
Development and Characterization of Inducible Astrocyte-Specific Aromatase Knockout Mice

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Generation of a Mouse Model of Inducible Astrocyte-Specific Aromatase Knockout
2.2. Induction of Aromatase Deletion in the GFAP-ARO-iKO Mice
2.3. Global Cerebral Ischemia
2.4. Tissue Sample Collection and Astrocyte Purification
2.5. Cell Lysates Preparation and Western Blotting
2.6. Immunohistochemistry (IHC)
2.7. Statistical Analyses
3. Results
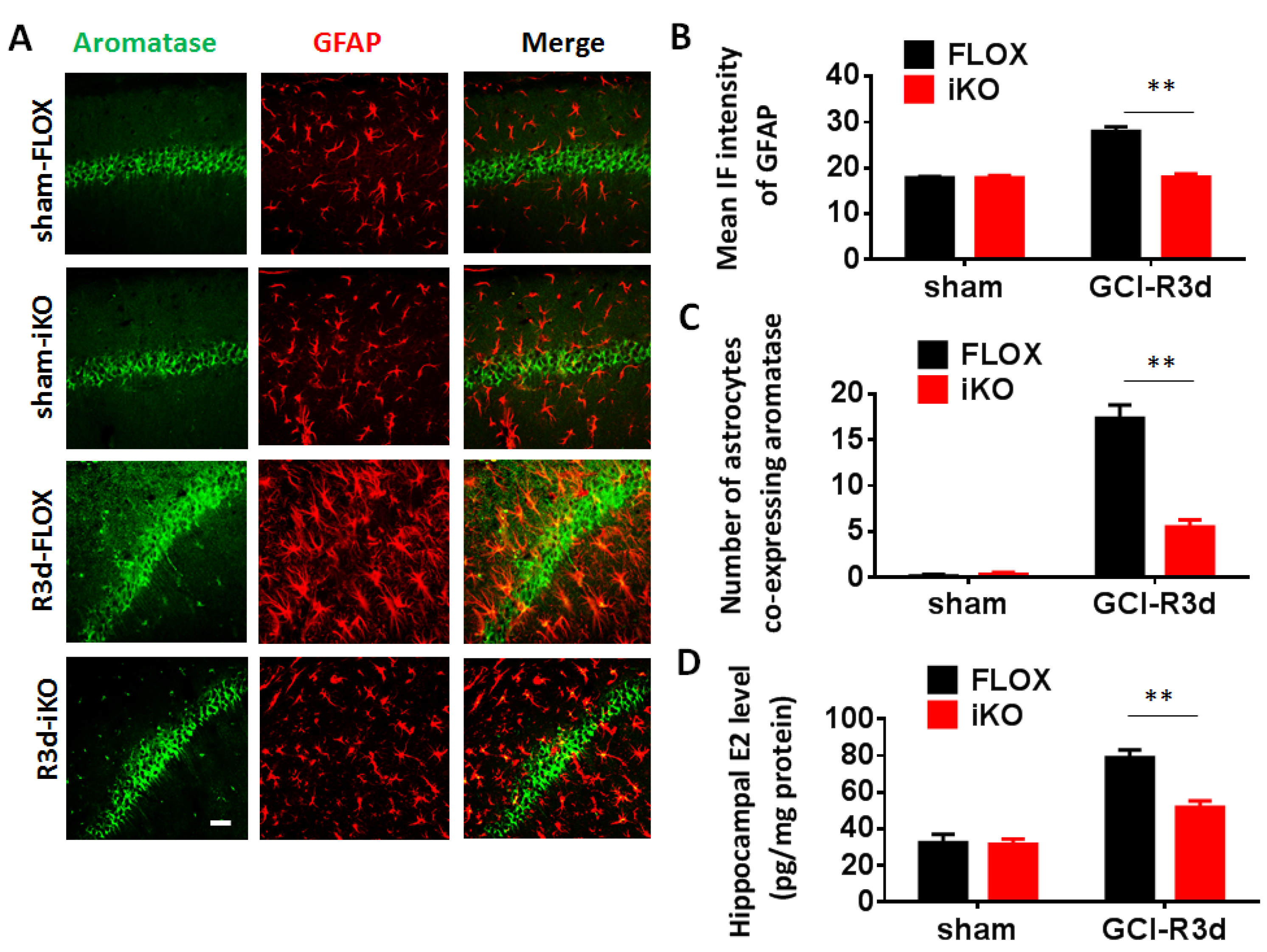
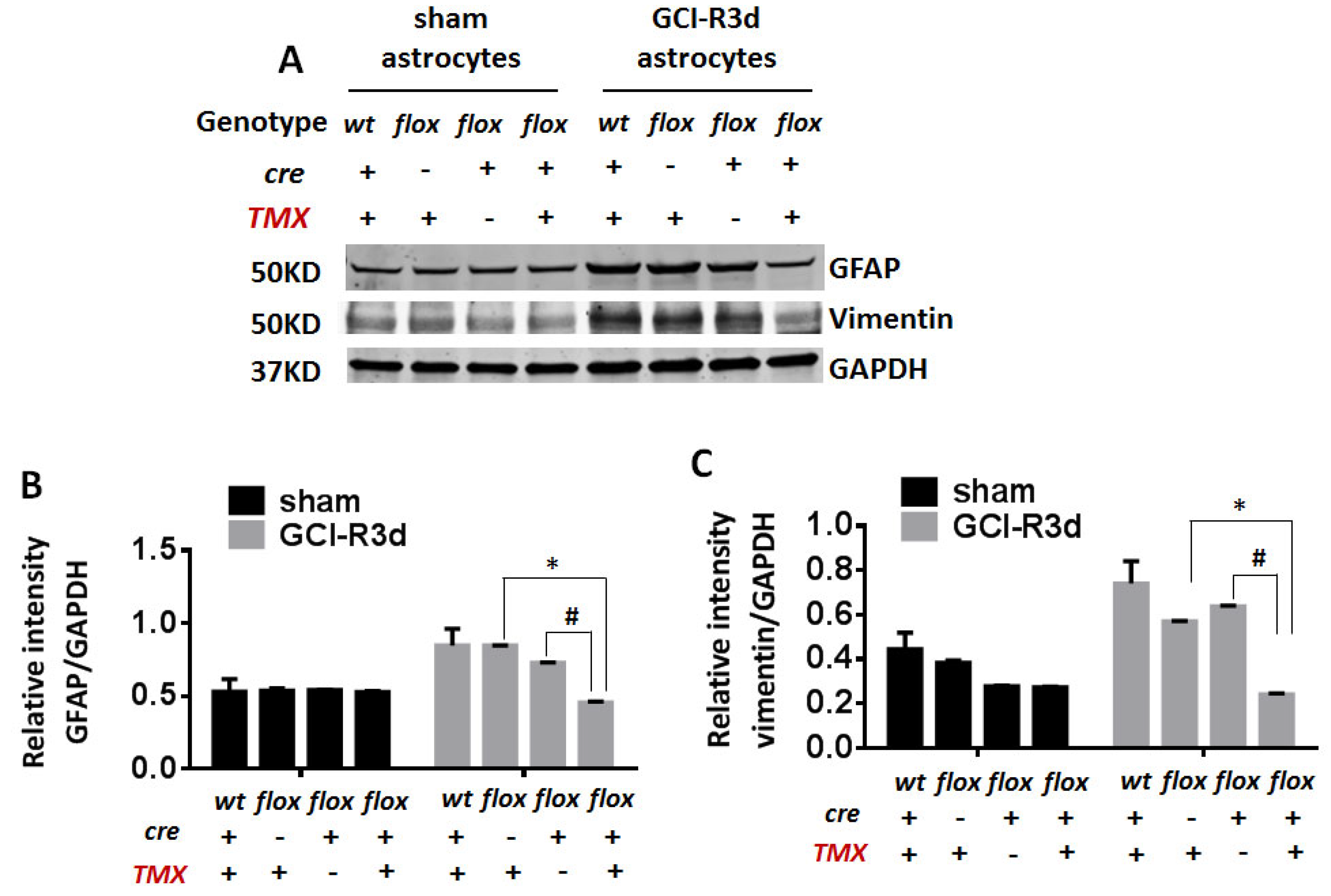
Generation and Characterization of GFAP-ARO-iKO Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blakemore, J.; Naftolin, F. Aromatase: Contributions to Physiology and Disease in Women and Men. Physiology 2016, 31, 258–269. [Google Scholar] [CrossRef]
- Simpson, E.R.; Clyne, C.; Rubin, G.; Boon, W.C.; Robertson, K.; Britt, K.; Speed, C.; Jones, M. Aromatase--A brief overview. Annu. Rev. Physiol. 2002, 64, 93–127. [Google Scholar] [CrossRef]
- Brocca, M.E.; Garcia-Segura, L.M. Non-reproductive Functions of Aromatase in the Central Nervous System under Physiological and Pathological Conditions. Cell. Mol. Neurobiol. 2018, 39, 473–481. [Google Scholar] [CrossRef]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A Transmembrane Intracellular Estrogen Receptor Mediates Rapid Cell Signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef]
- Kumar, A.; Bean, L.A.; Rani, A.; Jackson, T.; Foster, T.C. Contribution of estrogen receptor subtypes, ERalpha, ERbeta, and GPER1 in rapid estradiol-mediated enhancement of hippocampal synaptic transmission in mice. Hippocampus 2015, 25, 1556–1566. [Google Scholar] [CrossRef]
- Spencer-Segal, J.L.; Tsuda, M.C.; Mattei, L.; Waters, E.M.; Romeo, R.D.; Milner, T.A.; McEwen, B.S.; Ogawa, S. Estradiol acts via estrogen receptors alpha and beta on pathways important for synaptic plasticity in the mouse hippocampal formation. Neuroscience 2012, 202, 131–146. [Google Scholar] [CrossRef]
