
Effects of Fumarate and Nitroglycerin on In Vitro Rumen Fermentation, Methane and Hydrogen Production, and on Microbiota

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Sample Collection and Chemical Analysis
2.4. Sample Collection and Chemical Analysis
2.5. DNA Extraction and Real-Time PCR
2.6. 16S rRNA Gene Sequencing and Data Analysis
2.7. Statistical Analysis
3. Results
3.1. Total Gas, Hydrogen and Methane Production
3.2. Fermentation Characteristics
3.3. The Quantification of Protozoa, Bacteria, Anaerobic Fungi and Archaea
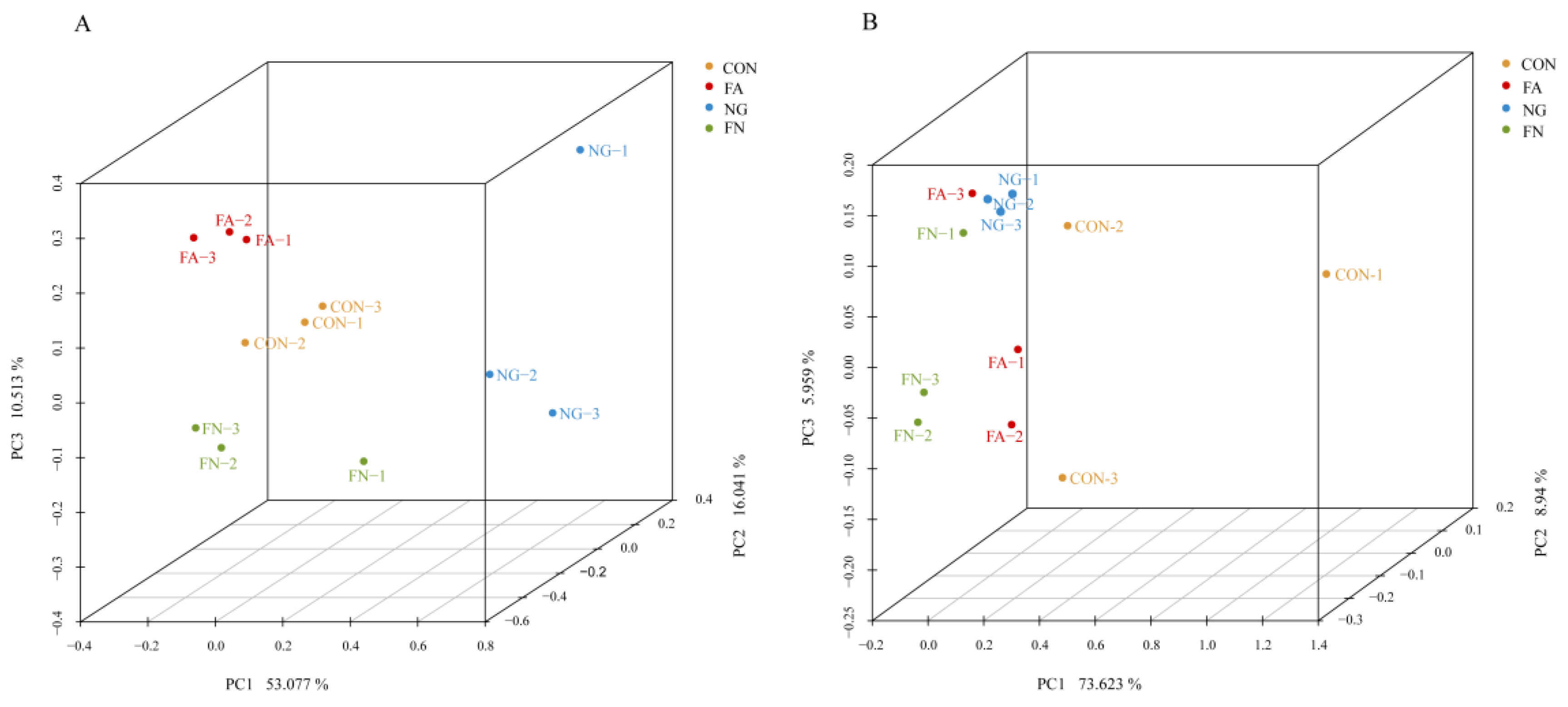
3.4. Bacterial Community
3.5. Archaeal Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Microbes | Primers | Sequences | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| Bacteria | F R | CCTACGGGAGGCAGCAG ATTACCGCGGCTGCTGG | 60 | [48] |
| Archaea | F R | GTGCTCCCCCGCCAATTCCT GCGGTGTGTGCAAGGAGC | 59 | [49] |
| Protozoa | F R | GCTTTCGWTGGTAGTGTATT CTTGCCCTCYAATCGTWCT | 55 | [50] |
| Fungi | F R | GAGGAAGTAAAAGTCGTAACAAGGTTTC CAAATTCACAAAGGGTAGGATGATT | 60 | [51] |
References
- Mizrahi, I.; Wallace, R.J.; Moraïs, S. The rumen microbiome: Balancing food security and environmental impacts. Nat. Rev. Microbiol. 2021, 19, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.J.; Henderson, B.; Makkar, H.; Hristov, A.N.; Oosting, S. Tackling Climate Change through Livestock: A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitta, D.; Indugu, N.; Narayan, K.; Hennessy, M. Symposium review: Understanding the role of the rumen microbiome in enteric methane mitigation and productivity in dairy cows. J. Dairy Sci. 2022, 105, 8569–8585. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.A.; Johnson, D. Methane Emissions from Cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef]
- Ocko, I.B.; Sun, T.; Shindell, D.; Oppenheimer, M.; Hristov, A.N.; Pacala, S.W.; Mauzerall, D.L.; Xu, Y.; Hamburg, S.P. Acting rapidly to deploy readily available methane mitigation measures by sector can immediately slow global warming. Environ. Res. Lett. 2021, 16, 054042. [Google Scholar] [CrossRef]
- Eckard, R.J.; Grainger, C.; de Klein, C.A.M. Options for the abatement of methane and nitrous oxide from ruminant production: A review. Livest. Sci. 2010, 130, 47–56. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [Green Version]
- Abecia, L.; Toral, P.; Martín-García, A.; Martínez, G.; Tomkins, N.; Molina-Alcaide, E.; Newbold, C.; Yáñez-Ruiz, D. Effect of bromochloromethane on methane emission, rumen fermentation pattern, milk yield, and fatty acid profile in lactating dairy goats. J. Dairy Sci. 2012, 95, 2027–2036. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.A.; Wallace, R.J.; Rowe, A.; Price, J.; Yáñez-Ruiz, D.R.; Murray, P.; Newbold, C.J. Encapsulated fumaric acid as a feed ingredient to decrease ruminal methane emissions. Anim. Feed. Sci. Technol. 2009, 152, 62–71. [Google Scholar] [CrossRef]
