


Adaptability of Wild-Growing Tulips of Greece: Uncovering Relationships between Soil Properties, Rhizosphere Fungal Morphotypes and Nutrient Content Profiles

 ,
,  ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Focal Greek Tulips
2.2. Collection of Samplings
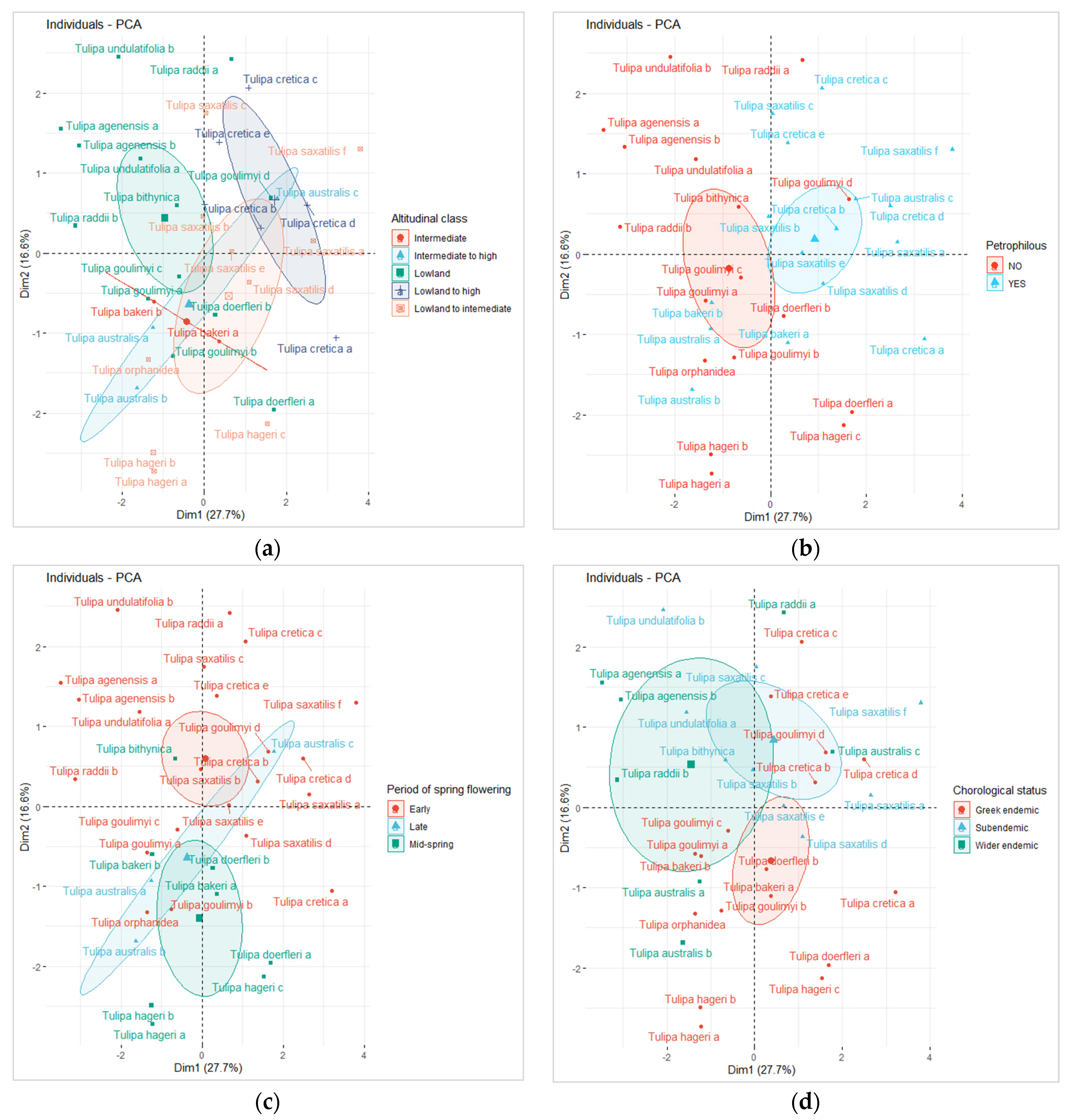
2.3. Classification of Studied Species into Functional Groups
- Tulips of mainland Greece and insular tulips of Greece (North Aegean tulips and Cretan tulips);
- Alien naturalized tulips and wild-growing Greek native tulips;
- Greek endemic tulips, sub-endemic tulips, and tulips of wider distribution range;
- Non-threatened and threatened tulips (critically endangered, endangered and vulnerable);
- Tulips of small altitudinal range (lowland tulips, tulips of intermediate altitudes), tulips of wide altitudinal range (tulips of lowland to intermediate altitudes, tulips of intermediate to high altitudes) and tulips of very wide altitudinal range (tulips occurring from lowlands to high altitudes);
- Tulips associated with agricultural habitat types, tulips associated with natural habitat types, and tulips occurring in both agricultural and natural habitat types;
- Rock-dwelling tulips and tulips not occurring in rocky habitats;
- Segetal tulips and tulips not occurring in agricultural habitats;
- Early flowering tulips, mid-spring flowering tulips, and late flowering tulips.
