

Transcriptome Analysis of the Immortal Human Keratinocyte HaCaT Cell Line Damaged by Tritiated Water

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line Culture
2.2. Cell Viability Assay
2.3. Radiation Treatment
2.4. RNA Extraction
2.5. cDNA Library Preparation and Sequencing
2.6. Sequencing Data Filtering and Reference Genome Comparisons
2.7. Bioinformatics Analysis
2.8. Western Blot Verification
2.9. Statistical Analysis
3. Results
3.1. Tritiated Water Impaired the Viability of HacaT Cell
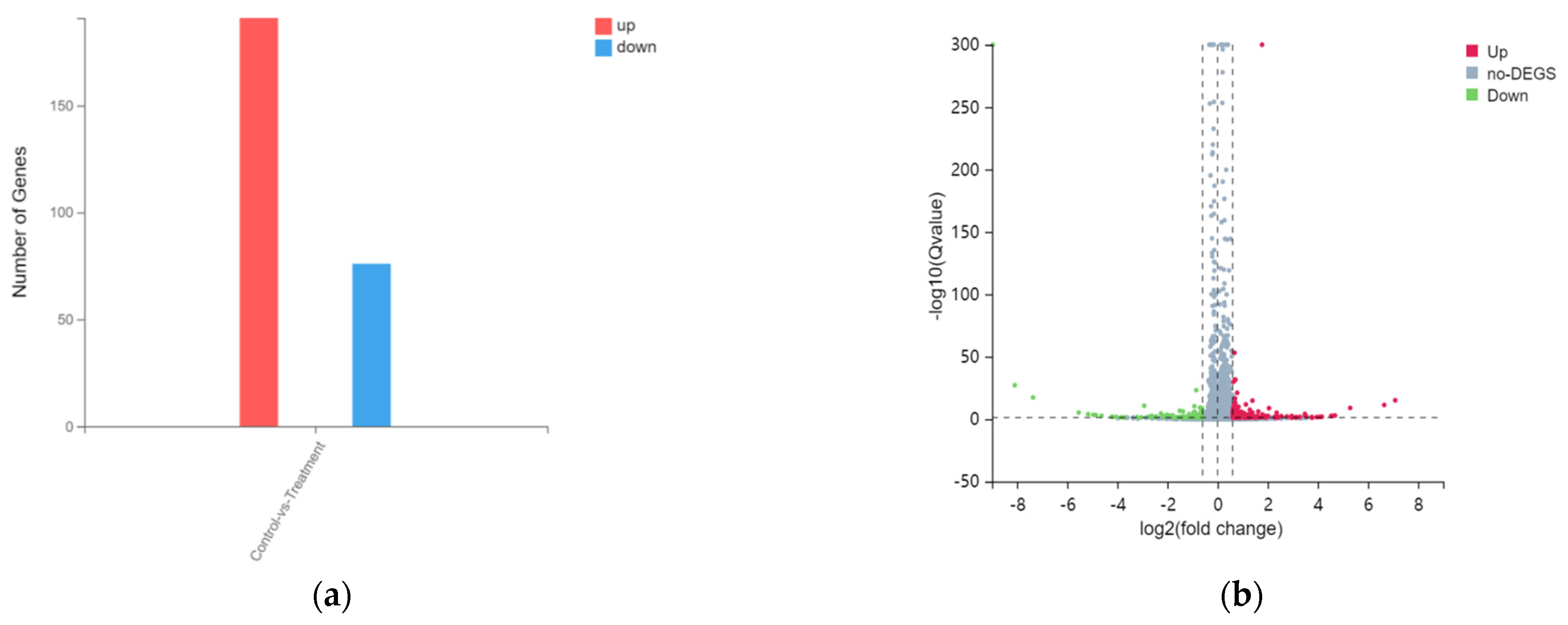
3.2. Overview on Transcriptome Sequencing Data Quality and Difference Analysis of HacaT Cells Exposed to Tritiated Water
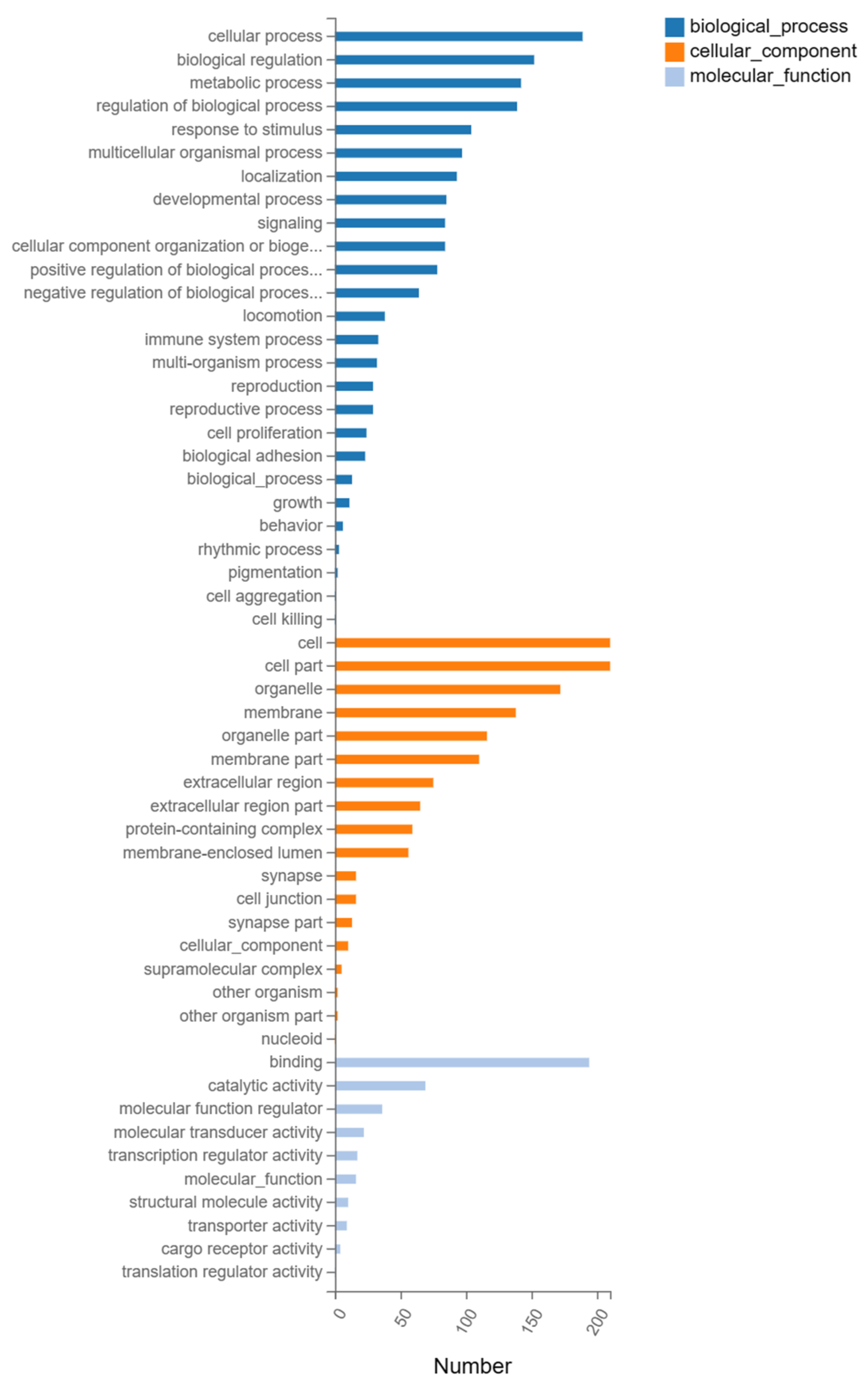
3.3. GO Enrichment Analysis of DEGs
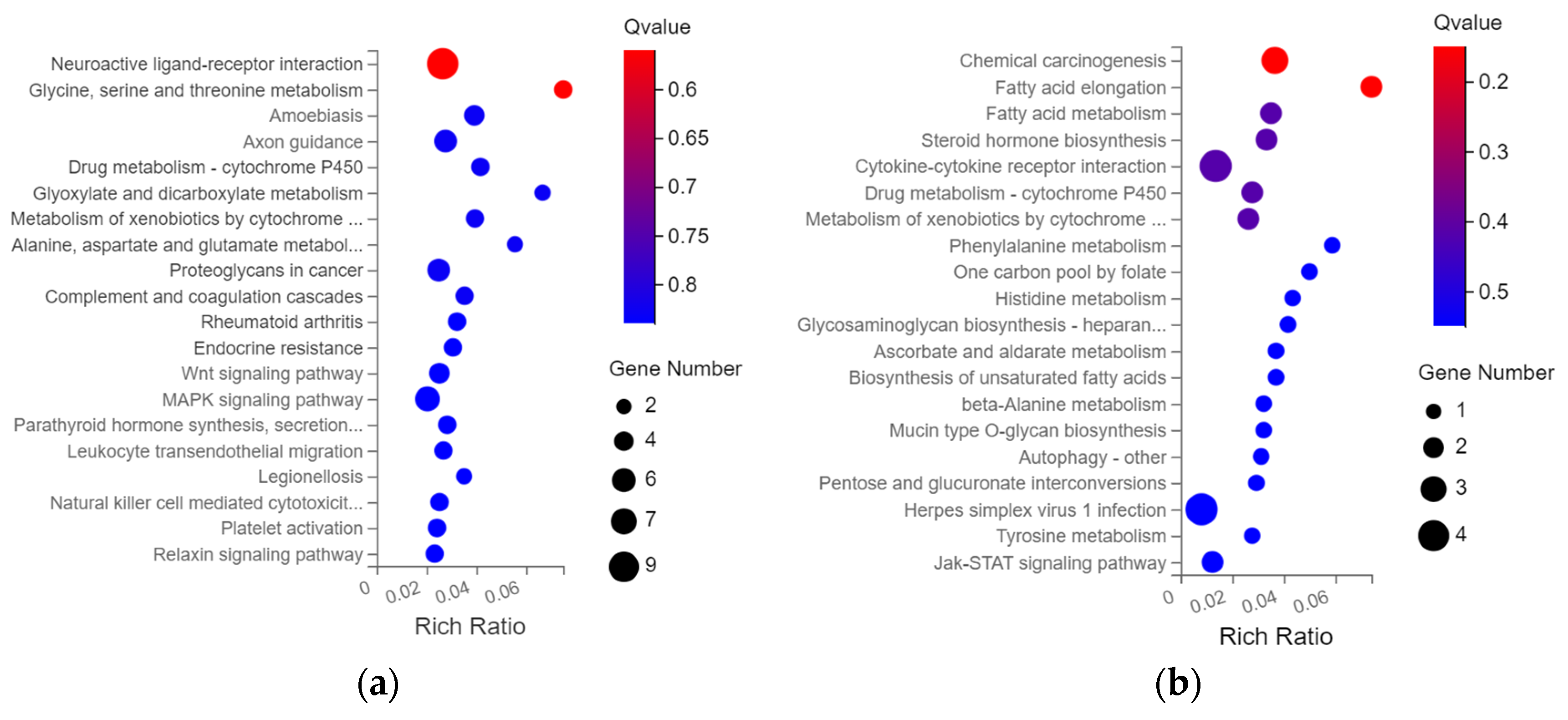
3.4. KEGG Pathway Enrichment Analysis
3.5. Tritiated Water Induced Protein Expression Differences in HacaT Cell
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bailly, B.P.; Dumas, F.; Voiseux, C.; Morillon, M.; Oms, P.E.; Solier, L. Dissolved Radiotracers and Numerical Modeling in North European Continental Shelf Dispersion Studies (1982–2016): Databases, Methods and Applications. Water 2020, 12, 1667. [Google Scholar] [CrossRef]
- Povinec, P.P.; Aoyama, M.; Biddulph, D.; Breier, R.; Buesseler, K.; Chang, C.C.; Golser, R.; Hou, X.L.; Jeskovsky, M.; Jull, A.J.T.; et al. Cesium, iodine and tritium in NW Pacific waters—A comparison of the Fukushima impact with global fallout. Biogeosciences 2013, 10, 5481–5496. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.B.; Shultz, C.; Stuart, M.; Festarin, A. Tritium uptake in rainbow trout (Oncorhynchus mykiss): HTO and OBT-spiked feed exposures simultaneously. Appl. Radiat. Isot. 2015, 98, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Alloni, D.; Cutaia, C.; Mariotti, L.; Friedland, W.; Ottolenghi, A. Modeling dose deposition and DNA damage due to low-energy β(–) emitters. Radiat. Res. 2014, 182, 322–330. [Google Scholar] [CrossRef]
- Yang, H.Y. Safety and Protection of Tritium; Atomic Energy Press: Beijing, China, 1997. [Google Scholar]
