


Genome Study of α-, β-, and γ-Carbonic Anhydrases from the Thermophilic Microbiome of Marine Hydrothermal Vent Ecosystems

 ,
,  ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of α-, β-, and γ-CA Sequences
2.2. Phylogenetic Analysis
2.3. Identification of α-, β-, and γ-CA Genes on the MGEs
2.3.1. Integrons
2.3.2. Genomic Islands (GIs)
2.3.3. Integrative Conjugative Elements (ICEs)
2.3.4. Transposable Elements (TEs), Phages, and Plasmids
3. Results
3.1. Identification of α-, β-, and γ-CA and Protein Sequences
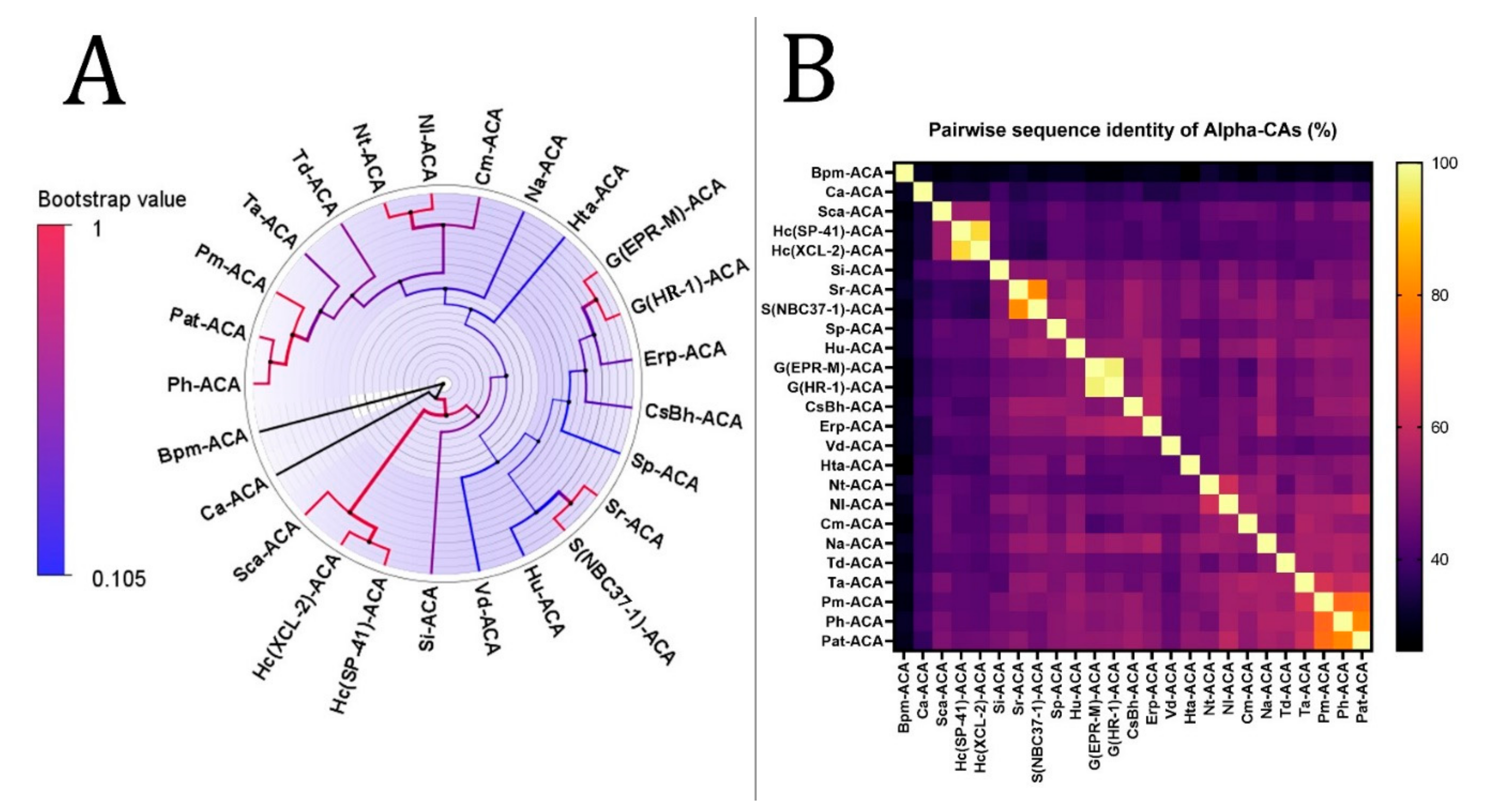
3.2. Phylogenetic Analysis
3.3. Identification of α-, β-, and γ-CA Genes on MGEs
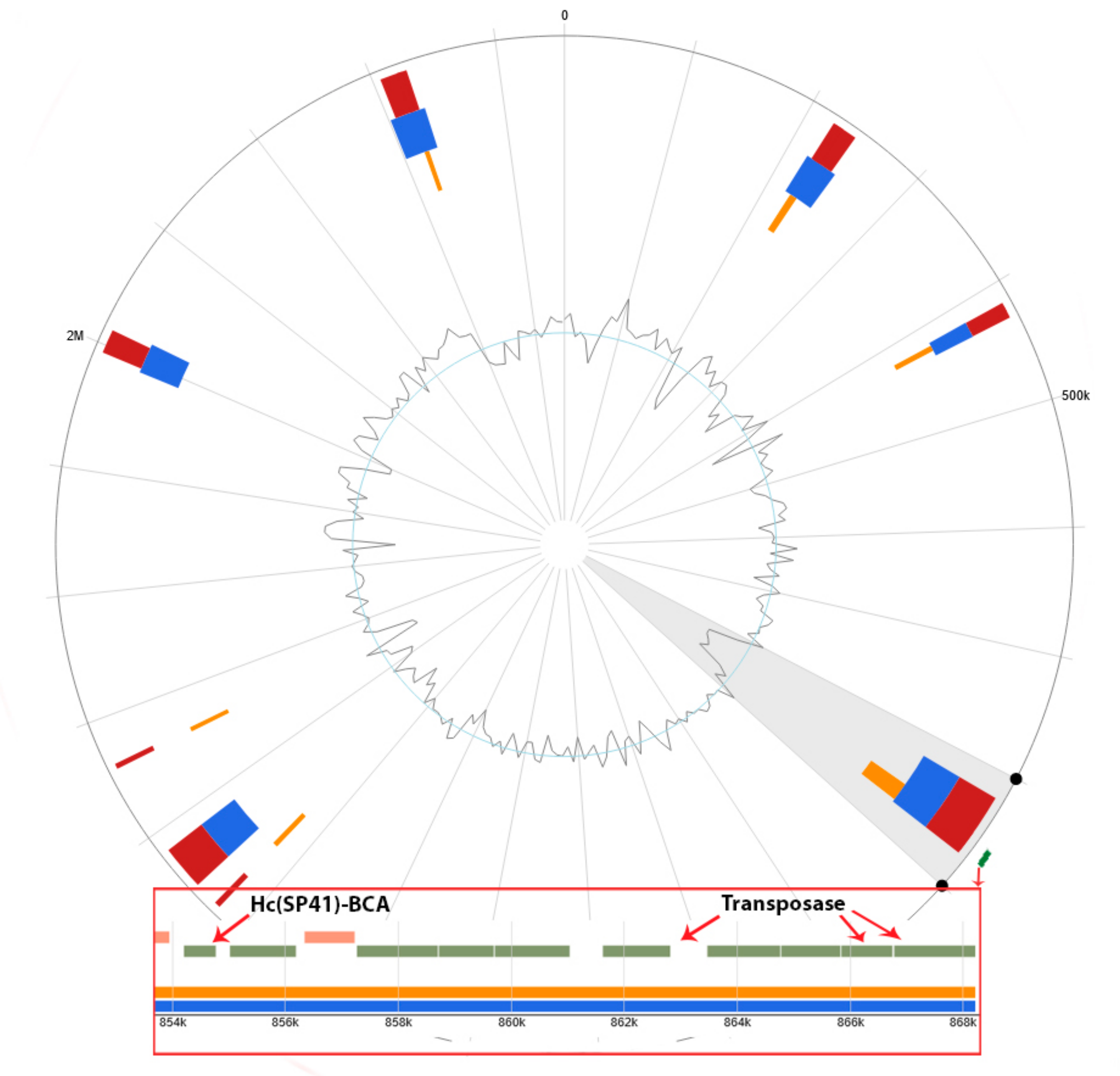
3.3.1. Integrons
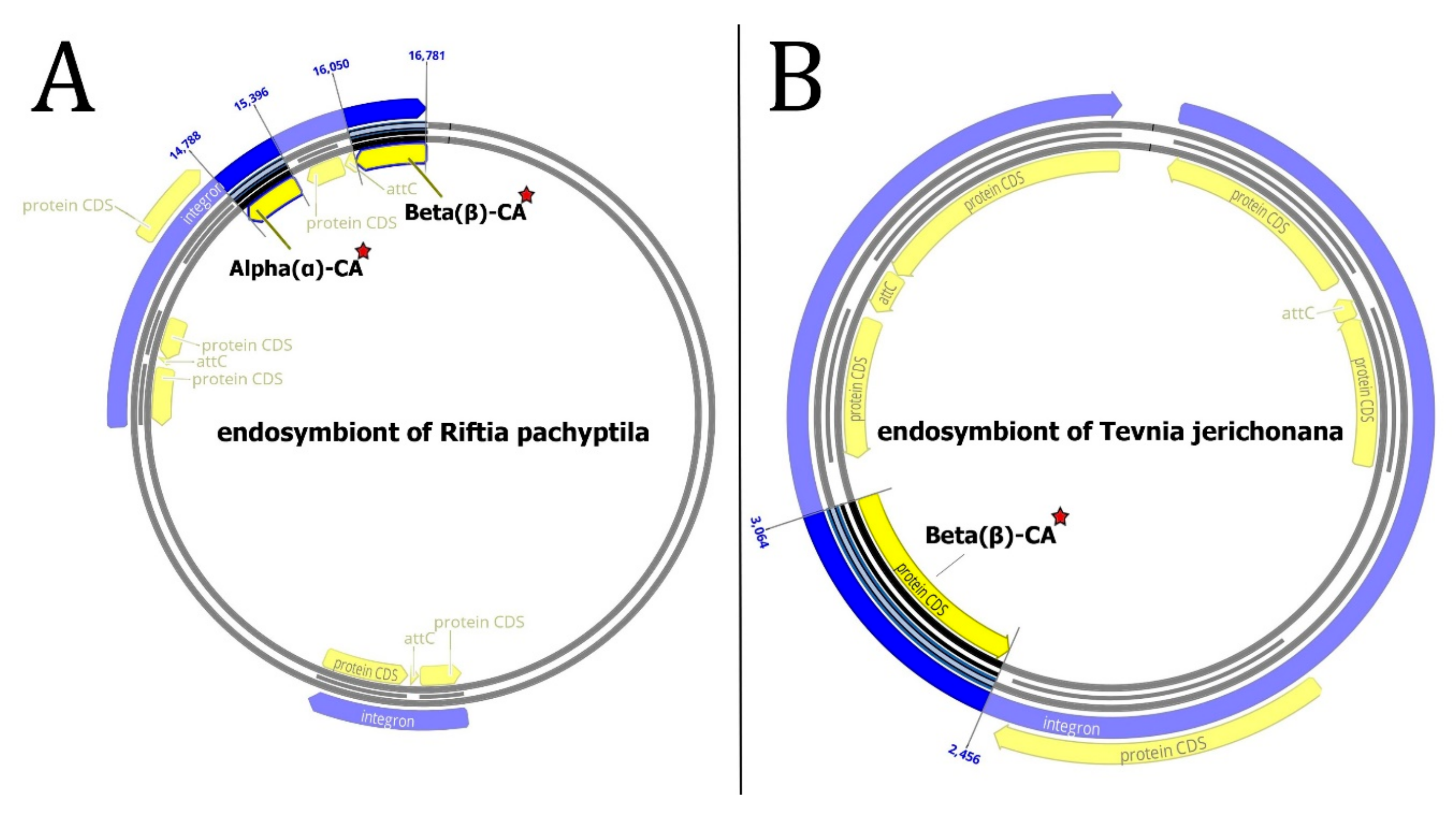
3.3.2. Genomic Islands (GIs)
3.3.3. Integrative Conjugative Elements (ICEs), Transposable Elements (TEs), Phages, and Plasmids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lonsdale, P. Clustering of suspension-feeding macrobenthos near abyssal hydrothermal vents at oceanic spreading centers. Deep Sea Res. 1977, 24, 857–863. [Google Scholar] [CrossRef]
- Corliss, J.B.; Dymond, J.; Gordon, L.I.; Edmond, J.M.; von Herzen, R.P.; Ballard, R.D.; Green, K.; Williams, D.; Bainbridge, A.; Crane, K. Submarine thermal springs on the Galapagos Rift. Science 1979, 203, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Colín-García, M.; Heredia, A.; Cordero, G.; Camprubí, A.; Negrón-Mendoza, A.; Ortega-Gutiérrez, F.; Beraldi, H.; Ramos-Bernal, S. Hydrothermal vents and prebiotic chemistry: A review. Bol. Soc. Geol. Mex. 2016, 68, 599–620. [Google Scholar] [CrossRef]
- Shitashima, K. CO2 supply from deep-sea hydrothermal systems. Waste Manag. 1998, 17, 385–390. [Google Scholar] [CrossRef]
- Maren, T.H. Carbonic anhydrase: Chemistry, physiology, and inhibition. Physiol. Rev. 1967, 47, 595–781. [Google Scholar] [CrossRef]
- Smith, K.S.; Jakubzick, C.; Whittam, T.S.; Ferry, J.G. Carbonic anhydrase is an ancient enzyme widespread in prokaryotes. Proc. Natl. Acad. Sci. USA 1999, 96, 15184–15189. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta-and gamma-carbonic anhydrases from Bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzym. Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef]
- Zolfaghari Emameh, R.; Kuuslahti, M.; Näreaho, A.; Sukura, A.; Parkkila, S. Innovative molecular diagnosis of T richinella species based on β-carbonic anhydrase genomic sequence. Microb. Biotechnol. 2016, 9, 172–179. [Google Scholar] [CrossRef]
- Zolfaghari Emameh, R.; Barker, H.R.; Syrjänen, L.; Urbański, L.; Supuran, C.T.; Parkkila, S. Identification and inhibition of carbonic anhydrases from nematodes. J. Enzym. Inhib. Med. Chem. 2016, 31, 176–184. [Google Scholar] [CrossRef]
- Emameh, R.Z.; Kuuslahti, M.; Vullo, D.; Barker, H.R.; Supuran, C.T.; Parkkila, S. Ascaris lumbricoides β carbonic anhydrase: A potential target enzyme for treatment of ascariasis. Parasites Vectors 2015, 8, 479. [Google Scholar] [CrossRef]
- Emameh, R.Z.; Barker, H.; Hytönen, V.P.; Tolvanen, M.E.; Parkkila, S. Beta carbonic anhydrases: Novel targets for pesticides and anti-parasitic agents in agriculture and livestock husbandry. Parasites Vectors 2014, 7, 403. [Google Scholar] [CrossRef] [PubMed]
- Zolfaghari Emameh, R.; Kuuslahti, M.; Nosrati, H.; Lohi, H.; Parkkila, S. Assessment of databases to determine the validity of beta- and gamma-carbonic anhydrase sequences from vertebrates. BMC Genom. 2020, 21, 352. [Google Scholar] [PubMed]
- Zolfaghari Emameh, R.; Hosseini, S.N.; Parkkila, S. Application of beta and gamma carbonic anhydrase sequences as tools for identification of bacterial contamination in the whole genome sequence of inbred Wuzhishan minipig (Sus scrofa) annotated in databases. Database 2021, 2021, baab029. [Google Scholar] [PubMed]
- Lindskog, S. Structure and mechanism of carbonic anhydrase. Pharmacol. Ther. 1997, 74, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Akocak, S.; Supuran, C.T. Activation of α-, β-, γ-δ-, ζ-and η-class of carbonic anhydrases with amines and amino acids: A review. J. Enzym. Inhib. Med. Chem. 2019, 34, 1652–1659. [Google Scholar] [CrossRef]
- Del Prete, S.; Nocentini, A.; Supuran, C.T.; Capasso, C. Bacterial ι-carbonic anhydrase: A new active class of carbonic anhydrase identified in the genome of the Gram-negative bacterium Burkholderia territorii. J. Enzym. Inhib. Med. Chem. 2020, 35, 1060–1068. [Google Scholar] [CrossRef]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. An overview of the bacterial carbonic anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef]
- Ferraroni, M.; Del Prete, S.; Vullo, D.; Capasso, C.; Supuran, C.T. Crystal structure and kinetic studies of a tetrameric type II β-carbonic anhydrase from the pathogenic bacterium Vibrio cholerae. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 2449–2456. [Google Scholar] [CrossRef]
- Iverson, T.M.; Alber, B.E.; Kisker, C.; Ferry, J.G.; Rees, D.C. A closer look at the active site of γ-class carbonic anhydrases: High-resolution crystallographic studies of the carbonic anhydrase from Methanosarcina thermophila. Biochemistry 2000, 39, 9222–9231. [Google Scholar] [CrossRef]