2.4. Soil and Plant Analysis
2.5. Statistical Analysis
3. Results
3.1. Soil Properties of the Studied Samples
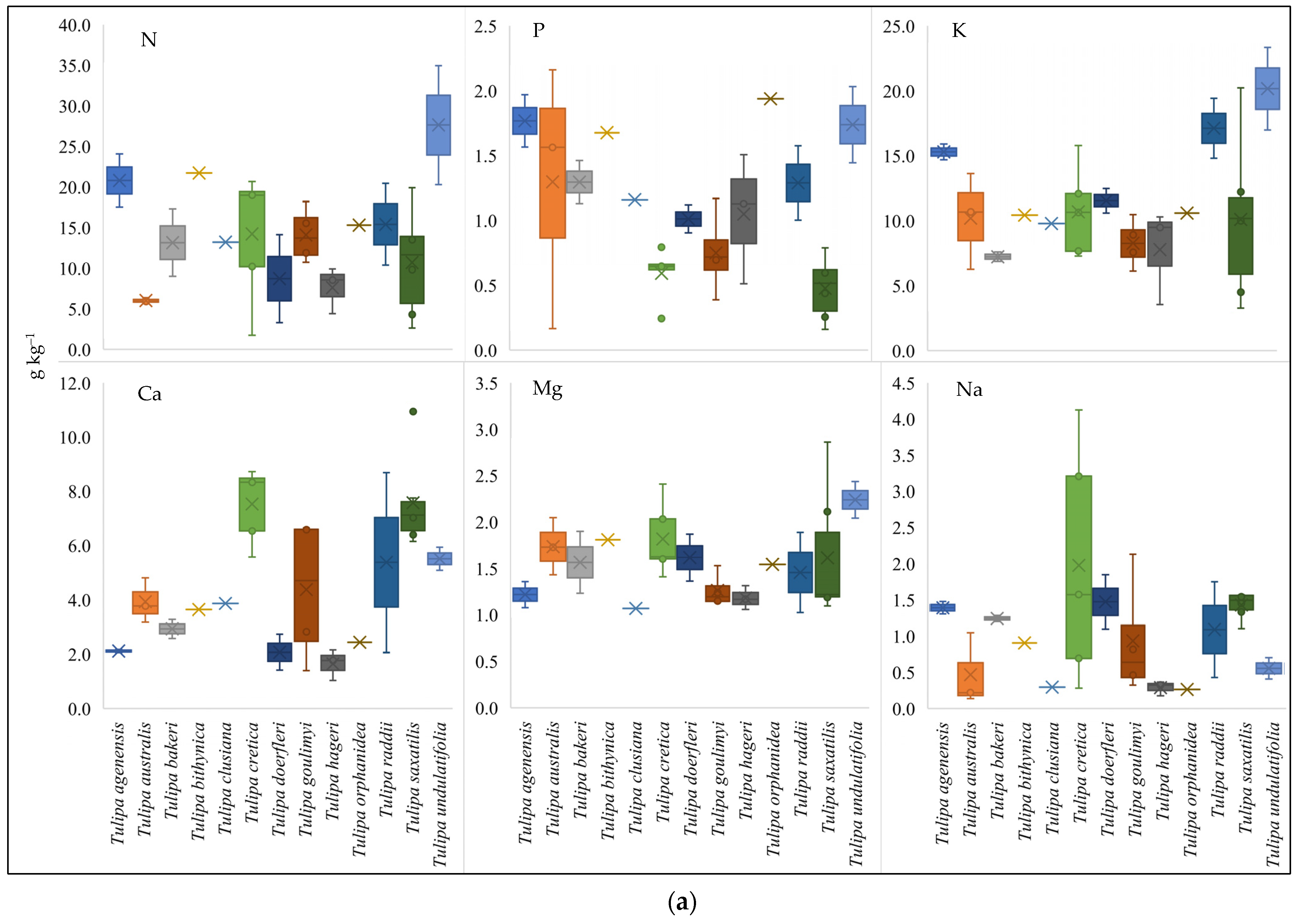
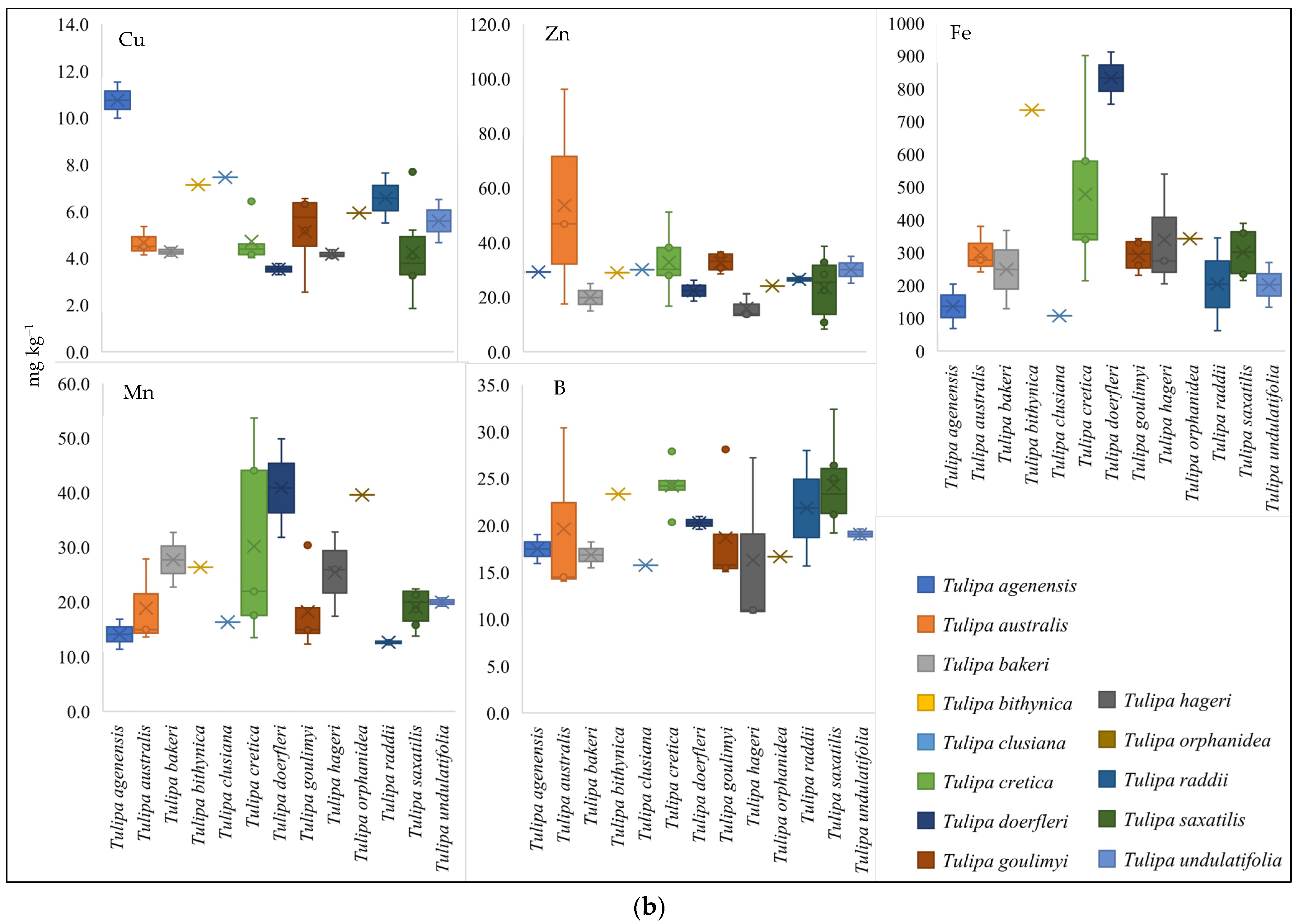
3.2. Nutritional Status of the Studied Greek Tulip Species
3.3. Interrelationships between Tulips’ Essential Macro- and Micro-Nutrients Content
3.4. Relationships between Rhizosphere’s Arbuscular Mycorrhizal Fungi (AMF) Spore Morphotypes, Soil Parameters, and Tulips Nutrients Content
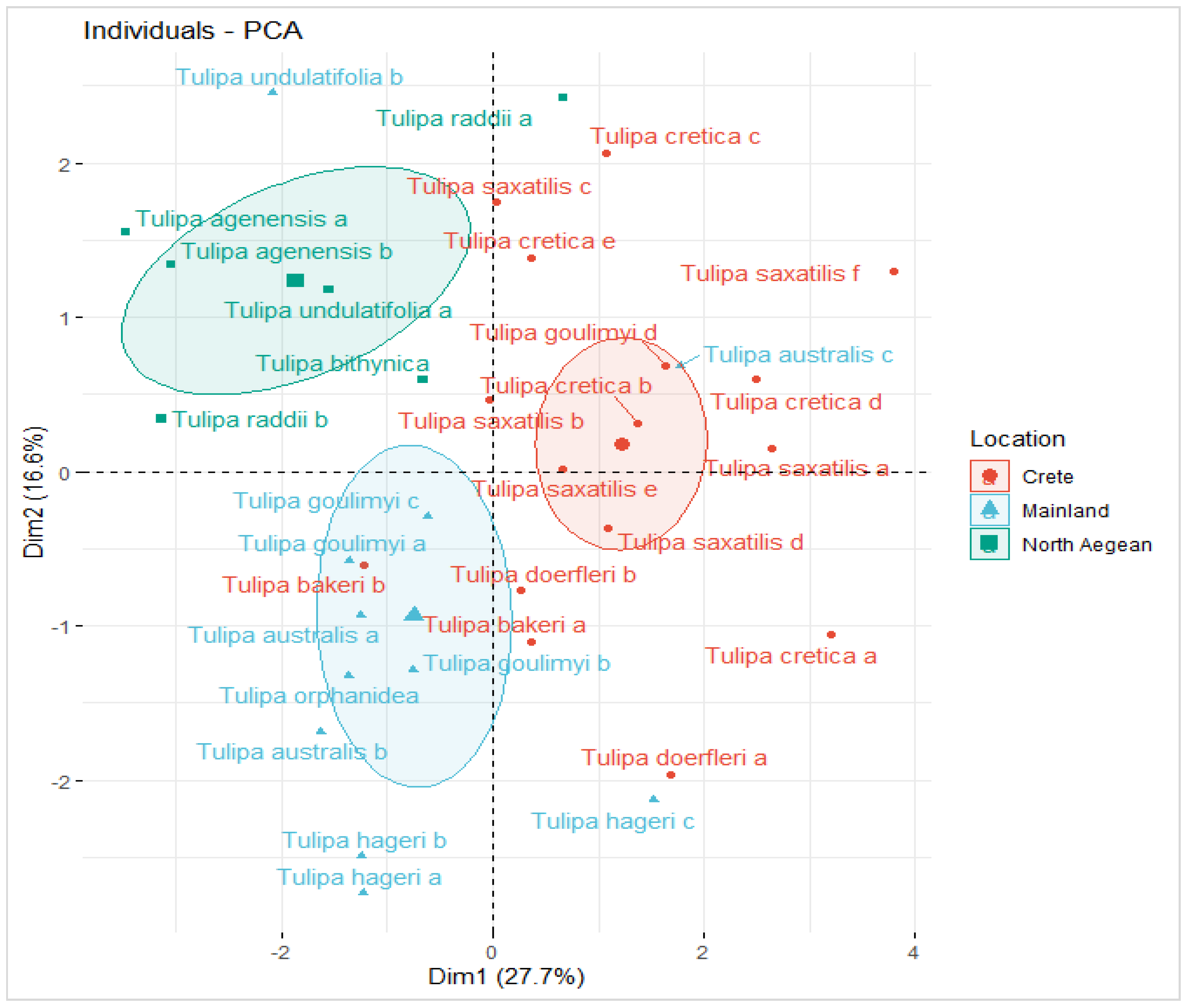
3.5. Nutrients Variability of Wild-Growing Greek Tulips and Functional Types Distribution
3.6. Effects of Soil Properties on Greek Tulips’ Above-Ground Macro- and Micro-Nutrients Content Variability
3.7. Evaluating Differences in Nutrients Profile and in Soil Properties between Different Spatial Units
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krigas, N.; Lykas, C.; Ipsilantis, I.; Matsi, T.; Weststrand, S.; Havström, M.; Tsoktouridis, G. Greek Tulips: Worldwide Electronic Trade over the Internet, Global Ex Situ Conservation and Current Sustainable Exploitation Challenges. Plants 2021, 10, 580. [Google Scholar] [CrossRef]
- Strid, A. Atlas of the Aegean Flora, Part 1: Text & Plates; Part 2: Maps; Englera, Volume 33; Botanic Garden and Botanical Museum Berlin, Freie Universität Berlin: Berlin, Germany, 2016; ISBN 978-3-921800-97-3 (Volume 1); 978-3-921800-98-0 (Volume 2). [Google Scholar]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Strid, A.; Dimopoulos, P. Extinction Risk Assessment of the Greek Endemic Flora. Biology 2021, 10, 195. [Google Scholar] [CrossRef]
- Phitos, D.; Strid, A.; Snogerup, S.; Greuter, W. The Red Data Book of Rare and Threatened Plants of Greece; World Wide Fund for Nature: Gland, Switzerland, 1995; ISBN 9789607506047. [Google Scholar]
- Phitos, D.; Constantinidis, T.H.; Kamari, G. The Red Data Book of Rare and Threatened Plants of Greece; Hellenic Βotanical Society: Patra, Greece, 2009; Volume II, ISBN 978-960-9407-09-0. [Google Scholar]
- Zhang, J.; Ren, T.; Yang, J.; Xu, L.; Li, M.; Zhang, Y.; Han, X.; He, N. Leaf Multi-Element Network Reveals the Change of Species Dominance Under Nitrogen Deposition. Front. Plant Sci. 2021, 12, 580340. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, H., Marschner, P., Eds.; Elsevier: London, UK; Academic Press: Waltham, MA, USA, 2012; ISBN 978-0-12-384905-2. [Google Scholar]
- Morgan, J.; Connolly, E. Plant-soil interactions: Nutrient uptake. Nat. Educ. Knowl. 2013, 4, 2. [Google Scholar]
- Zhang, C.; Hiradate, S.; Kusumoto, Y.; Morita, S.; Koyanagi, T.F.; Chu, Q.; Watanabe, T. Ionomic Responses of Local Plant Species to Natural Edaphic Mineral Variations. Front. Plant Sci. 2021, 12, 614613. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Azuma, T. Ionomic Variation in Leaves of 819 Plant Species Growing in the Botanical Garden of Hokkaido University, Japan. J. Plant. Res. 2021, 134, 291–304. [Google Scholar] [CrossRef]
- Gupta, N.; Yadav, K.K.; Kumar, V.; Kumar, S.; Chadd, R.P.; Kumar, A. Trace Elements in Soil-Vegetables Interface: Translocation, Bioaccumulation, Toxicity and Amelioration—A Review. Sci. Total Environ. 2019, 651, 2927–2942. [Google Scholar] [CrossRef] [PubMed]
- Likar, M.; Vogel-Mikuš, K.; Potisek, M.; Hančević, K.; Radić, T.; Nečemer, M.; Regvar, M. Importance of Soil and Vineyard Management in the Determination of Grapevine Mineral Composition. Sci. Total Environ. 2015, 505, 724–731. [Google Scholar] [CrossRef]