- Audette-Stuart, M.; Kim, S.; McMullin, D.; Festarini, A.; Yankovich, T.; Carr, J.; Mulpuru, S. Adaptive response in frogs chronically exposed to low doses of ionizing radiation in the environment. J. Environ. Radioact. 2011, 102, 566–573. [Google Scholar] [CrossRef]
- Arcanjo, C.; Armant, O.; Floriani, M.; Cavalie, I.; Camilleri, V.; Simon, O.; Orjollet, D.; Adam-Guillermin, C.; Gagnaire, B. Tritiated water exposure disrupts myofibril structure and induces mis-regulation of eye opacity and DNA repair genes in zebrafish early life stages. Aquat. Toxicol. 2018, 200, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Stuart, M.; Festarini, A.; Schleicher, K.; Tan, E.; Kim, S.B.; Wen, K.; Gawlik, J.; Ulsh, B. Biological effects of tritium on fish cells in the concentration range of international drinking water standards. Int. J. Radiat. Biol. 2016, 92, 563–571. [Google Scholar] [CrossRef]
- Tawn, E.J.; Curwen, G.B.; Riddell, A.E. Chromosome aberrations in workers occupationally exposed to tritium. J. Radiol. Prot. 2018, 38, N9–N16. [Google Scholar] [CrossRef]
- Yin, H.; Bhattacharjee, D.; Roy, G.; Fujimoto, N.; Nakatani, T.; Ito, A. Tumorigenesis in Infant C3H/HeN Mice Exposed to Tritiated Water (HTO). J. Radiat. Res. 2002, 43, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Seyama, T.; Yamamoto, O.; Kinomura, A.; Yokoro, K. Carcinogenic Effects of Tritiated Water (HTO) in Mice: In Comparison to those of Neutrons and Gamma-rays. J. Radiat. Res. 1991, 32, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Balonov, M.I.; Muksinova, K.N.; Mushkacheva, G.S. Tritium Radiobiological Effects in Mammals. Health Phys. 1993, 65, 713–726. [Google Scholar] [CrossRef]
- Cui, F.M.; Liu, L.; Zheng, L.L.; Bao, G.L.; Tu, Y.; Sun, L.; He, Y.M. The Role of miR-34a in Tritiated Water Toxicity in Human Umbilical Vein Endothelial Cells. Dose-Response 2016, 14, 155932581663858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.B.; Liu, Y.T.; Li, Z.Y.; Wu, Z.J.; Zhang, M.; Xue, P.J.; Chen, Q. Tritiated Water Induces Toxicity in Human Umbilical Vein Vascular Endothelial Cells via IL8. Dose-Response 2020, 18, 155932582093854. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, C.S.; Choi, S.I.; Kim, R.K.; Kim, J.Y.; Nam, S.Y.; Kim, I.G. Tissue distribution, excretion and effects on genotoxicity of tritium following oral administration to rats. Nucl. Eng. Technol. 2018, 51, 303–309. [Google Scholar] [CrossRef]
