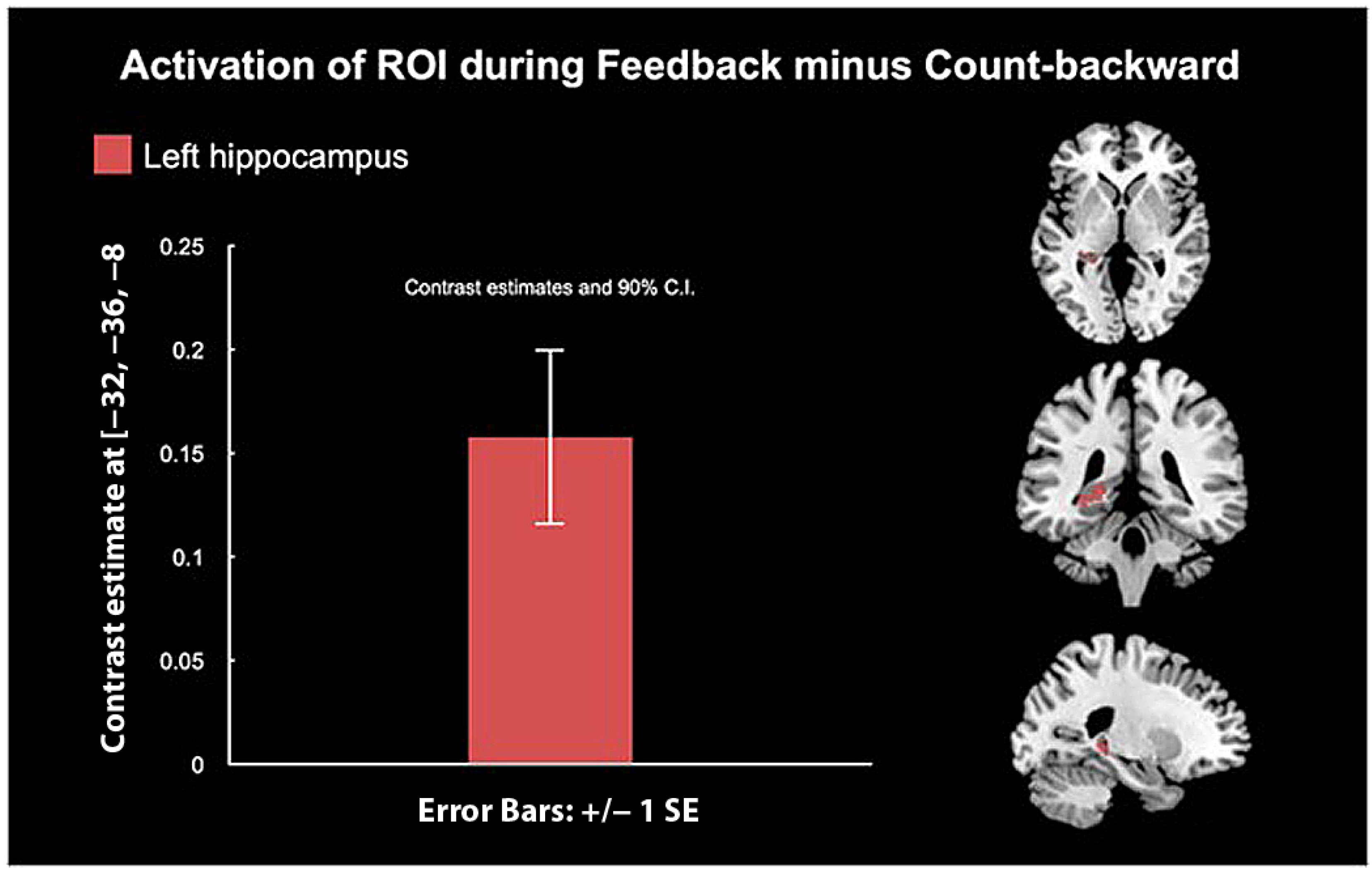
3.3. “Happy” Word Neurofeedback Task: Whole-Brain Analysis
4. Discussion
4.1. Amygdala and Hippocampus (AMYHIPPO) and Anterior Cingulate Cortex (ACC) Activity during Neurofeedback
4.2. Whole-Brain Analysis
4.3. Diagnosis and Brain Activity during Neurofeedback
5. Limitations and Future Studies
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Avenevoli, S.; Swendsen, J.; He, J.P.; Burstein, M.; Merikangas, K.R. Major depression in the national comorbidity survey-adolescent supplement: Prevalence, correlates, and treatment. J. Am. Acad. Child. Adolesc. Psychiatry 2015, 54, 37–44.e32. [Google Scholar] [CrossRef] [PubMed]
- Halfin, A. Depression: The benefits of early and appropriate treatment. Am. J. Manag. Care 2007, 13, S92–S97. [Google Scholar] [PubMed]
- Weisz, J.R.; Kuppens, S.; Ng, M.Y.; Eckshtain, D.; Ugueto, A.M.; Vaughn-Coaxum, R.; Jensen-Doss, A.; Hawley, K.M.; Krumholz Marchette, L.S.; Chu, B.C.; et al. What five decades of research tells us about the effects of youth psychological therapy: A multilevel meta-analysis and implications for science and practice. Am. Psychol. 2017, 72, 79–117. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Lifshitz, M.; Zalsman, G.; Giner, L.; Oquendo, M.A. Can we really prevent suicide? Curr. Psychiatry Rep. 2012, 14, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Rush, A.J.; Trivedi, M.H.; Wisniewski, S.R.; Nierenberg, A.A.; Stewart, J.W.; Warden, D.; Niederehe, G.; Thase, M.E.; Lavori, P.W.; Lebowitz, B.D.; et al. Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: A STAR*D report. Am. J. Psychiatry 2006, 163, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, E.; Gaynes, B.; Burda, B.U.; Williams, C.; Whitlock, E.P. U.S. Preventive Services Task Force Evidence Syntheses, formerly Systematic Evidence Reviews. In Screening for Suicide Risk in Primary Care: A Systematic Evidence Review for the U.S. Preventive Services Task Force; Agency for Healthcare Research and Quality (US): Rockville, MD, USA, 2013. [Google Scholar]
- Maalouf, F.T.; Brent, D.A. Child and adolescent depression intervention overview: What works, for whom and how well? Child Adolesc. Psychiatr. Clin. N. Am. 2012, 21, 299–312. [Google Scholar] [CrossRef]
- Hetrick, S.E.; Cox, G.R.; Witt, K.G.; Bir, J.J.; Merry, S.N. Cognitive behavioural therapy (CBT), third-wave CBT and interpersonal therapy (IPT) based interventions for preventing depression in children and adolescents. Cochrane Database Syst. Rev. 2016, 2016, CD003380. [Google Scholar] [CrossRef]
- De Silva, S.; Parker, A.; Purcell, R.; Callahan, P.; Liu, P.; Hetrick, S. Mapping the evidence of prevention and intervention studies for suicidal and self-harming behaviors in young people. Crisis 2013, 34, 223–232. [Google Scholar] [CrossRef]
- Cha, C.B.; Franz, P.J.; Guzmán, E.M.; Glenn, C.R.; Kleiman, E.M.; Nock, M.K. Annual Research Review: Suicide among youth—Epidemiology, (potential) etiology, and treatment. J. Child. Psychol. Psychiatry 2018, 59, 460–482. [Google Scholar] [CrossRef]
- Witte, T.K.; Timmons, K.A.; Fink, E.; Smith, A.R.; Joiner, T.E. Do major depressive disorder and dysthymic disorder confer differential risk for suicide? J. Affect. Disord. 2009, 115, 69–78. [Google Scholar] [CrossRef]
- Wang, P.S.; Simon, G.; Kessler, R.C. The economic burden of depression and the cost-effectiveness of treatment. Int. J. Methods Psychiatr. Res. 2003, 12, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Pezawas, L.; Wittchen, H.U.; Pfister, H.; Angst, J.; Lieb, R.; Kasper, S. Recurrent brief depressive disorder reinvestigated: A community sample of adolescents and young adults. Psychol. Med. 2003, 33, 407–418. [Google Scholar] [CrossRef]
- Jonsson, U.; Bohman, H.; von Knorring, L.; Olsson, G.; Paaren, A.; von Knorring, A.L. Mental health outcome of long-term and episodic adolescent depression: 15-year follow-up of a community sample. J. Affect. Disord. 2011, 130, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Shi, D.; Gao, Y.; Xu, J. Functional assessment of prefrontal lobes in patients with major depression disorder using a dual-mode technique of 3D-arterial spin labeling and (18)F-fluorodeoxyglucose positron emission tomography/computed tomography. Exp. Ther. Med. 2017, 14, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Schecklmann, M.; Dresler, T.; Beck, S.; Jay, J.T.; Febres, R.; Haeusler, J.; Jarczok, T.A.; Reif, A.; Plichta, M.M.; Ehlis, A.C.; et al. Reduced prefrontal oxygenation during object and spatial visual working memory in unpolar and bipolar depression. Psychiatry Res. 2011, 194, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Werner-Seidler, A.; Moulds, M.L. Recalling positive self-defining memories in depression: The impact of processing mode. Memory 2014, 22, 525–535. [Google Scholar] [CrossRef]