- Zolfaghari Emameh, R.; Barker, H.R.; Tolvanen, M.E.; Parkkila, S.; Hytönen, V.P. Horizontal transfer of β-carbonic anhydrase genes from prokaryotes to protozoans, insects, and nematodes. Parasites Vectors 2016, 9, 152. [Google Scholar] [CrossRef] [PubMed]
- Emameh, R.Z.; Barker, H.R.; Hytönen, V.P.; Parkkila, S. Involvement of β-carbonic anhydrase genes in bacterial genomic islands and their horizontal transfer to protists. Appl. Environ. Microbiol. 2018, 84, e00771-18. [Google Scholar]
- Andersson, J.O. Lateral gene transfer in eukaryotes. Cell. Mol. Life Sci. 2005, 62, 1182–1197. [Google Scholar] [CrossRef] [PubMed]
- Emamalipour, M.; Seidi, K.; Zununi Vahed, S.; Jahanban-Esfahlan, A.; Jaymand, M.; Majdi, H.; Amoozgar, Z.; Chitkushev, L.; Javaheri, T.; Jahanban-Esfahlan, R. Horizontal gene transfer: From evolutionary flexibility to disease progression. Front. Cell Dev. Biol. 2020, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Dutta, C.; Pan, A. Horizontal gene transfer and bacterial diversity. J. Biosci. 2002, 27, 27. [Google Scholar] [CrossRef] [PubMed]
- Frost, L.S.; Leplae, R.; Summers, A.O.; Toussaint, A. Mobile genetic elements: The agents of open source evolution. Nat. Rev. Microbiol. 2005, 3, 722–732. [Google Scholar] [CrossRef]
- Bellanger, X.; Payot, S.; Leblond-Bourget, N.; Guédon, G. Conjugative and mobilizable genomic islands in bacteria: Evolution and diversity. FEMS Microbiol. Rev. 2014, 38, 720–760. [Google Scholar] [CrossRef]
- Johnson, C.M.; Grossman, A.D. Integrative and conjugative elements (ICEs): What they do and how they work. Annu. Rev. Genet. 2015, 49, 577–601. [Google Scholar] [CrossRef]
- Watson, B.N.; Staals, R.H.; Fineran, P.C. CRISPR-Cas-mediated phage resistance enhances horizontal gene transfer by transduction. mBio 2018, 9, e02406-17. [Google Scholar] [CrossRef]
- Muñoz-López, M.; García-Pérez, J.L. DNA transposons: Nature and applications in genomics. Curr. Genom. 2010, 11, 115–128. [Google Scholar] [CrossRef]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Messier, N.; Roy, P.H. Integron integrases possess a unique additional domain necessary for activity. J. Bacteriol. 2001, 183, 6699–6706. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Recchia, G.D.; Scaramuzzi, C.; Collis, C.M.; Stokes, H.; Hall, R.M. Definition of the attI1 site of class 1 integrons. Microbiology 2000, 146, 2855–2864. [Google Scholar] [CrossRef] [PubMed]
- Collis, C.M.; Hall, R.M. Expression of antibiotic resistance genes in the integrated cassettes of integrons. Antimicrob. Agents Chemother. 1995, 39, 155–162. [Google Scholar] [CrossRef]
- Alvarez-Martinez, C.E.; Christie, P.J. Biological diversity of prokaryotic type IV secretion systems. Microbiol. Mol. Biol. Rev. 2009, 73, 775–808. [Google Scholar] [CrossRef]
- Domsic, J.F.; McKenna, R. Sequestration of carbon dioxide by the hydrophobic pocket of the carbonic anhydrases. Biochim. Biophys. Acta-Proteins Proteom. 2010, 1804, 326–331. [Google Scholar] [CrossRef]
- Mahillon, J.; Chandler, M. Insertion sequences. Microbiol. Mol. Biol. Rev. 1998, 62, 725–774. [Google Scholar] [CrossRef]
- Fisher, R.M.; Henry, L.M.; Cornwallis, C.K.; Kiers, E.T.; West, S.A. The evolution of host-symbiont dependence. Nat. Commun. 2017, 8, 15973. [Google Scholar] [CrossRef]
- Hall, R.J.; Whelan, F.J.; McInerney, J.O.; Ou, Y.; Domingo-Sananes, M.R. Horizontal Gene Transfer as a Source of Conflict and Cooperation in Prokaryotes. Front Microbiol 2020, 11, 1569. [Google Scholar]
- Sabine, C.L.; Tanhua, T. Estimation of anthropogenic CO2 inventories in the ocean. Ann. Rev. Mar. Sci. 2010, 2, 175–198. [Google Scholar] [CrossRef]
- De Goeyse, S.; Webb, A.E.; Reichart, G.-J.; De Nooijer, L.J. Carbonic anhydrase is involved in calcification by the benthic foraminifer Amphistegina lessonii. Biogeosciences 2021, 18, 393–401. [Google Scholar] [CrossRef]
- Manyumwa, C.V.; Emameh, R.Z.; Tastan Bishop, Ö. Alpha-carbonic anhydrases from hydrothermal vent sources as potential carbon dioxide sequestration agents: In silico sequence, structure and dynamics analyses. Int. J. Mol. Sci. 2020, 21, 8066. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.C.; Ferreira, V.; Zhang, X.; Anjos, L.; Félix, R.C.; Batista, F.M.; Power, D.M. Evolution and diversity of alpha-carbonic anhydrases in the mantle of the Mediterranean mussel (Mytilus galloprovincialis). Sci. Rep. 2019, 9, 10400. [Google Scholar] [CrossRef] [PubMed]
- Parisi, G.; Perales, M.; Fornasari, M.; Colaneri, A.; Schain, N.; Casati, D.; Zimmermann, S.; Brennicke, A.; Araya, A.; Ferry, J. Gamma carbonic anhydrases in plant mitochondria. Plant Mol. Biol. 2004, 55, 193–207. [Google Scholar] [CrossRef]
- Smith, K.S.; Ferry, J.G. Prokaryotic carbonic anhydrases. FEMS Microbiol. Rev. 2000, 24, 335–366. [Google Scholar] [CrossRef]
- Dobrinski, K.P.; Longo, D.L.; Scott, K.M. The carbon-concentrating mechanism of the hydrothermal vent chemolithoautotroph Thiomicrospira crunogena. J. Bacteriol. 2005, 187, 5761–5766. [Google Scholar] [CrossRef]