- Zhou, W.; Liu, G.; Xing, W. Variations of Multi-Elements in Wetland Plants on the Tibetan Plateau Are Mainly Determined by Environmental Factors. Ecol. Indic. 2023, 146, 109807. [Google Scholar] [CrossRef]
- Sadiqi, S.; Hamza, M.; Ali, F.; Alam, S.; Shakeela, Q.; Ahmed, S.; Ayaz, A.; Ali, S.; Saqib, S.; Ullah, F.; et al. Molecular Characterization of Bacterial Isolates from Soil Samples and Evaluation of their Antibacterial Potential against MDRS. Molecules 2022, 27, 6281. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. The Key Roles of ROS and RNS as a Signaling Molecule in Plant–Microbe Interactions. Antioxidants 2023, 12, 268. [Google Scholar] [CrossRef]
- Cole, D.W. Soil Nutrient Supply in Natural and Managed Forests. Plant Soil 1995, 168–169, 43–53. [Google Scholar] [CrossRef]
- Hestrin, R.; Hammer, E.C.; Mueller, C.W.; Lehmann, J. Synergies between Mycorrhizal Fungi and Soil Microbial Communities Increase Plant Nitrogen Acquisition. Commun. Biol. 2019, 2, 233. [Google Scholar] [CrossRef] [PubMed]
- Krigas, N.; Tsoktouridis, G.; Anestis, I.; Khabbach, A.; Libiad, M.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Lamchouri, F.; Tsiripidis, I.; Tsiafouli, M.A.; et al. Exploring the Potential of Neglected Local Endemic Plants of Three Mediterranean Regions in the Ornamental Sector: Value Chain Feasibility and Readiness Timescale for Their Sustainable Exploitation. Sustainability 2021, 13, 2539. [Google Scholar] [CrossRef]
- Johnson, C.N.; Balmford, A.; Brook, B.W.; Buettel, J.C.; Galetti, M.; Guangchun, L.; Wilmshurst, J.M. Biodiversity Losses and Conservation Responses in the Anthropocene. Science 2017, 356, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Bhaduri, D.; Sihi, D.; Bhowmik, A.; Verma, B.C.; Munda, S.; Dari, B. A Review on Effective Soil Health Bio-Indicators for Ecosystem Restoration and Sustainability. Front. Microbiol. 2022, 13, 938481. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analysis of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Rhoades, J.D. Salinity: Electrical conductivity and total dissolved solids. In Methods of Soil Analysis, Part 3: Chemical Methods; SSSA Book Series 5; Sparks, D.L., Ed.; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 417–435. [Google Scholar]
- Kuo, S. Phosphorus. In Methods of Soil Analysis, Part 3: Chemical Methods; SSSA Book Series 5; Sparks, D.L., Ed.; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 869–919. [Google Scholar]
- Mulvaney, R.L. Nitrogen-Inorganic Forms. In Methods of Soil Analysis, Part 3: Chemical Methods; SSSA Book Series, 5; Sparks, D.L., Ed.; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 1123–1184. [Google Scholar]
- Thomas, G.W. Exchangeable Cations. In Agronomy Monographs; Page, A.L., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 2015; pp. 159–165. [Google Scholar]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Keren, R. Boron. In Methods of Soil Analysis, Part 3: Chemical Methods; SSSA Book Series, 5; Sparks, D.L., Ed.; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 603–626. [Google Scholar]
- Mills, A.H.; Benton, A.J.; Jones, J.B., Jr. Plant Analysis Handbook II: A Practical Sampling, Preparation, Analysis and Interpretation Guide; MicroMacro Publishing, Inc.: Athens, GA, USA, 1996. [Google Scholar]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis, Part 3: Chemical Methods; SSSA Book Series, 5; Sparks, D.L., Ed.; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Sylvia, D.M. Vesicular-Arbuscular Mycorrhizal Fungi. In Methods of Soil Analysis: Part 2 Microbiological and Biochemical Properties; SSSA Book, Series; Weaver, R.W., Angle, S., Bottomley, P., Bezdicek, D., Smith, S., Tabatabai, A., Wollum, A., Eds.; Soil Science Society of America: Madison, WI, USA, 2018; pp. 351–378. ISBN 978-0-89118-865-0. [Google Scholar]