- Werner-Seidler, A.; Moulds, M.L. Autobiographical memory characteristics in depression vulnerability: Formerly depressed individuals recall less vivid positive memories. Cogn. Emot. 2011, 25, 1087–1103. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, M.; Yaroslavsky, I.; Rottenberg, J.; George, C.J.; Baji, I.; Benak, I.; Dochnal, R.; Halas, K.; Kiss, E.; Vetro, A.; et al. Mood repair via attention refocusing or recall of positive autobiographical memories by adolescents with pediatric-onset major depression. J. Child. Psychol. Psychiatry 2015, 56, 1108–1117. [Google Scholar] [CrossRef]
- Renner, F.; Siep, N.; Lobbestael, J.; Arntz, A.; Peeters, F.P.; Huibers, M.J. Neural correlates of self-referential processing and implicit self-associations in chronic depression. J. Affect. Disord. 2015, 186, 40–47. [Google Scholar] [CrossRef]
- Sebastian, C.; Burnett, S.; Blakemore, S.J. Development of the self-concept during adolescence. Trends Cogn. Sci. 2008, 12, 441–446. [Google Scholar] [CrossRef]
- Mochcovitch, M.D.; da Rocha Freire, R.C.; Garcia, R.F.; Nardi, A.E. A systematic review of fMRI studies in generalized anxiety disorder: Evaluating its neural and cognitive basis. J. Affect. Disord. 2014, 167, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Gee, D.G.; Hanson, C.; Caglar, L.R.; Fareri, D.S.; Gabard-Durnam, L.J.; Mills-Finnerty, C.; Goff, B.; Caldera, C.J.; Lumian, D.S.; Flannery, J.; et al. Experimental evidence for a child-to-adolescent switch in human amygdala-prefrontal cortex communication: A cross-sectional pilot study. Dev. Sci. 2022, 25, e13238. [Google Scholar] [CrossRef] [PubMed]
- Davey, C.G.; Whittle, S.; Harrison, B.J.; Simmons, J.G.; Byrne, M.L.; Schwartz, O.S.; Allen, N.B. Functional brain-imaging correlates of negative affectivity and the onset of first-episode depression. Psychol. Med. 2015, 45, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Laxmi, T.R.; Chattarji, S. Functional connectivity from the amygdala to the hippocampus grows stronger after stress. J. Neurosci. 2013, 33, 7234–7244. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.P.; Strange, B.A.; Dolan, R.J. Encoding of emotional memories depends on amygdala and hippocampus and their interactions. Nat. Neurosci. 2004, 7, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Young, K.D.; Zotev, V.; Phillips, R.; Misaki, M.; Yuan, H.; Drevets, W.C.; Bodurka, J. Real-time FMRI neurofeedback training of amygdala activity in patients with major depressive disorder. PLoS ONE 2014, 9, e88785. [Google Scholar] [CrossRef] [PubMed]
- Young, K.D.; Zotev, V.; Phillips, R.; Misaki, M.; Drevets, W.C.; Bodurka, J. Amygdala real-time functional magnetic resonance imaging neurofeedback for major depressive disorder: A review. Psychiatry Clin. Neurosci. 2018, 72, 466–481. [Google Scholar] [CrossRef] [PubMed]
- Young, K.D.; Misaki, M.; Harmer, C.J.; Victor, T.; Zotev, V.; Phillips, R.; Siegle, G.J.; Drevets, W.C.; Bodurka, J. Real-Time Functional Magnetic Resonance Imaging Amygdala Neurofeedback Changes Positive Information Processing in Major Depressive Disorder. Biol. Psychiatry 2017, 82, 578–586. [Google Scholar] [CrossRef]
- Linden, D.E.; Habes, I.; Johnston, S.J.; Linden, S.; Tatineni, R.; Subramanian, L.; Sorger, B.; Healy, D.; Goebel, R. Real-time self-regulation of emotion networks in patients with depression. PLoS ONE 2012, 7, e38115. [Google Scholar] [CrossRef]
- Mehler, D.M.A.; Sokunbi, M.O.; Habes, I.; Barawi, K.; Subramanian, L.; Range, M.; Evans, J.; Hood, K.; Luhrs, M.; Keedwell, P.; et al. Targeting the affective brain-a randomized controlled trial of real-time fMRI neurofeedback in patients with depression. Neuropsychopharmacology 2018, 43, 2578–2585. [Google Scholar] [CrossRef]
- Zotev, V.; Bodurka, J. Effects of simultaneous real-time fMRI and EEG neurofeedback in major depressive disorder evaluated with brain electromagnetic tomography. Neuroimage Clin. 2020, 28, 102459. [Google Scholar] [CrossRef] [PubMed]
