
Advances in Viral Aquatic Animal Disease Knowledge: The Molecular Methods’ Contribution


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Nucleocytoplasmic Large DNA Virus
2.1. Iridoviridae
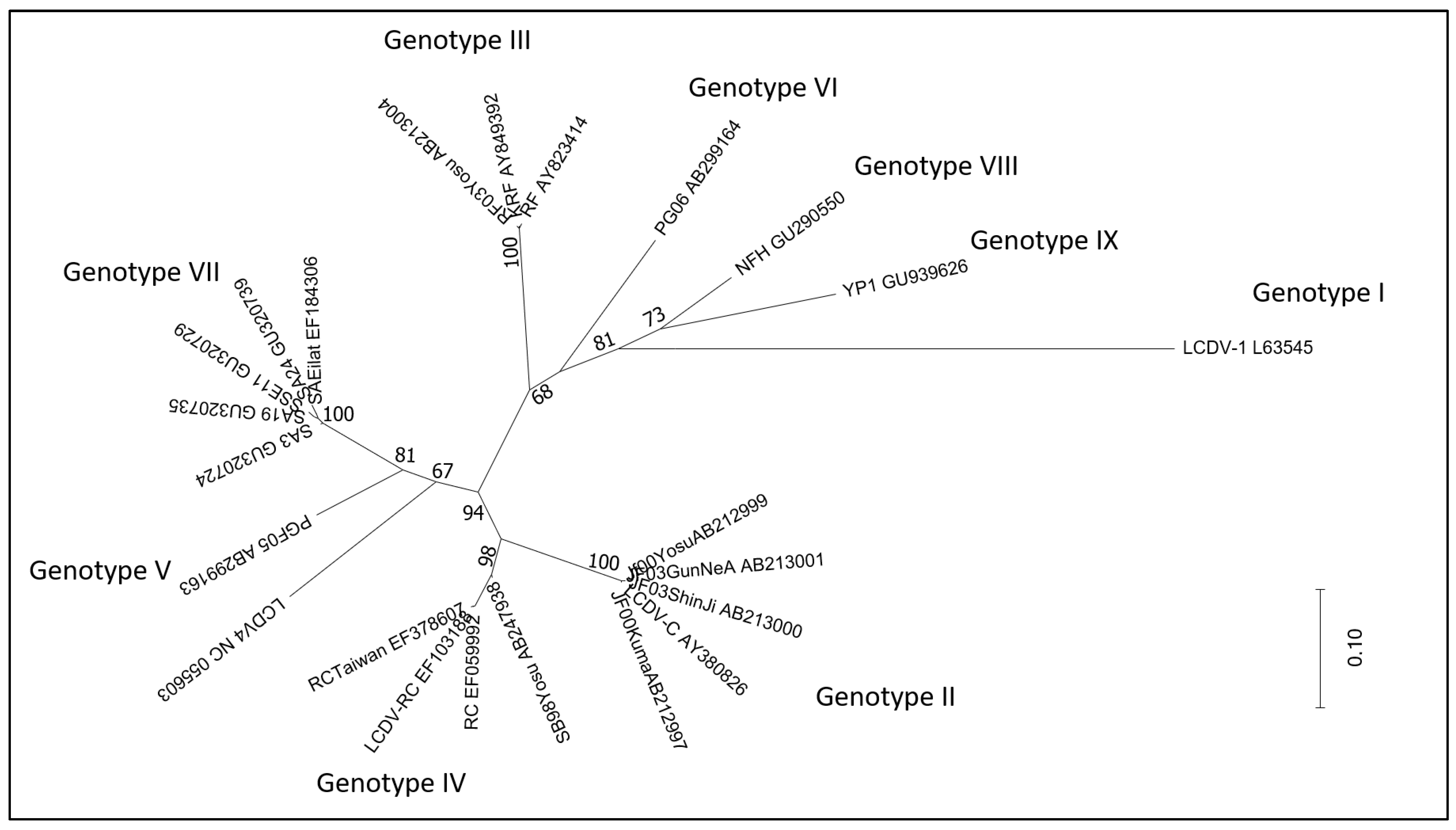
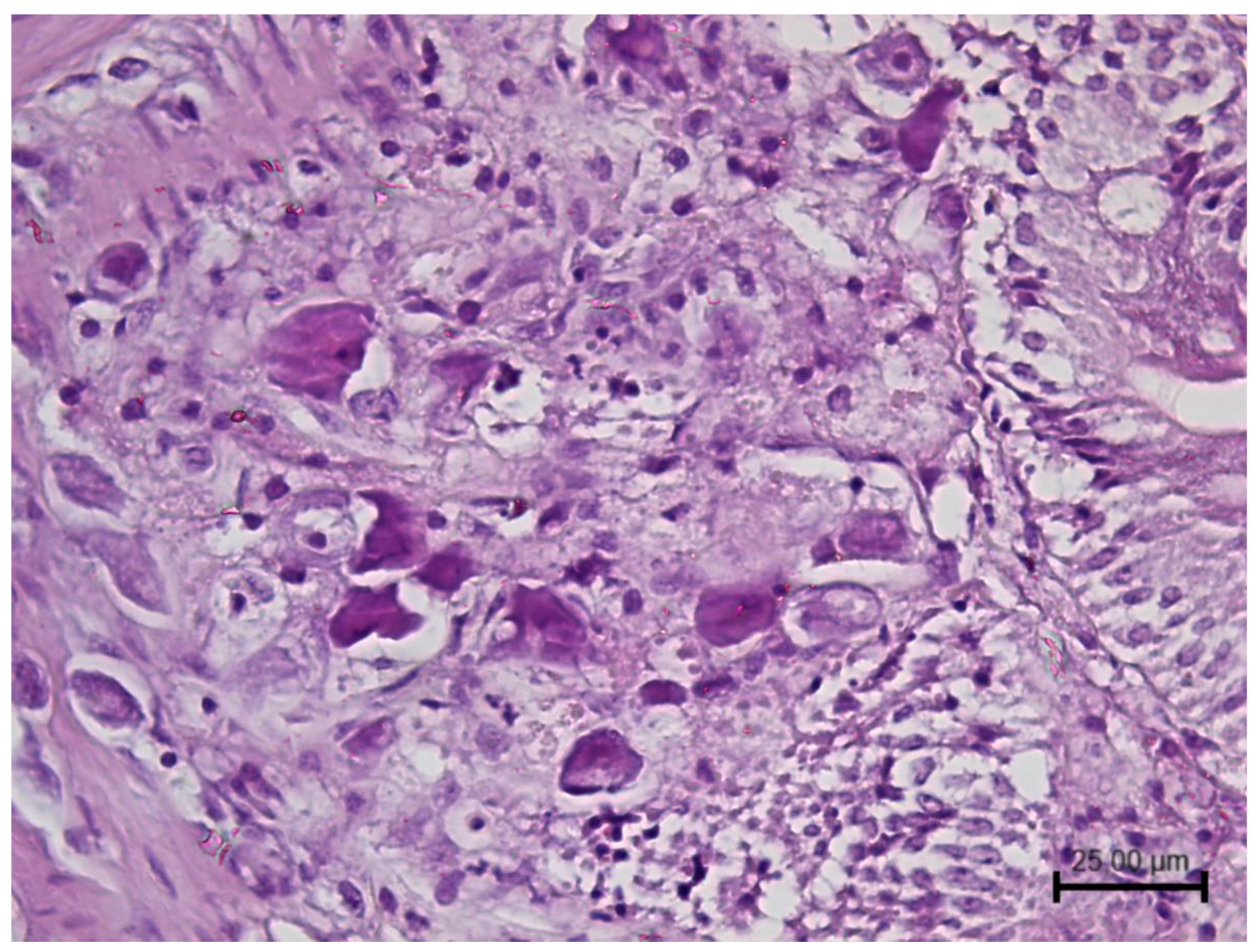
2.1.1. Lymphocystivirus
2.1.2. Megalocytivirus
2.2. Sturgeon NCLDVs
3. Herpesvirales
3.1. Alloherpesviridae
3.1.1. Cyprinivirus
3.1.2. Sturgeon Herpesviruses
3.1.3. Ictalurivirus
3.2. Malacoherpesviridae
4. Nodaviridae
Betanodavirus
5. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fonseca, A.; Laguardia-Nascimento, M.; Scotá Ferreira, A.; Pinto, C.; Pereira Freitas, T.; Rivetti Júnior, A.; Ferreira Homem, V.; Camargos, M. Detection of Megalocytivirus in Oreochromis Niloticus and Pseudoplatystoma Corruscans in Brazil. Dis. Aquat. Organ. 2022, 149, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, S.; Afzal, M.; Sikandar, S.; Afzal, I. Polymerase Chain Reaction. In Genetic Engineering—A Glimpse of Techniques and Applications; Jamal, F., Ed.; IntechOpen: London, UK, 2020; ISBN 978-1-78985-179-3. [Google Scholar]
- Jacquet, S.; Miki, T.; Noble, R.; Peduzzi, P.; Wilhelm, S. Viruses in Aquatic Ecosystems: Important Advancements of the Last 20 Years and Prospects for the Future in the Field of Microbial Oceanography and Limnology. Adv. Oceanogr. Limnol. 2010, 1, 97–141. [Google Scholar] [CrossRef]
- Pinheiro, A.C.A.S.; Volpe, E.; Principi, D.; Prosperi, S.; Ciulli, S. Development of a Multiplex RT-PCR Assay for Simultaneous Detection of the Major Viruses That Affect Rainbow Trout (Oncorhynchus Mykiss). Aquac. Int. 2016, 24, 115–125. [Google Scholar] [CrossRef]
- Errani, F.; Volpe, E.; Riera-Ferrer, E.; Caffara, M.; Padrós, F.; Gustinelli, A.; Fioravanti, M.; Ciulli, S. Development and Diagnostic Validation of a One-Step Multiplex RT-PCR Assay as a Rapid Method to Detect and Identify Nervous Necrosis Virus (NNV) and Its Variants Circulating in the Mediterranean. PLoS ONE 2022, 17, e0273802. [Google Scholar] [CrossRef] [PubMed]
- Kubista, M.; Andrade, J.M.; Bengtsson, M.; Forootan, A.; Jonák, J.; Lind, K.; Sindelka, R.; Sjöback, R.; Sjögreen, B.; Strömbom, L.; et al. The Real-Time Polymerase Chain Reaction. Mol. Asp. Med. 2006, 27, 95–125. [Google Scholar] [CrossRef]
- Ciulli, S.; Pinheiro, A.C.d.A.S.; Volpe, E.; Moscato, M.; Jung, T.S.; Galeotti, M.; Stellino, S.; Farneti, R.; Prosperi, S. Development and Application of a Real-Time PCR Assay for the Detection and Quantitation of Lymphocystis Disease Virus. J. Virol. Methods 2015, 213, 164–173. [Google Scholar] [CrossRef]
- Morley, A.A. Digital PCR: A Brief History. Biomol. Detect. Quantif. 2014, 1, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Zhang, Z.; Jing, H.; Zhang, M.; Wu, S.; Lin, X. Development of a Novel Droplet Digital PCR Assay for the Sensitive Detection of Carp Edema Virus. Aquaculture 2021, 545, 737162. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, F.; Li, Q.; Wang, L.; Fan, C. Isothermal Amplification of Nucleic Acids. Chem. Rev. 2015, 115, 12491–12545. [Google Scholar] [CrossRef]
- Yu, Q.; Liu, M.-Z.; Xiao, H.-H.; Yi, Y.; Cheng, H.; Putra, D.-F.; Li, S.-Q.; Li, P.-F. Selection and Characterization of Aptamers for Specific Detection of Iridovirus Disease in Cultured Hybrid Grouper (Epinephelus Fuscoguttatus♀ × E. Lanceolatus♂). Chin. J. Anal. Chem. 2020, 48, 650–661. [Google Scholar] [CrossRef]
- Pfankuche, V.M.; Hahn, K.; Bodewes, R.; Hansmann, F.; Habierski, A.; Haverkamp, A.-K.; Pfaender, S.; Walter, S.; Baechlein, C.; Postel, A.; et al. Comparison of Different In Situ Hybridization Techniques for the Detection of Various RNA and DNA Viruses. Viruses 2018, 10, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassidy, A.; Jones, J. Developments in in Situ Hybridisation. Methods S. Diego Calif. 2014, 70, 39–45. [Google Scholar] [CrossRef]
- Jansen, G.J.; Wiersma, M.; van Wamel, W.J.B.; Wijnberg, I.D. Direct Detection of SARS-CoV-2 Antisense and Sense Genomic RNA in Human Saliva by Semi-Autonomous Fluorescence in Situ Hybridization: A Proxy for Contagiousness? PLoS ONE 2021, 16, e0256378. [Google Scholar] [CrossRef]
