



Towards an Understanding of Large-Scale Biodiversity Patterns on Land and in the Sea


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
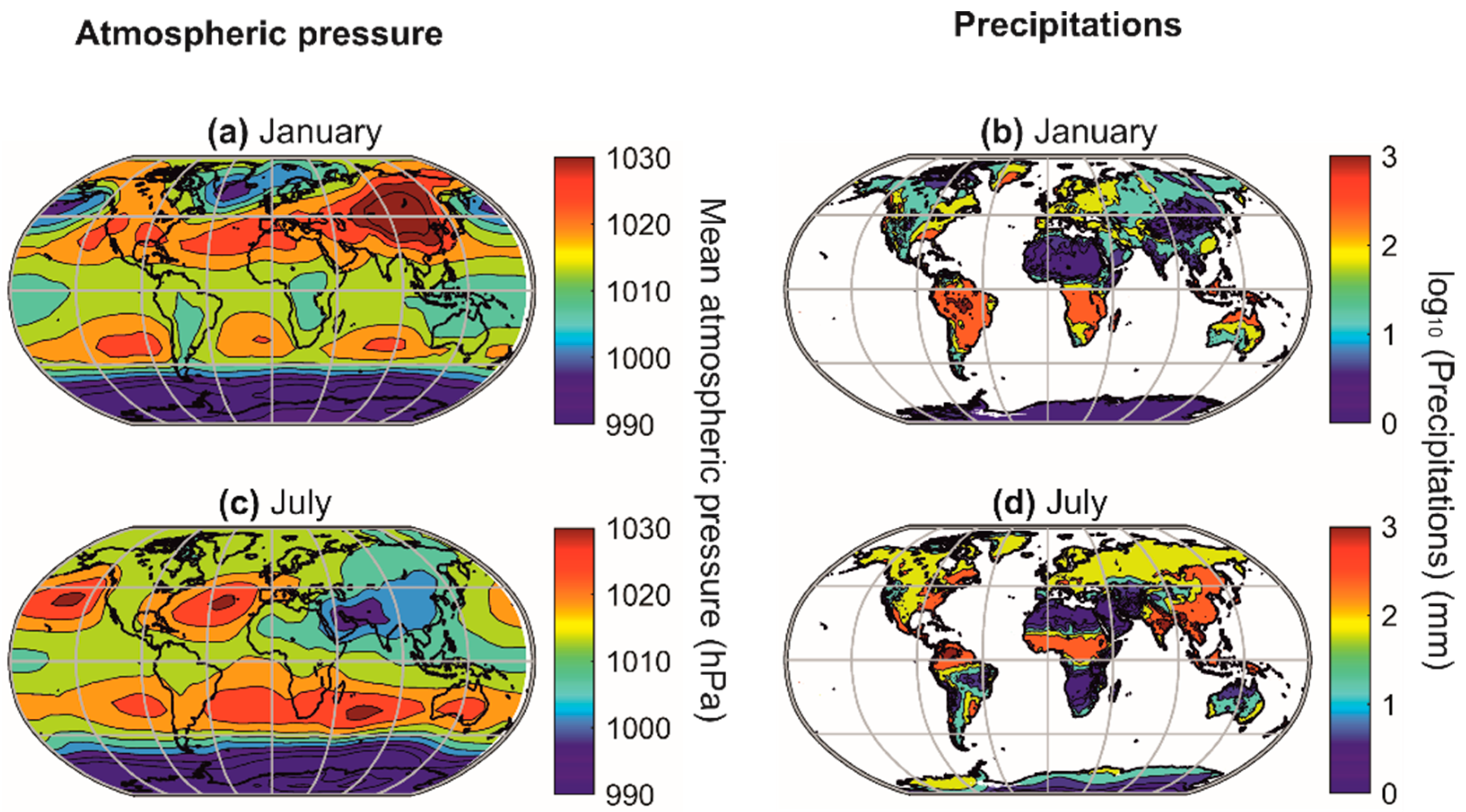
2. Patterns of Variability in Nature
3. The Difficult Identification of Patterns in Ecology
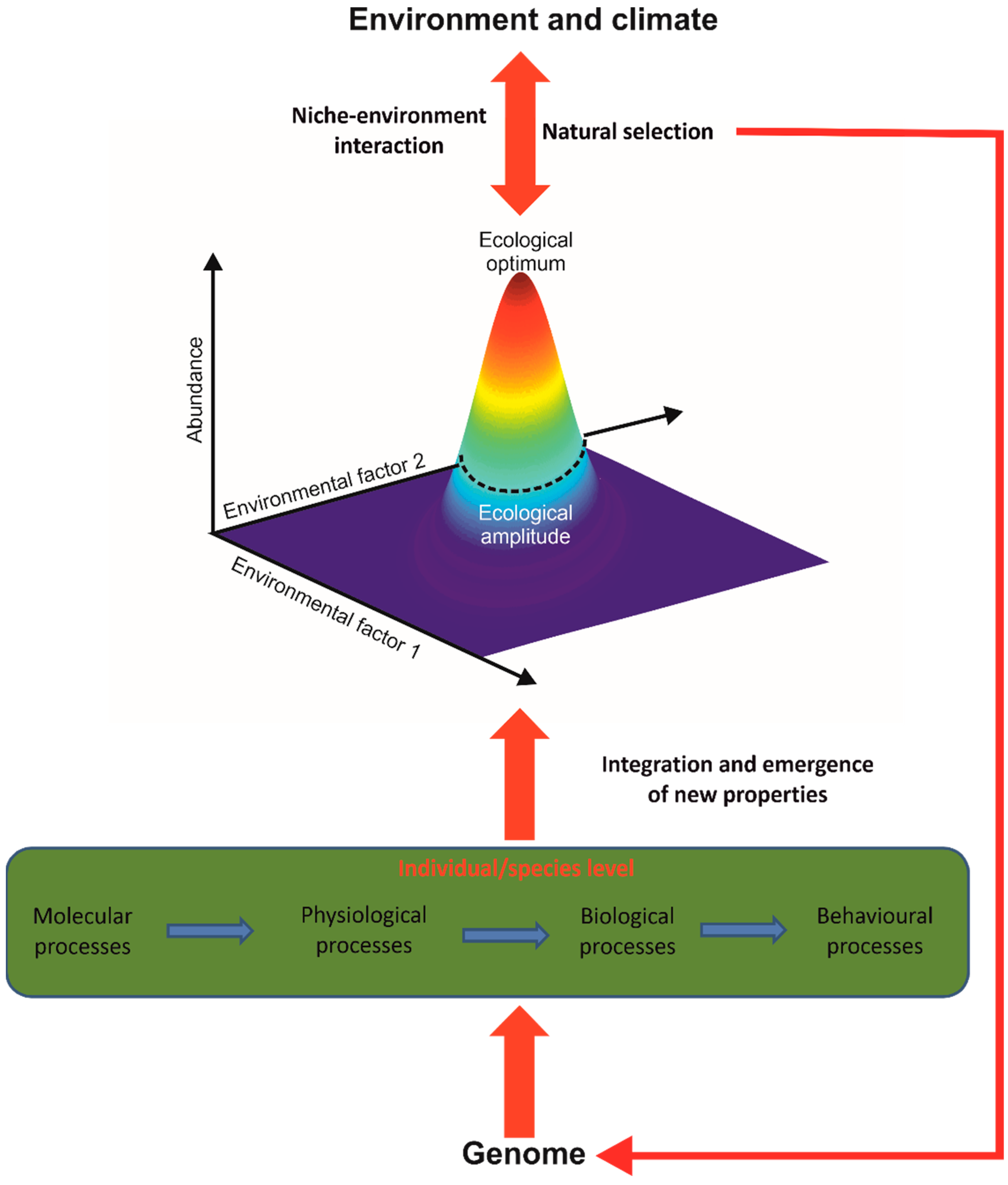
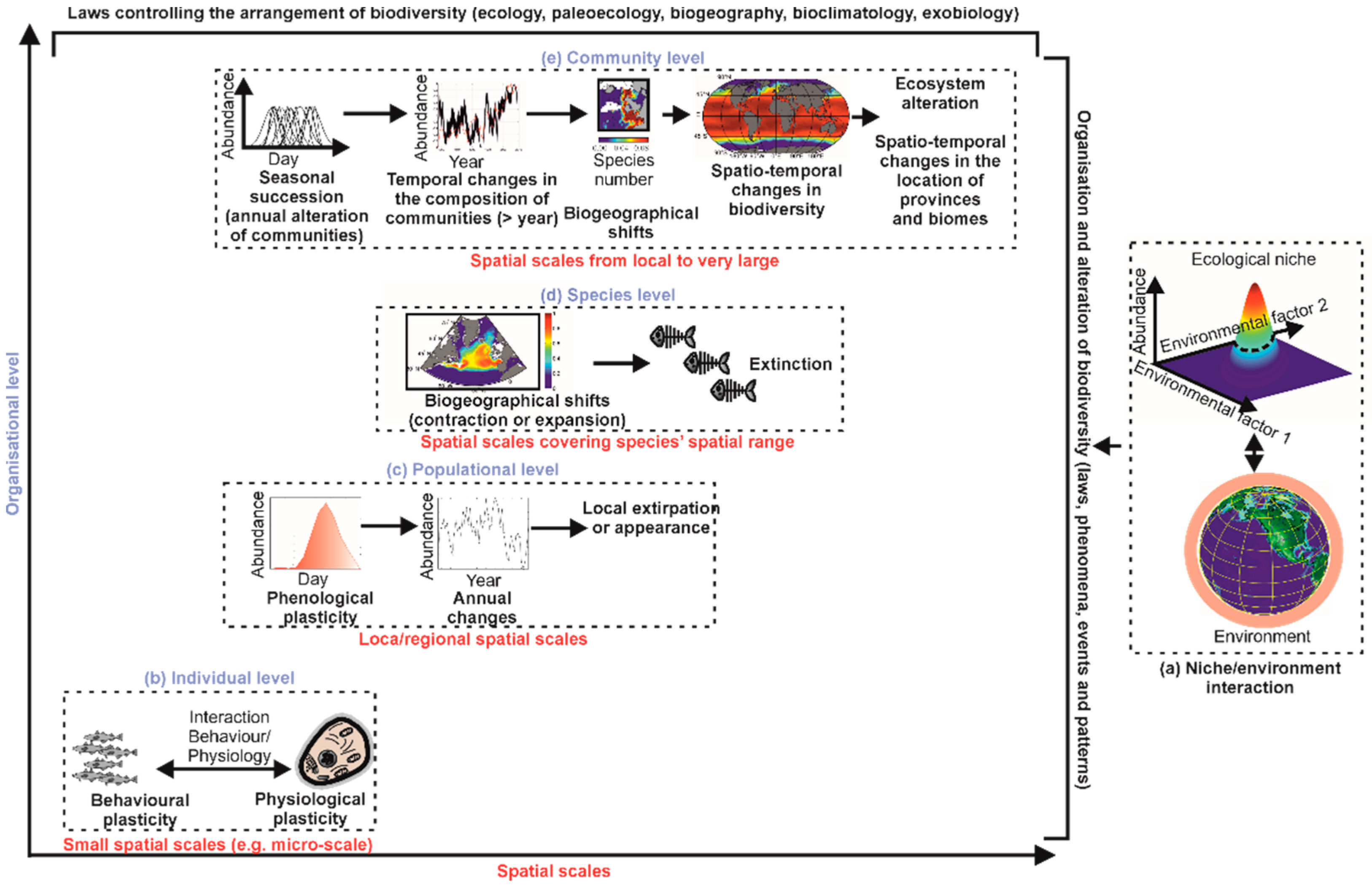
4. Towards a Better Understanding of Principles of Biodiversity Organisation and Climate Change Biology
5. Large-Scale Biodiversity Patterns
5.1. A Brief Overview of the Main Hypotheses or Theories That Have Attempted to Explain Large-Scale Biodiversity Patterns