- Miroshnichenko, M.L.; Kostrikina, N.A.; Chernyh, N.A.; Pimenov, N.V.; Tourova, T.P.; Antipov, A.N.; Spring, S.; Stackebrandt, E.; Bonch-Osmolovskaya, E.A. Caldithrix abyssi gen. nov., sp. nov., a nitrate-reducing, thermophilic, anaerobic bacterium isolated from a Mid-Atlantic Ridge hydrothermal vent, represents a novel bacterial lineage. Int. J. Syst. Evol. Microbiol. 2003, 53, 323–329. [Google Scholar] [CrossRef]
- Voordeckers, J.W.; Starovoytov, V.; Vetriani, C. Caminibacter mediatlanticus sp. nov., a thermophilic, chemolithoautotrophic, nitrate-ammonifying bacterium isolated from a deep-sea hydrothermal vent on the Mid-Atlantic Ridge. Int. J. Syst. Evol. Microbiol. 2005, 55, 773–779. [Google Scholar] [CrossRef]
- Ward, M.E.; Shields, J.D.; Van Dover, C.L. Parasitism in species of Bathymodiolus (Bivalvia: Mytilidae) mussels from deep-sea seep and hydrothermal vents. Dis. Aquat. 2004, 62, 1–16. [Google Scholar] [CrossRef]
- Klose, J.; Polz, M.F.; Wagner, M.; Schimak, M.P.; Gollner, S.; Bright, M. Endosymbionts escape dead hydrothermal vent tubeworms to enrich the free-living population. Proc. Natl. Acad. Sci. USA 2015, 112, 11300–11305. [Google Scholar] [CrossRef]
- L’Haridon, S.; Reysenbach, A.-L.; Tindall, B.; Schönheit, P.; Banta, A.; Johnsen, U.; Schumann, P.; Gambacorta, A.; Stackebrandt, E.; Jeanthon, C. Desulfurobacterium atlanticum sp. nov., Desulfurobacterium pacificum sp. nov. and Thermovibrio guaymasensis sp. nov., three thermophilic members of the Desulfurobacteriaceae fam. nov., a deep branching lineage within the Bacteria. Int. J. Syst. Evol. Microbiol. 2006, 56, 2843–2852. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Perner, M. Reasons for Thiomicrospira crunogena’s recalcitrance towards previous attempts to detect its hydrogen consumption ability. Environ. Microbiol. Rep. 2016, 8, 53–57. [Google Scholar] [CrossRef]
- Takai, K.; Nealson, K.H.; Horikoshi, K. Hydrogenimonas thermophila gen. nov., sp. nov., a novel thermophilic, hydrogen-oxidizing chemolithoautotroph within the ε-Proteobacteria, isolated from a black smoker in a Central Indian Ridge hydrothermal field. Int. J. Syst. Evol. Microbiol. 2004, 54, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Mino, S.; Shiotani, T.; Nakagawa, S.; Takai, K.; Sawabe, T. Hydrogenimonas urashimensis sp. nov., a hydrogen-oxidizing chemolithoautotroph isolated from a deep-sea hydrothermal vent in the Southern Mariana Trough. Syst. Appl. Microbiol. 2021, 44, 126170. [Google Scholar] [CrossRef] [PubMed]
- Shiotani, T.; Mino, S.; Sato, W.; Nishikawa, S.; Yonezawa, M.; Sievert, S.M.; Sawabe, T. Nitrosophilus alvini gen. nov., sp. nov., a hydrogen-oxidizing chemolithoautotroph isolated from a deep-sea hydrothermal vent in the East Pacific Rise, inferred by a genome-based taxonomy of the phylum “Campylobacterota”. PLoS ONE 2020, 15, e0241366. [Google Scholar] [CrossRef]
- Nakagawa, S.; Takai, K.; Inagaki, F.; Horikoshi, K.; Sako, Y. Nitratiruptor tergarcus gen. nov., sp. nov. and Nitratifractor salsuginis gen. nov., sp. nov., nitrate-reducing chemolithoautotrophs of the ε-Proteobacteria isolated from a deep-sea hydrothermal system in the Mid-Okinawa Trough. Int. J. Syst. Evol. Microbiol. 2005, 55, 925–933. [Google Scholar] [CrossRef]
- François, D.X.; Godfroy, A.; Mathien, C.; Aubé, J.; Cathalot, C.; Lesongeur, F.; L’Haridon, S.; Philippon, X.; Roussel, E.G. Persephonella atlantica sp. nov.: How to adapt to physico-chemical gradients in high temperature hydrothermal habitats. Syst. Appl. Microbiol. 2021, 44, 126176. [Google Scholar] [CrossRef]
- Nakagawa, S.; Takai, K.; Horikoshi, K.; Sako, Y. Persephonella hydrogeniphila sp. nov., a novel thermophilic, hydrogen-oxidizing bacterium from a deep-sea hydrothermal vent chimney. Int. J. Syst. Evol. Microbiol. 2003, 53, 863–869. [Google Scholar] [CrossRef]
- Götz, D.; Banta, A.; Beveridge, T.; Rushdi, A.; Simoneit, B.; Reysenbach, A. Persephonella marina gen. nov., sp. nov. and Persephonella guaymasensis sp. nov., two novel, thermophilic, hydrogen-oxidizing microaerophiles from deep-sea hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2002, 52, 1349–1359. [Google Scholar]
- Waite, D.W.; Vanwonterghem, I.; Rinke, C.; Parks, D.H.; Zhang, Y.; Takai, K.; Sievert, S.M.; Simon, J.; Campbell, B.J.; Hanson, T.E. Comparative genomic analysis of the class Epsilonproteobacteria and proposed reclassification to Epsilonbacteraeota (phyl. nov.). Front. Microbiol. 2017, 8, 682. [Google Scholar] [CrossRef]
- Xie, S.; Wang, S.; Li, D.; Shao, Z.; Lai, Q.; Wang, Y.; Wei, M.; Han, X.; Jiang, L. Sulfurovum indicum sp. nov., a novel hydrogen-and sulfur-oxidizing chemolithoautotroph isolated from a deep-sea hydrothermal plume in the Northwestern Indian Ocean. Int. J. Syst. Evol. Microbiol. 2021, 71, 004748. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Suzuki, M.; Nakagawa, S.; Miyazaki, M.; Suzuki, Y.; Inagaki, F.; Horikoshi, K. Sulfurimonas paralvinellae sp. nov., a novel mesophilic, hydrogen-and sulfur-oxidizing chemolithoautotroph within the Epsilonproteobacteria isolated from a deep-sea hydrothermal vent polychaete nest, reclassification of Thiomicrospira denitrificans as Sulfurimonas denitrificans comb. nov. and emended description of the genus Sulfurimonas. Int. J. Syst. Evol. Microbiol. 2006, 56, 1725–1733. [Google Scholar] [PubMed]