- Arzul, I.; Nicolas, J.-L.; Davison, A.J.; Renault, T. French Scallops: A New Host for Ostreid Herpesvirus-1. Virology 2001, 290, 342–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE CHAPTER 2.4.9 Infection with Ostreid Herpesvirus 1 Microvariant. In Manual of Diagnostic Tests for Aquatic Animals; OIE: Paris, France, 2013.
- Snow, M. The Contribution of Molecular Epidemiology to the Understanding and Control of Viral Diseases of Salmonid Aquaculture. Vet. Res. 2011, 42, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tengs, T.; Rimstad, E. Emerging Pathogens in the Fish Farming Industry and Sequencing-Based Pathogen Discovery. Dev. Comp. Immunol. 2017, 75, 109–119. [Google Scholar] [CrossRef]
- Clouthier, S.; Anderson, E.; Kurath, G.; Breyta, R. Molecular Systematics of Sturgeon Nucleocytoplasmic Large DNA Viruses. Mol. Phylogenet. Evol. 2018, 128, 26–37. [Google Scholar] [CrossRef]
- Iyer, L.M.; Balaji, S.; Koonin, E.V.; Aravind, L. Evolutionary Genomics of Nucleo-Cytoplasmic Large DNA Viruses. Virus Res. 2006, 117, 156–184. [Google Scholar] [CrossRef] [Green Version]
- Chinchar, V.G.; Waltzek, T.B.; Subramaniam, K. Ranaviruses and Other Members of the Family Iridoviridae: Their Place in the Virosphere. Virology 2017, 511, 259–271. [Google Scholar] [CrossRef]
- Wolf, K.; Gravell, M.; Malsberger, R.G. Lymphocystis Virus: Isolation and Propagation in Centrarchid Fish Cell Lines. Science 1966, 151, 1004–1005. [Google Scholar] [CrossRef]
- Kurita, J.; Nakajima, K. Megalocytiviruses. Viruses 2012, 4, 521–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, M.; Miller, D.; Hoverman, J. Ecology and Pathology of Amphibian Ranaviruses. Dis. Aquat. Organ. 2009, 87, 243–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jancovich, J.K.; Chinchar, V.G.; Hyatt, A.D.; Miyazaki, T.; Williams, T.; Zhang, Q.Y. Family Iridoviridae. In Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 193–210. [Google Scholar]
- Volpatti, D.; Ciulli, S. Chapter 15 Lymphocystis Virus Disease. In Aquaculture Pathophysiology Volume I. Fish Diseases; Elsevier Academic Press: New York, NY, USA, 2022; pp. 201–216. [Google Scholar]
- Doszpoly, A.; Kaján, G.L.; Puentes, R.; Perretta, A. Complete Genome Sequence and Analysis of a Novel Lymphocystivirus Detected in Whitemouth Croaker (Micropogonias Furnieri): Lymphocystis Disease Virus 4. Arch. Virol. 2020, 165, 1215–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkaroun, J.; Bergmann, S.M.; Römer-Oberdörfer, A.; Demircan, M.D.; Tamer, C.; Kachh, G.R.; Weidmann, M. New Insights into Lymphocystis Disease Virus Genome Diversity. Viruses 2022, 14, 2741. [Google Scholar] [CrossRef]
- Mönttinen, H.A.M.; Bicep, C.; Williams, T.A.; Hirt, R.P. The Genomes of Nucleocytoplasmic Large DNA Viruses: Viral Evolution Writ Large. Microb. Genom. 2021, 7, 000649. [Google Scholar] [CrossRef]
- Kitamura, S.-I.; Jung, S.-J.; Kim, W.-S.; Nishizawa, T.; Yoshimizu, M.; Oh, M.-J. A New Genotype of Lymphocystivirus, LCDV-RF, from Lymphocystis Diseased Rockfish. Arch. Virol. 2006, 151, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Cano, I.; Valverde, E.J.; Lopez-Jimena, B.; Alonso, M.C.; Garcia-Rosado, E.; Sarasquete, C.; Borrego, J.J.; Castro, D. A New Genotype of Lymphocystivirus Isolated from Cultured Gilthead Seabream, Sparus Aurata L., and Senegalese Sole, Solea Senegalensis (Kaup). J. Fish Dis. 2010, 33, 695–700. [Google Scholar] [CrossRef]
- Shawky, M.; Taha, E.; Ahmed, B.; Mahmoud, M.A.; Abdelaziz, M.; Faisal, M.; Yousif, A. Initial Evidence That Gilthead Seabream (Sparus Aurata L.) Is a Host for Lymphocystis Disease Virus Genotype I. Animals 2021, 11, 3032. [Google Scholar] [CrossRef]
- Kvitt, H.; Heinisch, G.; Diamant, A. Detection and Phylogeny of Lymphocystivirus in Sea Bream Sparus Aurata Based on the DNA Polymerase Gene and Major Capsid Protein Sequences. Aquaculture 2008, 275, 58–63. [Google Scholar] [CrossRef]
- Wu, R.-H.; Tang, X.-Q.; Sheng, X.-Z.; Zhan, W.-B. Tissue Distribution of the 27.8 KDa Receptor and Its Dynamic Expression in Response to Lymphocystis Disease Virus Infection in Flounder (Paralichthys Olivaceus). J. Comp. Pathol. 2015, 153, 324–332. [Google Scholar] [CrossRef]
