
Mesenchymal Stem Cells and Begacestat Mitigate Amyloid-β 25–35-Induced Cognitive Decline in Rat Dams and Hippocampal Deteriorations in Offspring


 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Surgical Procedure
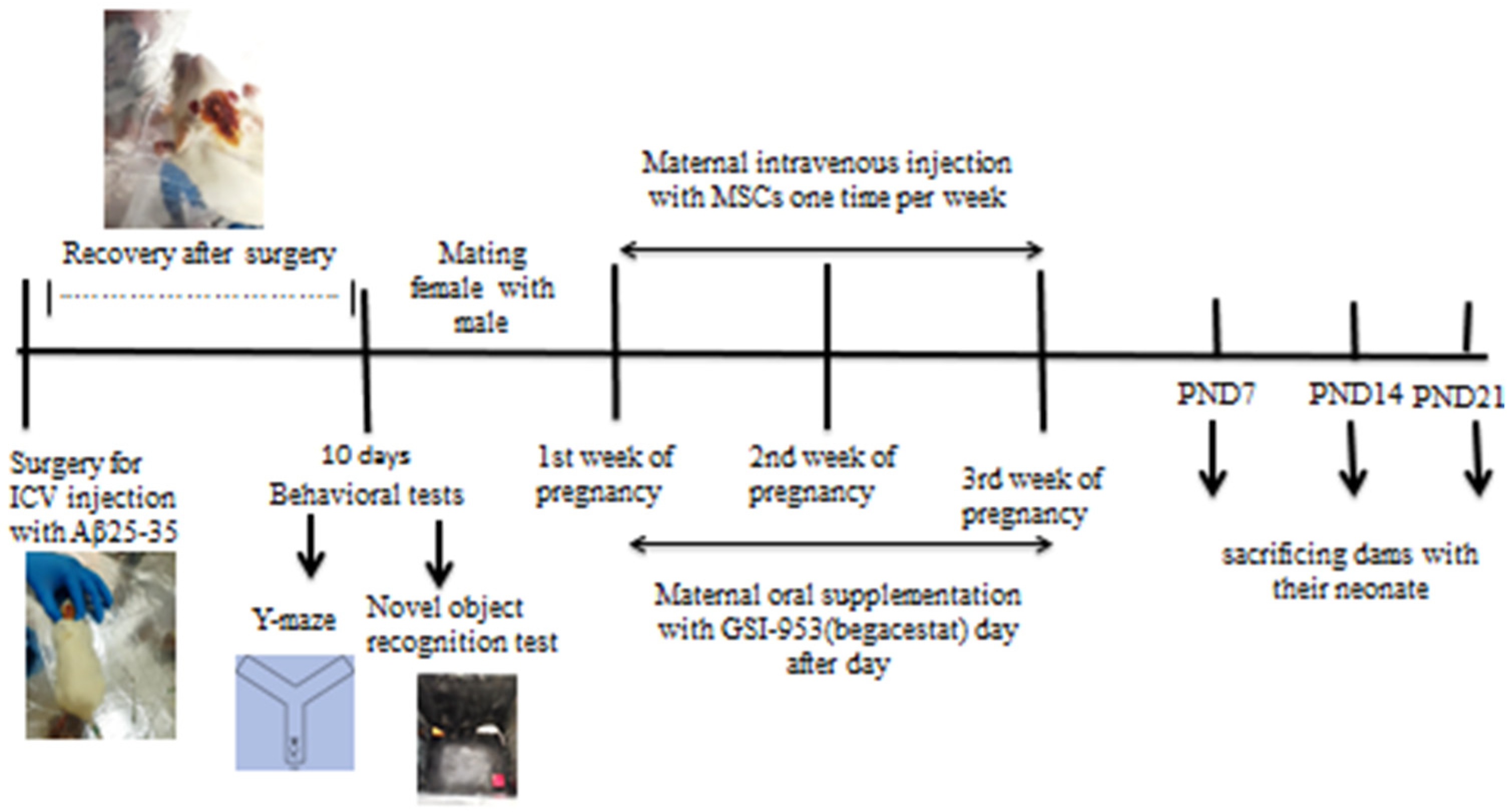
2.3. Animal Grouping: (Figure 1)