- Cox, A.E.; Joern, B.C.; Brouder, S.M.; Gao, D. Plant-Available Potassium Assessment with a Modified Sodium Tetraphenylboron Method. Soil Sci. Soc. Am. J. 1999, 63, 902–911. [Google Scholar] [CrossRef]
- Bilias, F.; Barbayiannis, N. Potassium Availability: An Approach Using Thermodynamic Parameters Derived from Quantity-Intensity Relationships. Geoderma 2019, 338, 355–364. [Google Scholar] [CrossRef]
- Wallor, E.; Kersebaum, K.-C.; Lorenz, K.; Gebbers, R. Soil State Variables in Space and Time: First Steps towards Linking Proximal Soil Sensing and Process Modelling. Precision Agric. 2019, 20, 313–334. [Google Scholar] [CrossRef]
- Kravchenko, A.N.; Harrigan, T.M.; Bailey, B.B. Soil electrical conductivity as a covariate to improve the efficiency of field experiments. Trans. ASAE 2005, 48, 1353–1357. [Google Scholar] [CrossRef]
- Zhao, N.; Liu, H.; Wang, Q.; Wang, R.; Xu, Z.; Jiao, C.; Zhu, J.; Yu, G.; He, N. Root Elemental Composition in Chinese Forests: Implications for Biogeochemical Niche Differentiation. Funct. Ecol. 2018, 32, 40–49. [Google Scholar] [CrossRef]
- Zhang, S.-B.; Zhang, J.-L.; Slik, J.W.F.; Cao, K.-F. Leaf Element Concentrations of Terrestrial Plants across China Are Influenced by Taxonomy and the Environment: Leaf Elements of Chinese Biomes. Glob. Ecol. Biogeogr. 2012, 21, 809–818. [Google Scholar] [CrossRef]
- Ibourki, M.; Ait Bouzid, H.; Bijla, L.; Sakar, E.H.; Asdadi, A.; Laknifli, A.; El Hammadi, A.; Gharby, S. Mineral Profiling of Twenty Wild and Cultivated Aromatic and Medicinal Plants Growing in Morocco. Biol. Trace Elem. Res. 2022, 200, 4880–4889. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Urayama, M.; Shinano, T.; Okada, R.; Osaki, M. Application of Ionomics to Plant and Soil in Fields under Long-Term Fertilizer Trials. SpringerPlus 2015, 4, 781. [Google Scholar] [CrossRef]
- Tariq, M.; Mott, C.J.B. Effect of Boron on the Behavior of Nutrients in Soil-Plant Systems—A Review. Asian J. Plant Sci. 2006, 6, 195–202. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2008. [Google Scholar]
- Miransari, M. Arbuscular Mycorrhizal Fungi and Nitrogen Uptake. Arch. Microbiol. 2011, 193, 77–81. [Google Scholar] [CrossRef]
- Karagiannidis, N.; Nikolaou, N.; Ipsilantis, I.; Zioziou, E. Effects of Different N Fertilizers on the Activity of Glomus Mosseae and on Grapevine Nutrition and Berry Composition. Mycorrhiza 2007, 18, 43–50. [Google Scholar] [CrossRef]
- Egerton-Warburton, L.M.; Allen, E.B. Shifts in arbuscular mycorrhizal communities along an anthropogenic nitrogen deposition gradient. Ecol. Appl. 2000, 10, 484–496. [Google Scholar] [CrossRef]
- Dias, T.; Stürmer, S.L.; Chaves, S.; Fidalgo, C.; Tenreiro, R.; Correia, P.; Carvalho, L.; Martins-Loução, M.-A.; Sheppard, L.J.; Cruz, C. Species of Arbuscular Mycorrhizal Fungal Spores Can Indicate Increased Nitrogen Availability in Mediterranean-Type Ecosystems. In Nitrogen Deposition, Critical Loads and Biodiversity; Sutton, M.A., Mason, K.E., Sheppard, L.J., Sverdrup, H., Haeuber, R., Hicks, W.K., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 259–266. ISBN 978-94-007-7938-9. [Google Scholar]
- Zheng, Z.; Ma, P.; Li, J.; Ren, L.; Bai, W.; Tian, Q.; Sun, W.; Zhang, W. Arbuscular Mycorrhizal Fungal Communities Associated with Two Dominant Species Differ in Their Responses to Long-term Nitrogen Addition in Temperate Grasslands. Funct. Ecol. 2018, 32, 1575–1588. [Google Scholar] [CrossRef]