- van Zijderveld, S.; Gerrits, W.; Apajalahti, J.; Newbold, J.; Dijkstra, J.; Leng, R.; Perdok, H. Nitrate and sulfate: Effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J. Dairy Sci. 2010, 93, 5856–5866. [Google Scholar] [CrossRef] [Green Version]
- Melgar, A.; Lage, C.; Nedelkov, K.; Räisänen, S.; Stefenoni, H.; Fetter, M.; Chen, X.; Oh, J.; Duval, S.; Kindermann, M.; et al. Enteric methane emission, milk production, and composition of dairy cows fed 3-nitrooxypropanol. J. Dairy Sci. 2021, 104, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Schilde, M.; von Soosten, D.; Hüther, L.; Meyer, U.; Zeyner, A.; Dänicke, S. Effects of 3-nitrooxypropanol and varying concentrate feed proportions in the ration on methane emission, rumen fermentation and performance of periparturient dairy cows. Arch. Anim. Nutr. 2021, 75, 79–104. [Google Scholar] [CrossRef]
- Kinley, R.D.; Martinez-Fernandez, G.; Matthews, M.K.; de Nys, R.; Magnusson, M.; Tomkins, N.W. Mitigating the carbon footprint and improving productivity of ruminant livestock agriculture using a red seaweed. J. Clean. Prod. 2020, 259, 120836. [Google Scholar] [CrossRef]
- Stefenoni, H.; Räisänen, S.; Cueva, S.; Wasson, D.; Lage, C.; Melgar, A.; Fetter, M.; Smith, P.; Hennessy, M.; Vecchiarelli, B.; et al. Effects of the macroalga Asparagopsis taxiformis and oregano leaves on methane emission, rumen fermentation, and lactational performance of dairy cows-Sciencedirect. J. Dairy Sci. 2021, 104, 4157–4173. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-W.; Chen, Y.-S.; Cheng, Y.-H.; Yang, C.-M.J.; Chang, C.-T.; Cheng, Y.-S. Effects of fumarate on ruminal ammonia accumulation and fiber digestion in vitro and nutrient utilization in dairy does. J. Dairy Sci. 2010, 93, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Meng, Z.; Wang, J.; Cheng, Y.; Zhu, W. Effect of nitrooxy compounds with different molecular structures on the rumen methanogenesis, metabolic profile, and methanogenic community. Curr. Microbiol. 2017, 74, 891–898. [Google Scholar] [CrossRef]
- Xie, F.; Zhang, L.; Jin, W.; Meng, Z.; Cheng, Y.; Wang, J.; Zhu, W. Methane emission, rumen fermentation, and microbial community response to a nitrooxy compound in low-quality forage fed Hu sheep. Curr. Microbiol. 2019, 76, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Ungerfeld, E.M.; Kohn, R.A.; Wallace, R.J.; Newbold, C.J. A meta-analysis of fumarate effects on methane production in ruminal batch cultures. J. Anim. Sci. 2007, 85, 2556–2563. [Google Scholar] [CrossRef]