- Volpe, E.; Farneti, R.; Moscato, M.; Prosperi, S.; Ciulli, S. Distribuzione Di Lymphocystivirus in Organi Target e Non Target Di Orate (Sparus Aurata) Naturalmente Infette. Ittiopatologia 2015, 12, 129–137. Available online: https://core.ac.uk/display/226695907 (accessed on 2 February 2023).
- Cano, I.; Ferro, P.; Alonso, M.C.; Sarasquete, C.; Garcia-Rosado, E.; Borrego, J.J.; Castro, D. Application of in Situ Detection Techniques to Determine the Systemic Condition of Lymphocystis Disease Virus Infection in Cultured Gilt-Head Seabream, Sparus Aurata L. J. Fish Dis. 2009, 32, 143–150. [Google Scholar] [CrossRef]
- Valverde, E.J.; Cano, I.; Labella, A.; Borrego, J.J.; Castro, D. Application of a New Real-Time Polymerase Chain Reaction Assay for Surveillance Studies of Lymphocystis Disease Virus in Farmed Gilthead Seabream. BMC Vet. Res. 2016, 12, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Yue, Z.; Liu, H.; Liang, C.; Zheng, X.; Zhao, Y.; Chen, X.; Xiao, X.; Chen, C. Development and Evaluation of a Loop-Mediated Isothermal Amplification Assay for Rapid Detection of Lymphocystis Disease Virus. J. Virol. Methods 2010, 163, 378–384. [Google Scholar] [CrossRef]
- Valverde, E.J.; Borrego, J.J.; Sarasquete, M.C.; Ortiz-Delgado, J.B.; Castro, D. Target Organs for Lymphocystis Disease Virus Replication in Gilthead Seabream (Sparus Aurata). Vet. Res. 2017, 48, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, J.; Waltzek, T.; Subramaniam, K.; Yun, S.; Groff, J.; Anderson, I.; Chong, R.; Shirley, I.; Schuh, J.; Handlinger, J.; et al. Detection of Infectious Spleen and Kidney Necrosis Virus (ISKNV) and Turbot Reddish Body Iridovirus (TRBIV) from Archival Ornamental Fish Samples. Dis. Aquat. Organ. 2016, 122, 105–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waltzek, T.; Marty, G.; Alfaro, M.; Bennett, W.; Garver, K.; Haulena, M.; Weber ES, I.; Hedrick, R. Systemic Iridovirus from Threespine Stickleback Gasterosteus Aculeatus Represents a New Megalocytivirus Species (Family Iridoviridae). Dis. Aquat. Organ. 2012, 98, 41–56. [Google Scholar] [CrossRef] [Green Version]
- de Groof, A.; Guelen, L.; Deijs, M.; van der Wal, Y.; Miyata, M.; Ng, K.S.; van Grinsven, L.; Simmelink, B.; Biermann, Y.; Grisez, L.; et al. A Novel Virus Causes Scale Drop Disease in Lates Calcarifer. PLoS Pathog. 2015, 11, e1005074. [Google Scholar] [CrossRef] [Green Version]
- Halaly, M.A.; Subramaniam, K.; Koda, S.A.; Popov, V.L.; Stone, D.; Way, K.; Waltzek, T.B. Characterization of a Novel Megalocytivirus Isolated from European Chub (Squalius Cephalus). Viruses 2019, 11, 440. [Google Scholar] [CrossRef] [Green Version]
- Canuti, M.; Eis-Huebinger, A.M.; Deijs, M.; de Vries, M.; Drexler, J.F.; Oppong, S.K.; Müller, M.A.; Klose, S.M.; Wellinghausen, N.; Cottontail, V.M.; et al. Two Novel Parvoviruses in Frugivorous New and Old World Bats. PLoS ONE 2011, 6, e29140. [Google Scholar] [CrossRef]
- Tan, L.V.; Van Doorn, H.R.; Van der Hoek, L.; Minh Hien, V.; Jebbink, M.F.; Quang Ha, D.; Farrar, J.; Van Vinh Chau, N.; de Jong, M.D. Random PCR and Ultracentrifugation Increases Sensitivity and Throughput of VIDISCA for Screening of Pathogens in Clinical Specimens. J. Infect. Dev. Ctries. 2011, 5, 142–148. [Google Scholar] [CrossRef] [Green Version]
- van der Hoek, L.; Pyrc, K.; Jebbink, M.F.; Vermeulen-Oost, W.; Berkhout, R.J.M.; Wolthers, K.C.; Wertheim-van Dillen, P.M.E.; Kaandorp, J.; Spaargaren, J.; Berkhout, B. Identification of a New Human Coronavirus. Nat. Med. 2004, 10, 368–373. [Google Scholar] [CrossRef]
- Inouye, K.; Yamano, K.; Maeno, Y.; Nakajima, K.; Matsuoka, M.; Wada, Y.; Sorimachi, M. Iridovirus Infection of Cultured Red Sea Bream, Pagrus major. Fish Pathol. 1992, 27, 19–27. [Google Scholar] [CrossRef]
- Stephens, F.J.; Jones, J.B.; Hillier, P. Ornamental Fish Testing Project: Final Report; Government of Western Australia Department of Fisheries and Fisheries Research Division WA Marine Research Laboratories: Perth, WA, Australia, 2009.
- OIE CHAPTER 2.3.7 Red Sea Bream Iridoviral Disease. In Manual of Diagnostic Tests for Aquatic Animals; OIE: Paris, France, 2021; pp. 332–343.