- Engler-Chiurazzi, E.B.; Brown, C.M.; Povroznik, J.M.; Simpkins, J.W. Estrogens as neuroprotectants: Estrogenic actions in the context of cognitive aging and brain injury. Prog. Neurobiol. 2016, 157, 188–211. [Google Scholar] [CrossRef]
- Brann, D.W.; Lu, Y.; Wang, J.; Zhang, Q.; Thakkar, R.; Sareddy, G.R.; Pratap, U.P.; Tekmal, R.R.; Vadlamudi, R.K. Brain-derived estrogen and neural function. Neurosci. Biobehav. Rev. 2021, 132, 793–817. [Google Scholar] [CrossRef]
- Naftolin, F.; Ryan, K.J.; Petro, Z. Aromatization of androstenedione by the diencephalon. J. Clin. Endocrinol. Metab. 1971, 33, 368–370. [Google Scholar] [CrossRef]
- Callard, G.V.; Petro, Z.; Ryan, K.J. Phylogenetic Distribution of Aromatase and Other Androgen-Converting Enzymes in the Central Nervous System. Endocrinology 1978, 103, 2283–2290. [Google Scholar] [CrossRef]
- Stoffel-Wagner, B.; Watzka, M.; Schramm, J.; Bidlingmaier, F.; Klingmüller, D. Expression of CYP19 (aromatase) mRNA in different areas of the human brain. J. Steroid Biochem. Mol. Biol. 1999, 70, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-G.; Wang, R.; Tang, H.; Dong, Y.; Chan, A.; Sareddy, G.R.; Vadlamudi, R.K.; Brann, D.W. Brain-derived estrogen exerts anti-inflammatory and neuroprotective actions in the rat hippocampus. Mol. Cell. Endocrinol. 2014, 389, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Azcoitia, I.; Yague, J.G.; Garcia-Segura, L.M. Estradiol synthesis within the human brain. Neuroscience 2011, 191, 139–147. [Google Scholar] [CrossRef]
- Yague, J.G.; Muñoz, A.; de Monasterio-Schrader, P.; DeFelipe, J.; Garcia-Segura, L.M.; Azcoitia, I. Aromatase expression in the human temporal cortex. Neuroscience 2006, 138, 389–401. [Google Scholar] [CrossRef]
- Yague, J.G.; Azcoitia, I.; DeFelipe, J.; Garcia-Segura, L.M.; Muñoz, A. Aromatase expression in the normal and epileptic human hippocampus. Brain Res. 2010, 1315, 41–52. [Google Scholar] [CrossRef]
- Naftolin, F.; Horvath, T.L.; Jakab, R.L.; Leranth, C.; Harada, N.; Balthazart, J. Aromatase Immunoreactivity in Axon Terminals of the Vertebrate Brain. An immunocytochemical study on quail, rat, monkey and human tissues. Neuroendocrinology 1996, 63, 149–155. [Google Scholar] [CrossRef]