- Zhou, Y.; Jin, W.; Xie, F.; Mao, S.; Cheng, Y.; Zhu, W. The role of Methanomassiliicoccales in trimethylamine metabolism in the rumen of dairy cows. Animal 2021, 15, 100259. [Google Scholar] [CrossRef]
- Yáñez-Ruiz, D.; Bannink, A.; Dijkstra, J.; Kebreab, E.; Morgavi, D.; O’kiely, P.; Reynolds, C.; Schwarm, A.; Shingfield, K.; Yu, Z.; et al. Design, implementation and interpretation of in vitro batch culture experiments to assess enteric methane mitigation in ruminants—A review. Anim. Feed. Sci. Technol. 2016, 216, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Martinez Fernandez, G.; Abecia, L.; Arco, A.; Cantalapiedra-Hijar, G.; Martín-García, A.I.; Molina-Alcaide, E.; Kindermann, M.; Duval, S.; Yáñez-Ruiz, D.R. Effects of ethyl-3-nitrooxy propionate and 3-nitrooxypropanol on ruminal fermentation, microbial abundance, and methane emissions in sheep. J. Dairy Sci. 2014, 97, 3790–3799. [Google Scholar] [CrossRef] [Green Version]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S.; McAllan, A.B.; France, J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed. Sci. Technol. 1994, 48, 185–197. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Jin, D.; Zhao, S.; Zheng, N.; Bu, D.; Beckers, Y.; Denman, S.E.; McSweeney, C.S.; Wang, J. Differences in ureolytic bacterial composition between the rumen digesta and rumen wall based on urec gene classification. Front. Microbiol. 2017, 8, 385. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Mitter, E.K.; de Freitas, J.R.; Germida, J.J. Bacterial root microbiome of plants growing in oil sands reclamation covers. Front. Microbiol. 2017, 8, 849. [Google Scholar] [CrossRef] [Green Version]
- Melgar, A.; Harper, M.; Oh, J.; Giallongo, F.; Young, M.; Ott, T.; Duval, S.; Hristov, A. Effects of 3-nitrooxypropanol on rumen fermentation, lactational performance, and resumption of ovarian cyclicity in dairy cows. J. Dairy Sci. 2020, 103, 410–432. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Meng, Q.; Yu, Z. Effects of methanogenic inhibitors on methane production and abundances of methanogens and cellulolytic bacteria in in vitro ruminal cultures. Appl. Environ. Microbiol. 2011, 77, 2634–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Pérez, A.; Okine, E.; Guan, L.; Duval, S.; Kindermann, M.; Beauchemin, K. Effects of 3-nitrooxypropanol on methane production using the rumen simulation technique (Rusitec). Anim. Feed. Sci. Technol. 2015, 209, 98–109. [Google Scholar] [CrossRef]
- Yang, C.; Mao, S.; Long, L.; Zhu, W. Effect of disodium fumarate on microbial abundance, ruminal fermentation and methane emission in goats under different forage: Concentrate ratios. Animal 2012, 6, 1788–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, M.A.; Reddy, C.A. Hydrogenase activity and the H2-fumarate electron transport system in Bacteroides fragilis. J. Bacteriol. 1977, 131, 922–928. [Google Scholar] [CrossRef] [Green Version]