- Kurita, J.; Nakajima, K.; Hirono, I.; Aoki, T. Polymerase Chain Reaction (PCR) Amplification of DNA of Red Sea Bream [Pagrus Major] Iridovirus (RSIV). Fish Pathol. Jpn. 1998, 33, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Rimmer, A.E.; Becker, J.A.; Tweedie, A.; Whittington, R.J. Development of a Quantitative Polymerase Chain Reaction (QPCR) Assay for the Detection of Dwarf Gourami Iridovirus (DGIV) and Other Megalocytiviruses and Comparison with the Office International Des Epizooties (OIE) Reference PCR Protocol. Aquaculture 2012, 358–359, 155–163. [Google Scholar] [CrossRef]
- Caipang, C.M.A.; Haraguchi, I.; Ohira, T.; Hirono, I.; Aoki, T. Rapid Detection of a Fish Iridovirus Using Loop-Mediated Isothermal Amplification (LAMP). J. Virol. Methods 2004, 121, 155–161. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, C.; Huang, J.; Jia, K.; Chen, X.; Liu, H. Rapid Diagnosis of Turbot Reddish Body Iridovirus in Turbot Using the Loop-Mediated Isothermal Amplification Method. J. Virol. Methods 2009, 158, 18–23. [Google Scholar] [CrossRef]
- Ding, W.C.; Chen, J.; Shi, Y.H.; Lu, X.J.; Li, M.Y. Rapid and Sensitive Detection of Infectious Spleen and Kidney Necrosis Virus by Loop-Mediated Isothermal Amplification Combined with a Lateral Flow Dipstick. Arch. Virol. 2010, 155, 385–389. [Google Scholar] [CrossRef]
- Sukonta, T.; Senapin, S.; Meemetta, W.; Chaijarasphong, T. CRISPR-based Platform for Rapid, Sensitive and Field-deployable Detection of Scale Drop Disease Virus in Asian Sea Bass (Lates Calcarifer). J. Fish Dis. 2022, 45, 107–120. [Google Scholar] [CrossRef]
- Sudthongkong, C.; Miyata, M.; Miyazaki, T. Viral DNA Sequences of Genes Encoding the ATPase and the Major Capsid Protein of Tropical Iridovirus Isolates Which Are Pathogenic to Fishes in Japan, South China Sea and Southeast Asian Countries. Arch. Virol. 2002, 147, 2089–2109. [Google Scholar] [CrossRef]
- Adkinson, M.A.; Cambre, M.; Hedrick, R.P. Identification of an Iridovirus in Russian Sturgeon (Acipenser Guldenstadi) from Northern Europe. Bull. Eur. Assoc. Fish Pathol. UK 1998, 18, 29–32. [Google Scholar]
- Drennan, J.D.; Lapatra, S.E.; Samson, C.A.; Ireland, S.; Eversman, K.F.; Cain, K.D. Evaluation of Lethal and Non-Lethal Sampling Methods for the Detection of White Sturgeon Iridovirus Infection in White Sturgeon, Acipenser Transmontanus (Richardson). J. Fish Dis. 2007, 30, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Ciulli, S.; Volpe, E.; Sirri, R.; Passalacqua, P.L.; Cesa Bianchi, F.; Serratore, P.; Mandrioli, L. Outbreak of Mortality in Russian (Acipenser Gueldenstaedtii) and Siberian (Acipenser Baerii) Sturgeons Associated with Sturgeon Nucleo-Cytoplasmatic Large DNA Virus. Vet. Microbiol. 2016, 191, 27–34. [Google Scholar] [CrossRef]
- Clouthier, S.; VanWalleghem, E.; Anderson, E. Sturgeon Nucleo-Cytoplasmic Large DNA Virus Phylogeny and PCR Tests. Dis. Aquat. Organ. 2015, 117, 93–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigarré, L.; Lesne, M.; Lautraite, A.; Chesneau, V.; Leroux, A.; Jamin, M.; Boitard, P.M.; Toffan, A.; Prearo, M.; Labrut, S.; et al. Molecular Identification of Iridoviruses Infecting Various Sturgeon Species in Europe. J. Fish Dis. 2017, 40, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Audic, S.; Robert, C.; Abergel, C.; Renesto, P.; Ogata, H.; La Scola, B.; Suzan, M.; Claverie, J.-M. The 1.2-Megabase Genome Sequence of Mimivirus. Science 2004, 306, 1344–1350. [Google Scholar] [CrossRef]
- Abrahão, J.; Silva, L.; Silva, L.S.; Khalil, J.Y.B.; Rodrigues, R.; Arantes, T.; Assis, F.; Boratto, P.; Andrade, M.; Kroon, E.G.; et al. Tailed Giant Tupanvirus Possesses the Most Complete Translational Apparatus of the Known Virosphere. Nat. Commun. 2018, 9, 749. [Google Scholar] [CrossRef] [Green Version]
- Claverie, J.-M.; Abergel, C. Mimiviridae: An Expanding Family of Highly Diverse Large DsDNA Viruses Infecting a Wide Phylogenetic Range of Aquatic Eukaryotes. Viruses 2018, 10, 506. [Google Scholar] [CrossRef] [Green Version]
- Boutier, M.; Gao, Y.; Donohoe, O.; Vanderplasschen, A. Current Knowledge and Future Prospects of Vaccines against Cyprinid Herpesvirus 3 (CyHV-3). Fish Shellfish. Immunol. 2019, 93, 531–541. [Google Scholar] [CrossRef]
- Hanson, L.; Dishon, A.; Kotler, M. Herpesviruses That Infect Fish. Viruses 2011, 3, 2160–2191. [Google Scholar] [CrossRef] [Green Version]
- Poli, G.; Dall’Ara, P.; Martino, P.A.; Rosati, S. Microbiologia e Immunologia Veterinaria, 3rd ed.; Edra: Milan, Italy, 2017; ISBN 978-88-214-4227-8. [Google Scholar]
- Umene, K.; Sakaoka, H. Evolution of Herpes Simplex Virus Type 1 under Herpesviral Evolutionary Processes. Arch. Virol. 1999, 144, 637–656. [Google Scholar] [CrossRef]
- Sierra, E.; Fernández, A.; Fernández-Maldonado, C.; Sacchini, S.; Felipe-Jiménez, I.; Segura-Göthlin, S.; Colom-Rivero, A.; Câmara, N.; Puig-Lozano, R.; Rambaldi, A.M.; et al. Molecular Characterization of Herpesviral Encephalitis in Cetaceans: Correlation with Histopathological and Immunohistochemical Findings. Animals 2022, 12, 1149. [Google Scholar] [CrossRef]
- Lovy, J.; Friend, S.E. Cyprinid Herpesvirus-2 Causing Mass Mortality in Goldfish: Applying Electron Microscopy to Histological Samples for Diagnostic Virology. Dis. Aquat. Organ. 2014, 108, 1–9. [Google Scholar] [CrossRef]
- Doszpoly, A.; Benkő, M.; Bovo, G.; LaPatra, S.E.; Harrach, B. Comparative Analysis of a Conserved Gene Block from the Genome of the Members of the Genus Ictalurivirus. Intervirology 2011, 54, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.T.; Subramaniam, K.; Kelley, K.L.; Marcquenski, S.; Groff, J.; Waltzek, T.B. Genetic Characterization of Esocid Herpesvirus 1 (EsHV1). Dis. Aquat. Organ. 2016, 122, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sunarto, A.; McColl, K.A.; Crane, M.S.J.; Sumiati, T.; Hyatt, A.D.; Barnes, A.C.; Walker, P.J. Isolation and Characterization of Koi Herpesvirus (KHV) from Indonesia: Identification of a New Genetic Lineage: Indonesian KHV Characterization. J. Fish Dis. 2011, 34, 87–101. [Google Scholar] [CrossRef]