- Karaarslan, E.; Uyanöz, R.; Doğu, S. Morphological Identification of Vesicular-Arbuscular Mycorrhiza on Bulbous Plants (Taurus Mountain in Turkey). Arch. Biol. Sci. 2015, 67, 411–426. [Google Scholar] [CrossRef]
- Melo, C.D.; Walker, C.; Krüger, C.; Borges, P.A.V.; Luna, S.; Mendonça, D.; Fonseca, H.M.A.C.; Machado, A.C. Environmental Factors Driving Arbuscular Mycorrhizal Fungal Communities Associated with Endemic Woody Plant Picconiaazorica on Native Forest of Azores. Ann. Microbiol. 2019, 69, 1309–1327. [Google Scholar] [CrossRef]
- Black, C.A. Soil Fertility Evaluation and Control, 1st ed.; CRC Press: Boca Raton, FL, USA, 1993; ISBN 978-0-429-15979-4. [Google Scholar]
- van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal Fungal Diversity Determines Plant Biodiversity, Ecosystem Variability and Productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Jakobsen, S.T. Interaction between Plant Nutrients: IV. Interaction between Calcium and Phosphate. Acta Agric. Scand. B Soil Plant Sci. 1993, 43, 6–10. [Google Scholar] [CrossRef]
- Xie, K.; Cakmak, I.; Wang, S.; Zhang, F.; Guo, S. Synergistic and Antagonistic Interactions between Potassium and Magnesium in Higher Plants. Crop J. 2021, 9, 249–256. [Google Scholar] [CrossRef]
- Smeti, E.; Tsirtsis, G.; Skoulikidis, N.T. Geology Can Drive the Diversity–Ecosystem Functioning Relationship in River Benthic Diatoms by Selecting for Species Functional Traits. Biology 2023, 12, 81. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; Pipinis, E.; Kostas, S.; Stagiopoulou, R.; Gitsa, K.; Dariotis, E.; Avramakis, M.; Samartza, I.; Plastiras, I.; Kriemadi, E.; et al. Influence of Temperature on Seed Germination of Five Wild-Growing Tulipa Species of Greece Associated with Their Ecological Profiles: Implications for Conservation and Cultivation. Plants 2023, 12, 1574. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tulipa Species | IPEN Accession | Altitude (m) | Phytogeographical Unit of Greece | Latitude (North) | Longitude (East) |
|---|---|---|---|---|---|
| T. agenensis (a) | GR-BBGK-1-22,2 | 101 | North Aegean Islands | 38.33376 | 26.081543 |
| T. agenensis (b) | GR-BBGK-1-22,15 | 99 | North Aegean Islands | 38.333713 | 26.081833 |
| T. australis (a) | GR-BBGK-1-21,115 | 1554 | Mainland | 40.096552 | 21.114582 |
| T. australis (b) | GR-BBGK-1-21,116 | 1856 | Mainland | 40.096552 | 21.114582 |
| T. australis (c) | GR-BBGK-1-22,54 | 477 | Mainland | 37.809379 | 23.941191 |
| T. bakeri (a) | GR-BBGK-1-21,8 | 1057 | Crete | 35.330758 | 23.896977 |
| T. bakeri (b) | GR-BBGK-1-22,28 | 1065 | Crete | 35.331772 | 23.907389 |
| T. bithynica (a) | GR-BBGK-1-22,19 | 870 | North Aegean Islands | 39.161205 | 26.066789 |
| T. clusiana (a) | GR-BBGK-1-22,1 | 422 | North Aegean Islands | 38.304548 | 26.056388 |
| T. cretica (a) | GR-BBGK-1-21,79 | 12 | Crete | 34.957198 | 25.099697 |
| T. cretica (b) | GR-BBGK-1-21,83 | 57 | Crete | 35.549880 | 24.150415 |
| T. cretica (c) | GR-BBGK-1- 22,7 | 131 | Crete | 35.551054 | 24.147271 |
| T. cretica (d) | GR-BBGK-1-22,9 | 158 | Crete | 35.222128 | 26.212745 |
| T. cretica (e) | GR-BBGK-1-22,21 | 974 | Crete | 34.957373 | 25.100580 |
| T. doerfleri (a) | GR-BBGK-1-21,106 | 743 | Crete | 35.207935 | 24.560692 |
| T. doerfleri (b) | GR-BBGK-1-22,33 | 773 | Crete | 35.213769 | 24.567137 |
| T. goulimyi (a) | GR-BBGK-1-21,39 | 415 | Mainland * | 36.16800 | 22.96600 |