- Boutet, I.; Ripp, R.; Lecompte, O.; Dossat, C.; Corre, E.; Tanguy, A.; Lallier, F.H. Conjugating effects of symbionts and environmental factors on gene expression in deep-sea hydrothermal vent mussels. BMC Genom. 2011, 12, 530. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Gugliandolo, C.; Spanò, A.; Taurisano, V.; Di Donato, P.; Maugeri, T.L.; Nicolaus, B.; Arena, A. Poly-[Gamma]-Glutamic Acid from Bacillus Horneckiae Strain APA of Shallow Marine Vent Origin with Antiviral and Immunomodulatory Effects against Herpes Simplex Virus Type-2. J. Mar. Sci. Res. Dev. 2015, 5, 173. [Google Scholar] [CrossRef]
- Wu, Y.-H.; Xu, L.; Zhou, P.; Wang, C.-S.; Oren, A.; Xu, X.-W. Brevirhabdus pacifica gen. nov., sp. nov., isolated from deep-sea sediment in a hydrothermal vent field. Int. J. Syst. Evol. Microbiol. 2015, 65, 3645–3651. [Google Scholar] [CrossRef]
- Ponnudurai, R.; Sayavedra, L.; Kleiner, M.; Heiden, S.E.; Thürmer, A.; Felbeck, H.; Schlüter, R.; Sievert, S.M.; Daniel, R.; Schweder, T. Genome sequence of the sulfur-oxidizing Bathymodiolus thermophilus gill endosymbiont. Stand. Genom. Sci. 2017, 12, 50. [Google Scholar] [CrossRef]
- Grosche, A.; Sekaran, H.; Pérez-Rodríguez, I.; Starovoytov, V.; Vetriani, C. Cetia pacifica gen. nov., sp. nov., a chemolithoautotrophic, thermophilic, nitrate-ammonifying bacterium from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2015, 65, 1144–1150. [Google Scholar] [CrossRef]
- Slobodkina, G.; Kolganova, T.; Chernyh, N.; Querellou, J.; Bonch-Osmolovskaya, E.; Slobodkin, A. Deferribacter autotrophicus sp. nov., an iron (III)-reducing bacterium from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2009, 59, 1508–1512. [Google Scholar] [CrossRef]
- Cao, J.; Birien, T.; Gayet, N.; Huang, Z.; Shao, Z.; Jebbar, M.; Alain, K. Desulfurobacterium indicum sp. nov., a thermophilic sulfur-reducing bacterium from the Indian Ocean. Int. J. Syst. Evol. Microbiol. 2017, 67, 1665–1668. [Google Scholar] [CrossRef]
- Jiang, L.; Xu, H.; Shao, Z.; Long, M. Defluviimonas indica sp. nov., a marine bacterium isolated from a deep-sea hydrothermal vent environment. Int. J. Syst. Evol. Microbiol. 2014, 64, 2084–2088. [Google Scholar] [CrossRef]
- Hansen, M.; Perner, M. Hydrogenase gene distribution and H2 consumption ability within the Thiomicrospira lineage. Front. Microbiol. 2016, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Kaye, J.Z.; Marquez, M.C.; Ventosa, A.; Baross, J.A. Halomonas neptunia sp. nov., Halomonas sulfidaeris sp. nov., Halomonas axialensis sp. nov. and Halomonas hydrothermalis sp. nov.: Halophilic bacteria isolated from deep-sea hydrothermal-vent environments. Int. J. Syst. Evol. Microbiol. 2004, 54, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Lyu, J.; Shao, Z. Sulfur metabolism of Hydrogenovibrio thermophilus strain S5 and its adaptations to deep-sea hydrothermal vent environment. Front. Microbiol. 2017, 8, 2513. [Google Scholar] [CrossRef] [PubMed]
- Slobodkina, G.B.; Baslerov, R.V.; Novikov, A.A.; Viryasov, M.B.; Bonch-Osmolovskaya, E.A.; Slobodkin, A.I. Inmirania thermothiophila gen. nov., sp. nov., a thermophilic, facultatively autotrophic, sulfur-oxidizing gammaproteobacterium isolated from a shallow-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2016, 66, 701–706. [Google Scholar] [CrossRef]
- Nagata, R.; Takaki, Y.; Tame, A.; Nunoura, T.; Muto, H.; Mino, S.; Sawayama, S.; Takai, K.; Nakagawa, S. Lebetimonas natsushimae sp. nov., a novel strictly anaerobic, moderately thermophilic chemoautotroph isolated from a deep-sea hydrothermal vent polychaete nest in the Mid-Okinawa Trough. Syst. Appl. Microbiol. 2017, 40, 352–356. [Google Scholar] [CrossRef]
- Stewart, L.C.; Jung, J.-H.; Kim, Y.-T.; Kwon, S.-W.; Park, C.-S.; Holden, J.F. Methanocaldococcus bathoardescens sp. nov., a hyperthermophilic methanogen isolated from a volcanically active deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2015, 65, 1280–1283. [Google Scholar] [CrossRef]
- McAllister, S.M.; Davis, R.E.; McBeth, J.M.; Tebo, B.M.; Emerson, D.; Moyer, C.L. Biodiversity and emerging biogeography of the neutrophilic iron-oxidizing Zetaproteobacteria. Appl. Environ. Microbiol. 2011, 77, 5445–5457. [Google Scholar] [CrossRef]
- Takai, K.; Nealson, K.H.; Horikoshi, K. Methanotorris formicicus sp. nov., a novel extremely thermophilic, methane-producing archaeon isolated from a black smoker chimney in the Central Indian Ridge. Int. J. Syst. Evol. Microbiol. 2004, 54, 1095–1100. [Google Scholar] [CrossRef]
- Takeuchi, M.; Katayama, T.; Yamagishi, T.; Hanada, S.; Tamaki, H.; Kamagata, Y.; Oshima, K.; Hattori, M.; Marumo, K.; Nedachi, M. Methyloceanibacter caenitepidi gen. nov., sp. nov., a facultatively methylotrophic bacterium isolated from marine sediments near a hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2014, 64, 462–468. [Google Scholar] [CrossRef]
- Wang, H.; Li, H.; Shao, Z.; Liao, S.; Johnstone, L.; Rensing, C.; Wang, G. Genome sequence of deep-sea manganese-oxidizing bacterium Marinobacter manganoxydans MnI7-9. J. Bacteriol. 2012, 194, 899–900. [Google Scholar] [CrossRef]