- Garcia-Segura, L.M.; Wozniak, A.; Azcoitia, I.; Rodriguez, J.R.; Hutchison, R.E.; Hutchison, J.B. Aromatase expression by astrocytes after brain injury: Implications for local estrogen formation in brain repair. Neuroscience 1999, 89, 567–578. [Google Scholar] [CrossRef]
- Lu, Y.; Sareddy, G.R.; Wang, J.; Zhang, Q.; Tang, F.-L.; Pratap, U.P.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Neuron-Derived Estrogen Is Critical for Astrocyte Activation and Neuroprotection of the Ischemic Brain. J. Neurosci. 2020, 40, 7355–7374. [Google Scholar] [CrossRef]
- Mehos, C.J.; Nelson, L.H.; Saldanha, C.J. A Quantification of the Injury-Induced Changes in Central Aromatase, Oestrogenic Milieu and Steroid Receptor Expression in the Zebra Finch. J. Neuroendocr. 2016, 28, 12348. [Google Scholar] [CrossRef]
- Peterson, R.S.; Saldanha, C.J.; Schlinger, B.A. Rapid Upregulation of Aromatase mRNA and Protein Following Neural Injury in the Zebra Finch (Taeniopygia guttata ). J. Neuroendocr. 2001, 13, 317–323. [Google Scholar] [CrossRef]
- Wang, J.; Sareddy, G.R.; Lu, Y.; Pratap, U.P.; Tang, F.; Greene, K.M.; Meyre, P.L.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Astrocyte-Derived Estrogen Regulates Reactive Astrogliosis and is Neuroprotective following Ischemic Brain Injury. J. Neurosci. 2020, 40, 9751–9771. [Google Scholar] [CrossRef]
- Roselli, C.E.; Ellinwood, W.E.; Resko, J.A. Regulation of Brain Aromatase Activity in Rats. Endocrinology 1984, 114, 192–200. [Google Scholar] [CrossRef]
- Biegon, A.; Alexoff, D.L.; Kim, S.W.; Logan, J.; Pareto, D.; Schlyer, D.; Wang, G.J.; Fowler, J.S. Aromatase imaging with [N-methyl-11C]vorozole PET in healthy men and women. J. Nucl. Med. 2015, 56, 580–585. [Google Scholar] [CrossRef]
- Naftolin, F. Brain aromatization of androgens. J. Reprod. Med. 1994, 39, 257–261. [Google Scholar]
- Balthazart, J.; Baillien, M.; Charlier, T.D.; Ball, G.F. Calcium-dependent phosphorylation processes control brain aromatase in quail. Eur. J. Neurosci. 2003, 17, 1591–1606. [Google Scholar] [CrossRef]