- Young, K.D.; Siegle, G.J.; Zotev, V.; Phillips, R.; Misaki, M.; Yuan, H.; Drevets, W.C.; Bodurka, J. Randomized Clinical Trial of Real-Time fMRI Amygdala Neurofeedback for Major Depressive Disorder: Effects on Symptoms and Autobiographical Memory Recall. Am. J. Psychiatry 2017, 174, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Zotev, V.; Phillips, R.; Young, K.D.; Drevets, W.C.; Bodurka, J. Prefrontal control of the amygdala during real-time fMRI neurofeedback training of emotion regulation. PLoS ONE 2013, 8, e79184. [Google Scholar] [CrossRef] [PubMed]
- Quevedo, K.; Harms, M.; Sauder, M.; Scott, H.; Mohamed, S.; Thomas, K.M.; Schallmo, M.P.; Smyda, G. The neurobiology of self face recognition among depressed adolescents. J. Affect. Disord. 2018, 229, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Uher, R.; Payne, J.L.; Pavlova, B.; Perlis, R.H. Major depressive disorder in DSM-5: Implications for clinical practice and research of changes from DSM-IV. Depress. Anxiety 2014, 31, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Snyder, M.; White, P. Moods and memories: Elation, depression, and the remembering of the events of one’s life. J. Pers. 1982, 50, 149–167. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, S.J.; Hemsley, D.R. Depression and the accessibility of memories. A longitudinal study. Br. J. Psychiatry 1983, 142, 232–237. [Google Scholar] [CrossRef]
- Chen, C.; Takahashi, T.; Yang, S. Remembrance of happy things past: Positive autobiographical memories are intrinsically rewarding and valuable, but not in depression. Front. Psychol. 2015, 6, 222. [Google Scholar] [CrossRef]
- Dillon, D.G.; Dobbins, I.G.; Pizzagalli, D.A. Weak reward source memory in depression reflects blunted activation of VTA/SN and parahippocampus. Soc. Cogn. Affect. Neurosci. 2014, 9, 1576–1583. [Google Scholar] [CrossRef]
- Sugiura, M.; Yomogida, Y.; Mano, Y.; Sassa, Y.; Kambara, T.; Sekiguchi, A.; Kawashima, R. From social-signal detection to higher social cognition: An fMRI approach. Soc. Cogn. Affect. Neurosci. 2014, 9, 1303–1309. [Google Scholar] [CrossRef]
- Habel, U.; Windischberger, C.; Derntl, B.; Robinson, S.; Kryspin-Exner, I.; Gur, R.C.; Moser, E. Amygdala activation and facial expressions: Explicit emotion discrimination versus implicit emotion processing. Neuropsychologia 2007, 45, 2369–2377. [Google Scholar] [CrossRef]
- Fenker, D.B.; Schott, B.H.; Richardson-Klavehn, A.; Heinze, H.J.; Duzel, E. Recapitulating emotional context: Activity of amygdala, hippocampus and fusiform cortex during recollection and familiarity. Eur. J. Neurosci. 2005, 21, 1993–1999. [Google Scholar] [CrossRef] [PubMed]
- Devue, C.; Bredart, S. The neural correlates of visual self-recognition. Conscious Cogn. 2011, 20, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.J.; Yang, G.F.; Zhang, X.Y.; Wang, Y.F.; Liu, Y.; Zheng, G.; Lu, G.M.; Zhang, L.J.; Han, Y. Altered amygdala and hippocampus effective connectivity in mild cognitive impairment patients with depression: A resting-state functional MR imaging study with granger causality analysis. Oncotarget 2017, 8, 25021–25031. [Google Scholar] [CrossRef] [PubMed]
- Tahmasian, M.; Knight, D.C.; Manoliu, A.; Schwerthoffer, D.; Scherr, M.; Meng, C.; Shao, J.; Peters, H.; Doll, A.; Khazaie, H.; et al. Aberrant intrinsic connectivity of hippocampus and amygdala overlap in the fronto-insular and dorsomedial-prefrontal cortex in major depressive disorder. Front. Hum. Neurosci. 2013, 7, 639. [Google Scholar] [CrossRef] [PubMed]
- Hastings, R.S.; Parsey, R.V.; Oquendo, M.A.; Arango, V.; Mann, J.J. Volumetric analysis of the prefrontal cortex, amygdala, and hippocampus in major depression. Neuropsychopharmacology 2004, 29, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Benson, B.E.; Willis, M.W.; Ketter, T.A.; Speer, A.; Kimbrell, T.A.; Herscovitch, P.; George, M.S.; Post, R.M. Differential abnormalities of functional connectivity of the amygdala and hippocampus in unipolar and bipolar affective disorders. J. Affect. Disord. 2014, 168, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Belzung, C.; Willner, P.; Philippot, P. Depression: From psychopathology to pathophysiology. Curr. Opin. Neurobiol. 2015, 30, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.P.; Bittencourt-Hewitt, A.; Sebastian, C.L. Neurocognitive bases of emotion regulation development in adolescence. Dev. Cogn. Neurosci. 2015, 15, 11–25. [Google Scholar] [CrossRef]