- Nowosielska, E.M.; Cheda, A.; Zdanowski, R.; Lewicki, S.; Scott, B.R.; Janiak, M.K. Effect of internal contamination with tritiated water on the neoplastic colonies in the lungs, innate anti-tumour reactions, cytokine profile, and haematopoietic system in radioresistant and radiosensitive mice. Radiat. Environ. Biophys. 2018, 57, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yin, Y.; Liu, J.; Lu, B.; Wan, H.; Yang, L.; Wang, W.; Li, R. Hydrogen—Rich water attenuates the radiotoxicity induced by tritium exposure in vitro and in vivo. J. Radiat. Res. 2021, 62, 34–45. [Google Scholar] [CrossRef]
- Kamiguchi, Y.; Tateno, H.; Mikamo, K. Dose—Response relationship for the induction of structural chromosome aberrations in human spermatozoa after in vitro exposure to tritium beta-rays. Mutat. Res. 1990, 228, 125–131. [Google Scholar] [CrossRef]
- Wang, B.; Zhou, X.Y. Effects of prenatal exposure to low-dose beta radiation from tritiated water on the neurobehavior of mice. J. Radiat. Res. 1995, 36, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Selivanova, M.A.; Mogilnaya, O.A.; Badun, G.A.; Vydryakova, G.A.; Kuznetsov, A.M.; Kudryasheva, N.S. Effect of tritium on luminous marine bacteria and enzyme reactions. J. Environ. Radioact. 2013, 120, 19–25. [Google Scholar] [CrossRef]
- Kim, S.B.; Shultz, C.; Stuart, M.; McNamara, E.; Festarini, A.; Bureau, D.P. Organically bound tritium (OBT) formation in rainbow trout (Oncorhynchus mykiss): HTO and OBT-spiked food exposure experiments. Applied radiation and isotopes: Including data, instrumentation and methods for use in agriculture. Ind. Med. 2013, 72, 114–122. [Google Scholar] [CrossRef]
- Réty, C.; Gilbin, R.; Gomez, E. Induction of reactive oxygen species and algal growth inhibition by tritiated water with or without copper. Environ. Toxicol. 2010, 27, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Jaeschke, B.C.; Bradshaw, C. Bioaccumulation of tritiated water in phytoplankton and trophic transfer of organically bound tritium to the blue mussel, Mytilus edulis. J. Environ. Radioact. 2013, 115, 28–33. [Google Scholar] [CrossRef]