- Takai, K.; Inoue, A.; Horikoshi, K. Methanothermococcus okinawensis sp. nov., a thermophilic, methane-producing archaeon isolated from a Western Pacific deep-sea hydrothermal vent system. Int. J. Syst. Evol. Microbiol. 2002, 52, 1089–1095. [Google Scholar] [PubMed]
- Zhou, M.; Dong, B.; Shao, Z. Complete genome sequence of Marinobacter sp. LQ44, a haloalkaliphilic phenol-degrading bacterium isolated from a deep-sea hydrothermal vent. Mar. Genom. 2020, 50, 100697. [Google Scholar] [CrossRef]
- Handley, K.M.; Hery, M.; Lloyd, J.R. Marinobacter santoriniensis sp. nov., an arsenate-respiring and arsenite-oxidizing bacterium isolated from hydrothermal sediment. Int. J. Syst. Evol. Microbiol. 2009, 59, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Campbell, B.J.; Hanson, T.E.; Zhang, C.L.; Cary, S.C. Nautilia profundicola sp. nov., a thermophilic, sulfur-reducing epsilonproteobacterium from deep-sea hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2008, 58, 1598–1602. [Google Scholar] [CrossRef]
- Dong, C.; Lai, Q.; Chen, L.; Sun, F.; Shao, Z.; Yu, Z. Oceanibaculum pacificum sp. nov., isolated from hydrothermal field sediment of the south-west Pacific Ocean. Int. J. Syst. Evol. Microbiol. 2010, 60, 219–222. [Google Scholar] [CrossRef]
- Miroshnichenko, M.; L’haridon, S.; Jeanthon, C.; Antipov, A.; Kostrikina, N.; Tindall, B.; Schumann, P.; Spring, S.; Stackebrandt, E.; Bonch-Osmolovskaya, E. Oceanithermus profundus gen. nov., sp. nov., a thermophilic, microaerophilic, facultatively chemolithoheterotrophic bacterium from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2003, 53, 747–752. [Google Scholar] [CrossRef]
- Newton, I.; Woyke, T.; Auchtung, T.; Dilly, G.; Dutton, R.; Fisher, M.; Fontanez, K.; Lau, E.; Stewart, F.; Richardson, P. The Calyptogena magnifica chemoautotrophic symbiont genome. Science 2007, 315, 998–1000. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, H.; Zhou, W.; Wang, Y.; Zhou, H.; Chen, X. Sulfur metabolism pathways in Sulfobacillus acidophilus TPY, a gram-positive moderate thermoacidophile from a hydrothermal vent. Front. Microbiol. 2016, 7, 1861. [Google Scholar] [CrossRef]
- Inagaki, F.; Takai, K.; Nealson, K.H.; Horikoshi, K. Sulfurovum lithotrophicum gen. nov., sp. nov., a novel sulfur-oxidizing chemolithoautotroph within the ε-Proteobacteria isolated from Okinawa Trough hydrothermal sediments. Int. J. Syst. Evol. Microbiol. 2004, 54, 1477–1482. [Google Scholar] [CrossRef]
- Giovannelli, D.; Chung, M.; Staley, J.; Starovoytov, V.; Le Bris, N.; Vetriani, C. Sulfurovum riftiae sp. nov., a mesophilic, thiosulfate-oxidizing, nitrate-reducing chemolithoautotrophic epsilonproteobacterium isolated from the tube of the deep-sea hydrothermal vent polychaete Riftia pachyptila. Int. J. Syst. Evol. Microbiol. 2016, 66, 2697–2701. [Google Scholar] [CrossRef]
- Vetriani, C.; Speck, M.D.; Ellor, S.V.; Lutz, R.A.; Starovoytov, V. Thermovibrio ammonificans sp. nov., a thermophilic, chemolithotrophic, nitrate-ammonifying bacterium from deep-sea hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2004, 54, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Urios, L.; Cueff, V.; Pignet, P.; Barbier, G. Tepidibacter formicigenes sp. nov., a novel spore-forming bacterium isolated from a Mid-Atlantic Ridge hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2004, 54, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Hensley, S.A.; Jung, J.-H.; Park, C.-S.; Holden, J.F. Thermococcus paralvinellae sp. nov. and Thermococcus cleftensis sp. nov. of hyperthermophilic heterotrophs from deep-sea hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2014, 64, 3655–3659. [Google Scholar] [CrossRef]
- Martins, E.; Santos, R.S.; Bettencourt, R. Vibrio diabolicus challenge in Bathymodiolus azoricus populations from Menez Gwen and Lucky Strike hydrothermal vent sites. Fish. Shellfish Immunol. 2015, 47, 962–977. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, H.; Yoshida, T.; Takaki, Y.; Shimamura, S.; Nishi, S.; Harada, M.; Matsuyama, K.; Takishita, K.; Kawato, M.; Uematsu, K. Reduced genome of the thioautotrophic intracellular symbiont in a deep-sea clam, Calyptogena okutanii. Curr. Biol. 2007, 17, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhang, Z.; Li, X.; Zhang, X.; Cao, J.; Jebbar, M.; Alain, K.; Shao, Z. Anoxybacter fermentans gen. nov., sp. nov., a piezophilic, thermophilic, anaerobic, fermentative bacterium isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2015, 65, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Zammuto, V.; Fuchs, F.M.; Fiebrandt, M.; Stapelmann, K.; Ulrich, N.J.; Maugeri, T.L.; Pukall, R.; Gugliandolo, C.; Moeller, R. Comparing spore resistance of Bacillus strains isolated from hydrothermal vents and spacecraft assembly facilities to environmental stressors and decontamination treatments. Astrobiology 2018, 18, 1425–1434. [Google Scholar] [CrossRef]
- Wery, N.; Moricet, J.-M.; Cueff, V.; Jean, J.; Pignet, P.; Lesongeur, F.; Cambon-Bonavita, M.-A.; Barbier, G. Caloranaerobacter azorensis gen. nov., sp nov., an anaerobic thermophilic bacterium isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2001, 51, 1789–1796. [Google Scholar] [CrossRef]