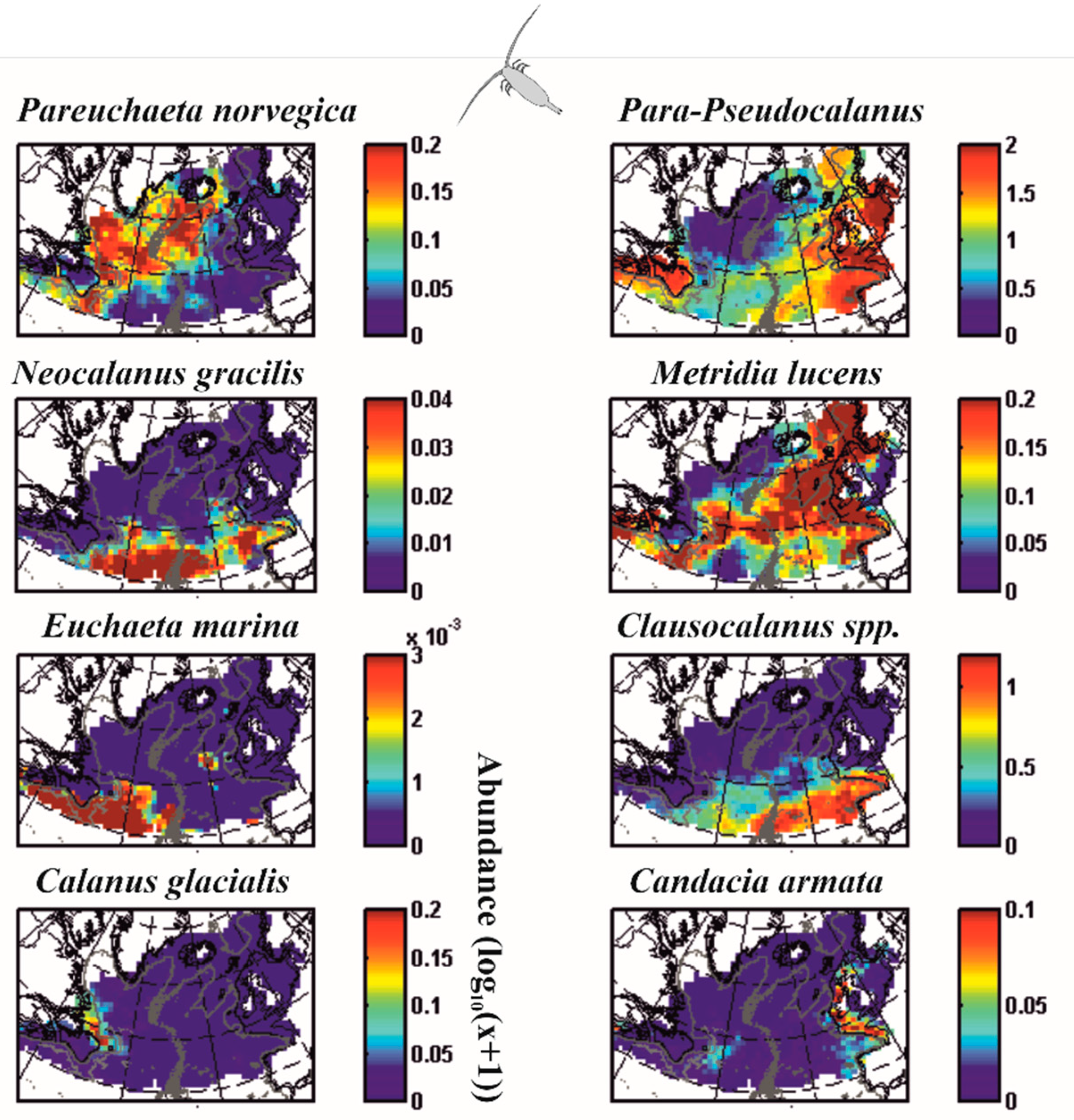
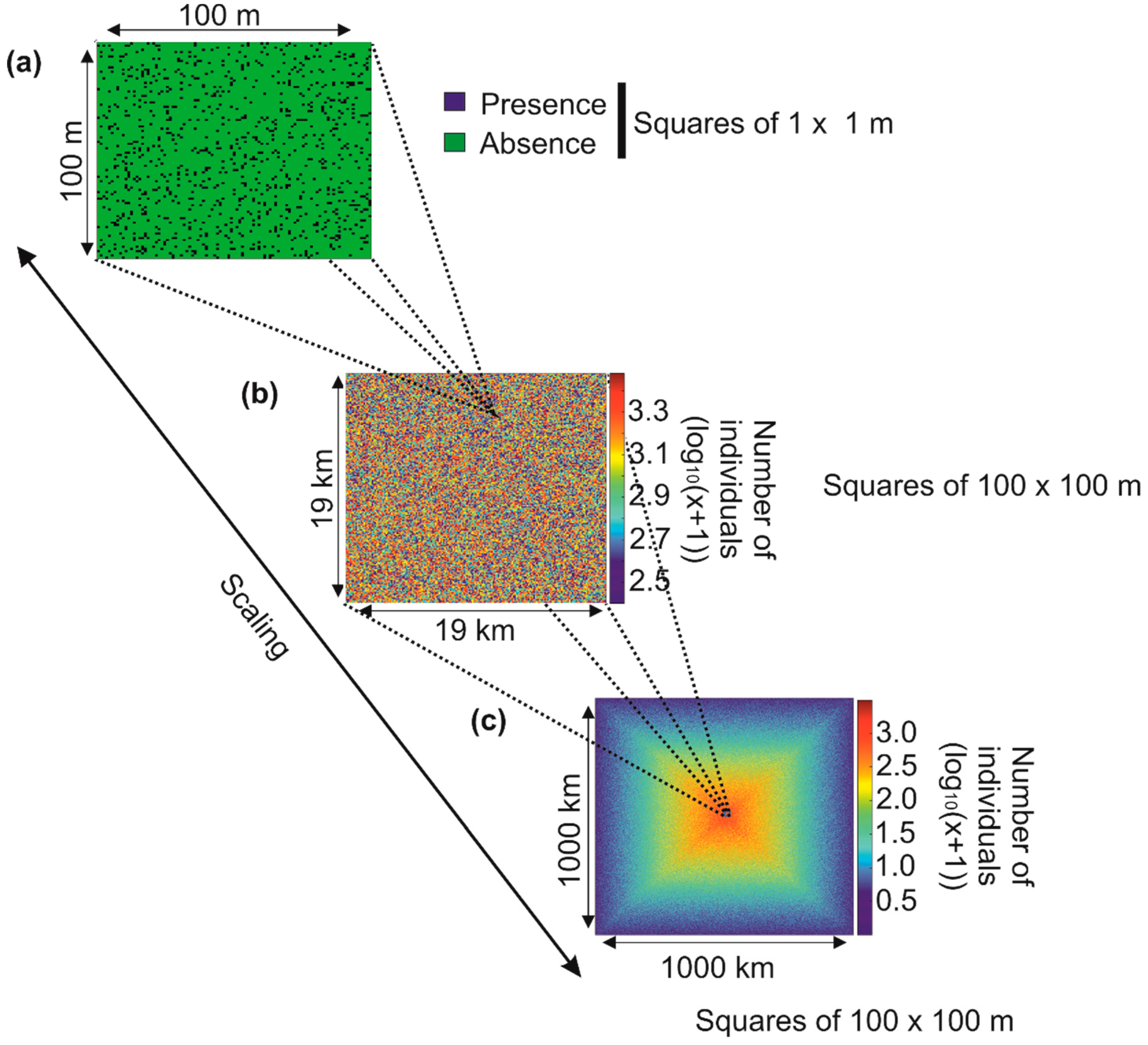
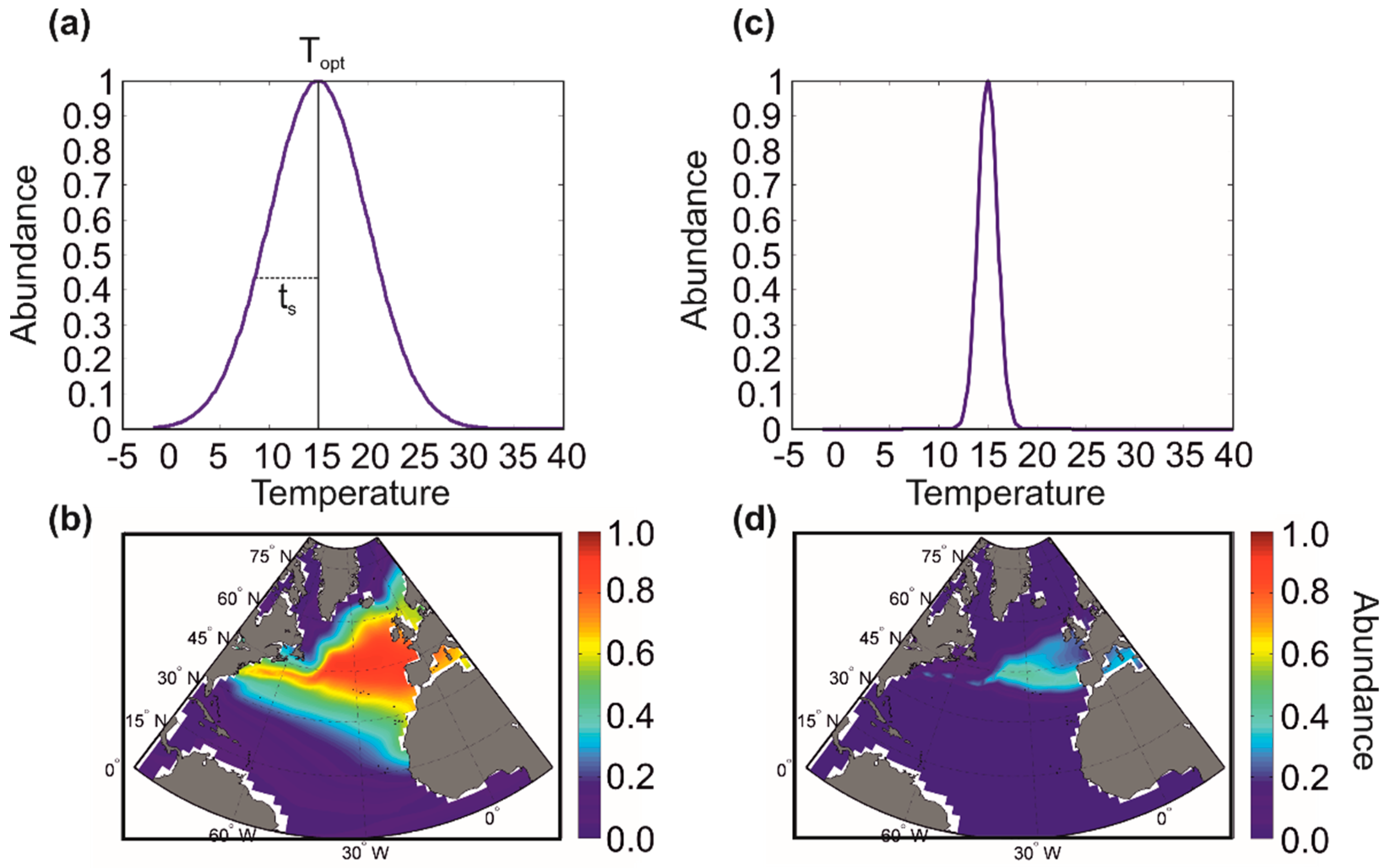
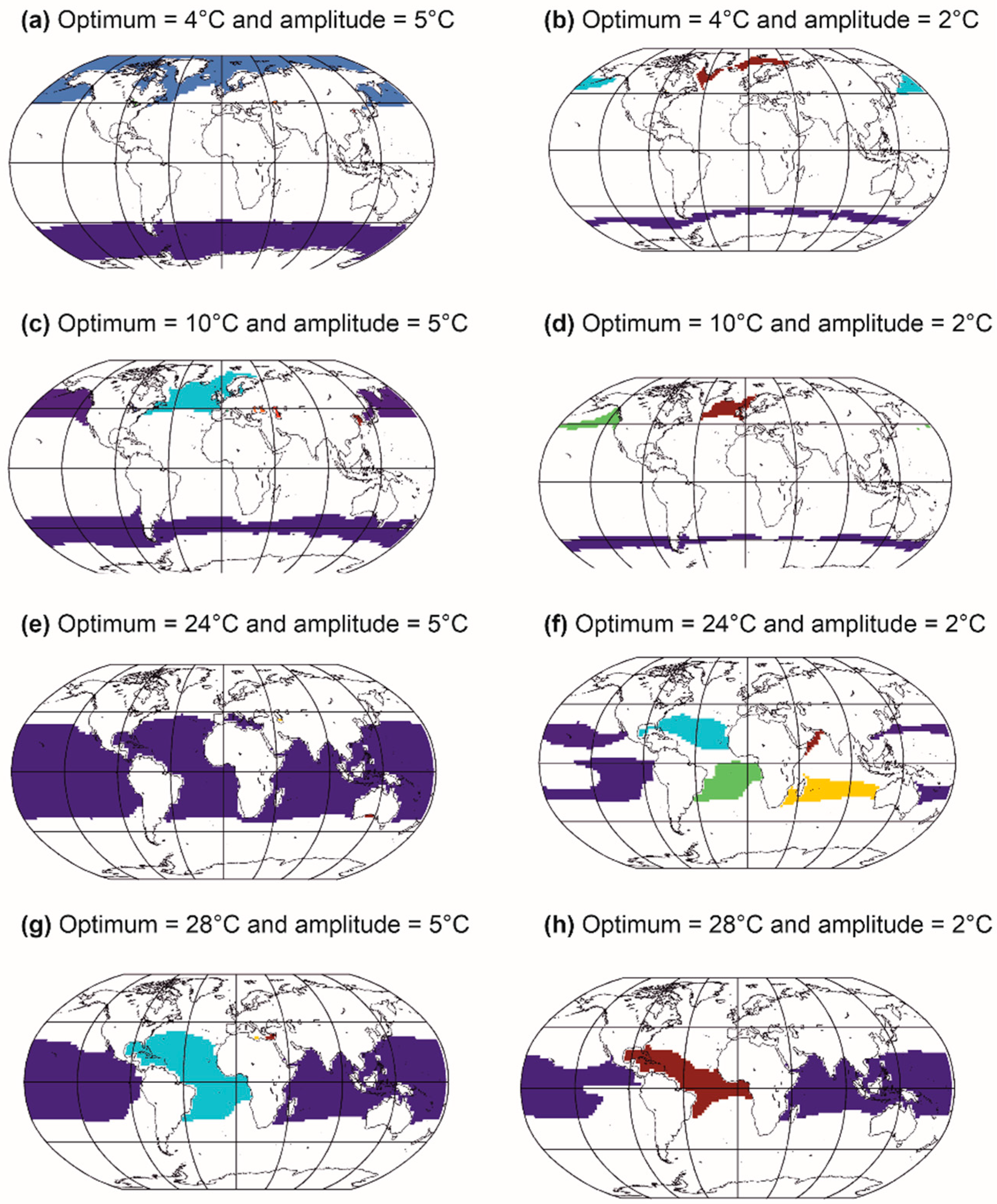
5.2. Modelling Biodiversity in METAL
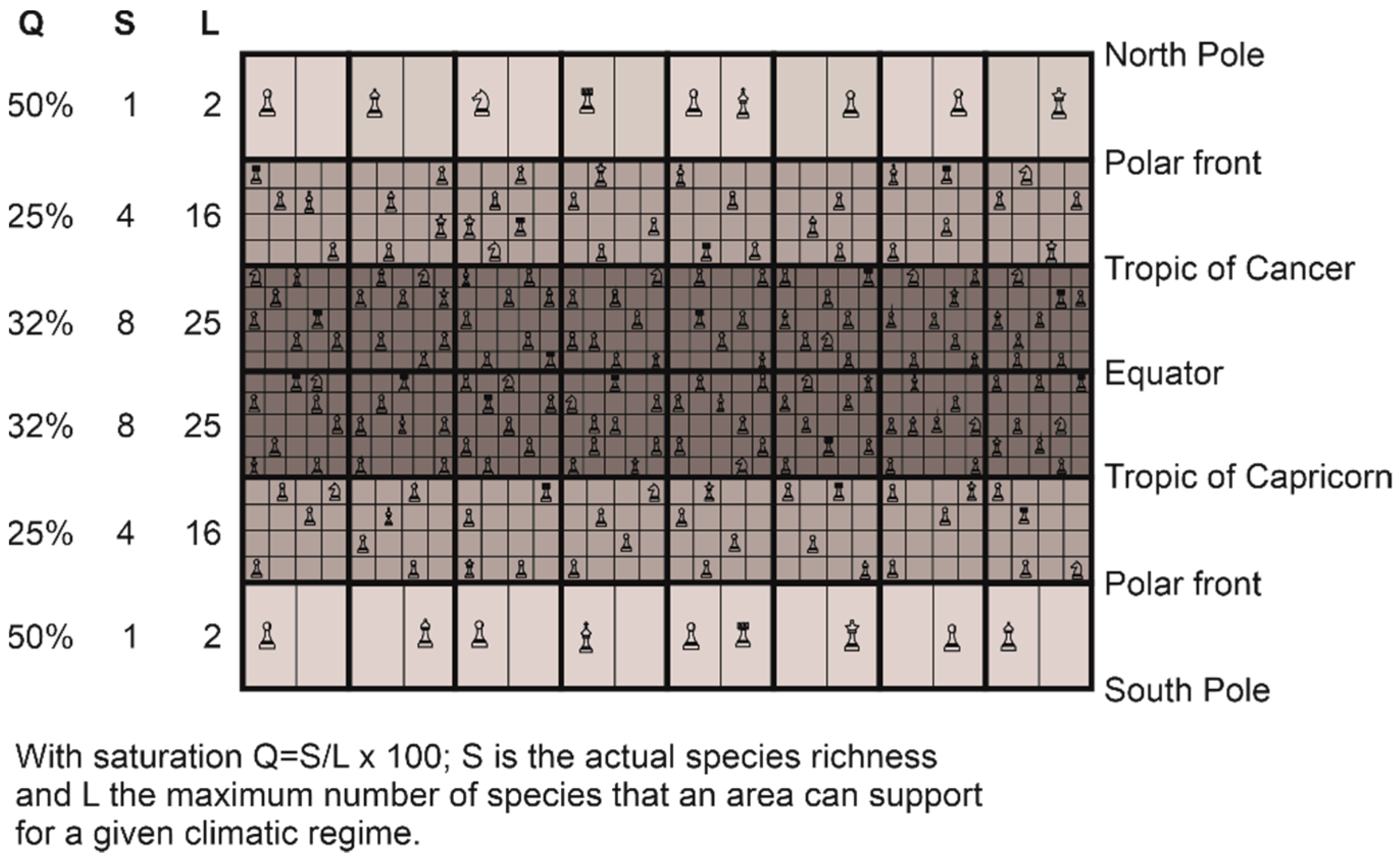
6. The Great Chessboard of Life
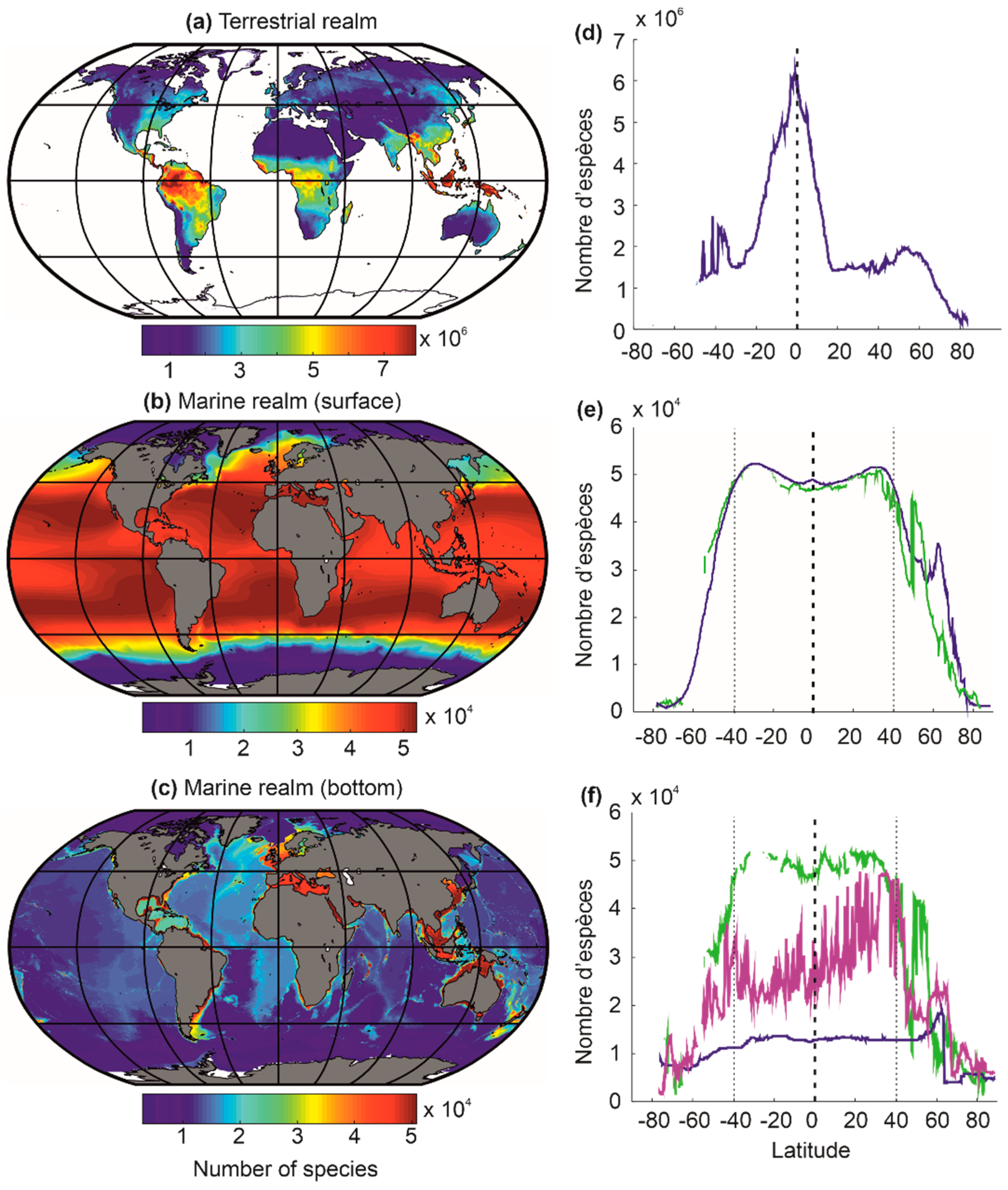
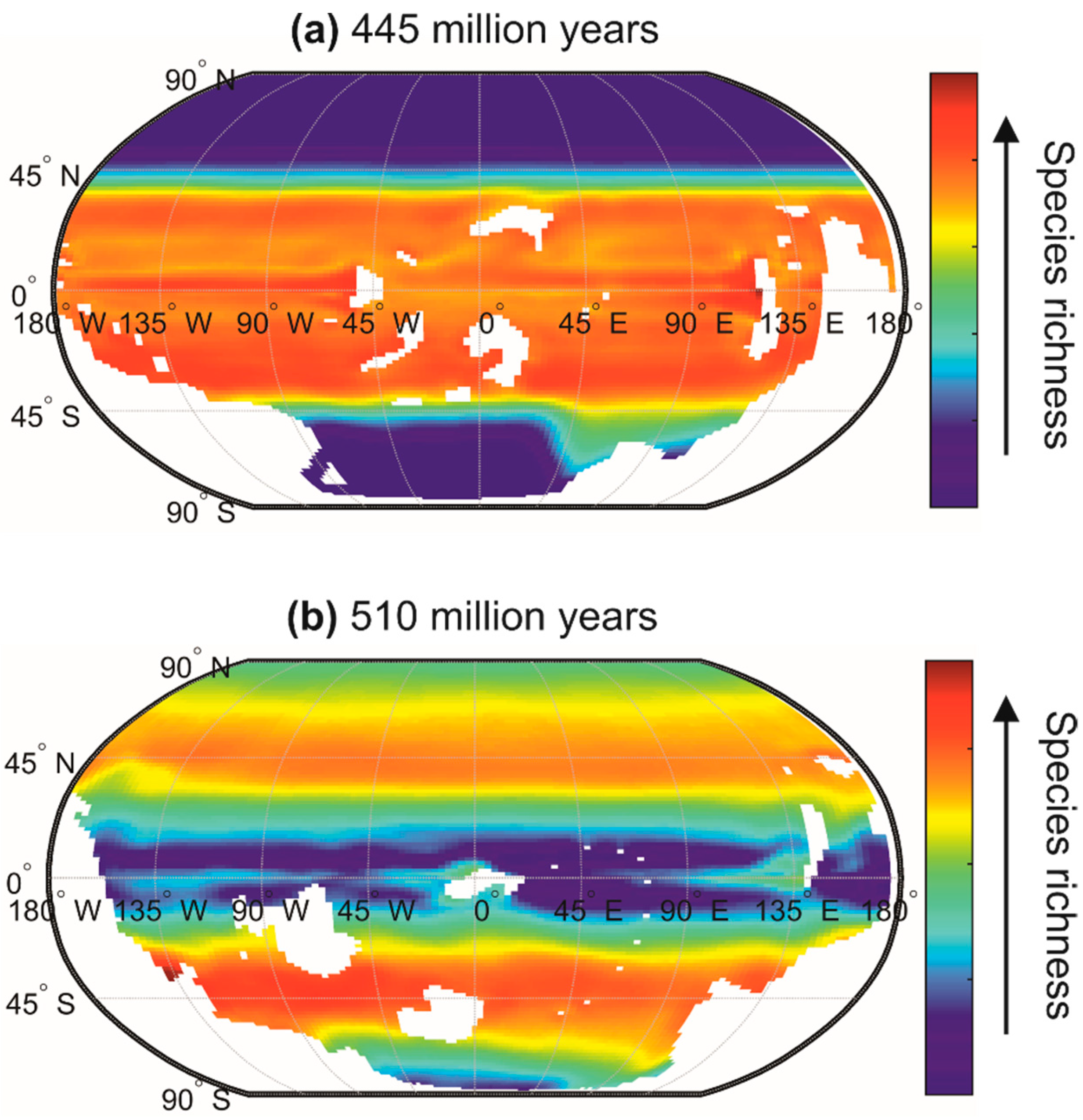
7. Differences in Latitudinal Biodiversity Gradients between the Terrestrial and the Marine Domains
8. Why Are There More Terrestrial Than Marine Species?
9. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purves, W.K.; Sadava, D.; Orians, G.H.; Heller, H.C. Life: The Science of Biology, 7th ed.; W. H. Freeman & Co.: New York, NY, USA, 2004; p. 1121. [Google Scholar]
- Calenbuhr, V. Complexity Science in the Context of Policymaking. In Science for Policy Handbook; Šucha, V., Sienkiewicz, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 118–127. [Google Scholar]
- Wolfa, Y.I.; Katsnelsonb, M.I.; Koonin, E.V. Physical foundations of biological complexity. Proc. Natl. Acad. Sci. USA 2018, 115, E8678–E8687. [Google Scholar] [CrossRef] [Green Version]
- Wickramasinghe, N.C.; Wickramasinghe, D.T.; Tout, C.A.; Lattanzio, J.C.; Steele, E.J. Cosmic biology in perspective. Astrophys. Space Sci. 2019, 364, 205. [Google Scholar] [CrossRef] [Green Version]
- Kump, L.R.; Kasting, J.F.; Crane, R.G. The Earth System; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Gould, S.J. Punctuated Equilibrium; Harvard University Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Stearns, S.C. Life history evolution: Successes, limitations, and prospects. Naturwissenschaften 2000, 87, 476–486. [Google Scholar] [CrossRef]
- Gould, S.J. Ever Since Darwin (Reflexions in Natural History); W.W. Norton and Company: New York, NY, USA, 1977. [Google Scholar]
- Jacob, F. Evolution and tinkering. Science 1977, 196, 1161–1166. [Google Scholar] [CrossRef] [Green Version]
- Monod, J. Chance and Necessity: An Essay on the Natural Philosophy of Modern Biology; Vintage Books: New York, NY, USA, 1971. [Google Scholar]
- Darwin, C. The Origin of Species by Means of Natural Selection; John Murray: London, UK, 1859. [Google Scholar]
- Dobzhansky, T. Nothing in biology makes sense except in the light of evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Kimura, M. The Neutral Theory of Molecular Evolution; Cambridge University Press: Cambridge, UK, 1983; p. 272. [Google Scholar]
- Mayr, E. Systematics and the Origin of Species from the Viewpoint of a Zoologist; Columbia University Press: New York, NY, USA, 1942. [Google Scholar]
- Müller, G.B. Evo–devo: Extending the evolutionary synthesis. Nat. Rev. Genet. 2007, 8, 943–949. [Google Scholar] [CrossRef]
- Pennisi, E. What determines species diversity? Science 2005, 309, 90. [Google Scholar] [CrossRef] [Green Version]
- Beaugrand, G. Marine Biodiversity, Climatic Variability and Global Change; Routledge: London, UK, 2015; p. 474. [Google Scholar]
- Naeem, S.; Loreau, M.; Inchausti, P. Biodiversity and ecosystem functioning: The emergence of a synthetic ecological framework. In Biodiversity and Ecosystem Functioning—Synthesis and Perspectives; loreau, M., Naeem, S., Inchausti, P., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 3–11. [Google Scholar]
- Tilman, D. The ecological consequences of changes in biodiversity: A search for general principles. Ecology 1999, 80, 1455–1474. [Google Scholar] [CrossRef]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of biodiversity loss on ocean ecosystem services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Beaugrand, G. Theoretical basis for predicting climate-induced abrupt shifts in the oceans. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20130264. [Google Scholar] [CrossRef] [Green Version]
- Beaugrand, G.; Mackas, D.; Goberville, E. Applying the concept of the ecological niche and a macroecological approach to understand how climate influences zooplankton: Advantages, assumptions, limitations and requirements. Prog. Oceanogr. 2013, 111, 75–90. [Google Scholar] [CrossRef]
- Beaugrand, G.; Goberville, E.; Luczak, C.; Kirby, R.R. Marine biological shifts and climate. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133350. [Google Scholar] [CrossRef]
- Beaugrand, G.; Kirby, R.R. Quasi-deterministic responses of marine species to climate change. Clim. Res. 2016, 69, 117–128. [Google Scholar] [CrossRef]
- Beaugrand, G.; Balembois, A.; Kléparski, L.; Kirby, R.R. Addressing the dichotomy of fishing and climate in fishery management with the FishClim model. Commun. Biol. 2022, 5, 1146. [Google Scholar] [CrossRef]