- Ungerfeld, E.; Kohn, R. The Role of Thermodynamics in the Control of Ruminal Fermentation. In Ruminant Physiology; Sejrsen, K., Hvelplund, T., Nielsen, M.O., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 55–85. [Google Scholar]
- Cord-Ruwisch, R.; Conrad, R.; Seitz, H.-J. The capacity of hydrogenotrophic anaerobic bacteria to compete for traces of hydrogen depends on the redox potential of the terminal electron acceptor. Arch. Microbiol. 1988, 149, 350–357. [Google Scholar] [CrossRef]
- Bayaru, E.; Kanda, S.; Kamada, T.; Itabashi, H.; Andoh, S.; Nishida, T.; Ishida, M.; Itoh, T.; Nagara, K.; Isobe, Y. Effect of fumaric acid on methane production, rumen fermentation and digestibility of cattle fed roughage alone. Nihon Chikusan Gakkaiho 2001, 72, 139–146. [Google Scholar] [CrossRef] [Green Version]
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef]
- Molano, G.; Knight, T.W.; Clark, H. Fumaric acid supplements have no effect on methane emissions per unit of feed intake in wether lambs. Aust. J. Exp. Agric. 2008, 48, 165–168. [Google Scholar] [CrossRef]
- Lee, S.S.; Hsu, J.-T.; Mantovani, H.C.; Russell, J.B. The effect of bovicin HC5, a bacteriocin from Streptococcus bovisHC5, on ruminal methane production in vitro. FEMS Microbiol. Lett. 2002, 217, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, S.; Zhang, G.; Zhu, W. Effect of disodium fumarate on ruminal metabolism and rumen bacterial communities as revealed by denaturing gradient gel electrophoresis analysis of 16S ribosomal DNA. Anim. Feed. Sci. Technol. 2008, 140, 293–306. [Google Scholar] [CrossRef]
- Ortiz-Chura, A.; Gere, J.; Marcoppido, G.; Depetris, G.; Cravero, S.; Faverín, C.; Pinares-Patiño, C.; Cataldi, A.; Cerón-Cucchi, M.E. Dynamics of the ruminal microbial ecosystem, and inhibition of methanogenesis and propiogenesis in response to nitrate feeding to Holstein calves. Anim. Nutr. 2021, 7, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Xue, C.; Liu, J.; Yin, Y.; Zhu, W.; Mao, S. Effects of disodium fumarate on in vitro rumen fermentation, the production of lipopolysaccharide and biogenic amines, and the rumen bacterial community. Curr. Microbiol. 2017, 74, 1337–1342. [Google Scholar] [CrossRef] [PubMed]
- Vlaeminck, B.; Fievez, V.; Tamminga, S.; Dewhurst, R.; van Vuuren, A.; De Brabander, D.; Demeyer, D. Milk odd- and branched-chain fatty acids in relation to the rumen fermentation pattern. J. Dairy Sci. 2006, 89, 3954–3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yohe, T.; Enger, B.; Wang, L.; Tucker, H.; Ceh, C.; Parsons, C.; Yu, Z.; Daniels, K. Short communication: Does early-life administration of a Megasphaera elsdenii probiotic affect long-term establishment of the organism in the rumen and alter rumen metabolism in the dairy calf? J. Dairy Sci. 2018, 101, 1747–1751. [Google Scholar] [CrossRef]