- Alain, K.; Pignet, P.; Zbinden, M.; Quillevere, M.; Duchiron, F.; Donval, J.-P.; Lesongeur, F.; Raguenes, G.; Crassous, P.; Querellou, J. Caminicella sporogenes gen. nov., sp. nov., a novel thermophilic spore-forming bacterium isolated from an East-Pacific Rise hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2002, 52, 1621–1628. [Google Scholar]
- Takai, K.; Kobayashi, H.; Nealson, K.H.; Horikoshi, K. Deferribacter desulfuricans sp. nov., a novel sulfur-, nitrate-and arsenate-reducing thermophile isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2003, 53, 839–846. [Google Scholar] [CrossRef]
- Chao, L.S.L.; Davis, R.E.; Moyer, C.L. Characterization of bacterial community structure in vestimentiferan tubeworm Ridgeia piscesae trophosomes. Mar. Ecol. 2007, 28, 72–85. [Google Scholar] [CrossRef]
- Won, Y.-J.; Hallam, S.J.; O’Mullan, G.D.; Pan, I.L.; Buck, K.R.; Vrijenhoek, R.C. Environmental acquisition of thiotrophic endosymbionts by deep-sea mussels of the genus Bathymodiolus. Appl. Environ. Microbiol. 2003, 69, 6785–6792. [Google Scholar] [CrossRef] [PubMed]
- Slobodkina, G.; Kolganova, T.; Querellou, J.; Bonch-Osmolovskaya, E.; Slobodkin, A. Geoglobus acetivorans sp. nov., an iron (III)-reducing archaeon from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 2009, 59, 2880–2883. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Stackebrandt, E. Three novel halotolerant and thermophilic Geobacillus strains from shallow marine vents. Syst. Appl. Microbiol. 2002, 25, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Roalkvam, I.; Bredy, F.; Baumberger, T.; Pedersen, R.-B.; Steen, I.H. Hypnocyclicus thermotrophus gen. nov., sp. nov. isolated from a microbial mat in a hydrothermal vent field. Int. J. Syst. Evol. Microbiol. 2015, 65, 4521–4525. [Google Scholar] [CrossRef]
- Van Dover, C. The Ecology of Deep-Sea Hydrothermal Vents; Princeton University Press: Princeton, NJ, USA, 2000. [Google Scholar]
- Erauso, G.; Reysenbach, A.-L.; Godfroy, A.; Meunier, J.-R.; Crump, B.; Partensky, F.; Baross, J.A.; Marteinsson, V.; Barbier, G.; Pace, N.R. Pyrococcus abyssi sp. nov., a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. Arch. Microbiol. 1993, 160, 338–349. [Google Scholar] [CrossRef]
- Marteinsson, V.T.; Bjornsdottir, S.H.; Bienvenu, N.; Kristjansson, J.K.; Birrien, J.-L. Rhodothermus profundi sp. nov., a thermophilic bacterium isolated from a deep-sea hydrothermal vent in the Pacific Ocean. Int. J. Syst. Evol. Microbiol. 2010, 60, 2729–2734. [Google Scholar] [CrossRef]
- Miroshnichenko, M.; Gongadze, G.; Rainey, F.; Kostyukova, A.; Lysenko, A.; Chernyh, N.; Bonch-Osmolovskaya, E. Thermococcus gorgonarius sp. nov. and Thermococcus pacificus sp. nov.: Heterotrophic extremely thermophilic archaea from New Zealand submarine hot vents. Int. J. Syst. Evol. Microbiol. 1998, 48, 23–29. [Google Scholar] [CrossRef]
- Marteinsson, V.T.; Birrien, J.-L.; Reysenbach, A.-L.; Vernet, M.; Marie, D.; Gambacorta, A.; Messner, P.; Sleytr, U.B.; Prieur, D. Thermococcus barophilus sp. nov., a new barophilic and hyperthermophilic archaeon isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 1999, 49, 351–359. [Google Scholar] [CrossRef]
- Duffaud, G.D.; d’Hennezel, O.B.; Peek, A.S.; Reysenbach, A.-L.; Kelly, R.M. Isolation and characterization of Thermococcus barossii, sp. nov., a hyperthermophilic archaeon isolated from a hydrothermal vent flange formation. Syst. Appl. Microbiol. 1998, 21, 40–49. [Google Scholar] [CrossRef]
- Jolivet, E.; l’Haridon, S.; Corre, E.; Forterre, P.; Prieur, D. Thermococcus gammatolerans sp. nov., a hyperthermophilic archaeon from a deep-sea hydrothermal vent that resists ionizing radiation. Int. J. Syst. Evol. Microbiol. 2003, 53, 847–851. [Google Scholar] [CrossRef]
- Gorlas, A.; Croce, O.; Oberto, J.; Gauliard, E.; Forterre, P.; Marguet, E. Thermococcusnautili sp. nov., a hyperthermophilic archaeon isolated from a hydrothermal deep-sea vent. Int. J. Syst. Evol. Microbiol. 2014, 64, 1802–1810. [Google Scholar] [CrossRef] [PubMed]
- González, J.M.; Kato, C.; Horikoshi, K. Thermococcus peptonophilus sp. nov., a fast-growing, extremely thermophilic archaebacterium isolated from deep-sea hydrothermal vents. Arch. Microbiol. 1995, 164, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Kwak, Y.S.; Akiba, T.; Kudo, T.; Horikoshi, K. Thermococcus profundus sp. nov., a new hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. Syst. Appl. Microbiol. 1994, 17, 232–236. [Google Scholar] [CrossRef]
- Dalmasso, C.; Oger, P.; Selva, G.; Courtine, D.; L’haridon, S.; Garlaschelli, A.; Roussel, E.; Miyazaki, J.; Reveillaud, J.; Jebbar, M. Thermococcus piezophilus sp. nov., a novel hyperthermophilic and piezophilic archaeon with a broad pressure range for growth, isolated from a deepest hydrothermal vent at the Mid-Cayman Rise. Syst. Appl. Microbiol. 2016, 39, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Slobodkin, A.; Tourova, T.; Kostrikina, N.; Chernyh, N.; Bonch-Osmolovskaya, E.; Jeanthon, C.; Jones, B. Tepidibacterthalassicus gen. nov., sp. nov., a novel moderately thermophilic, anaerobic, fermentative bacterium from a deep-sea hydrothermal vent. J. Med. Microbiol. 2003, 53, 1131–1134. [Google Scholar]