| T. goulimyi (b) | GR-BBGK-1-21,4 | 411 | Mainland * | 36.2531000 | 22.9677000 |
| T. goulimyi (c) | GR-BBGK-1-21,41 | 616 | Mainland | 36.8283000 | 22.9475000 |
| T. goulimyi (d) | GR-BBGK-1-22,25 | 177 | Crete | 35.503031 | 23.754020 |
| T. hageri (a) | GR-BBGK-1-21,67 | 516 | Mainland | 40.799011 | 23.389939 |
| T. hageri (b) | GR-BBGK-1-21,102 | 428 | Mainland | 40.421873 | 23.694688 |
| T. hageri (c) | GR-BBGK-1-22,55 | 544 | Mainland | 38.190514 | 23.794406 |
| T. orphanidea | GR-BBGK-1-21,44 | 876 | Mainland | 37.30900 | 22.42200 |
| T. raddii (a) | GR-BBGK-1-21,285 | 183 | North Aegean Islands | 38.28275 | 26.07461 |
| T. raddii (b) | GR-BBGK-1-22,3 | 381 | North Aegean Islands | 38.319157 | 26.055302 |
| T. saxatilis (a) | GR-BBGK-1-21,105 | 721 | Crete | 35.213210 | 24.564025 |
| T. saxatilis (b) | GR-BBGK-1-22,8 | 518 | Crete | 35.419784 | 23.740193 |
| T. saxatilis (c) | GR-BBGK-1-22,14 | 95 | Crete | 35.074496 | 25.806127 |
| T, saxatilis (d) | GR-BBGK-1-22,2 | 599 | Crete | 35.176982 | 24.997673 |
| T. saxatilis (e) | GR-BBGK-1-22,27 | 531 | Crete | 35.419784 | 23.740193 |
| T. saxatilis (f) | GR-BBGK-1-22,43 | 830 | Crete | 35.1696440 | 25.4882520 |
| T. undulatifolia (a) | GR-BBGK-1-22,4 | 25 | North Aegean Islands | 38.203085 | 26.030258 |
| T. undulatifolia (b) | GR-BBGK-1-22,11 | 445 | Mainland | 38.090000 | 23.230000 |
| Tulips (Tulipa spp.) | Phytogeographical Status | Chorological Status | Threatened Status | Altitudinal Class (Altitudinal Range in m) | Habitat Types | Rock-Dwelling | Segetal | Flowering |
|---|---|---|---|---|---|---|---|---|
| T. agenensis | Naturalized alien | Irano-Turanian (Chios Island, Greece) | No | Lowland (0–300) | Agricultural 1 | No | Yes | Early |
| T. australis | Wild-growing native | Mediterranean-SW Asiatic | No | Intermediate to high (500–2000) | Natural 2 | Yes | No | Late |
| T. bakeri | Wild-growing native | Greek endemic (Crete) | Yes (CR) | Intermediate (700–1300) | Agricultural and natural 3 | Yes | No | Mid-spring |
| T. bithynica | Wild-growing native | Subendemic (Lesvos, Greece-Anatolia) | No | Lowland (200–800) | Agricultural and natural 4 | No | No | Mid-spring |
| T. clusiana | Naturalized alien | Irano-Turanian (Chios Island, Greece) | No | Lowland (100–600) | Agricultural 5 | No | No | Early |
| T. cretica | Wild-growing native | Greek endemic (Crete) | Yes (EN) | Lowland to high (0–2100) | Natural 6 | Yes | No | Early |
| T. doerfleri | Wild-growing native | Greek endemic (Crete) | Yes (CR) | Lowland (400–800) | Agricultural 7 | No | Yes | Mid-spring |
| T. goulimyi | Wild-growing native | Greek endemic (Peloponnese, nearby islands to Crete) | Yes (VU) | Lowland (0–900) | Natural 8 | No | No | Early |
| T. hageri | Wild-growing native | Greek endemic (Sterea Hellas, Peloponnese and north Greece) | Yes (EN) | Lowland to intemediate (100–1200) | Agricultural and natural 9 | No | Yes | Mid-spring |
| T. orphanidea | Wild-growing native | Greek endemic (Sterea Hellas, Peloponnese) | Yes (EN) | Lowland to intemediate (700–1600) | Agricultural and natural 10 | No | Yes | Early |
| T. raddii | Naturalized alien | East Mediterranean (Chios island) | No | Lowland (0–400) | Agricultural 11 | No | Yes | Early |
| T. saxatilis | Wild-growing native | Subendemic (South Aegean, Anatolia) | No | Lowland to intemediate (200–1300) | Agricultural and natural 12 | Yes | No | Early |
| T. undulatifolia | Wild-growing native | Subendemic (Balkan-Anatolia) | Yes (VU) | Lowland (100–800) | Agricultural 13 and natural | No | Yes | Early |