- Waltzek, T.B.; Kelley, G.O.; Alfaro, M.E.; Kurobe, T.; Davison, A.J.; Hedrick, R.P. Phylogenetic Relationships in the Family Alloherpesviridae. Dis. Aquat. Organ. 2009, 84, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellett, P.E.; Davison, A.J.; Ederle, R.; Ehlers, B.; Hayward, G.S.; Lacoste, V.; Minson, A.C.; Nicholas, J.; Roizman, B.; Studdert, M.J.; et al. Order Herpesvirales. In Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 99–107. [Google Scholar]
- Gotesman, M.; Kattlun, J.; Bergmann, S.M.; El-Matbouli, M. CyHV-3: The Third Cyprinid Herpesvirus. Dis. Aquat. Organ. 2013, 105, 163–174. [Google Scholar] [CrossRef] [Green Version]
- McAllister, P.E.; Lidgerding, B.C.; Herman, R.L.; Hoyer, L.C.; Hankins, J. Viral Diseases of Fish: First Report of Carp Pox in Golden Ide (Leuciscus Idus) in North America. J. Wildl. Dis. 1985, 21, 199–204. [Google Scholar] [CrossRef]
- Hedrick, R.P.; Groff, J.M.; Okihiro, M.S.; McDowell, T.S. Herpesviruses Detected in Papillomatous Skin Growths of Koi Carp (Cyprinus Carpio). J. Wildl. Dis. 1990, 26, 578–581. [Google Scholar] [CrossRef]
- Sano, N.; Sano, M.; Sano, T.; Hondo, R. Herpesvirus Cyprini: Detection of the Viral Genome by in Situ Hybridization. J. Fish Dis. 1992, 15, 153–162. [Google Scholar] [CrossRef]
- Sano, N.; Moriwake, M.; Hondo, R.; Sano, T. Herpesvirus Cyprini: A Search for Viral Genome in Infected Fish by Infected Fish by in Situ Hybridization. J. Fish Dis. 1993, 16, 495–499. [Google Scholar] [CrossRef]
- Way, K.; Dixon, P.F. Koi Herpesvirus Disease. In Fish Viruses and Bacteria: Pathobiology and Protection; CABI Digital Library: Long Beach, CA, USA, 2017; pp. 115–127. ISBN 978-1-78064-778-4. [Google Scholar]
- Lievens, B.; Frans, I.; Heusdens, C.; Justé, A.; Jonstrup, S.P.; Lieffrig, F.; Willems, K.A. Rapid Detection and Identification of Viral and Bacterial Fish Pathogens Using a DNA Array-Based Multiplex Assay. J. Fish Dis. 2011, 34, 861–875. [Google Scholar] [CrossRef]
- Sirri, R.; Ciulli, S.; Barbé, T.; Volpe, E.; Lazzari, M.; Franceschini, V.; Errani, F.; Sarli, G.; Mandrioli, L. Detection of Cyprinid Herpesvirus 1 DNA in Cutaneous Squamous Cell Carcinoma of Koi Carp (Cyprinus Carpio). Vet. Dermatol. 2018, 29, 60-e24. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, K.; Kurita, J.; Kawana, M.; Kiryu, I.; Oseko, N.; Sano, M. Development of MRNA-Specific RT-PCR for the Detection of Koi Herpesvirus (KHV) Replication Stage. Dis. Aquat. Organ. 2012, 100, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zeng, L.; Zhang, H.; Zhou, Y.; Ma, J.; Fan, Y. Cyprinid Herpesvirus 2 Infection Emerged in Cultured Gibel Carp, Carassius Auratus Gibelio in China. Vet. Microbiol. 2013, 166, 138–144. [Google Scholar] [CrossRef]
- Jiang, N.; Xu, J.; Ma, J.; Fan, Y.; Zhou, Y.; Liu, W.; Zeng, L. Histopathology and Ultrastructural Pathology of Cyprinid Herpesvirus II (CyHV-2) Infection in Gibel Carp, Carassius Auratus Gibelio. Wuhan Univ. J. Nat. Sci. 2015, 20, 413–420. [Google Scholar] [CrossRef]
- Fichi, G.; Susini, F.; Cocumelli, C.; Cersini, A.; Salvadori, M.; Guarducci, M.; Cardeti, G. New Detection of Cyprinid Herpesvirus 2 in Mass Mortality Event of Carassius Carassius (L.), in Italy. J. Fish Dis. 2016, 39, 1523–1527. [Google Scholar] [CrossRef]
- Tang, R.; Lu, L.; Wang, B.; Yu, J.; Wang, H. Identification of the Immediate-Early Genes of Cyprinid Herpesvirus 2. Viruses 2020, 12, 994. [Google Scholar] [CrossRef]
- Wang, H.; Xu, L.; Lu, L. Detection of Cyprinid Herpesvirus 2 in Peripheral Blood Cells of Silver Crucian Carp, Carassius Auratus Gibelio (Bloch), Suggests Its Potential in Viral Diagnosis. J. Fish Dis. 2016, 39, 155–162. [Google Scholar] [CrossRef]
- Ding, Z.; Xia, S.; Zhao, Z.; Xia, A.; Shen, M.; Tang, J.; Xue, H.; Geng, X.; Yuan, S. Histopathological Characterization and Fluorescence in Situ Hybridization of Cyprinid Herpesvirus 2 in Cultured Prussian Carp, Carassius Auratus Gibelio in China. J. Virol. Methods 2014, 206, 76–83. [Google Scholar] [CrossRef]
- Ma, J.; Jiang, N.; LaPatra, S.E.; Jin, L.; Xu, J.; Fan, Y.; Zhou, Y.; Zeng, L. Establishment of a Novel and Highly Permissive Cell Line for the Efficient Replication of Cyprinid Herpesvirus 2 (CyHV-2). Vet. Microbiol. 2015, 177, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Kurita, J.; Ozaki, A.; Sano, M.; Fukuda, H.; Ototake, M. Growth of Cyprinid Herpesvirus 2 (CyHV-2) in Cell Culture and Experimental Infection of Goldfish Carassius Auratus. Dis. Aquat. Organ. 2013, 105, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Xu, D.; Lu, L. A Novel Cell Line Established from Caudal Fin Tissue of Carassius Auratus Gibelio Is Susceptible to Cyprinid Herpesvirus 2 Infection with the Induction of Apoptosis. Virus Res. 2018, 258, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Engelsma, M.Y.; Way, K.; Dodge, M.J.; Voorbergen-Laarman, M.; Panzarin, V.; Abbadi, M.; El-Matbouli, M.; Frank Skall, H.; Kahns, S.; Stone, D.M. Detection of Novel Strains of Cyprinid Herpesvirus Closely Related to Koi Herpesvirus. Dis. Aquat. Organ. 2013, 107, 113–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, A.; Merry, G.; Sadler, J. Detection of the Herpesviral Hematopoietic Necrosis Disease Agent (Cyprinid Herpesvirus 2) in Moribund and Healthy Goldfish: Validation of a Quantitative PCR Diagnostic Method. Dis. Aquat. Organ. 2006, 69, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Shi, X.; Yu, L.; Zheng, X.; Lan, W.; Jia, P.; Wang, J.; Liu, H. Development and Evaluation of a Loop-Mediated Isothermal Amplification Assay for Diagnosis of Cyprinid Herpesvirus 2. J. Virol. Methods 2013, 194, 206–210. [Google Scholar] [CrossRef]
- Preena, P.G.; Kumar, T.V.A.; Johny, T.K.; Dharmaratnam, A.; Swaminathan, T.R. Quick Hassle-Free Detection of Cyprinid Herpesvirus 2 (CyHV-2) in Goldfish Using Recombinase Polymerase Amplification-Lateral Flow Dipstick (RPA-LFD) Assay. Aquac. Int. 2022, 30, 1211–1220. [Google Scholar] [CrossRef]
- Wang, H.; Sun, M.; Xu, D.; Podok, P.; Xie, J.; Jiang, Y.; Lu, L. Rapid Visual Detection of Cyprinid Herpesvirus 2 by Recombinase Polymerase Amplification Combined with a Lateral Flow Dipstick. J. Fish Dis. 2018, 41, 1201–1206. [Google Scholar] [CrossRef]