- Quan, Y.; Lin, J.; Deng, B. The response of human mesenchymal stem cells to internal exposure to tritium β-rays. J. Radiat. Res. 2019, 60, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorob’eva, N.Y.; Kochetkov, O.A.; Pustovalova, M.V.; Grekhova, A.K.; Blokhina, T.M.; Yashkina, E.I.; Osipov, A.A.; Kabanov, D.I.; Surin, P.P.; Barchukov, V.G.; et al. Comparative Analysis of the Formation of γH2AX Foci in Human Mesenchymal Stem Cells Exposed to 3H-Thymidine, Tritium Oxide, and X-Rays Irradiation. Bull. Exp. Biol. Med. 2018, 166, 178–181. [Google Scholar] [CrossRef]
- Qiu, J.; Cai, E.; Wu, C.P.; Wang, M.M. Effects of Tritium β Irradiation on the Expression of Regulated Factors Associated with the Neural Cell Migration. Radiat. Prot. 2012, 32, 204–209. [Google Scholar]
- Zhou, X.; Gao, W.; Wang, B. A review of experimental studies on biological effects of tritium exposure. Zhonghua Yi Xue Za Zhi 1999, 79, 493–496. [Google Scholar]
- Little, J.B.; Nove, J. Sensitivity of human diploid fibroblast cell strains from various genetic disorders to acute and protracted radiation exposure. Radiat. Res. 1990, 123, 87–92. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA–Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Ajami, S.; Dadras, S.; Faghih, Z.; Shobeiri, S.S.; Mahdian, A. In Vitro evaluation of immediate cytotoxicity of resterilised orthodontic bands on HGF-1 cell line. Int. Orthod. 2021, 19, 500–504. [Google Scholar] [CrossRef]
- Gu, H.; Sun, Y.H.; Li, X.Z. Novel rRNA-depletion methods for total RNA sequencing and ribosome profiling developed for avian species. Poult. Sci. 2021, 100, 101321. [Google Scholar] [CrossRef]
- Cock, P.J.; Fields, C.J.; Goto, N.; Heuer, M.L.; Rice, P.M. The Sanger FASTQ file format for sequences with quality scores, and the Solexa/Illumina FASTQ variants. Nucleic Acids Res. 2010, 38, 1767–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, G.F.; Granotier, C.; Barroca, V.; Laurent, D.; Boussin, F.D.; Lewandowski, D.; Saintigny, Y.; Romeo, P.H. Tritium contamination of hematopoietic stem cells alters long—Term hematopoietic reconstitution. Int. J. Radiat. Biol. 2011, 87, 556–570. [Google Scholar] [CrossRef]