| Variable | Min. | Max. | Median | Mean | SD | CV% |
|---|---|---|---|---|---|---|
| Sand (%) | 20.8 | 78.2 | 49.6 | 48.9 | 13.8 | 28.2% |
| Silt (%) | 10.4 | 53.2 | 26.7 | 28.1 | 9.3 | 33.1% |
| Clay (%) | 6.4 | 47.0 | 18.2 | 23.0 | 12.8 | 55.4% |
| pH | 5.20 | 7.90 | 7.65 | 7.21 | 0.81 | 11.2% |
| CaCO3 (%) | 0.0 | 62.4 | 2.1 | 9.0 | 15.4 | 170.4% |
| OC (%) | 0.64 | 12.12 | 1.39 | 2.39 | 2.30 | 96.3% |
| Total N (%) | 0.08 | 0.80 | 0.16 | 0.22 | 0.16 | 72.6% |
| C/N | 6.50 | 15.30 | 9.60 | 9.60 | 2.00 | 20.8% |
| CEC (cmolc kg–1) | 7.2 | 56.9 | 24.4 | 24.3 | 12.5 | 51.7% |
| ECse (ds m−1) | 0.14 | 2.64 | 0.45 | 0.57 | 0.50 | 84.7% |
| Kex (mg kg–1) | 34 | 900 | 258 | 275 | 214 | 77.8% |
| Naex (mg kg–1) | 12.7 | 96.3 | 49.2 | 47.9 | 20.5 | 42.9% |
| Caex (mg kg–1) | 616 | 8472 | 3856 | 3663 | 1981 | 54.1% |
| Mgex (mg kg–1) | 97 | 884 | 225 | 357 | 256 | 71.6% |
| NO3-N (mg kg–1) | 1.1 | 98.7 | 7.3 | 13.0 | 17.5 | 134.5% |
| NH4-N (mg kg–1) | 2.6 | 48.3 | 11.8 | 12.5 | 8.2 | 65.1% |
| P-Olsen (mg kg–1) | 0.90 | 45.20 | 4.45 | 6.16 | 7.95 | 129.0% |
| Cu-DTPA (mg kg–1) | 0.64 | 2.77 | 1.26 | 1.42 | 0.60 | 39.4% |
| Zn-DTPA (mg kg–1) | 0.31 | 7.09 | 0.71 | 1.33 | 1.50 | 114.4% |
| Fe-DTPA (mg kg–1) | 3.6 | 95.0 | 20.5 | 27.7 | 23.7 | 85.7% |
| Mn-DTPA (mg kg–1) | 5.4 | 115.2 | 20.0 | 27.4 | 21.7 | 79.0% |
| B-HW (mg kg–1) | 0.25 | 1.81 | 0.56 | 0.67 | 0.40 | 53.6% |
| N | K | Na | Ca | Mg | Cu | Zn | Fe | Mn | P | |
|---|---|---|---|---|---|---|---|---|---|---|
| K | 0.31 a | |||||||||
| Na | 0.21 | −0.04 | ||||||||
| Ca | −0.17 | 0.09 | 0.38 * | |||||||
| Mg | −0.22 | −0.09 | 0.15 | 0.49 ** | ||||||
| Cu | 0.35 * | 0.11 | −0.17 | −0.14 | 0.10 | |||||
| Zn | 0.44 ** | 0.19 | 0.07 | −0.01 | −0.24 | 0.09 | ||||
| Fe | −0.28 | −0.36 * | 0.10 | 0.14 | 0.39 * | −0.34 | −0.10 | |||
| Mn | 0.01 | −0.31 a | 0.11 | −0.10 | 0.17 | −0.23 | 0.07 | 0.62 *** | ||
| P | 0.39 * | 0.40 * | −0.40 * | −0.57 *** | −0.13 | 0.46 ** | 0.22 | −0.27 | 0.02 | |
| B | −0.19 | −0.16 | 0.57 *** | 0.65 *** | 0.49 | −0.20 | −0.04 | 0.37 * | 0.15 | −0.65 *** |
| Dependent Variable (Y) | Independent Variable (X) | Model | r2 % | p-Value |
|---|---|---|---|---|
| N | Clay | Simple linear | 32.2 | 0.0005 |
| P | ECse, CEC | Multiple reciprocal-Y | 67.1 | 0.0000 |
| K | CaCO3, OC%, CEC | Multiple linear | 58.2 | 0.0000 |
| Ca | pH, ECse | Multiple linear | 49.6 | 0.0000 |
| Mg | ECse | Double squared | 43.1 | 0.0000 |
| Na | Naex | Double reciprocal | 56.4 | 0.0000 |
| Cu | Cu-DTPA | Double squared | 22.1 | 0.0050 |
| Zn | Zn-DTPA, pH, ECse | Multiple linear | 57.5 | 0.0000 |
| Fe | pH, OC% | Multiple linear | 21.6 | 0.0258 |
| Mn | pH, OC% | Multiple linear | 47.5 | 0.0001 |
| B | ECse | Squared-Y square root-X | 38.3 | 0.0001 |
| Location | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ν | Ρ | Κ | Ca | Mg | Na | |||||||
| (g kg−1) | ||||||||||||
| North Aegean Islands | 18.3 a | (1.8) | 1.49 a | (0.12) | 15.4 a | (1.8) | 3.94 b | (0.91) | 1.47 a | (0.17) | 0.93 b | (0.22) |
| Crete Island | 12.0 b | (1.6) | 0.68 b | (0.09) | 10.0 b | (1.0) | 6.27 a | (0.65) | 1.65 a | (0.13) | 1.63 a | (0.23) |
| Mainland Greece | 10.3 b | (1.5) | 1.24 a | (0.20) | 9.4 b | (1.1) | 3.27 b | (0.56) | 1.52 a | (0.13) | 0.43 b | (0.09) |
| p F-test | 0.021 | <0.001 | 0.011 | 0.007 | NS | <0.001 | ||||||
| Cu | Zn | Fe | Mn | B | AMF spore morphotypes | |||||||
| (mg kg−1) | ||||||||||||
| North Aegean Islands | 7.74 a | (0.91) | 27.9 a | (0.8) | 154 b | (44) | 16.6 b | (2.0) | 19.5 ab | (1. 8) | 4.50 a | (0.62) |