- Balthazart, J.; Baillien, M.; Ball, G.F. Rapid Control of Brain Aromatase Activity by Glutamatergic Inputs. Endocrinology 2006, 147, 359–366. [Google Scholar] [CrossRef]
- Kretz, O.; Fester, L.; Wehrenberg, U.; Zhou, L.; Brauckmann, S.; Zhao, S.; Prange-Kiel, J.; Naumann, T.; Jarry, H.; Frotscher, M.; et al. Hippocampal Synapses Depend on Hippocampal Estrogen Synthesis. J. Neurosci. 2004, 24, 5913–5921. [Google Scholar] [CrossRef]
- Grassi, S.; Tozzi, A.; Costa, C.; Tantucci, M.; Colcelli, E.; Scarduzio, M.; Calabresi, P.; Pettorossi, V.E. Neural 17beta-estradiol facilitates long-term potentiation in the hippocampal CA1 region. Neuroscience 2011, 192, 67–73. [Google Scholar] [CrossRef]
- Azcoitia, I.; Sierra, A.; Veiga, S.; Honda, S.-I.; Harada, N.; Garcia-Segura, L.M. Brain aromatase is neuroprotective. J. Neurobiol. 2001, 47, 318–329. [Google Scholar] [CrossRef]
- Wynne, R.D.; Saldanha, C.J. Glial Aromatization Decreases Neural Injury in the Zebra Finch (Taeniopygia guttata): Influence on Apoptosis. J. Neuroendocr. 2004, 16, 676–683. [Google Scholar] [CrossRef]
- Wynne, R.D.; Walters, B.J.; Bailey, D.J.; Saldanha, C.J. Inhibition of injury-induced glial aromatase reveals a wave of secondary degeneration in the songbird brain. Glia 2007, 56, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Brann, D.W.; Lu, Y.; Wang, J.; Sareddy, G.R.; Pratap, U.P.; Zhang, Q.; Tekmal, R.R.; Vadlamudi, R.K. Neuron-Derived Estrogen—A Key Neuromodulator in Synaptic Function and Memory. Int. J. Mol. Sci. 2021, 22, 13242. [Google Scholar] [CrossRef] [PubMed]
- McCullough, L.D.; Blizzard, K.; Simpson, E.R.; Öz, O.K.; Hurn, P.D. Aromatase Cytochrome P450 and Extragonadal Estrogen Play a Role in Ischemic Neuroprotection. J. Neurosci. 2003, 23, 8701–8705. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Sareddy, G.R.; Wang, J.; Wang, R.; Li, Y.; Dong, Y.; Zhang, Q.; Liu, J.; O’Connor, J.C.; Xu, J.; et al. Neuron-Derived Estrogen Regulates Synaptic Plasticity and Memory. J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 2792–2809. [Google Scholar] [CrossRef]