- Jha, A.N.; Dogra, Y.; Turner, A.; Millward, G.E. Impact of low doses of tritium on the marine mussel, Mytilus edulis: Genotoxic effects and tissue-specific bioconcentration. Mutat. Res. 2005, 586, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Gagnaire, B.; Arcanjo, C.; Cavalié, I.; Camilleri, V.; Simon, O.; Floriani, M.; Orjollet, D.; Adam-Guillermin, C. Tritiated Water Exposure in Zebrafish (Danio rerio): Effects on the Early-Life Stages. Environ. Toxicol. Chem. 2020, 39, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Krokosz, A.; Lichota, A.; Nowak, K.E.; Grebowski, J. Carbon nanoparticles as possible radioprotectors in biological systems. Radiat. Phys. Chem. 2016, 128, 143–150. [Google Scholar] [CrossRef]
- Deng, B.; Cheng, Q.; Du, Y.; Yang, Y. Cytogenetic Effects of Low Dose Tritiated Water in Human Peripheral Blood Lymphocytes. Chin. J. Radiol. Health 2016, 25, 6–10+14. [Google Scholar]
- Kaneski, C.R.; Hanover, J.A.; Schueler Hoffman, U.H. Generation of GLA-knockout human embryonic stem cell lines to model peripheral neuropathy in Fabry disease. Mol. Genet. Metab. Rep. 2022, 33, 100914. [Google Scholar] [CrossRef]
- Ebrahim, N.; El-Halim, H.E.A.; Helal, O.K.; El-Azab, N.E.; Badr, O.A.M.; Hassouna, A.; Saihati, H.A.A.; Aborayah, N.H.; Emam, H.T.; El-Wakeel, H.S.; et al. Effect of bone marrow mesenchymal stem cells-derived exosomes on diabetes-induced retinal injury: Implication of Wnt/ b-catenin signaling pathway. Biomed. Pharmacother. 2022, 154, 113554. [Google Scholar] [CrossRef]
- Jere, S.W.; Houreld, N.N. Regulatory Processes of the Canonical Wnt/β-Catenin Pathway and Photobiomodulation in Diabetic Wound Repair. Int. J. Mol. Sci. 2022, 23, 4210. [Google Scholar] [CrossRef]