- L’Haridon, S.; Miroshnichenko, M.; Kostrikina, N.; Tindall, B.; Spring, S.; Schumann, P.; Stackebrandt, E.; Bonch-Osmolovskaya, E.; Jeanthon, C. Vulcanibacillus modesticaldus gen. nov., sp. nov., a strictly anaerobic, nitrate-reducing bacterium from deep-sea hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2006, 56, 1047–1053. [Google Scholar] [CrossRef]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hall, R.M.; Brookes, D.E.; Stokes, H. Site-specific insertion of genes into integrons: Role of the 59-base element and determination of the recombination cross-over point. Mol. Microbiol. 1991, 5, 1941–1959. [Google Scholar] [CrossRef] [PubMed]
- Rowe-Magnus, D.A.; Guérout, A.-M.; Mazel, D. Super-integrons. Res. Microbiol. 1999, 150, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Rowe-Magnus, D.A.; Guerout, A.-M.; Ploncard, P.; Dychinco, B.; Davies, J.; Mazel, D. The evolutionary history of chromosomal super-integrons provides an ancestry for multiresistant integrons. Proc. Natl. Acad. Sci. USA 2001, 98, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Stokes, H.; O’Gorman, D.; Recchia, G.D.; Parsekhian, M.; Hall, R.M. Structure and function of 59-base element recombination sites associated with mobile gene cassettes. Mol. Microbiol. 1997, 26, 731–745. [Google Scholar] [CrossRef]
- Collis, C.M.; Hall, R.M. Gene cassettes from the insert region of integrons are excised as covalently closed circles. Mol. Microbiol. 1992, 6, 2875–2885. [Google Scholar] [CrossRef]
- Hocquet, D.; Llanes, C.; Thouverez, M.; Kulasekara, H.D.; Bertrand, X.; Plésiat, P.; Mazel, D.; Miller, S.I. Evidence for induction of integron-based antibiotic resistance by the SOS response in a clinical setting. PLoS Pathog. 2012, 8, e1002778. [Google Scholar] [CrossRef]
- Cury, J.; Jové, T.; Touchon, M.; Néron, B.; Rocha, E.P. Identification and analysis of integrons and cassette arrays in bacterial genomes. Nucleic Acids Res. 2016, 44, 4539–4550. [Google Scholar] [CrossRef]
- Hsiao, W.; Wan, I.; Jones, S.J.; Brinkman, F.S. IslandPath: Aiding detection of genomic islands in prokaryotes. Bioinformatics 2003, 19, 418–420. [Google Scholar] [CrossRef]
- Tu, Q.; Ding, D. Detecting pathogenicity islands and anomalous gene clusters by iterative discriminant analysis. FEMS Microbiol. Lett. 2003, 221, 269–275. [Google Scholar] [CrossRef]
- Rajan, I.; Aravamuthan, S.; Mande, S.S. Identification of compositionally distinct regions in genomes using the centroid method. Bioinformatics 2007, 23, 2672–2677. [Google Scholar] [CrossRef]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Group, S.F.U.R.C.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.F.; Almagro-Moreno, S.; Parent, M.A. Genomic islands are dynamic, ancient integrative elements in bacterial evolution. Trends Microbiol. 2009, 17, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, X.; Xie, Y.; Bi, D.; Sun, J.; Li, J.; Tai, C.; Deng, Z.; Ou, H.-Y. ICEberg 2.0: An updated database of bacterial integrative and conjugative elements. Nucleic Acids Res 2019, 47, D660–D665. [Google Scholar] [CrossRef] [PubMed]
- Bi, D.; Xu, Z.; Harrison, E.M.; Tai, C.; Wei, Y.; He, X.; Jia, S.; Deng, Z.; Rajakumar, K.; Ou, H.-Y. ICEberg: A web-based resource for integrative and conjugative elements found in Bacteria. Nucleic Acids Res. 2012, 40, D621–D626. [Google Scholar] [CrossRef]
- Johansson, M.H.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Campbell, A. Prophage insertion sites. Res. Microbiol. 2003, 154, 277–282. [Google Scholar] [CrossRef]
- Sousa, A.L.D.; Maués, D.; Lobato, A.; Franco, E.F.; Pinheiro, K.; Araújo, F.; Pantoja, Y.; Costa da Silva, A.L.D.; Morais, J.; Ramos, R.T. PhageWeb–Web interface for rapid identification and characterization of prophages in bacterial genomes. Front. Genet. 2018, 9, 644. [Google Scholar] [CrossRef]
- Consortium, T.U. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2020, 49, D480–D489. [Google Scholar]
- Domsic, J.F.; Avvaru, B.S.; Kim, C.U.; Gruner, S.M.; Agbandje-McKenna, M.; Silverman, D.N.; McKenna, R. Entrapment of carbon dioxide in the active site of carbonic anhydrase II. J. Biol. Chem. 2008, 283, 30766–30771. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef]
- Peters, J.E.; Fricker, A.D.; Kapili, B.J.; Petassi, M.T. Heteromeric transposase elements: Generators of genomic islands across diverse bacteria. Mol. Microbiol. 2014, 93, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Waack, S.; Keller, O.; Asper, R.; Brodag, T.; Damm, C.; Fricke, W.F.; Surovcik, K.; Meinicke, P.; Merkl, R. Score-based prediction of genomic islands in prokaryotic genomes using hidden Markov models. BMC Bioinform. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Li, X.; Jiang, L.; Gong, L.; Geslin, C.; Shao, Z. Virus diversity and interactions with hosts in deep-sea hydrothermal vents. Microbiome 2022, 10, 235. [Google Scholar] [PubMed]
- Gestal, A.M.; Liew, E.F.; Coleman, N.V. Natural transformation with synthetic gene cassettes: New tools for integron research and biotechnology. Microbiology 2011, 157, 3349–3360. [Google Scholar] [PubMed]