- Duin, E.C.; Wagner, T.; Shima, S.; Prakash, D.; Cronin, B.; Yáñez-Ruiz, D.R.; Duval, S.; Rümbeli, R.; Stemmler, R.T.; Thauer, R.K.; et al. Mode of action uncovered for the specific reduction of methane emissions from ruminants by the small molecule 3-nitrooxypropanol. Proc. Natl. Acad. Sci. USA 2016, 113, 6172–6177. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.U.; Schmitz-Esser, S.; Klevenhusen, F.; Podstatzky-Lichtenstein, L.; Wagner, M.; Zebeli, Q. Grain-rich diets differently alter ruminal and colonic abundance of microbial populations and lipopolysaccharide in goats. Anaerobe 2013, 20, 65–73. [Google Scholar] [CrossRef]
- Jeyanathan, J.; Kirs, M.; Ronimus, R.S.; Hoskin, S.O.; Janssen, P.H. Methanogen community structure in the rumens of farmed sheep, cattle and red deer fed different diets. FEMS Microbiol. Ecol. 2011, 76, 311–326. [Google Scholar] [CrossRef]
- Sylvester, J.T.; Karnati, S.K.R.; Yu, Z.; Morrison, M.; Firkins, J.L. Development of an assay to quantify rumen ciliate protozoal biomass in cows using real-time PCR. J. Nutr. 2004, 134, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef]

| Items | Groups 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | FA | NG | FN | FA | NG | NG * FA | ||
| Total gas (mL) | 203.67 c | 245.67 a | 201.67 c | 226.67 b | 5.46 | <0.001 | <0.001 | <0.001 |
| Hydrogen (mL) | 0.06 c | 0.11 c | 9.68 a | 6.38 b | 1.25 | <0.001 | <0.001 | <0.001 |
| Methane (mL) | 21.30 b | 24.07 a | 0.00 c | 0.00 c | 3.44 | 0.020 | <0.001 | 0.020 |
| Items | Groups 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | FA | NG | FN | FA | NG | NG * FA | ||
| pH | 6.63 | 6.34 | 6.21 | 6.59 | 0.03 | 0.053 | 0.848 | 0.187 |
| Total VFA (mmol/L) | 60.65 ab | 73.63 a | 45.32 b | 60.01 ab | 3.44 | 0.007 | 0.005 | 0.829 |
| Acetate (mmol/L) | 32.88 a | 36.87 a | 20.82 b | 25.06 b | 2.08 | 0.073 | <0.001 | 0.950 |
| Propionate (mmol/L) | 19.07 b | 26.61 a | 15.34 b | 23.93 a | 1.40 | <0.001 | 0.026 | 0.665 |
| A:P | 1.72 a | 1.39 b | 1.36 b | 1.05 c | 0.08 | 0.001 | <0.001 | 0.846 |
| Butyrate (mmol/L) | 6.79 | 7.66 | 5.80 | 8.13 | 0.65 | 0.280 | 0.855 | 0.612 |
| Isobutyrate (mmol/L) | 0.30 b | 0.48 a | 0.06 c | 0.19 bc | 0.03 | 0.001 | <0.001 | 0.354 |
| Valerate (mmol/L) | 1.24 b | 1.33 b | 3.30 a | 2.63 a | 0.17 | 0.192 | <0.001 | 0.099 |
| Isovalerate (mmol/L) | 0.37 b | 0.69 a | 0.01 c | 0.07 c | 0.05 | <0.001 | <0.001 | 0.001 |
| N-NH3 (mol/L) | 10.68 b | 12.62 a | 9.46 b | 9.52 b | 0.20 | 0.012 | <0.001 | 0.017 |