- Nelson, A.L.; Fontana, G.; Miclau, E.; Rongstad, M.; Murphy, W.; Huard, J.; Ehrhart, N.; Bahney, C. Therapeutic approaches to activate the canonical Wnt pathway for bone regeneration. J. Tissue Eng. Regen. Med. 2022, 16, 961–976. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, X.J.; Xing, J. Signal Transduction Pathways of EMT Induced by TGF–β, SHH, and WNT and Their Crosstalks. J. Clin. Med. 2016, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Ghahhari, N.M.; Babashah, S. Interplay between microRNAs and Wnt/β-catenin signalling pathway regulates epithelial-mesenchymal transition in cancer. Eur. J. Cancer 2015, 51, 1638–1649. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, S.; Yang, R.; Wang, Z.; Zhang, W.; Liu, H.; Huang, Y. ROS Promote Hypoxia-Induced Keratinocyte Epithelial-Mesenchymal Transition by Inducing SOX2 Expression and Subsequent Activation of Wnt/β–Catenin. Oxidative Med. Cell. Longev. 2022, 2022, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Tan, Z.; Yang, Y.; Deng, B.; Mu, L. Prolonged effect associated with inflammatory response observed after exposure to low dose of tritium β–rays. Int. J. Radiat. Biol. 2020, 96, 972–979. [Google Scholar] [CrossRef]
- Lee, H.; Jeong, A.J.; Ye, S.K. Highlighted STAT3 as a potential drug target for cancer therapy. BMB Rep. 2019, 52, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Chen, C.; Zhang, Y.; Zhang, L.; Mei, Y.; Long, X.; Tan, R.; Liang, W.; Sun, L. Acitretin modulates HaCaT cells proliferation through STAT1- and STAT3- dependent signaling. Saudi Pharm. J. 2017, 25, 620–624. [Google Scholar] [CrossRef]
- Park, J.W.; Lee, H.S.; Lim, Y.; Paik, J.H.; Kwon, O.K.; Kim, J.H.; Paryanto, I.; Yunianto, P.; Choi, S.; Oh, S.R.; et al. Rhododendron album Blume extract inhibits TNF–α/IFN–γ–induced chemokine production via blockade of NF—κB and JAK/STAT activation in human epidermal keratinocytes. Int. J. Mol. Med. 2018, 41, 3642–3652. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Raw Reads (M) | Total Clean Reads (M) | Total Clean Bases (Gb) | Clean Reads Q20 (%) | Clean Reads Q30 (%) | Clean Reads Ratio (%) |
|---|---|---|---|---|---|---|
| DZ1 | 44.85 | 42.87 | 6.43 | 97.7 | 93.88 | 95.59 |
| DZ2 | 44.65 | 42.99 | 6.45 | 97.54 | 93.48 | 96.28 |
| DZ3 | 47.33 | 45.43 | 6.81 | 97.66 | 93.67 | 95.98 |
| GJL1 | 47.33 | 44.85 | 6.73 | 97.81 | 94.19 | 94.78 |