- Beaugrand, G.; Kirby, R.R. How do marine pelagic species respond to climate change? Theories and observations. Annu. Rev. Mar. Sci. 2018, 10, 169–197. [Google Scholar] [CrossRef]
- Beaugrand, G.; Edwards, M.; Raybaud, V.; Goberville, E.; Kirby, R.R. Future vulnerability of marine biodiversity compared with contemporary and past changes. Nat. Clim. Change 2015, 5, 695–701. [Google Scholar] [CrossRef]
- Beaugrand, G.; Conversi, A.; Atkinson, A.; Cloern, J.; Chiba, S.; Fonda-Unami, S.; Kirby, R.R.; Greene, C.G.; Goberville, E.; Otto, S.A.; et al. Prediction of unprecedented biological shifts in the global ocean. Nat. Clim. Change 2019, 9, 237–243. [Google Scholar] [CrossRef]
- Beaugrand, G.; Kirby, R.R.; Goberville, E. The mathematical influence on global patterns of biodiversity. Ecol. Evol. 2020, 10, 6494–6511. [Google Scholar] [CrossRef]
- Beaugrand, G.; Luczak, C.; Goberville, E.; Kirby, R.R. Marine biodiversity and the chessboard of life. PLoS ONE 2018, 13, e0194006. [Google Scholar] [CrossRef] [Green Version]
- Hellweger, F.L.; Van Sebille, E.; Fredrick, N.D. Biogeographic patterns in ocean microbes emerge in a neutral agent-based model. Science 2014, 345, 1346–1349. [Google Scholar] [CrossRef]
- Lladó Fernández, S.; Větrovský, T.; Baldrian, P. The concept of operational taxonomic units revisited: Genomes of bacteria that are regarded as closely related are often highly dissimilar. Folia Microbiol. 2018, 64, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Cohan, F.M.; Koeppel, A.F. The origins of ecological diversity in prokaryotes. Curr. Biol. 2008, 18, R1024–R1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suen, G.; Goldman, B.S.; Welch, R.D. Predicting prokaryotic eological niches using genome sequence analysis. PLoS ONE 2007, 2, e743. [Google Scholar] [CrossRef] [PubMed]
- Hughes Martiny, J.B.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef]
- Lundberg, P.; Ranta, E.; Ripa, J.; Kaitala, V. Population variability in space and time. Trends Ecol. Evol. 2000, 15, 460–464. [Google Scholar] [CrossRef]
- Russell, F.S. A summary of the observations of the occurence of planktonic stages of fish off Plymouth 1924–1972. J. Mar. Biol. Assoc. U. K. 1973, 53, 347–355. [Google Scholar] [CrossRef]
- Edwards, M.; Beaugrand, G.; Helaouët, P.; Alheit, J.; Coombs, S.H. Marine ecosystem response to the Atlantic Multidecadal Oscillation. PLoS ONE 2013, 8, e57212. [Google Scholar] [CrossRef] [Green Version]
- Alheit, J.; Hagen, E. Long-term forcing of European herring and sardine populations. Fish. Oceanogr. 1997, 6, 130–139. [Google Scholar] [CrossRef]
- Poole, K.G. A review of the Canada Lynx, Lynx canadensis, in Canada. Can. Field-Nat. 2003, 117, 360–376. [Google Scholar] [CrossRef] [Green Version]
- Lomolino, M.V.; Riddle, B.R.; Brown, J.H. Biogeography, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2006; p. 845. [Google Scholar]
- Ricklefs, R.E. Evolutionary diversification, coevolution between populations and their antagonists, and the filling of niche space. Proc. Natl. Acad. Sci. USA 2010, 107, 1265–1272. [Google Scholar] [CrossRef] [Green Version]
- van der Spoel, S. A biosystematic basis for pelagic biodiversity. Bijdr. Tot De Dierkd. 1994, 64, 3–31. [Google Scholar] [CrossRef] [Green Version]
- Angel, M.V. Pelagic biodiversity. In Marine Biodiversity: Patterns and Processes; Ormond, R.F.G., Gage, J.D., Angel, M.V., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 35–68. [Google Scholar]
- Rex, M.A.; Etter, R.J.; Stuart, C.T. Large-scale patterns of species diversity in the deep-sea benthos. In Marine Biodiversity: Patterns and Processes; Ormond, R.F.G., Gage, J.D., Angel, M.V., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 94–121. [Google Scholar]
- Whittaker, R.H. Communities and Ecosystems, 2nd ed.; Macmillan: New York, NY, USA, 1975. [Google Scholar]
- Beaugrand, G.; Ibañez, F.; Lindley, J.A. An overview of statistical methods applied to the CPR data. Prog. Oceanogr. 2003, 58, 235–262. [Google Scholar] [CrossRef]
- Faillettaz, R.; Beaugrand, G.; Goberville, E.; Kirby, R.R. Atlantic Multidecadal Oscillations drive the basin-scale distribution of Atlantic bluefin tuna. Sci. Adv. 2019, 5, eaar6993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caracciolo, M.; Beaugrand, G.; Hélaouët, P.; Gevaert, F.; Edwards, M.; Lizon, F.; Kléparski, L.; Goberville, E. Annual phytoplankton succession results from niche-environment interaction. J. Plankton Res. 2021, 43, 85–102. [Google Scholar] [CrossRef]