| MCP (mg/dL) | 3.33 a | 3.35 a | 2.99 c | 3.16 b | 0.03 | <0.001 | <0.001 | <0.001 |
| Lactate (mmol/L) | 0.81 | 0.63 | 0.72 | 0.71 | 0.02 | 0.258 | 0.988 | 0.276 |
| Items | Groups 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | FA | NG | FN | FA | NG | NG * FA | ||
| Bacteria (log10/L) | 9.31 | 9.32 | 9.48 | 9.42 | 0.03 | 0.647 | 0.053 | 0.538 |
| Protozoa (log10/L) | 6.55 | 6.14 | 6.01 | 6.85 | 0.16 | 0.488 | 0.778 | 0.063 |
| Anaerobic fungi (log10/L) | 6.41 | 6.82 | 6.73 | 6.74 | 0.10 | 0.336 | 0.567 | 0.351 |
| Archaea (log10/L) | 7.93 bc | 8.35 a | 7.64 c | 8.06 ab | 0.09 | 0.001 | 0.011 | 0.971 |
| Items | Groups 1 | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | FA | NG | FN | |||
| Bacterial | ||||||
| Reads | 31,504.7 | 33,801.3 | 32,442.3 | 32,255.3 | 477.6 | 0.439 |
| Coverage | 0.99 | 0.99 | 0.99 | 0.99 | 0.00 | 0.532 |
| Chao 1 | 2108.0 | 2208.0 | 1723.0 | 2052.0 | 85.39 | 0.144 |
| Shannon | 6.27 a | 6.17 a | 5.36 b | 6.16 a | 0.14 | 0.043 |
| Simpson | 0.01 | 0.01 | 0.05 | 0.01 | 0.01 | 0.063 |
| Archaea | ||||||
| Reads | 50,258.0 | 54,952.0 | 34,009.0 | 50,166.0 | 3133.5 | 0.060 |
| Coverage | 0.98 | 0.98 | 0.97 | 0.98 | 0.00 | 0.006 |
| Chao 1 | 2744.0 | 2870.0 | 2891.0 | 3039.0 | 92.67 | 0.790 |
| Shannon | 4.78 | 4.90 | 4.77 | 5.00 | 0.08 | 0.639 |
| Simpson | 0.04 | 0.04 | 0.04 | 0.03 | 0.00 | 0.532 |
| Items | Groups 1 | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | FA | NG | FN | |||
| Bacteroidetes | 39.10 ab | 45.33 ab | 25.68 b | 55.39 a | 3.84 | 0.015 |
| Firmicutes | 36.18 b | 30.31 b | 56.25 a | 30.05 b | 3.73 | 0.009 |
| Proteobacteria | 16.51 a | 16.77 a | 7.69 b | 6.36 b | 1.63 | 0.004 |
| Spirochaetae | 2.20 ab | 2.42 a | 0.48 b | 2.65 a | 0.32 | 0.026 |
| Actinobacteria | 1.93 b | 1.00 b | 6.54 a | 1.89 b | 0.73 | 0.004 |
| Candidate_division_SR1 | 0.90 | 0.82 | 0.49 | 1.17 | 0.11 | 0.158 |
| Candidate_division_TM7 | 1.14 a | 0.62 b | 0.56 b | 0.49 b | 0.09 | 0.006 |
| Cyanobacteria | 0.82 | 1.25 | 1.50 | 1.20 | 0.19 | 0.702 |
| Items | Groups 1 | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| CON | FA | NG | FN | ||||
| Phylum | Genus | ||||||
| Bacteroidetes | Prevotella | 22.14 | 27.05 | 14.46 | 34.5 | 2.84 | 0.053 |
| Rikenellaceae_RC9_gut_group | 6.56 | 8.36 | 5.62 | 7.37 | 0.46 | 0.187 | |
| Unclassified Prevotellaceae | 3.77 | 3.35 | 2.14 | 6.71 | 0.72 | 0.075 | |
| Unclassified BS11_gut_group | 2.31 a | 2.58 a | 0.78 b | 0.96 b | 0.26 | 0.002 | |
| Unclassified S24-7 | 1.81 | 0.68 | 1.32 | 2.27 | 0.25 | 0.108 | |
| Unclassified RF16 | 1.78 | 2.13 | 1.03 | 2.43 | 0.24 | 0.192 | |
| Firmicutes | Unclassified Ruminococcaceae | 7.62 a | 6.00 ab | 3.92 b | 4.26 ab | 0.55 | 0.030 |