- Hare, T.A.; Tottenham, N.; Galvan, A.; Voss, H.U.; Glover, G.H.; Casey, B.J. Biological substrates of emotional reactivity and regulation in adolescence during an emotional go-nogo task. Biol. Psychiatry 2008, 63, 927–934. [Google Scholar] [CrossRef]
- Quevedo, K.; Liu, G.; Teoh, J.Y.; Ghosh, S.; Zeffiro, T.; Ahrweiler, N.; Zhang, N.; Wedan, R.; Oh, S.; Guercio, G.; et al. Neurofeedback and neuroplasticity of visual self-processing in depressed and healthy adolescents: A preliminary study. Dev. Cogn. Neurosci. 2019, 40, 100707. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, N.; Siegle, G.J.; Jones, N.P.; Kamishima, K.; Thompson, W.K.; Gross, J.J.; Ohira, H. Feeling bad about screwing up: Emotion regulation and action monitoring in the anterior cingulate cortex. Cogn. Affect. Behav. Neurosci. 2011, 11, 354–371. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Masaoka, Y.; Kawamura, M.; Yoshida, M.; Koiwa, N.; Yoshikawa, A.; Kubota, S.; Ida, M.; Ono, K.; Izumizaki, M. Left Posterior Orbitofrontal Cortex Is Associated With Odor-Induced Autobiographical Memory: An fMRI Study. Front. Psychol. 2018, 9, 687. [Google Scholar] [CrossRef] [PubMed]
- Klumpp, H.; Keutmann, M.K.; Fitzgerald, D.A.; Shankman, S.A.; Phan, K.L. Resting state amygdala-prefrontal connectivity predicts symptom change after cognitive behavioral therapy in generalized social anxiety disorder. Biol. Mood Anxiety Disord. 2014, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Ahrweiler, N.; Santana-Gonzalez, C.; Zhang, N.; Quandt, G.; Ashtiani, N.; Liu, G.; Engstrom, M.; Schultz, E.; Liengswangwong, R.; Teoh, J.Y.; et al. Neural Activity Associated with Symptoms Change in Depressed Adolescents following Self-Processing Neurofeedback. Brain. Sci. 2022, 12, 1128. [Google Scholar] [CrossRef] [PubMed]
- Quevedo, K.; Yuan Teoh, J.; Engstrom, M.; Wedan, R.; Santana-Gonzalez, C.; Zewde, B.; Porter, D.; Cohen Kadosh, K. Amygdala Circuitry During Neurofeedback Training and Symptoms’ Change in Adolescents With Varying Depression. Front. Behav. Neurosci. 2020, 14, 110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Raya, J.; Morfini, F.; Urban, Z.; Pagliaccio, D.; Yendiki, A.; Auerbach, R.P.; Bauer, C.C.C.; Whitfield-Gabrieli, S. Reducing default mode network connectivity with mindfulness-based fMRI neurofeedback: A pilot study among adolescents with affective disorder history. Mol. Psychiatry 2023, 28, 2540–2548. [Google Scholar] [CrossRef]
- Hashemian, P.; Sadjadi, S. Evaluation of neurofeedback therapy in adolescents with major depressive disorder who take fluoxetine. J. Psychiatry 2015, 18, 180. [Google Scholar]
- Nawa, N.E.; Ando, H. Effective connectivity within the ventromedial prefrontal cortex-hippocampus-amygdala network during the elaboration of emotional autobiographical memories. NeuroImage 2019, 189, 316–328. [Google Scholar] [CrossRef]
- De Voogd, L.D.; Klumpers, F.; Fernandez, G.; Hermans, E.J. Intrinsic functional connectivity between amygdala and hippocampus during rest predicts enhanced memory under stress. Psychoneuroendocrinology 2017, 75, 192–202. [Google Scholar] [CrossRef]
- Zyss, T. Expert decisions concerning disability pensions in the area of psychiatry--in view of a court-appointed expert. Arch. Med. Sadowej Kryminol. 2005, 55, 338–348. [Google Scholar] [PubMed]
- Luyten, P.; Fonagy, P. The stress-reward-mentalizing model of depression: An integrative developmental cascade approach to child and adolescent depressive disorder based on the Research Domain Criteria (RDoC) approach. Clin. Psychol. Rev. 2018, 64, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Dolcos, F.; LaBar, K.S.; Cabeza, R. Remembering one year later: Role of the amygdala and the medial temporal lobe memory system in retrieving emotional memories. Proc. Natl. Acad. Sci. USA 2005, 102, 2626–2631. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, L.; Comte, A.; Tatu, L.; Millot, J.L.; Moulin, T.; Medeiros de Bustos, E. The role of the amygdala in the perception of positive emotions: An “intensity detector”. Front. Behav. Neurosci. 2015, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Krause-Utz, A.; Winter, D.; Niedtfeld, I.; Schmahl, C. The latest neuroimaging findings in borderline personality disorder. Curr. Psychiatry Rep. 2014, 16, 438. [Google Scholar] [CrossRef] [PubMed]