- Casper, K.B.; McCarthy, K.D. GFAP-positive progenitor cells produce neurons and oligodendrocytes throughout the CNS. Mol. Cell. Neurosci. 2006, 31, 676–684. [Google Scholar] [CrossRef]
- Jahn, H.M.; Kasakow, C.V.; Helfer, A.; Michely, J.; Verkhratsky, A.; Maurer, H.H.; Scheller, A.; Kirchhoff, F. Refined protocols of tamoxifen injection for inducible DNA recombination in mouse astroglia. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Cheong, R.Y.; Porteous, R.; Chambon, P.; Abraham, I.; Herbison, A.E. Effects of neuron-specific estrogen receptor (ER) alpha and ERbeta deletion on the acute estrogen negative feedback mechanism in adult female mice. Endocrinology 2014, 155, 1418–1427. [Google Scholar] [CrossRef]
- de Pablo, Y.; Nilsson, M.; Pekna, M.; Pekny, M. Intermediate filaments are important for astrocyte response to oxidative stress induced by oxygen–glucose deprivation and reperfusion. Histochemistry 2013, 140, 81–91. [Google Scholar] [CrossRef]
- Zhao, S.-C.; Ma, L.-S.; Chu, Z.-H.; Xu, H.; Wu, W.-Q.; Liu, F. Regulation of microglial activation in stroke. Acta Pharmacol. Sin. 2017, 38, 445–458. [Google Scholar] [CrossRef]
- Li, T.; Zhao, J.; Xie, W.; Yuan, W.; Guo, J.; Pang, S.; Gan, W.-B.; Gómez-Nicola, D.; Zhang, S. Specific depletion of resident microglia in the early stage of stroke reduces cerebral ischemic damage. J. Neuroinflammation 2021, 18, 1–15. [Google Scholar] [CrossRef]
- McEwen, B.; Akama, K.; Alves, S.; Brake, W.G.; Bulloch, K.; Lee, S.; Li, C.; Yuen, G.; Milner, T.A. Tracking the estrogen receptor in neurons: Implications for estrogen-induced synapse formation. Proc. Natl. Acad. Sci. USA 2001, 98, 7093–7100. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.M.; Thompson, L.I.; Patel, P.; Gonzales, A.D.; Ye, H.; Filardo, E.J.; Clegg, D.J.; Gorecka, J.; Akama, K.T.; McEwen, B.S.; et al. G-Protein-Coupled Estrogen Receptor 1 Is Anatomically Positioned to Modulate Synaptic Plasticity in the Mouse Hippocampus. J. Neurosci. 2015, 35, 2384–2397. [Google Scholar] [CrossRef] [PubMed]