| Crete Island | 4.24 b | (0.35) | 26.5 a | (2.8) | 414 a | (62) | 27.1 a | (3.1) | 23.1 a | (1.1) | 2.75 b | (0.49) |
| Mainland Greece | 5.23 b | (0.31) | 33.0 a | (7.1) | 316 ab | (28) | 19.6 ab | (2.2) | 17.3 b | (1.9) | 2.18 b | (0.46) |
| p F-test | <0.001 | NS | 0.023 | 0.048 | 0.022 | 0.044 | ||||||
| Location | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE |
| Clay (%) | pH | CaCO3 (%) | OC (%) | N-Total (%) | CEC (cmolc kg−1) | ECse (ds m−1) | ||||||||
| North Aegean Islands | 31.9 a | (4.5) | 7.51 a | (0.22) | 21.6 a | (7.1) | 1.86 a | (0.62) | 0.18 a | (0.04) | 35.2 a | (1.6) | 0.61 a | (0.11) |
| Crete Island | 17.1 b | (2.6) | 7.29 a | (0.17) | 8.78 ab | (4.03) | 2.81 a | (0.73) | 0.25 a | (0.05) | 21.6 b | (3.6) | 0.71 a | (0.15) |
| Mainland Greece | 25.9 ab | (3.9) | 6.90 a | (0.31) | 1.40 b | (0.64) | 2.10 a | (0.48) | 0.21 a | (0.04) | 21.1 b | (3.0) | 0.33 a | (0.06) |
| p F-test | 0.020 | NS | 0.020 | NS | NS | 0.029 | NS | |||||||
| Location | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | ||
| NO3-N | NH4-N | P-Olsen | Κ-ex | Ca-ex | Mg-ex | |||||||||
| (mg kg−1) | ||||||||||||||
| North Aegean Islands | 13.6 a | (4.3) | 7.81 b | (1.14) | 6.10 a | (2.55) | 473 a | (114) | 5103 a | (222) | 627 a | (104) | ||
| Crete Island | 15.4 a | (5.9) | 10.9 ab | (1.5) | 6.81 a | (2.71) | 227 b | (42) | 3665 ab | (563) | 215 c | (25) | ||
| Mainland Greece | 9.23 a | (2.62) | 14.5 a | (1.3) | 5.25 a | (0.58) | 218 b | (43) | 2744 b | (501) | 392 b | (81) | ||
| p F-test | NS | 0.020 | NS | 0.043 | <0.001 | 0.012 | ||||||||
| Na-ex | Cu-DTPA | Zn-DTPA | Fe-DTPA | Mn-DTPA | B-HW | |||||||||
| (mg kg−1) | ||||||||||||||
| North Aegean Islands | 54.0 a | (5.1) | 1.87 a | (0.24) | 1.15 a | (0.30) | 17.2 a | (5.3) | 14.1 b | (2.9) | 0.74 a | (0.18) | ||
| Crete Island | 52.9 a | (4.62) | 1.26 b | (0.09) | 1.46 a | (0.38) | 27.2 a | (5.6) | 27.8 ab | (3.4) | 0.69 a | (0.08) | ||
| Mainland Greece | 31.9 b | (5.9) | 1.39 ab | (0.20) | 1.26 a | (0.59) | 35.0 a | (8.7) | 35.5 a | (9.7) | 0.60 a | (0.11) | ||
| p F-test | <0.001 | 0.049 | NS | NS | NS (0.123) | NS | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilias, F.; Karagianni, A.-G.; Ipsilantis, I.; Samartza, I.; Krigas, N.; Tsoktouridis, G.; Matsi, T. Adaptability of Wild-Growing Tulips of Greece: Uncovering Relationships between Soil Properties, Rhizosphere Fungal Morphotypes and Nutrient Content Profiles. Biology 2023, 12, 605. https://doi.org/10.3390/biology12040605
Bilias F, Karagianni A-G, Ipsilantis I, Samartza I, Krigas N, Tsoktouridis G, Matsi T. Adaptability of Wild-Growing Tulips of Greece: Uncovering Relationships between Soil Properties, Rhizosphere Fungal Morphotypes and Nutrient Content Profiles. Biology. 2023; 12(4):605. https://doi.org/10.3390/biology12040605
Chicago/Turabian StyleBilias, Fotis, Anastasia-Garyfallia Karagianni, Ioannis Ipsilantis, Ioulietta Samartza, Nikos Krigas, Georgios Tsoktouridis, and Theodora Matsi. 2023. "Adaptability of Wild-Growing Tulips of Greece: Uncovering Relationships between Soil Properties, Rhizosphere Fungal Morphotypes and Nutrient Content Profiles" Biology 12, no. 4: 605. https://doi.org/10.3390/biology12040605
APA StyleBilias, F., Karagianni, A. -G., Ipsilantis, I., Samartza, I., Krigas, N., Tsoktouridis, G., & Matsi, T. (2023). Adaptability of Wild-Growing Tulips of Greece: Uncovering Relationships between Soil Properties, Rhizosphere Fungal Morphotypes and Nutrient Content Profiles. Biology, 12(4), 605. https://doi.org/10.3390/biology12040605