- Yang, J.; Wen, J.; Xiao, S.; Wei, C.; Yu, F.; Roengjit, P.; Lu, L.; Wang, H. Complete Genome and Molecular Characterization of a New Cyprinid Herpesvirus 2 (CyHV-2) SH-01 Strain Isolated from Cultured Crucian Carp. Viruses 2022, 14, 2068. [Google Scholar] [CrossRef]
- Chong, R. Chapter 14 Koi Herpesvirus Disease. In Aquaculture Pathophysiology Volume I. Fish Diseases; Elsevier Academic Press: New York, NY, USA, 2022; pp. 189–199. [Google Scholar]
- Hanson, L.; Doszpoly, A.; Van Beurden, S.J.; de Oliveira Viadanna, P.H.; Waltzek, T.B. Chapter 9 Alloherpesviruses of Fish. In Aquaculture Virology; Elsevier Academic Press: New York, NY, USA, 2016; pp. 153–172. [Google Scholar]
- Bercovier, H.; Fishman, Y.; Nahary, R.; Sinai, S.; Zlotkin, A.; Eyngor, M.; Gilad, O.; Eldar, A.; Hedrick, R.P. Cloning of the Koi Herpesvirus (KHV) Gene Encoding Thymidine Kinase and Its Use for a Highly Sensitive PCR Based Diagnosis. BMC Microbiol. 2005, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, W.L.; Mullis, L.; LaPatra, S.E.; Groff, J.M.; Goodwin, A. Detection of Koi Herpesvirus DNA in Tissues of Infected Fish. J. Fish Dis. 2002, 25, 171–178. [Google Scholar] [CrossRef]
- Yuasa, K.; Sano, M.; Kurita, J.; Ito, T.; Iida, T. Improvement of a PCR Method with the Sph I-5 Primer Set for the Detection of Koi Herpesvirus (KHV). Fish Pathol. 2005, 40, 37–39. [Google Scholar] [CrossRef]
- Yuasa, K.; Sano, M. Koi Herpesvirus: Status of Outbreaks, Diagnosis, Surveillance, and Research. Isr. J. Aquac. Bamidgeh 2009, 61, 169–179. [Google Scholar] [CrossRef]
- OIE CHAPTER 2.3.6 Infection with Koi Herpesvirus. In Manual of Diagnostic Tests for Aquatic Animals; OIE: Paris, France, 2022.
- Yuasa, K.; Kawana, M.; Ito, T.; Kiryu, I.; Oseko, N.; Sano, M. Intra Vitam Assays for Detecting Fish Infected with Cyprinid Herpesvirus 3 (CyHV-3). Dis. Aquat. Organ. 2022, 149, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Soliman, H.; El-Matbouli, M. Rapid Detection and Differentiation of Carp Oedema Virus and Cyprinid Herpes Virus-3 in Koi and Common Carp. J. Fish Dis. 2018, 41, 761–772. [Google Scholar] [CrossRef]
- Boutier, M.; Ronsmans, M.; Rakus, K.; Jazowiecka-Rakus, J.; Vancsok, C.; Morvan, L.; Peñaranda, M.M.D.; Stone, D.M.; Way, K.; van Beurden, S.J.; et al. Cyprinid Herpesvirus 3: An Archetype of Fish Alloherpesviruses. Adv. Virus Res. 2015, 93, 161–256. [Google Scholar] [CrossRef] [PubMed]
- Cano, I.; Worswick, J.; Mulhearn, B.; Stone, D.; Wood, G.; Savage, J.; Paley, R. A Seasonal Study of Koi Herpesvirus and Koi Sleepy Disease Outbreaks in the United Kingdom in 2018 Using a Pond-Side Test. Animals 2021, 11, 459. [Google Scholar] [CrossRef]
- Thorstensen, M.J.; Vandervelde, C.A.; Bugg, W.S.; Michaleski, S.; Vo, L.; Mackey, T.E.; Lawrence, M.J.; Jeffries, K.M. Non-Lethal Sampling Supports Integrative Movement Research in Freshwater Fish. Front. Genet. 2022, 13, 795355. [Google Scholar] [CrossRef]
- Monaghan, S.J.; Thompson, K.D.; Adams, A.; Kempter, J.; Bergmann, S.M. Examination of the Early Infection Stages of Koi Herpesvirus (KHV) in Experimentally Infected Carp, Cyprinus Carpio L. Using in Situ Hybridization. J. Fish Dis. 2015, 38, 477–489. [Google Scholar] [CrossRef]
- Doszpoly, A.; Kovács, E.R.; Bovo, G.; LaPatra, S.E.; Harrach, B.; Benkő, M. Molecular Confirmation of a New Herpesvirus from Catfish (Ameiurus Melas) by Testing the Performance of a Novel PCR Method, Designed to Target the DNA Polymerase Gene of Alloherpesviruses. Arch. Virol. 2008, 153, 2123–2127. [Google Scholar] [CrossRef]
- Doszpoly, A.; Shchelkunov, I. Partial Genome Analysis of Siberian Sturgeon Alloherpesvirus Suggests Its Close Relation to AciHV-2—Short Communication. Acta Vet. Hung. 2010, 58, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Radosavljević, V.; Milićević, V.; Maksimović-Zorić, J.; Veljović, L.; Nešić, K.; Pavlović, M.; Pelić, D.L.; Marković, Z. Sturgeon Diseases in Aquaculture. Arch. Vet. Med. 2019, 12, 5–20. [Google Scholar] [CrossRef]
- Doszpoly, A.; Kalabekov, I.M.; Breyta, R.; Shchelkunov, I.S. Isolation and Characterization of an Atypical Siberian Sturgeon Herpesvirus Strain in Russia: Novel North American Acipenserid Herpesvirus 2 Strain in Europe? J. Fish Dis. 2017, 40, 1363–1372. [Google Scholar] [CrossRef]
- Kelley, G.O.; Waltzek, T.B.; McDowell, T.S.; Yun, S.C.; LaPatra, S.E.; Hedrick, R.P. Genetic Relationships among Herpes-Like Viruses Isolated from Sturgeon. J. Aquat. Anim. Health 2005, 17, 297–303. [Google Scholar] [CrossRef]
- Kurobe, T.; Kelley, G.O.; Waltzek, T.B.; Hedrick, R.P. Revised Phylogenetic Relationships among Herpesviruses Isolated from Sturgeons. J. Aquat. Anim. Health 2008, 20, 96–102. [Google Scholar] [CrossRef]
- Ciulli, S.; Volpe, E.; Sirri, R.; Tura, G.; Errani, F.; Zamperin, G.; Toffan, A.; Silvi, M.; Renzi, A.; Abbadi, M.; et al. Multifactorial Causes of Chronic Mortality in Juvenile Sturgeon (Huso Huso). Animals 2020, 10, 1866. [Google Scholar] [CrossRef]
- Hedrick, R.; McDowell, T.; Gilad, O.; Adkison, M.; Bovo, G. Systemic Herpes-like Virus in Catfish Ictalurus Melas (Italy) Differs from Ictalurid Herpesvirus 1 (North America). Dis. Aquat. Organ. 2003, 55, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Alborali, L.; Paternello, P.; Lavazza, A.; Caggiano, M.; Lombardi, G. Descrizione Di Un Episodio Di Linfocisti Nel Sarago (Diplodus Puntazzo). Boll. Soc. Ital. Patol. Ittica 1996, 20, 2–7. [Google Scholar]
- Thompson, D.J.; Khoo, L.H.; Wise, D.J.; Hanson, L.A. Evaluation of Channel Catfish Virus Latency on Fingerling Production Farms in Mississippi. J. Aquat. Anim. Health 2005, 17, 211–215. [Google Scholar] [CrossRef]