| Unclassified Christensenellaceae | 4.09 | 2.76 | 3.13 | 2.63 | 0.31 | 0.370 | |
| Streptococcus2 | 3.40 b | 2.20 b | 26.94 a | 5.69 b | 3.40 | <0.050 | |
| Butyrivibrio | 2.80 | 2.00 | 3.18 | 2.20 | 0.36 | 0.689 | |
| Succiniclasticum | 2.20 | 2.43 | 1.12 | 1.48 | 0.21 | 0.068 | |
| Ruminococcus | 2.13 | 1.30 | 2.38 | 1.30 | 0.19 | 0.063 | |
| Quinella | 1.63 | 2.15 | 1.12 | 1.53 | 0.18 | 0.238 | |
| Proteobacteria | Succinivibrio | 8.33 a | 8.94 a | 4.53 ab | 2.31 b | 0.98 | 0.017 |
| Unclassified Succinivibrionaceae | 3.93 | 4.32 | 1.71 | 2.2 | 0.42 | 0.064 | |
| Ruminobacter | 3.77 a | 2.32 ab | 1.08 b | 1.48 b | 0.35 | 0.006 | |
| Spirochaetae | Treponema | 2.18 ab | 2.38 ab | 0.47 b | 2.64 a | 0.32 | 0.027 |
| Actinobacteria | Bifidobacterium | 1.05 b | 0.48 b | 5.66 a | 1.46 b | 0.67 | 0.024 |
| Items | Groups 1 | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | FA | NG | FN | |||
| Methanobacteriales | 99.57 a | 96.59 b | 98.43 a | 90.06 c | 1.13 | <0.001 |
| Methanomassiliicoccales | 0.43 c | 3.41 b | 1.57 c | 9.94 a | 1.13 | <0.001 |
| Items | Groups 1 | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | FA | NG | FN | |||
| Methanobrevibacter gottschalkii clade | 74.93 a | 68.46 ab | 68.11 ab | 61.89 b | 2.17 | 0.002 |
| Methanobrevibacter boviskoreani clade | 6.35 | 1.20 | 0.84 | 0.91 | 1.68 | 0.055 |
| Methanobrevibacter ruminantium clade | 13.47 | 20.28 | 23.88 | 22.49 | 2.53 | 0.082 |
| Methanosphaera sp. ISO3-F5 | 4.02 | 6.47 | 5.46 | 4.60 | 0.65 | 0.268 |
| Group12 sp. ISO4-H5 | 0.27 b | 1.57 b | 0.78 b | 6.45 a | 1.08 | 0.019 |
| Group9 sp. ISO4-G1 | 0.06 c | 1.05 b | 0.40 c | 2.30 a | 0.37 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhao, S.; Meng, Z.; Gao, Y.; Miao, J.; Mao, S.; Jin, W. Effects of Fumarate and Nitroglycerin on In Vitro Rumen Fermentation, Methane and Hydrogen Production, and on Microbiota. Biology 2023, 12, 1011. https://doi.org/10.3390/biology12071011
Li J, Zhao S, Meng Z, Gao Y, Miao J, Mao S, Jin W. Effects of Fumarate and Nitroglycerin on In Vitro Rumen Fermentation, Methane and Hydrogen Production, and on Microbiota. Biology. 2023; 12(7):1011. https://doi.org/10.3390/biology12071011
Chicago/Turabian StyleLi, Jichao, Shengwei Zhao, Zhenxiang Meng, Yunlong Gao, Jing Miao, Shengyong Mao, and Wei Jin. 2023. "Effects of Fumarate and Nitroglycerin on In Vitro Rumen Fermentation, Methane and Hydrogen Production, and on Microbiota" Biology 12, no. 7: 1011. https://doi.org/10.3390/biology12071011
APA StyleLi, J., Zhao, S., Meng, Z., Gao, Y., Miao, J., Mao, S., & Jin, W. (2023). Effects of Fumarate and Nitroglycerin on In Vitro Rumen Fermentation, Methane and Hydrogen Production, and on Microbiota. Biology, 12(7), 1011. https://doi.org/10.3390/biology12071011