| GJL2 | 47.33 | 45.14 | 6.77 | 97.85 | 94.28 | 95.37 |
| GJL3 | 47.33 | 45.17 | 6.78 | 97.58 | 93.55 | 95.45 |
| Part | ID | Description | Gene Ratio | BgRatio | p Value | geneID |
|---|---|---|---|---|---|---|
| BP | GO:0009812 | flavonoid metabolic process | 2/10 | 15/18862 | 2.65 × 10−5 | 445329/54576 |
| BP | GO:0006805 | xenobiotic metabolic process | 2/10 | 120/18862 | 1.75 × 10−3 | 445329/54576 |
| BP | GO:0071466 | cellular response to xenobiotic stimulus | 2/10 | 125/18862 | 1.89 × 10−3 | 445329/54576 |
| CC | GO:0000782 | telomere cap complex | 1/11 | 13/19520 | 7.30 × 10−3 | 7015 |
| CC | GO:0034045 | phagophore assembly site membrane | 1/11 | 15/19520 | 8.42 × 10−3 | 8987 |
| CC | GO:0005697 | telomerase holoenzyme complex | 1/11 | 22/19520 | 1.23 × 10−2 | 7015 |
| MF | GO:0003964 | RNA−directed DNA polymerase activity | 1/11 | 12/18337 | 7.18 × 10−3 | 7015 |
| MF | GO:0001972 | retinoic acid binding | 1/11 | 20/18337 | 1.19 × 10−2 | 54576 |
| MF | GO:0070034 | telomerase RNA binding | 1/11 | 22/18337 | 1.31 × 10−2 | 7015 |
| MF | GO:0001223 | transcription coactivator binding | 1/11 | 26/18337 | 1.55 × 10−2 | 7015 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhou, Y.; Wu, H.; Yan, Z.; Chen, J.; Song, W. Transcriptome Analysis of the Immortal Human Keratinocyte HaCaT Cell Line Damaged by Tritiated Water. Biology 2023, 12, 405. https://doi.org/10.3390/biology12030405
Zhang Y, Zhou Y, Wu H, Yan Z, Chen J, Song W. Transcriptome Analysis of the Immortal Human Keratinocyte HaCaT Cell Line Damaged by Tritiated Water. Biology. 2023; 12(3):405. https://doi.org/10.3390/biology12030405
Chicago/Turabian StyleZhang, Yan, Yuanyuan Zhou, Hui Wu, Zhuna Yan, Jinwu Chen, and Wencheng Song. 2023. "Transcriptome Analysis of the Immortal Human Keratinocyte HaCaT Cell Line Damaged by Tritiated Water" Biology 12, no. 3: 405. https://doi.org/10.3390/biology12030405
APA StyleZhang, Y., Zhou, Y., Wu, H., Yan, Z., Chen, J., & Song, W. (2023). Transcriptome Analysis of the Immortal Human Keratinocyte HaCaT Cell Line Damaged by Tritiated Water. Biology, 12(3), 405. https://doi.org/10.3390/biology12030405