- Gray, W.L.; Williams, R.J.; Jordan, R.L.; Griffin, B.R. Detection of Channel Catfish Virus DNA in Latently Infected Catfish. J. Gen. Virol. 1999, 80, 1817–1822. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, A.E.; Marecaux, E. Validation of a QPCR Assay for the Detection of Ictalurid Herpesvirus-2 (IcHV-2) in Fish Tissues and Cell Culture Supernatants. J. Fish Dis. 2010, 33, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Hao, K.; Liu, H.-Y.; Chen, X.-H.; Yuan, J.-F.; Li, L.-J.; Bian, W.-J.; Zhao, Z. Real-Time fluorescent loop mediated isothermal amplification for detection of channel catfish virus. Acta Hydrobiol. Sin. 2021, 45, 541–546. [Google Scholar] [CrossRef]
- Barbosa-Solomieu, V.; Miossec, L.; Vázquez-Juárez, R.; Ascencio-Valle, F.; Renault, T. Diagnosis of Ostreid Herpesvirus 1 in Fixed Paraffin-Embedded Archival Samples Using PCR and in Situ Hybridisation. J. Virol. Methods 2004, 119, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Batista, F.M.; Arzul, I.; Pepin, J.-F.; Ruano, F.; Friedman, C.S.; Boudry, P.; Renault, T. Detection of Ostreid Herpesvirus 1 DNA by PCR in Bivalve Molluscs: A Critical Review. J. Virol. Methods 2007, 139, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Carter, M.J. Enterically Infecting Viruses: Pathogenicity, Transmission and Significance for Food and Waterborne Infection. J. Appl. Microbiol. 2005, 98, 1354–1380. [Google Scholar] [CrossRef]
- Errani, F.; Ponti, M.; Volpe, E.; Ciulli, S. Spatial and Seasonal Variability of Human and Fish Viruses in Mussels inside and Offshore of Ravenna’s Harbour (Adriatic Sea, Italy). J. Appl. Microbiol. 2021, 130, 994–1008. [Google Scholar] [CrossRef]
- Volpe, E.; Pagnini, N.; Serratore, P.; Ciulli, S. Fate of Redspotted Grouper Nervous Necrosis Virus (RGNNV) in Experimentally Challenged Manila Clam Ruditapes Philippinarum. Dis. Aquat. Organ. 2017, 125, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Furhmann, M.; Patirana, E.; de Kantzou, M.; Hick, P. Chapter 63 Ostreid Herpesvirus Disease. In Aquaculture Pathophysiology Volume II. Crustacean and Molluscan Diseases; Elsevier Academic Press: New York, NY, USA, 2022; pp. 473–488. ISBN 978-0-323-95434-1. [Google Scholar]
- Evans, O.; Paul-Pont, I.; Whittington, R.J. Detection of Ostreid Herpesvirus 1 Microvariant DNA in Aquatic Invertebrate Species, Sediment and Other Samples Collected from the Georges River Estuary, New South Wales, Australia. Dis. Aquat. Organ. 2017, 122, 247–255. [Google Scholar] [CrossRef]
- Bueno, R.; Perrott, M.; Dunowska, M.; Brosnahan, C.; Johnston, C. In Situ Hybridization and Histopathological Observations during Ostreid Herpesvirus-1-Associated Mortalities in Pacific Oysters Crassostrea Gigas. Dis. Aquat. Organ. 2016, 122, 43–55. [Google Scholar] [CrossRef]
- Toldrà, A.; Furones, M.D.; O’Sullivan, C.K.; Campàs, M. Detection of Isothermally Amplified Ostreid Herpesvirus 1 DNA in Pacific Oyster (Crassostrea Gigas) Using a Miniaturised Electrochemical Biosensor. Talanta 2020, 207, 120308. [Google Scholar] [CrossRef]
- Dotto-Maurel, A.; Pelletier, C.; Morga, B.; Jacquot, M.; Faury, N.; Dégremont, L.; Bereszczynki, M.; Delmotte, J.; Escoubas, J.-M.; Chevignon, G. Evaluation of Tangential Flow Filtration Coupled to Long-Read Sequencing for Ostreid Herpesvirus Type 1 Genome Assembly. Microb. Genom. 2022, 8, mgen000895. [Google Scholar] [CrossRef]
- Trancart, S.; Tweedie, A.; Liu, O.; Paul-Pont, I.; Hick, P.; Houssin, M.; Whittington, R.J. Diversity and Molecular Epidemiology of Ostreid Herpesvirus 1 in Farmed Crassostrea Gigas in Australia: Geographic Clusters and Implications for “Microvariants” in Global Mortality Events. Virus Res. 2023, 323, 198994. [Google Scholar] [CrossRef] [PubMed]
- Thiéry, R.; Johnson, K.L.; Nakai, T.; Schneemann, A.; Bonami, J.R.; Lightner, D.V. Family Nodaviridae. In Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 1061–1067. [Google Scholar]
- Bandín, I.; Souto, S. Betanodavirus and VER Disease: A 30-Year Research Review. Pathogens 2020, 9, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpe, E.; Gustinelli, A.; Caffara, M.; Errani, F.; Quaglio, F.; Fioravanti, M.L.; Ciulli, S. Viral Nervous Necrosis Outbreaks Caused by the RGNNV/SJNNV Reassortant Betanodavirus in Gilthead Sea Bream (Sparus Aurata) and European Sea Bass (Dicentrarchus Labrax). Aquaculture 2020, 523, 735155. [Google Scholar] [CrossRef]
- Schneemann, A.; Reddy, V.; Johnson, J.E. The Structure and Function of Nodavirus Particles: A Paradigm for Understanding Chemical Biology. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 1998; Volume 50, pp. 381–446. ISBN 978-0-12-039850-8. [Google Scholar]
- Nishizawa, T.; Mori, K.-I.; Furuhashi, M.; Nakai, T.; Furusawa, I.; Muroga, K. Comparison of the Coat Protein Genes of Five Fish Nodaviruses, the Causative Agents of Viral Nervous Necrosis in Marine Fish. J. Gen. Virol. 1995, 76, 1563–1569. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic Classification of Fish Nodaviruses by Molecular Phylogenetic Analysis of the Coat Protein Gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toffolo, V.; Negrisolo, E.; Maltese, C.; Bovo, G.; Belvedere, P.; Colombo, L.; Valle, L.D. Phylogeny of Betanodaviruses and Molecular Evolution of Their RNA Polymerase and Coat Proteins. Mol. Phylogenet. Evol. 2007, 43, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Ho, K.L.; Gabrielsen, M.; Beh, P.L.; Kueh, C.L.; Thong, Q.X.; Streetley, J.; Tan, W.S.; Bhella, D. Structure of the Macrobrachium Rosenbergii Nodavirus: A New Genus within the Nodaviridae? PLoS Biol. 2018, 16, e3000038. [Google Scholar] [CrossRef] [Green Version]
- NaveenKumar, S.; Shekar, M.; Karunasagar, I.; Karunasagar, I. Genetic Analysis of RNA1 and RNA2 of Macrobrachium Rosenbergii Nodavirus (MrNV) Isolated from India. Virus Res. 2013, 173, 377–385. [Google Scholar] [CrossRef]
- Johnson, K.L.; Moore, J.S. Nodaviruses of Invertebrates and Fish (Nodaviridae). In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 819–826. ISBN 978-0-12-814516-6. [Google Scholar]