- Kléparski, L.; Beaugrand, G.; Edwards, M.; Schmitt, F.G.; Kirby, R.R.; Breton, E.; Gevaert, F.; Maniez, E. Morphological traits, niche-environment interaction and temporal changes in diatoms. Prog. Oceanogr. 2022, 201, 102747. [Google Scholar] [CrossRef]
- Levin, S.A. The problem of pattern and scale in ecology. Ecology 1992, 73, 1943–1967. [Google Scholar] [CrossRef]
- Haury, L.R.; McGowan, J.A.; Wiebe, P.H. Patterns and processes in the time-space scales of plankton distributions. In Spatial Pattern in Plankton Communities; Steele, J.H., Ed.; Plenun Press: New York, NY, USA, 1978; pp. 277–327. [Google Scholar]
- Wiens, J.A. Spatial scaling in ecology. Funct. Ecol. 1989, 1989, 385–397. [Google Scholar] [CrossRef]
- Hutchinson, G.E. The Ecological Theater and the Evolutionary Play; Yale University Press: New Haven, CT, USA, 1965; p. 172. [Google Scholar]
- Bachelard, G. L’activité Rationaliste de la Physique Contemporaine, 2nd ed.; Presses Universitaires de France: Paris, France, 1965; p. 223. [Google Scholar]
- Zacaï, A.; Monnet, C.; Pohl, A.; Beaugrand, G.; Mullins, G.; Kröck, D.; Servais, T. Truncated bimodal latitudinal diversity gradient in early Paleozoic phytoplankton. Sci. Adv. 2021, 7, eabd6709. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Hutchinson, G.E. An Introduction to Population Ecology; Yale University Press: New Haven, CT, USA, 1978; p. 260. [Google Scholar]
- Frontier, S.; Pichot-Viale, D.; Leprêtre, A.; Davoult, D.; Luczak, C. Ecosystèmes. Structure, Fonctionnement et Évolution, 3rd ed.; Dunod: Paris, France, 2004; p. 549. [Google Scholar]
- Morin, E. Introduction à la Pensée Complexe; Editions du Seuil: Paris, France, 2005; p. 158. [Google Scholar]
- Araújo, M.B.; Peterson, A.T. Uses and misuses of bioclimatic envelope modelling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Sillero, N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Model. 2011, 222, 1343–1346. [Google Scholar] [CrossRef]
- Goberville, E.; Beaugrand, G.; Hautekeete, N.-C.; Piquot, Y.; Luczak, C. Uncertainties in species distribution projections and general circulation models. Ecol. Evol. 2015, 5, 1100–1116. [Google Scholar] [CrossRef] [PubMed]
- Raybaud, V.; Beaugrand, G.; Goberville, E.; Delebecq, G.; Destombe, C.; Valero, M.; Davoult, D.; Morin, P.; Gevaert, F. Decline in Kelp in West Europe and Climate. PLoS ONE 2013, 8, e66044. [Google Scholar] [CrossRef] [Green Version]
- Rombouts, I.; Beaugrand, G.; Dauvin, J.-C. Potential changes in benthic macrofaunal distributions from the English Channel simulated under climate change scenarios. Estuar. Coast. Shelf Sci. 2012, 99, 153–161. [Google Scholar] [CrossRef]
- Schickele, A.; Goberville, E.; Leroy, B.; Beaugrand, G.; Hattab, T.; Francour, P.; Raybaud, V. Redistribution of small pelagic fish in Europe and Climate Change. Fish Fish. 2021, 22, 212–225. [Google Scholar] [CrossRef]
- Peterson, A.T.; Cobos, M.E.; Jiménez-García, D. Major challenges for correlational ecological niche model projections to future climate conditions: Climate change, ecological niche models, and uncertainty. Ann. N. Y. Acad. Sci. 2018, 1429, 66–77. [Google Scholar] [CrossRef]
- Ehrlén, J.; Morris, W.F. Predicting changes in the distribution and abundance of species under environmental change. Ecol. Lett. 2015, 18, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Helaouët, P.; Beaugrand, G. Physiology, ecological niches and species distribution. Ecosystems 2009, 12, 1235–1245. [Google Scholar] [CrossRef]
- Pauly, D. Ecosystem impacts of the world’s marine fisheries. Glob. Chang. Newsl. 2003, 55, 21–23. [Google Scholar]
- Jennings, S.; Kaiser, M.J.; Reynolds, J.D. Marine Fisheries Ecology; Blackwell Science Ltd.: Oxford, UK, 2001; p. 417. [Google Scholar]
- Koch, G.W.; Vitousek, P.M.; Steffen, W.L.; Walker, B.H. Terrestrial transects for global change research. Vegetatio 1992, 121, 53–65. [Google Scholar] [CrossRef]
- Raven, P.H.; Johnson, G.B.; Mason, K.A.; Losos, J.B.; Singer, S.R. Biologie; Deboeck Supérieur: Louvain-la-Neuve, Belgium, 2017; p. 1282. [Google Scholar]
- Costello, M.J.; May, R.M.; Stork, N.E. Can we name Earth’s species before they go extinct? Science 2013, 339, 413–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, C.; Tittensor, D.P.; Adl, S.; Simpson, A.G.B.; Worm, B. How Many Species Are There on Earth and in the Ocean? PLoS Biol. 2011, 9, e1001127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appeltans, W.; Ahyong, S.T.; Anderson, G.; Angel, M.V.; Artois, T.; Bailly, N.; Bamber, R.; Barber, A.; Bartsch, I.; Berta, A.; et al. The magnitude of global marine species biodiversity. Curr. Biol. 2012, 22, 2189–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erwin, T.L. Tropical forests: Their richness in Coleoptera and other arthropod species. Coleopt. Bull. 1982, 36, 74–75. [Google Scholar]