- Jaworska, N.; Yang, X.R.; Knott, V.; MacQueen, G. A review of fMRI studies during visual emotive processing in major depressive disorder. World J. Biol. Psychiatry 2015, 16, 448–471. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yan, J.; Wang, D.; Sun, M.; Zhu, Y.; Zhu, X.; Jiang, P.; Yin, R.; Zhao, L. Magnetic resonance study of the structure and function of the hippocampus and amygdala in patients with depression. Chin. Med. J. 2014, 127, 3610–3615. [Google Scholar]
- Nejad, A.B.; Fossati, P.; Lemogne, C. Self-referential processing, rumination, and cortical midline structures in major depression. Front. Hum. Neurosci. 2013, 7, 666. [Google Scholar] [CrossRef]
- Lemogne, C.; le Bastard, G.; Mayberg, H.; Volle, E.; Bergouignan, L.; Lehericy, S.; Allilaire, J.F.; Fossati, P. In search of the depressive self: Extended medial prefrontal network during self-referential processing in major depression. Soc. Cogn. Affect. Neurosci. 2009, 4, 305–312. [Google Scholar] [CrossRef]
- Otterpohl, N.; Wild, E. Cross-lagged relations among parenting, children’s emotion regulation, and psychosocial adjustment in early adolescence. J. Clin. Child Adolesc. Psychol. 2015, 44, 93–108. [Google Scholar] [CrossRef]
- Jones, J.D.; Brett, B.E.; Ehrlich, K.B.; Lejuez, C.W.; Cassidy, J. Maternal Attachment Style and Responses to Adolescents’ Negative Emotions: The Mediating Role of Maternal Emotion Regulation. Parent Sci. Pract. 2014, 14, 235–257. [Google Scholar] [CrossRef] [PubMed]
- Siffert, A.; Schwarz, B. Parental conflict resolution styles and children’s adjustment: Children’s appraisals and emotion regulation as mediators. J. Genet. Psychol. 2011, 172, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.; Birmaher, B.; Brent, D.; Rao, U.; Flynn, C.; Moreci, P.; Williamson, D.; Ryan, N. Schedule for Affective Disorders and Schizophrenia for School-Age Children-Present and Lifetime Version (K-SADS-PL): Initial reliability and validity data. J. Am. Acad. Child. Adolesc. Psychiatry 1997, 36, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Poznanski, E.O.; Cook, S.C.; Carroll, B.J. A depression rating scale for children. Pediatrics 1979, 64, 442–450. [Google Scholar] [CrossRef]
- Sorger, B.; Scharnowski, F.; Linden, D.E.J.; Hampson, M.; Young, K.D. Control freaks: Towards optimal selection of control conditions for fMRI neurofeedback studies. NeuroImage 2019, 186, 256–265. [Google Scholar] [CrossRef]
- Malatesta-Magai, C. Emotional socialization: Its role in personality and developmental psychopathology. In Internalizing and Externalizing Expressions of Dysfunction; Rochester Symposium on Developmental Psychopathology; Lawrence Erlbaum Associates, Inc.: Hillsdale, NJ, USA, 1991; Volume 2, pp. 203–224. [Google Scholar]
- Peirce, J.W.; Gray, J.R.; Simpson, S.; MacAskill, M.R.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J. Psy-choPy2: Experiments in behavior made easy. Behav. Res. Methods 2019, 51, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Hinds, O.; Ghosh, S.; Thompson, T.W.; Yoo, J.J.; Whitfield-Gabrieli, S.; Triantafyllou, C.; Gabrieli, J.D. Computing moment-to-moment BOLD activation for real-time neurofeedback. NeuroImage 2011, 54, 361–368. [Google Scholar] [CrossRef]
- Woodward, T.S.; Cairo, T.A.; Ruff, C.C.; Takane, Y.; Hunter, M.A.; Ngan, E.T. Functional connectivity reveals load dependent neural systems underlying encoding and maintenance in verbal working memory. Neuroscience 2006, 139, 317–325. [Google Scholar] [CrossRef]
- Maldjian, J.A.; Laurienti, P.J.; Kraft, R.A.; Burdette, J.H. An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. NeuroImage 2003, 19, 1233–1239. [Google Scholar] [CrossRef]
- Ashburner, J.; Barnes, G.; Chen, C.-C.; Daunizeau, J.; Flandin, G.; Friston, K.; Kiebel, S.; Kilner, J.; Litvak, V.; Moran, R. SPM12 Manual; Wellcome Trust Centre for Neuroimaging: London, UK, 2014; Volume 2464. [Google Scholar]
- Cox, R.W. AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Comput. Biomed. Res. 1996, 29, 162–173. [Google Scholar] [CrossRef]
- Young, K.D.; Siegle, G.J.; Misaki, M.; Zotev, V.; Phillips, R.; Drevets, W.C.; Bodurka, J. Altered task-based and resting-state amygdala functional connectivity following real-time fMRI amygdala neurofeedback training in major depressive disorder. Neuroimage Clin. 2018, 17, 691–703. [Google Scholar] [CrossRef]
- Keller, M.; Zweerings, J.; Klasen, M.; Zvyagintsev, M.; Iglesias, J.; Mendoza Quinones, R.; Mathiak, K. fMRI Neurofeedback-Enhanced Cognitive Reappraisal Training in Depression: A Double-Blind Comparison of Left and Right vlPFC Regulation. Front. Psychiatry 2021, 12, 715898. [Google Scholar] [CrossRef] [PubMed]