- Mori, K.; Nakai, T.; Muroga, K.; Arimoto, M.; Mushiake, K.; Furusawa, I. Properties of a New Virus Belonging to Nodaviridae Found in Larval Striped Jack (Pseudocaranx Dentex) with Nervous Necrosis. Virology 1992, 187, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Munday, B.L.; Kwang, J.; Moody, N. Betanodavirus Infections of Teleost Fish: A Review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Iwamoto, T.; Okinaka, Y.; Mise, K.; Mori, K.-I.; Arimoto, M.; Okuno, T.; Nakai, T. Identification of Host-Specificity Determinants in Betanodaviruses by Using Reassortants between Striped Jack Nervous Necrosis Virus and Sevenband Grouper Nervous Necrosis Virus. J. Virol. 2004, 78, 1256–1262. [Google Scholar] [CrossRef] [Green Version]
- Johansen, R.; Sommerset, I.; Tørud, B.; Korsnes, K.; Hjortaas, M.J.; Nilsen, F.; Nerland, A.H.; Dannevig, B.H. Characterization of Nodavirus and Viral Encephalopathy and Retinopathy in Farmed Turbot, Scophthalmus Maximus (L.). J. Fish Dis. 2004, 27, 591–601. [Google Scholar] [CrossRef]
- Gagné, N.; Johnson, S.C.; Cook-Versloot, M.; MacKinnon, A.M.; Olivier, G. Molecular Detection and Characterization of Nodavirus in Several Marine Fish Species from the Northeastern Atlantic. Dis. Aquat. Organ. 2004, 62, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Kwon, W.J.; Min, J.G.; Kim, K.I.; Jeong, H.D. Complete Genome Sequence and Pathogenic Analysis of a New Betanodavirus Isolated from Shellfish. J. Fish Dis. 2019, 42, 519–531. [Google Scholar] [CrossRef]
- Frerichs, G.N.; Rodger, H.D.; Peric, Z. Cell Culture Isolation of Piscine Neuropathy Nodavirus from Juvenile Sea Bass, Dicentrarchus Labrax. J. Gen. Virol. 1996, 77 Pt 9, 2067–2071. [Google Scholar] [CrossRef]
- Zrnčić, S.; Padros, F.; Mladineo, I.; Fioravanti, M.-L.; Gustinelli, A.; Palenzuela, O.; Toffan, A.; Cuilli, S.; Fouz, B.; Breton, A.L.; et al. Bottlenecks in Diagnostics of Mediterranean Fish Diseases; European Association of Fish Pathologists: Aberdeen City, UK, 2022; Volume 40. [Google Scholar]
- Baud, M.; Cabon, J.; Salomoni, A.; Toffan, A.; Panzarin, V.; Bigarré, L. First Generic One Step Real-Time Taqman RT-PCR Targeting the RNA1 of Betanodaviruses. J. Virol. Methods 2015, 211, 1–7. [Google Scholar] [CrossRef]
- Sung, C.-H.; Lu, J.-K. Reverse Transcription Loop-Mediated Isothermal Amplification for Rapid and Sensitive Detection of Nervous Necrosis Virus in Groupers. J. Virol. Methods 2009, 159, 206–210. [Google Scholar] [CrossRef]
- Xu, H.-D.; Feng, J.; Guo, Z.-X.; Ou, Y.-J.; Wang, J.-Y. Detection of Red-Spotted Grouper Nervous Necrosis Virus by Loop-Mediated Isothermal Amplification. J. Virol. Methods 2010, 163, 123–128. [Google Scholar] [CrossRef]
- Hwang, J.; Suh, S.-S.; Park, M.; Oh, M.-J.; Kim, J.-O.; Lee, S.; Lee, T.-K. Detection of Coat Protein Gene of Nervous Necrosis Virus Using Loop-Mediated Isothermal Amplification. Asian Pac. J. Trop. Med. 2016, 9, 235–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toffan, A.; Pascoli, F.; Pretto, T.; Panzarin, V.; Abbadi, M.; Buratin, A.; Quartesan, R.; Gijón, D.; Padrós, F. Viral Nervous Necrosis in Gilthead Sea Bream (Sparus Aurata) Caused by Reassortant Betanodavirus RGNNV/SJNNV: An Emerging Threat for Mediterranean Aquaculture. Sci. Rep. 2017, 7, 46755. [Google Scholar] [CrossRef] [Green Version]
- Olveira, J.G.; Souto, S.; Dopazo, C.P.; Thiéry, R.; Barja, J.L.; Bandín, I. Comparative Analysis of Both Genomic Segments of Betanodaviruses Isolated from Epizootic Outbreaks in Farmed Fish Species Provides Evidence for Genetic Reassortment. J. Gen. Virol. 2009, 90, 2940–2951. [Google Scholar] [CrossRef]
- Panzarin, V.; Fusaro, A.; Monne, I.; Cappellozza, E.; Patarnello, P.; Bovo, G.; Capua, I.; Holmes, E.C.; Cattoli, G. Molecular Epidemiology and Evolutionary Dynamics of Betanodavirus in Southern Europe. Infect. Genet. Evol. 2012, 12, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Biasini, L.; Berto, P.; Abbadi, M.; Buratin, A.; Toson, M.; Marsella, A.; Toffan, A.; Pascoli, F. Pathogenicity of Different Betanodavirus RGNNV/SJNNV Reassortant Strains in European Sea Bass. Pathogens 2022, 11, 458. [Google Scholar] [CrossRef]
- Krishnan, R.; Kim, J.-O.; Qadiri, S.S.N.; Kim, J.-O.; Oh, M.-J. Early Viral Uptake and Host-Associated Immune Response in the Tissues of Seven-Band Grouper Following a Bath Challenge with Nervous Necrosis Virus. Fish Shellfish. Immunol. 2020, 103, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Qi, J.; He, L.; Luo, H.; Lin, J.; Qiu, F.; Wang, Q.; Zheng, L. Isolation and Identification of a New Strain of Nervous Necrosis Virus from the Big-Belly Seahorse Hippocampus Abdominalis. Virol. J. 2022, 19, 109. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, S.; Singh, I.S.B.; Puthumana, J. Status in Molluscan Cell Line Development in Last One Decade (2010–2020): Impediments and Way Forward. Cytotechnology 2022, 74, 433–457. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.E.J.; Bufalino, M.R.; Christie, A.E.; Frischer, M.E.; Soin, T.; Tsui, C.K.M.; Hanner, R.H.; Smagghe, G. Misidentification of OLGA-PH-J/92, Believed to Be the Only Crustacean Cell Line. In Vitro Cell. Dev. Biol. Anim. 2011, 47, 665–674. [Google Scholar] [CrossRef]
- OIE CHAPTER 2.3.8 Infection with Salmon Alphavirus. In Manual of Diagnostic Tests for Aquatic Animals; OIE: Paris, France, 2021; pp. 344–359.
- OIE CHAPTER 2.3.10 Infection with Viral Haemorrhagic Septicaemia Virus. In Manual of Diagnostic Tests for Aquatic Animals; OIE: Paris, France, 2021; pp. 377–401.


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volpe, E.; Errani, F.; Mandrioli, L.; Ciulli, S. Advances in Viral Aquatic Animal Disease Knowledge: The Molecular Methods’ Contribution. Biology 2023, 12, 466. https://doi.org/10.3390/biology12030466
Volpe E, Errani F, Mandrioli L, Ciulli S. Advances in Viral Aquatic Animal Disease Knowledge: The Molecular Methods’ Contribution. Biology. 2023; 12(3):466. https://doi.org/10.3390/biology12030466
Chicago/Turabian StyleVolpe, Enrico, Francesca Errani, Luciana Mandrioli, and Sara Ciulli. 2023. "Advances in Viral Aquatic Animal Disease Knowledge: The Molecular Methods’ Contribution" Biology 12, no. 3: 466. https://doi.org/10.3390/biology12030466
APA StyleVolpe, E., Errani, F., Mandrioli, L., & Ciulli, S. (2023). Advances in Viral Aquatic Animal Disease Knowledge: The Molecular Methods’ Contribution. Biology, 12(3), 466. https://doi.org/10.3390/biology12030466