- May, R.M. How Many Species are There on Earth? Science 1988, 241, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Thackeray, S.J.; Henrys, P.A.; Hemming, D.; Bell, J.R.; Botham, M.S.; Burthe, S.; Helaouet, P.; Johns, D.G.; Jones, I.D.; Leech, D.I.; et al. Phenological sensitivity to climate across taxa and trophic levels. Nature 2016, 535, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Parmesan, C.; Matthews, J. Biological impacts of climate change. In Principles of Concervation Biology; Groom, M.J., Meffe, G.K., Carroll, C.R., Eds.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2006; pp. 333–360. [Google Scholar]
- Beaugrand, G.; Brander, K.M.; Lindley, J.A.; Souissi, S.; Reid, P.C. Plankton effect on cod recruitment in the North Sea. Nature 2003, 426, 661–664. [Google Scholar] [CrossRef]
- Beaugrand, G.; Reid, P.C.; Ibañez, F.; Lindley, J.A.; Edwards, M. Reorganisation of North Atlantic marine copepod biodiversity and climate. Science 2002, 296, 1692–1694. [Google Scholar] [CrossRef] [Green Version]
- Post, E.; Peterson, R.O.; Stenseth, N.C.; McLaren, B.E. Ecosystem consequences of wolf behavioural response to climate. Nature 1999, 1999, 905–907. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Brown, C.J.; Sydeman, W.J.; Kiessling, W.; Schoeman, D.S.; Moore, P.J.; Brander, K.; Bruno, J.F.; Buckley, L.B.; Burrows, M.T.; et al. Global imprint of climate change on marine life. Nat. Clim. Change 2013, 3, 919–925. [Google Scholar] [CrossRef]
- Kennedy, D.; Norman, C. What don’t we know? Science 2005, 309, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tittensor, D.T.; Mora, C.; Jetz, W.; Lotze, H.K.; Ricard, D.; Berghe, E.V.; Worm, B. Global patterns and predictors of marine biodiversity across taxa. Nature 2010, 466, 1098–1101. [Google Scholar] [CrossRef]
- Rex, M.A.; Stuart, C.T.; Hessler, R.R.; Allen, R.R.; Sanders, H.L.; Wilson, G.D.F. Global-scale latitudinal patterns of species diversity in the deep-sea benthos. Nature 1993, 365, 636–639. [Google Scholar] [CrossRef]
- Pianka, E.R. Latitudinal gradients in species diversity: A review of concepts. Am. Nat. 1966, 100, 33–46. [Google Scholar] [CrossRef]
- Roy, K.; Jablonski, D.; Valentine, J.W.; Rosenberg, G. Marine latitudinal diversty gradients: Tests of causal hypotheses. Proc. Natl. Acad. Sci. USA 1998, 95, 3699–3702. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Diniz-Filho, J.A.F.; Bini, L.M.; Araujo, M.B.; Field, R.; Hortal, J.; Kerr, J.T.; Rahbek, C.; Rodriguez, M.A.; Sanders, N.J. Metabolic theory and diversity gradients: Where do we go from here? Ecology 2007, 88, 1898–1902. [Google Scholar] [CrossRef]
- Currie, D.J. Energy and large-scale patterns of animal- and plant- species richness. Am. Nat. 1991, 137, 27–49. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal gradients in species diversity: The search for the primary cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, M.L.; Sandlin, E.A. Species diversity and latitudes: Listening to area’s signal. Oikos 1997, 80, 172–176. [Google Scholar] [CrossRef]
- Colwell, R.K.; Hurtt, G.C. Nonbiological gradients in species richness and a spurious rapoport effect. Am. Nat. 1994, 144, 570–595. [Google Scholar] [CrossRef]
- Allen, A.P.; Brown, J.H.; Gillooly, J.F. Global biodiversity, biochemical kinetics, and the enrgetic-equivalence rule. Science 2002, 297, 1545–1548. [Google Scholar] [CrossRef] [PubMed]
- Hubbell, S.P. The Unified Neutral Theoy of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Beaugrand, G.; Rombouts, I.; Kirby, R.R. Towards an understanding of the pattern of biodiversity in the oceans. Glob. Ecol. Biogeogr. 2013, 22, 440–449. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, Y.-G.; Zhang, C.-Y.; Wang, T.-R.; Su, T.-H.; Huang, P.-H.; Meng, H.-H.; Li, J. Latitudinal Diversity Gradient in the Changing World: Retrospectives and Perspectives. Diversity 2022, 14, 334. [Google Scholar] [CrossRef]
- Willig, M.R.; Presley, S.J. Latitudinal Gradients of Biodiversity: Theory and Empirical Patterns; Elsevier: Oxford, UK, 2018; Volume 3. [Google Scholar]
- Muenchow, J.; Dieker, P.; Kluge, J.; Kessler, M.; von Wehrden, H. A review of ecological gradient research in the Tropics: Identifying research gaps, future directions, and conservation priorities. Biodivers. Conserv. 2018, 27, 273–285. [Google Scholar] [CrossRef]