- Lorenzetti, V.; Melo, B.; Basilio, R.; Suo, C.; Yucel, M.; Tierra-Criollo, C.J.; Moll, J. Emotion Regulation Using Virtual Environments and Real-Time fMRI Neurofeedback. Front. Neurol. 2018, 9, 390. [Google Scholar] [CrossRef] [PubMed]
- Paret, C.; Kluetsch, R.; Ruf, M.; Demirakca, T.; Hoesterey, S.; Ende, G.; Schmahl, C. Down-regulation of amygdala activation with real-time fMRI neurofeedback in a healthy female sample. Front. Behav. Neurosci. 2014, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Linhartova, P.; Latalova, A.; Kosa, B.; Kasparek, T.; Schmahl, C.; Paret, C. fMRI neurofeedback in emotion regulation: A literature review. NeuroImage 2019, 193, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Barry, D.N.; Clark, I.A.; Maguire, E.A. The relationship between hippocampal subfield volumes and autobiographical memory persistence. Hippocampus 2021, 31, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, V.; Cavallina, C.; Colucci, P.; Santori, A.; Macri, S.; McGaugh, J.L.; Campolongo, P. Enhanced brain activity associated with memory access in highly superior autobiographical memory. Proc. Natl. Acad. Sci. USA 2018, 115, 7795–7800. [Google Scholar] [CrossRef] [PubMed]
- Zotev, V.; Krueger, F.; Phillips, R.; Alvarez, R.P.; Simmons, W.K.; Bellgowan, P.; Drevets, W.C.; Bodurka, J. Self-regulation of amygdala activation using real-time FMRI neurofeedback. PLoS ONE 2011, 6, e24522. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.J.; Boehm, S.G.; Healy, D.; Goebel, R.; Linden, D.E. Neurofeedback: A promising tool for the self-regulation of emotion networks. NeuroImage 2010, 49, 1066–1072. [Google Scholar] [CrossRef]
- Johnston, S.; Linden, D.E.; Healy, D.; Goebel, R.; Habes, I.; Boehm, S.G. Upregulation of emotion areas through neurofeedback with a focus on positive mood. Cogn. Affect. Behav. Neurosci. 2011, 11, 44–51. [Google Scholar] [CrossRef]
- Bylsma, L.M.; Yaroslavsky, I.; Rottenberg, J.; Kiss, E.; Kapornai, K.; Halas, K.; Dochnal, R.; Lefkovics, E.; Baji, I.; Vetromicron, A.; et al. Familiality of mood repair responses among youth with and without histories of depression. Cogn. Emot. 2016, 30, 807–816. [Google Scholar] [CrossRef]
- Aldao, A.; Nolen-Hoeksema, S.; Schweizer, S. Emotion-regulation strategies across psychopathology: A meta-analytic review. Clin. Psychol. Rev. 2010, 30, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Werner-Seidler, A.; Moulds, M.L. Characteristics of self-defining memory in depression vulnerability. Memory 2012, 20, 935–948. [Google Scholar] [CrossRef] [PubMed]
- Breiter, H.C.; Etcoff, N.L.; Whalen, P.J.; Kennedy, W.A.; Rauch, S.L.; Buckner, R.L.; Strauss, M.M.; Hyman, S.E.; Rosen, B.R. Response and habituation of the human amygdala during visual processing of facial expression. Neuron 1996, 17, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Maza, M.T.; Fox, K.A.; Kwon, S.J.; Flannery, J.E.; Lindquist, K.A.; Prinstein, M.J.; Telzer, E.H. Association of Habitual Checking Behaviors on Social Media With Longitudinal Functional Brain Development. JAMA Pediatr. 2023, 177, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Ghobadi-Azbari, P.; Mahdavifar Khayati, R.; Ekhtiari, H. Habituation or sensitization of brain response to food cues: Temporal dynamic analysis in an functional magnetic resonance imaging study. Front. Hum. Neurosci. 2023, 17, 1076711. [Google Scholar] [CrossRef] [PubMed]
- Gyurak, A.; Gross, J.J.; Etkin, A. Explicit and implicit emotion regulation: A dual-process framework. Cogn. Emot. 2011, 25, 400–412. [Google Scholar] [CrossRef]
- Fuentes, A.; Desrocher, M. The effects of gender on the retrieval of episodic and semantic components of autobiographical memory. Memory 2013, 21, 619–632. [Google Scholar] [CrossRef]
- Noakes, M.A.; Rinaldi, C.M. Age and Gender Differences in Peer Conflict. J. Youth Adolesc. 2006, 35, 881–891. [Google Scholar] [CrossRef]
- Nolen-Hoeksema, S. Emotion regulation and psychopathology: The role of gender. Annu. Rev. Clin. Psychol. 2012, 8, 161–187. [Google Scholar] [CrossRef]
- Alegria, A.A.; Wulff, M.; Brinson, H.; Barker, G.J.; Norman, L.J.; Brandeis, D.; Stahl, D.; David, A.S.; Taylor, E.; Giampietro, V.; et al. Real-time fMRI neurofeedback in adolescents with attention deficit hyperactivity disorder. Hum. Brain Mapp. 2017, 38, 3190–3209. [Google Scholar] [CrossRef] [PubMed]