- Goffredi, S.; Childress, J.; Desaulniers, N.; Lee, R.; Lallier, F.; Hammond, D. Inorganic carbon acquisition by the hydrothermal vent tubeworm Riftia pachyptila depends upon high external PCO2 and upon proton-equivalent ion transport by the worm. J. Exp. Biol. 1997, 200, 883–896. [Google Scholar] [CrossRef] [PubMed]
- De Cian, M.C.; Bailly, X.; Morales, J.; Strub, J.M.; Van Dorsselaer, A.; Lallier, F.H. Characterization of carbonic anhydrases from Riftia pachyptila, a symbiotic invertebrate from deep-sea hydrothermal vents. Proteins Struct. Funct. Bioinf. 2003, 51, 327–339. [Google Scholar] [CrossRef] [PubMed]
- De Cian, M.-C.; Andersen, A.C.; Bailly, X.; Lallier, F.H. Expression and localization of carbonic anhydrase and ATPases in the symbiotic tubeworm Riftia pachyptila. J. Exp. Biol. 2003, 206, 399–409. [Google Scholar] [CrossRef]
- Gonnella, G.; Adam, N.; Perner, M. Horizontal acquisition of hydrogen conversion ability and other habitat adaptations in the Hydrogenovibrio strains SP-41 and XCL-2. BMC Genom. 2019, 20, 339. [Google Scholar] [CrossRef]
- Fabry, V.J.; Seibel, B.A.; Feely, R.A.; Orr, J.C. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES J. Mar. Sci. 2008, 65, 414–432. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Integron Type | Integrase | CA Gene |
|---|---|---|---|
| Cycloclasticus sp. symbiont of Bathymodiolus heckerae Endosymbiont of Riftia pachyptila (vent Ph05) Sulfurovum sp. NBC37-1 | In0 | Intersection tyr intI | - |
| Integron 1: CALIN Integron 2: CALIN | - | α-CA β-CA | |
| Integron 1: CALIN Integron 2: CALIN | - | - | |
| Caldithrix abyssi Hydrogenovibrio crunogenus SP-41 | CALIN | - | - |
| Integron 1: CALIN Integron 2: CALIN Integron 3: Complete Integron 4: CALIN | Intersection tyr intI | - | |
| Thiomicrospira crunogena XCL-2 Bathymodiolus thermophilus thioautotrophic gill symbiont Endosymbiont of Tevnia jerichonana Halomonas sulfidaeris strain SST4 | Integron 1: CALIN Integron 2: CALIN | - | - |
| Integron 1: CALIN Integron 2: CALIN Integron 3: CALIN | - | - | |
| CALIN | - | β-CA | |
| CALIN | - | - | |
| Marinobacter sp. LQ44 | Integron 1: CALIN Integron 2: In0 | Intersection tyr intI | - |
| Sulfurimonas autotrophica | Integron 1: In0 Integron 2: CALIN | Intersection tyr intI | - |
| Endosymbiont of Bathymodiolus septemdierum | Integron 1: CALIN Integron 2: CALIN Integron 3: In0 Integron 4: CALIN Integron 5: CALIN | Intersection tyr intI | - |
| Hydrogenovibrio thermophilus Thermococcus barophilus strain CH5 | Complete | Intersection tyr intI | - |
| CALIN | - | - | |
| Cycloclasticus sp. symbiont of Bathymodiolus heckerae | In0 | Intersection tyr intI | - |
| Endosymbiont of Riftia pachyptila (vent Ph05) | Integron 1: CALIN Integron 2: CALIN | - | α-CA β-CA |
| Sulfurovum sp. NBC37-1 | Integron 1: CALIN Integron 2: CALIN | - | - |
| Caldithrix abyssi | CALIN | - | - |
| Hydrogenovibrio crunogenus SP-41 | Integron 1: CALIN Integron 2: CALIN Integron 3: Complete Integron 4: CALIN | Intersection tyr intI | - |
| Thiomicrospira crunogena XCL-2 | Integron 1: CALIN Integron 2: CALIN | - | - |
| Bathymodiolus thermophilus thioautotrophic gill symbiont | Integron 1: CALIN Integron 2: CALIN Integron 3: CALIN | - | - |
| Endosymbiont of Tevnia jerichonana | CALIN | - | β-CA |
| Halomonas sulfidaeris strain SST4 | CALIN | - | - |
| Marinobacter sp. LQ44 | Integron 1: CALIN Integron 2: In0 | Intersection tyr intI | - |
| Sulfurimonas autotrophica | Integron 1: In0 Integron 2: CALIN | Intersection tyr intI | - |
| Endosymbiont of Bathymodiolus septemdierum | Integron 1: CALIN Integron 2: CALIN Integron 3: In0 Integron 4: CALIN Integron 5: CALIN | Intersection tyr intI | - |
| Hydrogenovibrio thermophilus | Complete | Intersection tyr intI | - |
| Thermococcus barophilus strain CH5 | CALIN | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gheibzadeh, M.S.; Manyumwa, C.V.; Tastan Bishop, Ö.; Shahbani Zahiri, H.; Parkkila, S.; Zolfaghari Emameh, R. Genome Study of α-, β-, and γ-Carbonic Anhydrases from the Thermophilic Microbiome of Marine Hydrothermal Vent Ecosystems. Biology 2023, 12, 770. https://doi.org/10.3390/biology12060770
Gheibzadeh MS, Manyumwa CV, Tastan Bishop Ö, Shahbani Zahiri H, Parkkila S, Zolfaghari Emameh R. Genome Study of α-, β-, and γ-Carbonic Anhydrases from the Thermophilic Microbiome of Marine Hydrothermal Vent Ecosystems. Biology. 2023; 12(6):770. https://doi.org/10.3390/biology12060770
Chicago/Turabian StyleGheibzadeh, Mohammad Sadegh, Colleen Varaidzo Manyumwa, Özlem Tastan Bishop, Hossein Shahbani Zahiri, Seppo Parkkila, and Reza Zolfaghari Emameh. 2023. "Genome Study of α-, β-, and γ-Carbonic Anhydrases from the Thermophilic Microbiome of Marine Hydrothermal Vent Ecosystems" Biology 12, no. 6: 770. https://doi.org/10.3390/biology12060770
APA StyleGheibzadeh, M. S., Manyumwa, C. V., Tastan Bishop, Ö., Shahbani Zahiri, H., Parkkila, S., & Zolfaghari Emameh, R. (2023). Genome Study of α-, β-, and γ-Carbonic Anhydrases from the Thermophilic Microbiome of Marine Hydrothermal Vent Ecosystems. Biology, 12(6), 770. https://doi.org/10.3390/biology12060770