- Hawkins, B.A. Ecology’s oldest pattern? Trends Ecol. Evol. 2001, 16, 470. [Google Scholar] [CrossRef]
- Brown, J.H. Why are there so many species in the tropics? J. Biogeogr. 2014, 41, 8–22. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H. Two decades of homage to Santa Rosalia: Toward a general theory of diversity. Am. Zool. 1981, 21, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, G.E. Homage to Santa Rosalia, or why are there so many kinds of animals? Am. Nat. 1959, 93, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E. A comprehensive framework for global patterns in biodiversity. Ecol. Lett. 2004, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Benton, M.J. Origins of Biodiversity. PLoS Biol. 2016, 14, e2000724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995; p. 436. [Google Scholar]
- Colwell, R.K.; Lees, D.C. The mid-domain effect: Geometric constraints on the geography of species richness. Trends Ecol. Evol. 2000, 15, 70–76. [Google Scholar] [CrossRef]
- Colwell, R.K.; Gotelli, N.J.; Rahbek, C.; Entsminger, G.L.; Farrell, C.; Graves, G.R. Peaks, plateaus, canyons, and craters: The complex geometry of simple mid-domain effect models. Evol. Ecol. Res. 2009, 11, 355–370. [Google Scholar]
- Simpson, G.G. Species density of North American recent mammals. Syst. Zool. 1964, 13, 57–73. [Google Scholar] [CrossRef]
- Jablonski, D.; Roy, K.; Valentine, J.W. Out of the Tropics: Evolutionary dynamics of the latitudinal diversity gradient. Science 2006, 314, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Dowle, E.J.; Morgan-Richards, M.; Trewick, S.A. Molecular evolution and the latitudinal biodiversity gradient. Heredity 2013, 110, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Crame, J.A. Pattern and proceses in marine biogeography: A view from the poles. In Fontiers of Biogeography I: New Directions in the Geography of Nature; Lomolino, M.V., Heaney, L.R., Eds.; Sinauer Associates: Sunderland, MA, USA, 2004; pp. 272–292. [Google Scholar]
- Economo, E.P.; Narula, N.; Friedman, N.R.; Weiser, M.D.; Guénard, B. Macroecology and macroevolution of the latitudinal diversity gradient in ants. Nat. Commun. 2018, 9, 1778. [Google Scholar] [CrossRef] [Green Version]
- Moreau, C.S.; Bell, C.D. Testing the museum versus cradle tropical biological diversity hypothesis: Phylogeny, diversification, and ancestral biogeographic range evolution of the ants. Evolution 2013, 67, 2240–2257. [Google Scholar] [CrossRef]
- Fine, P.V.A. Ecological and evolutionary drivers of geographic variation in species diversity. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 369–392. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, T.; O’Meara, B.C.; Beaulieu, J.M. Retiring “Cradles” and “Museums” of Biodiversity. Am. Nat. 2022, 199, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Scholl, J.P.; Wiens, J.J. Diversification rates and species richness across the tree of life. Proc. R. Soc. B 2016, 283, 20161334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillooly, J.; Allen, A.P.; West, G.B.; Brown, J.H. The rate of DNA evolution: Effects of body size and temperature on the molecular clock. Proc. Natl. Acad. Sci. USA 2005, 102, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Davies, T.J.; Barraclough, T.G.; Savolainen, V.; Chase, M.W. Environmental causes for plant biodiversity gradients. Philos. Trans. R. Soc. B 2004, 359, 1645–1656. [Google Scholar] [CrossRef]
- Connell, J.H.; Orias, E. The ecological regulation of species diversity. Am. Nat. 1964, 98, 399–414. [Google Scholar] [CrossRef]
- Schonbeck, M.W.; Norton, T.A. Factors controlling the upper limits of fucoid algae on the shore. J. Exp. Mar. Biol. Ecol. 1978, 31, 303–313. [Google Scholar] [CrossRef]
- Sousa, W.P. The role of disturbance in natural communities. Annu. Rev. Ecol. Evol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Stein, A.; Kreft, H. Terminology and quantification of environmental heterogeneity in species-richness research. Biol. Rev. 2015, 90, 815–836. [Google Scholar] [CrossRef]
- Hortal, J.; Triantis, K.A.; Meiri, S.; Thébault, E.; Sfenthourakis, S. Island species richness increases with habitat diversity. Am. Nat. 2009, 174, E205–E217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.R.G.; Hawkins, B.A. The global biodiversity gradient. In Frontiers of Biogeography: New Directions in the Geography of Nature; Lomolino, M.V., Heaney, L.R., Eds.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2004; pp. 171–190. [Google Scholar]