- Engler-Chiurazzi, E.B.; Singh, M.; Simpkins, J.W. Reprint of: From the 90׳s to now: A brief historical perspective on more than two decades of estrogen neuroprotection. Brain Res. 2016, 1645, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.-Z.; Shi, F.; Hu, J.; He, S.-M.; Ding, Q.; Ma, L.-T. GPER1 mediates estrogen-induced neuroprotection against oxygen-glucose deprivation in the primary hippocampal neurons. Neuroscience 2016, 328, 117–126. [Google Scholar] [CrossRef]
- Traystman, R.J. Animal Models of Focal and Global Cerebral Ischemia. ILAR J. 2003, 44, 85–95. [Google Scholar] [CrossRef]
- McGinn, M.J.; Povlishock, J.T. Pathophysiology of Traumatic Brain Injury. Neurosurg. Clin. N. Am. 2016, 27, 397–407. [Google Scholar] [CrossRef]
- Arevalo, M.-A.; Azcoitia, I.; Garcia-Segura, L.M. The neuroprotective actions of oestradiol and oestrogen receptors. Nat. Rev. Neurosci. 2014, 16, 17–29. [Google Scholar] [CrossRef]
- Duncan, K.A.; Saldanha, C.J. Central aromatization: A dramatic and responsive defense against threat and trauma to the vertebrate brain. Front. Neuroendocr. 2020, 56, 100816. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Pratap, U.P.; Lu, Y.; Sareddy, G.R.; Tekmal, R.R.; Vadlamudi, R.K.; Brann, D.W. Development and Characterization of Inducible Astrocyte-Specific Aromatase Knockout Mice. Biology 2023, 12, 621. https://doi.org/10.3390/biology12040621
Wang J, Pratap UP, Lu Y, Sareddy GR, Tekmal RR, Vadlamudi RK, Brann DW. Development and Characterization of Inducible Astrocyte-Specific Aromatase Knockout Mice. Biology. 2023; 12(4):621. https://doi.org/10.3390/biology12040621
Chicago/Turabian StyleWang, Jing, Uday P. Pratap, Yujiao Lu, Gangadhara R. Sareddy, Rajeshwar R. Tekmal, Ratna K. Vadlamudi, and Darrell W. Brann. 2023. "Development and Characterization of Inducible Astrocyte-Specific Aromatase Knockout Mice" Biology 12, no. 4: 621. https://doi.org/10.3390/biology12040621
APA StyleWang, J., Pratap, U. P., Lu, Y., Sareddy, G. R., Tekmal, R. R., Vadlamudi, R. K., & Brann, D. W. (2023). Development and Characterization of Inducible Astrocyte-Specific Aromatase Knockout Mice. Biology, 12(4), 621. https://doi.org/10.3390/biology12040621