- Werner, K.H.; Roberts, N.A.; Rosen, H.J.; Dean, D.L.; Kramer, J.H.; Weiner, M.W.; Miller, B.L.; Levenson, R.W. Emotional reactivity and emotion recognition in frontotemporal lobar degeneration. Neurology 2007, 69, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Weiner, K.S.; Zilles, K. The anatomical and functional specialization of the fusiform gyrus. Neuropsychologia 2016, 83, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Zaki, J.; Davis, J.I.; Ochsner, K.N. Overlapping activity in anterior insula during interoception and emotional experience. NeuroImage 2012, 62, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Fiori, S.; Zendler, C.; Hauser, T.K.; Lidzba, K.; Wilke, M. Assessing motor, visual and language function using a single 5-minute fMRI paradigm: Three birds with one stone. Brain Imaging Behav. 2018, 12, 1775–1785. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Wade, A.R. The negative BOLD signal unmasked. Neuron 2002, 36, 993–995. [Google Scholar] [CrossRef]
- Boddaert, N.; Zilbovicius, M. Functional neuroimaging and childhood autism. Pediatr. Radiol. 2002, 32, 1–7. [Google Scholar] [CrossRef]
- Pelphrey, K.A.; Carter, E.J. Charting the typical and atypical development of the social brain. Dev. Psychopathol. 2008, 20, 1081–1102. [Google Scholar] [CrossRef]
- Heller, A.S.; Casey, B.J. The neurodynamics of emotion: Delineating typical and atypical emotional processes during adolescence. Dev. Sci. 2016, 19, 3–18. [Google Scholar] [CrossRef]
- Weschsler, D. Wechsler Abbreviated Scale of Intelligence; Harcourt Brace & Company: The Psychological Corporation: New York, NY, USA, 1999. [Google Scholar]
- Reynolds, W.M. Suicidal Ideation and Depression in Adolescents—Assessment and Research. Proc. XXIV Int. Congr. Psychol. 1989, 9, 125–135. [Google Scholar]
- Harter, S. The Perceived Competence Scale for Children. Child. Dev. 1982, 53, 87–97. [Google Scholar] [CrossRef]
- Beck, A.T.; Steer, R.A.; Brown, G.K. Manual for the Beck Depression Inventory—II; Psychological Corporation: San Antonio, TX, USA, 1996. [Google Scholar]
- Carver, C.S.; White, T.L. Behavioral inhibition, behavioral activation, and affective responses to impending reward and punishment: The BIS/BAS scales. J. Personal. Soc. Psychol. 1994, 67, 319–333. [Google Scholar] [CrossRef]
- Robin, A.; Foster, S.L. Negotiating parent adolescent conflict: A Behavioral-Family System Approach; Guilford Press: New york, NY, USA, 1989. [Google Scholar]
- Gratz, K.L. Measurement of deliberate self-harm: Preliminary data on the Deliberate Self-Harm Inventory. J. Psychopathol. Behav. Assess. 2001, 23, 253–263. [Google Scholar] [CrossRef]
- Treynor, W.; Gonzalez, R.; Nolen-Hoeksema, S. Rumination Reconsidered: A Psychometric Analysis. Cogn. Ther. Res. 2003, 27, 247–259. [Google Scholar] [CrossRef]
- Angold, A.; Costello, E.J.; Messer, S.C.; Pickles, A.; Winder, F.; Silver, D. The development of a short questionnaire for use in epidemiological studies of depression in children and adolescents. Int. J. Methods Psychiatr. Res. 1995, 5, 237–249. [Google Scholar]
- Messer, S.C.; Angold, A.; Costello, E.J.; Loeber, R.; Van Kammen, W.; Stouthamer-Loeber, M. Development of a short questionnaire for use in epidemiological studies of depression in children and adolescents: Factor composition and structure across development. Int. J. Methods Psychiatr. Res. 1995, 5, 251–262. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controls | Depressed | Statistics | |
|---|---|---|---|
| N = 19 | N = 34 | ||
| AgeS1: M (SD) | 16.26 (1.19) | 16.08 (1.27) | F(1, 48) = 0.26 |
| AgeS2: M (SD) | 16.35 (1.23) | 16.11 (1.25) | F(1, 48) = 0.45 |
| IQ: M (SD) | 115.32 (9.12) | 108.35 (10.84) | F(1, 51) = 5.61 * |
| Sex | χ2(1) = 0.31 | ||
| Male | 7 (36.84%) | 10 (29.41%) | |
| Female | 12 (63.16%) | 24 (70.59%) | |
| Puberty: median (IQR) | 4.75 (0.50) | 5.00 (0.50) | U(19, 33) = 276.5 |
| Ethnicity | χ2(4) = 7.69 | ||
| White | 14 (73.68%) | 27 (79.41%) | |
| African American/Black | 0 | 2 (5.88%) | |
| American Indian | 0 | 2 (5.88%) | |
| Asian | 3 (15.79%) | 0 | |
| Other | 2 (10.53%) | 3 (8.82%) | |
| Family Structure | χ2(3) = 2.80 | ||
| Married | 15 (78.95%) | 22 (64.71%) | |
| Living with partner | 1 (5.26%) | 3 (8.82%) | |
| Separated-Divorced | 3 (15.79%) | 5 (14.71%) | |
| Single | 0 | 4 (11.76%) | |
| Income | χ2(2) = 3.90 | ||
| =>35 K | 0 | 6 (17.65%) | |
| 35–75 K | 7 (36.84%) | 9 (26.47%) | |