- Emerson, B.C.; Kolm, N. Species diversity can drive speciation. Nature 2005, 434, 1015–1017. [Google Scholar] [CrossRef]
- Erwin, D.H. Macroevolution of ecosystem engineering, niche construction and diversity. Trends Ecol. Evol. 2008, 23, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Erwin, D.H. Seeds of diversity. Science 2005, 308, 1752–1753. [Google Scholar] [CrossRef] [PubMed]
- Cadena, C.D.; Ricklefs, R.E.; Jiménez, I.; Bermingham, E. Is speciation driven by species diversity? Nature 2005, 438, E1–E2. [Google Scholar] [CrossRef] [PubMed]
- Emerson, B.C.; Kolm, N. Emerson & Kolm reply. Nature 2005, 438, E2. [Google Scholar]
- Stehli, F.G.; Douglas, R.G.; Newell, R.G. Generation and maintenance of gradients in taxonomic diversity. Science 1969, 164, 947–949. [Google Scholar] [CrossRef]
- Rombouts, I.; Beaugrand, G.; Ibañez, F.; Gasparini, S.; Chiba, S.; Legendre, L. Global latitudinal variations in marine copepod diversity and environmental factors. Proc. R. Soc. B 2009, 276, 3053–3062. [Google Scholar] [CrossRef] [Green Version]
- Romdal, T.S.; Araújo, M.B.; Rahbek, C. Life on a tropical planet: Niche conservatism and the global diversity gradient. Glob. Ecol. Biogeogr. 2013, 22, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Crisp, M.D.; Cook, L.G. Phylogenetic niche conservatism: What are the underlying evolutionary and ecological causes? New Phytol. 2012, 196, 681–694. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ecthoterms. Proc. R. Soc. B 2011, 278, 1823–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaugrand, G.; Ibañez, F.; Lindley, J.A. Geographical distribution and seasonal and diel changes of the diversity of calanoid copepods in the North Atlantic and North Sea. Mar. Ecol. Prog. Ser. 2001, 219, 205–219. [Google Scholar] [CrossRef]
- Jablonski, D.; Huang, S.; Roy, K.; Valentine, J.W. Shaping the latitudinal diversity gradient: New perspectives from a synthesis of paleobiology and biogeography. Am. Nat. 2017, 189, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brayard, A.; Escarguel, G.; Bucher, H. Latitudinal gradient of taxonomic richness: Combined outcome of temperature and geographic mid-domains effects? J. Zool. Syst. Evol. Res. 2005, 43, 178–188. [Google Scholar] [CrossRef]
- Kléparski, L.; Beaugrand, G. The species chromatogram, a new graphical method to represent, characterise and compare the ecological niches of different species. Ecol. Evol. 2022, 12, e8830. [Google Scholar] [CrossRef]
- Speck, L. Caractérisation De La Biodiversité Marine à L’aide De La Théorie METAL; Université du Littoral Côte d’Opale: Wimereux, France, 2022; p. 22. [Google Scholar]
- Goberville, E.; Hautekèete, N.-C.; Kirby, R.R.; Piquot, Y.; Luczak, C.; Beaugrand, G. Climate change and the ash dieback crisis. Sci. Rep. 2016, 6, 35303. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.H. The Ecological Implications of Body Size; Cambridge University Press: Cambridge, UK, 1983; p. 329. [Google Scholar]
- Damuth, J. Population density and body size in mammals. Nature 1981, 290, 699–700. [Google Scholar] [CrossRef]
- Damuth, J. Of size and abundance. Nature 1991, 351, 268–269. [Google Scholar] [CrossRef]
- Brown, J.H. On the relationship between abundance and distribution of species. Am. Nat. 1984, 124, 255–279. [Google Scholar] [CrossRef]
- Norris, R.D.; Hull, P.M. The temporal dimension of marine speciation. Evol. Ecol. 2012, 26, 393–415. [Google Scholar] [CrossRef]
- Briggs, J.C. Modes of speciation: Marine indo-west Pacific. Bull. Mar. Sci. 1999, 65, 645–656. [Google Scholar]
- Pierrot-Bults, A.C.; Van der Spoel, S. Speciation in macrozooplankton. In Zoogeography and Diversity of Plankton; Van der Spoel, S., Pierrot-Bults, A.C., Eds.; John Wiley: New York, NY, USA, 1979; pp. 144–167. [Google Scholar]
- Momigliano, P.; Jokinen, H.; Fraimout, A.; Florin, A.-B.; Norkko, A.; Merilä, A. Extraordinarily rapid speciation in a marine fish. Proc. Natl. Acad. Sci. USA 2017, 114, 6074–6079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Visser, J. Transition zones and salp speciation. In Pelagic Biogeography; Pierrot-Bults, A.C., Van der Spoel, S., Zahuranec, B.J., Johnson, R.K., Eds.; Unesco: Paris, France, 1985; pp. 266–269. [Google Scholar]
- Knowlton, N. Sibling species in the sea. Annu. Rev. Ecol. Evol. Syst. 1993, 24, 189–216. [Google Scholar] [CrossRef]
- Gause, G.F. The Struggle for Coexistence; Williams and Wilkins: Baltimore, MD, USA, 1934. [Google Scholar]
- Rabosky, D.L.; Hurlbert, A.H. Species richness at continental sclaes is dominated by ecological limits. Am. Nat. 2015, 185, 572–583. [Google Scholar] [CrossRef]
- McManus, M.A.; Woodson, C.B. Plankton distribution and ocean dispersal. J. Exp. Biol. 2012, 215, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Sexton, P.F.; Norris, R.D. Dispersal and biogeography of marine plankton: Long-distance dispersal of the foraminifer Truncorotalia truncatulinoides. Geology 2008, 36, 899–902. [Google Scholar] [CrossRef]
- Machac, A.; Zrzavy, J.; Smrckova, J.; Storch, D. Temperature dependence of evolutionary diversification: Differences between two contrasting model taxa support the metabolic theory of ecology. J. Evol. Biol. 2012, 25, 2449–2456. [Google Scholar] [CrossRef]
- Rohde, K. Nonequilibrium Ecology; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Benton, M.J. Diversification and extinction in the history of life. Science 1995, 268, 52–58. [Google Scholar] [CrossRef]
- Morlon, H. Phylogenetic approaches for studying diversification. Ecol. Lett. 2014, 17, 508–525. [Google Scholar] [CrossRef]
- Berta, A.; Adam, P. The evolutionary biology of pinnipeds. In Secondary Adaptation of Tetrapods to Life in the Water; de Buffrenil, V., Mazin, J.-M., Eds.; Verlag Dr Frederich Pfeil: Munchen, Germany, 2001; pp. 235–260. [Google Scholar]
- Wiens, J.J.; Donoghue, M.J. Historical biogeography, ecology and species richness. Trends Ecol. Evol. 2004, 19, 639–644. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Porter, E.E. Does herbivore diversity depend on plant diversity? The case of California butterflies. Am. Nat. 2003, 161, 40–49. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Robinson, P.J.; Henderson-Sellers, A. Contemporary Climatology; Pearson Education Limited: Edinburgh, UK, 1999; p. 315. [Google Scholar]
- Bonan, G. Ecological Climatology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2008; p. 550. [Google Scholar]
- Mannion, P.D.; Upchurch, P.; Benson, R.B.J.; Goswami, A. The latitudinal biodiversity gradient through deep time. Trends Ecol. Evol. 2014, 29, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, R.M. Biological diversity: Differences between land and sea. Philos. Trans. R. Soc. Lond. B 1994, 343, 105–111. [Google Scholar]
- Rozhnov, S.V. Appearance and evolution of marine benthic communities in the early Palaeozoic. Paleontol. J. 2005, 40, S444–S452. [Google Scholar] [CrossRef]
- May, R.M. Conceptual aspects of the quantification of the extent of biological diversity. Philos. Trans. R. Soc. Lond. B 1994, 345, 13–20. [Google Scholar]
- Benton, M.J. Biodiversity on land and in the sea. Geol. J. 2001, 36, 211–230. [Google Scholar] [CrossRef]
- Román-Palacios, C.; Moraga-López, D.; Wiens, J.J. The origins of global biodiversity on land, sea and freshwater. Ecol. Lett. 2022, 25, 1376–1386. [Google Scholar] [CrossRef]
- Pisani, D.; Poling, L.L.; Lyons-Weiler, M.; Hedges, S.B. The colonization of land by animals: Molecular phylogeny and divergence times among arthropods. BMC Biol. 2004, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Grassle, C.F.; Maciolek, N.J. Deep-sea species richness: Regional and local diversity estimates from quantitative bottom samples. Am. Nat. 1992, 139, 313–341. [Google Scholar] [CrossRef]
- Snelgrove, P.V.R. Getting to the bottom of marine biodiversity: Sedimentary habitats. Bioscience 1999, 49, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Kléparski, L.; Beaugrand, G.; Kirby, R.R. How do plankton species coexist in an apparently unstructured environment? Biol. Lett. 2022, 18, 20220207. [Google Scholar] [CrossRef] [PubMed]
- Briggs, J.C. Global Biogeography; Elsevier: Amsterdam, The Netherland, 1995; Volume 14. [Google Scholar]
- Swanborn, D.J.B.; Huvenne, V.A.I.; Pittman, S.J.; Rogers, A.D.; Taylor, M.L.; Woodall, L.C. Mapping, quantifying and comparing seascape heterogeneity of Southwest Indian Ridge seamounts. Landsc. Ecol. 2023, 38, 185–203. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beaugrand, G. Towards an Understanding of Large-Scale Biodiversity Patterns on Land and in the Sea. Biology 2023, 12, 339. https://doi.org/10.3390/biology12030339
Beaugrand G. Towards an Understanding of Large-Scale Biodiversity Patterns on Land and in the Sea. Biology. 2023; 12(3):339. https://doi.org/10.3390/biology12030339
Chicago/Turabian StyleBeaugrand, Grégory. 2023. "Towards an Understanding of Large-Scale Biodiversity Patterns on Land and in the Sea" Biology 12, no. 3: 339. https://doi.org/10.3390/biology12030339
APA StyleBeaugrand, G. (2023). Towards an Understanding of Large-Scale Biodiversity Patterns on Land and in the Sea. Biology, 12(3), 339. https://doi.org/10.3390/biology12030339