| +>75 K | 12 (63.16%) | 19 (55.88%) | |
| Medication | |||
| Antidepressants | 0 | 26 | |
| Antipsychotics | 0 | 2 | |
| Mood stabilizers | 0 | 0 | |
| Anxiolytic | 0 | 10 | |
| Depression Severity: M (SD)/median (IQR) | 19.21 (3.56)/18 (2.5) | 49.85 (16.14)/48.5 (28.5) | F(1, 51) = 66.06 ** |
| Parental Support: M (SD) | 37.95 (5.02) | 34.82 (7.14) | F(1, 50) = 1.45 |
| Predictors of Mean AMYHIPPO Activity during the Neurofeedback Task’s Time Series | |||||
|---|---|---|---|---|---|
| Effect | Estimate γ | SE γ | df | t | p |
| Intercept | 0.01 | 0.03 | 212.53 | 0.43 | 0.67 |
| Group | |||||
| Controls | 0.16 | 0.05 | 203.96 | 3.50 | <0.01 |
| Depressed | |||||
| Condition | |||||
| Count Backwards | |||||
| Neurofeedback | 0.05 | 0.02 | 364 | 2.96 | <0.01 |
| Group by Linear Slope | |||||
| Controls × Linear Slope | −0.24 | 0.01 | 364 | −4.02 | <0.01 |
| Depressed × Linear Slope | 0.002 | 0.005 | 364 | 0.52 | 0.60 |
| Predictors of Mean ACC Activity during the Neurofeedback Task’s Time Series | |||||
| Intercept | −0.526 | 0.21 | 59.25 | −2.51 | <0.05 |
| Condition | |||||
| Count Backwards | |||||
| Neurofeedback | 0.114 | 0.047 | 349.24 | 2.45 | <0.05 |
| Gender | |||||
| Female | 0.17 | 0.072 | 50.23 | 2.38 | <0.05 |
| Male | |||||
| Medication Presence | |||||
| No Medication | 0.223 | 0.093 | 102.06 | 2.40 | <0.05 |
| Medication Present | |||||
| Group by Linear Slope | |||||
| Controls × Linear Slope | −0.051 | 0.015 | 393.87 | −3.50 | <0.01 |
| Depressed × Linear Slope | −0.001 | 0.012 | 395.90 | −0.07 | 0.94 |
| Parental Support | 0.014 | 0.005 | 49.71 | 2.69 | <0.01 |
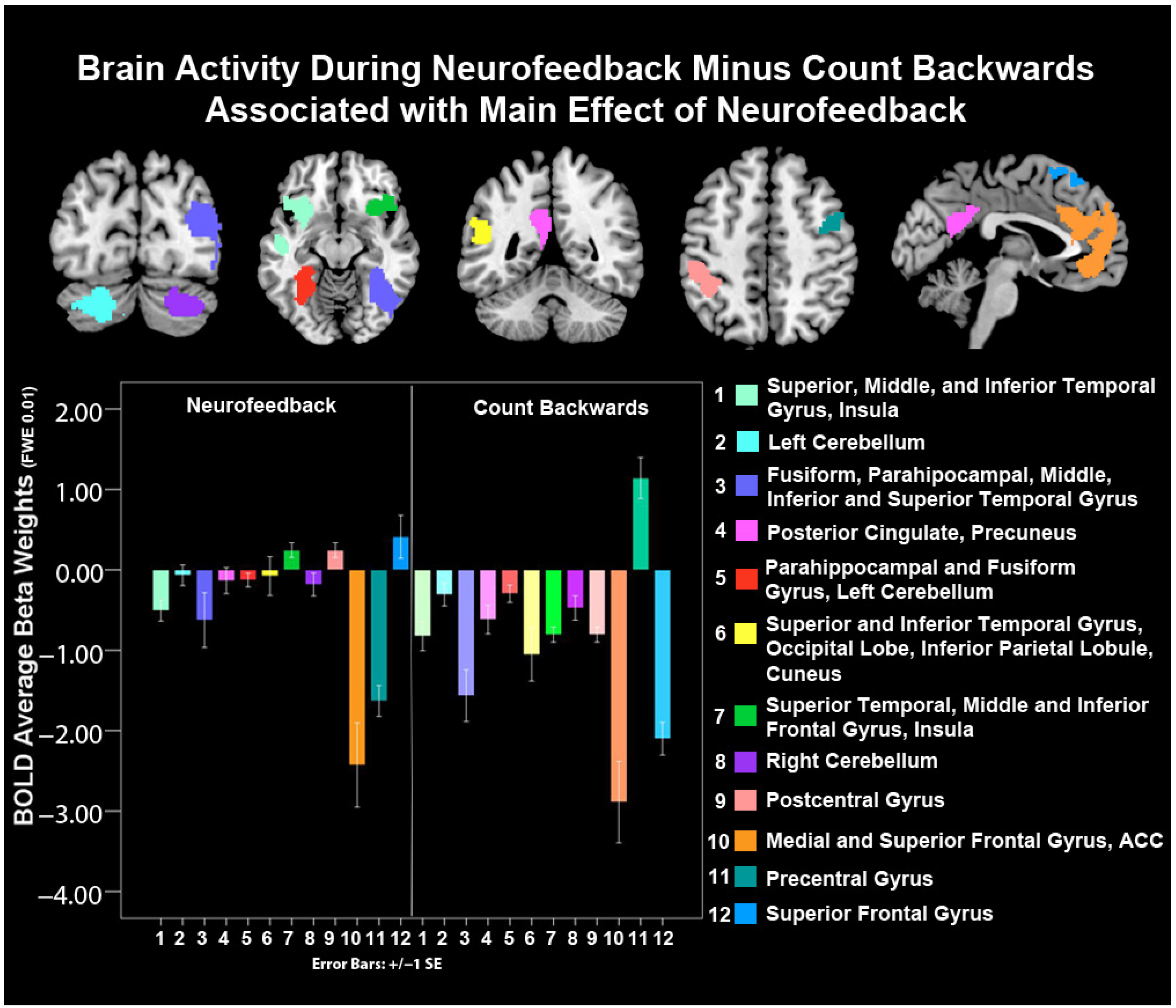
| Cluster Size (K) | Hemisphere | MNI Coordinates | F | p(K) | |||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Main Effect of Neurofeedback | |||||||
| Superior, Middle, and Inferior Temporal Gyrus, Insula BA 13, 21, 38, 47 | 1323 | Left | −28 | 12 | −18 | 137.12 | <0.001 |
| Left Cerebellum | 602 | Left | −20 | −74 | −36 | 108.83 | <0.001 |
| Fusiform, Parahippocampal, Middle, Inferior and Superior Temporal Gyrus, BA 13, 19, 21, 22, 36, 37, 39, 40 | 3925 | Right | 38 | −86 | 20 | 108.13 | <0.001 |
| Posterior Cingulate Cortex, Precuneus, BA 23, 30, 31 | 546 | Left | −06 | −50 | 22 | 104.23 | <0.001 |
| Parahippocampal and Fusiform Gyrus, Left Cerebellum, BA 19, 36, 37 | 768 | Left | −28 | −34 | −18 | 97.14 | <0.001 |
| Superior and Inferior Temporal Gyrus, Occipital Lobe, Inferior Parietal Lobule, Cuneus BA 19, 22, 27, 39, 40 | 2126 | Left | −44 | −80 | 10 | 96.35 | <0.001 |
| Superior Temporal, Middle and Inferior Frontal Gyrus, Insula, BA 47, 13, 45, 38, 46 | 1097 | Right | 36 | 26 | 00 | 97.66 | <0.001 |
| Right Cerebellum | 477 | Right | 28 | −74 | −34 | 77.71 | <0.001 |
| Postcentral Gyrus, BA, 2, 40 | 504 | Left | −46 | −36 | 44 | 74.45 | <0.001 |
| Medial and Superior Frontal Gyrus, ACC, BA 9, 10, 11, 24, 32 | 1862 | Left and Right | −06 | 56 | 14 | 73.38 | <0.001 |
| Precentral Gyrus, BA 6, 9 | 234 | Right | 38 | −04 | 46 | 68.21 | <0.001 |
| Superior Frontal Gyrus, BA, 6, 8 | 362 | Left and Right | 02 | 18 | 56 | 63.95 | <0.001 |
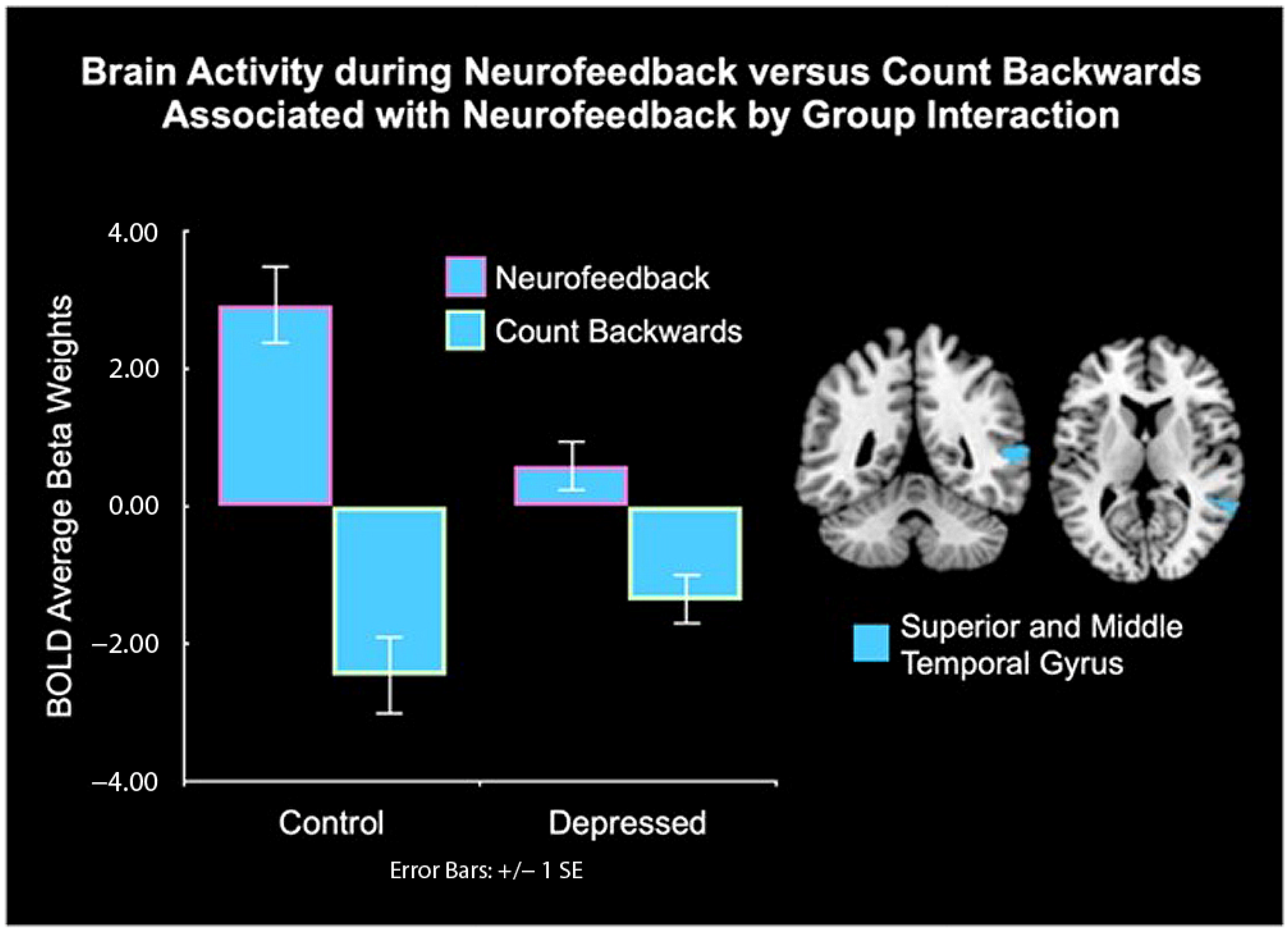
| Group by Condition Interaction | |||||||
| Superior and Middle Temporal Gyrus, BA 21, 22 | 163 | Right | 54 | −48 | 08 | 17.67 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, G.H.; Oh, S.; Schneider, C.; Teoh, J.Y.; Engstrom, M.; Santana-Gonzalez, C.; Porter, D.; Quevedo, K. Neurofeedback and Affect Regulation Circuitry in Depressed and Healthy Adolescents. Biology 2023, 12, 1399. https://doi.org/10.3390/biology12111399
Nguyen GH, Oh S, Schneider C, Teoh JY, Engstrom M, Santana-Gonzalez C, Porter D, Quevedo K. Neurofeedback and Affect Regulation Circuitry in Depressed and Healthy Adolescents. Biology. 2023; 12(11):1399. https://doi.org/10.3390/biology12111399
Chicago/Turabian StyleNguyen, Giang H., Sewon Oh, Corey Schneider, Jia Y. Teoh, Maggie Engstrom, Carmen Santana-Gonzalez, David Porter, and Karina Quevedo. 2023. "Neurofeedback and Affect Regulation Circuitry in Depressed and Healthy Adolescents" Biology 12, no. 11: 1399. https://doi.org/10.3390/biology12111399
APA StyleNguyen, G. H., Oh, S., Schneider, C., Teoh, J. Y., Engstrom, M., Santana-Gonzalez, C., Porter, D., & Quevedo, K. (2023). Neurofeedback and Affect Regulation Circuitry in Depressed and Healthy Adolescents. Biology, 12(11), 1399. https://doi.org/10.3390/biology12111399