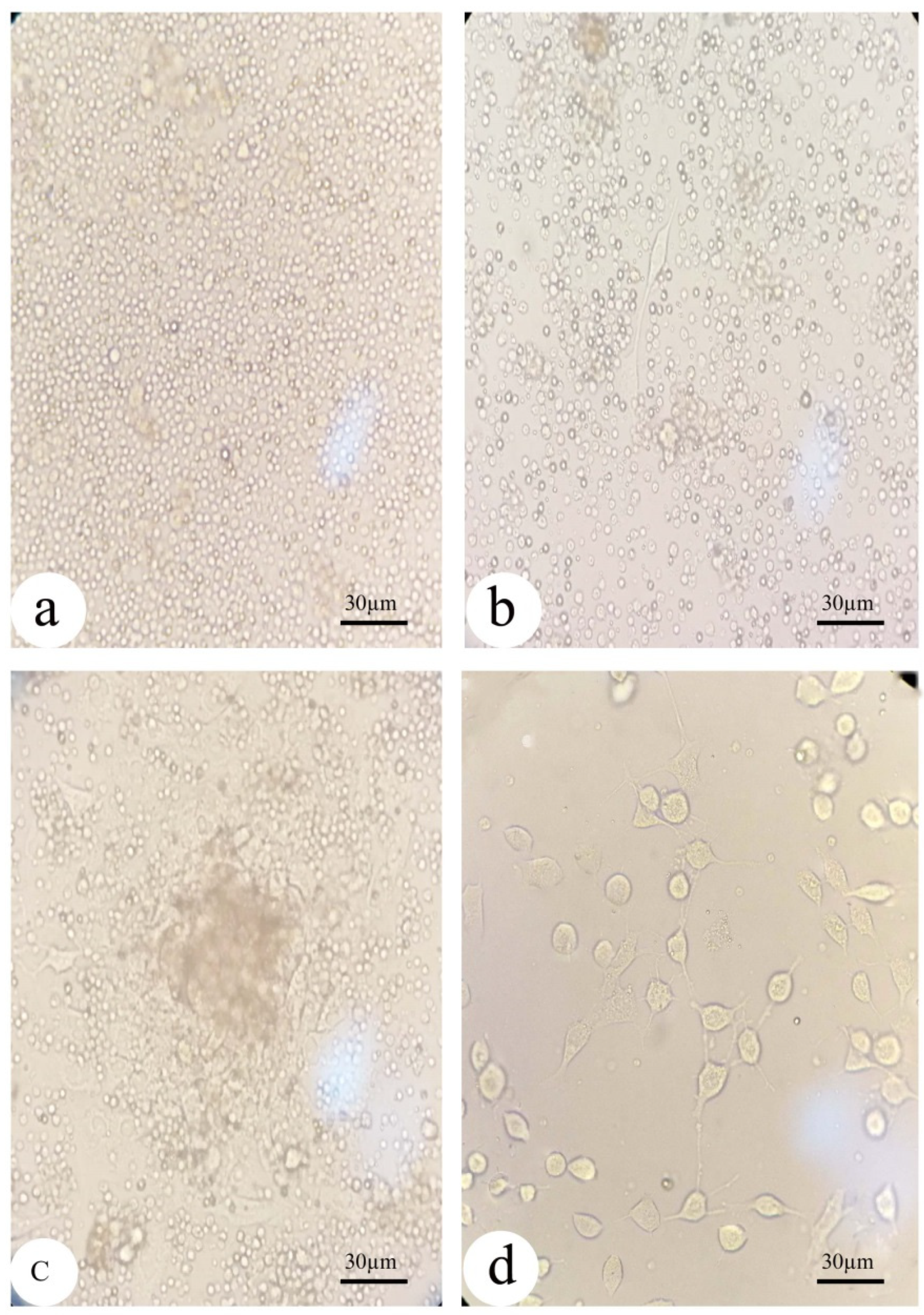
2.4. Isolation of BM-MSCs from Rats: (Figure 2)

2.5. Behavioral Tests
2.5.1. Y-Maze Test
2.5.2. Novel Object Recognition Test
2.6. Histological Analysis of the Newborn Hippocampus
2.7. Congo Red Staining
2.8. Immunohistochemistry
2.9. ELISA Assay
2.10. qRT-PCR
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
3.1. Aβ 25–35 i.c.v. Injection Impaired Working and Object Recognition Memories in Adult Female Rats
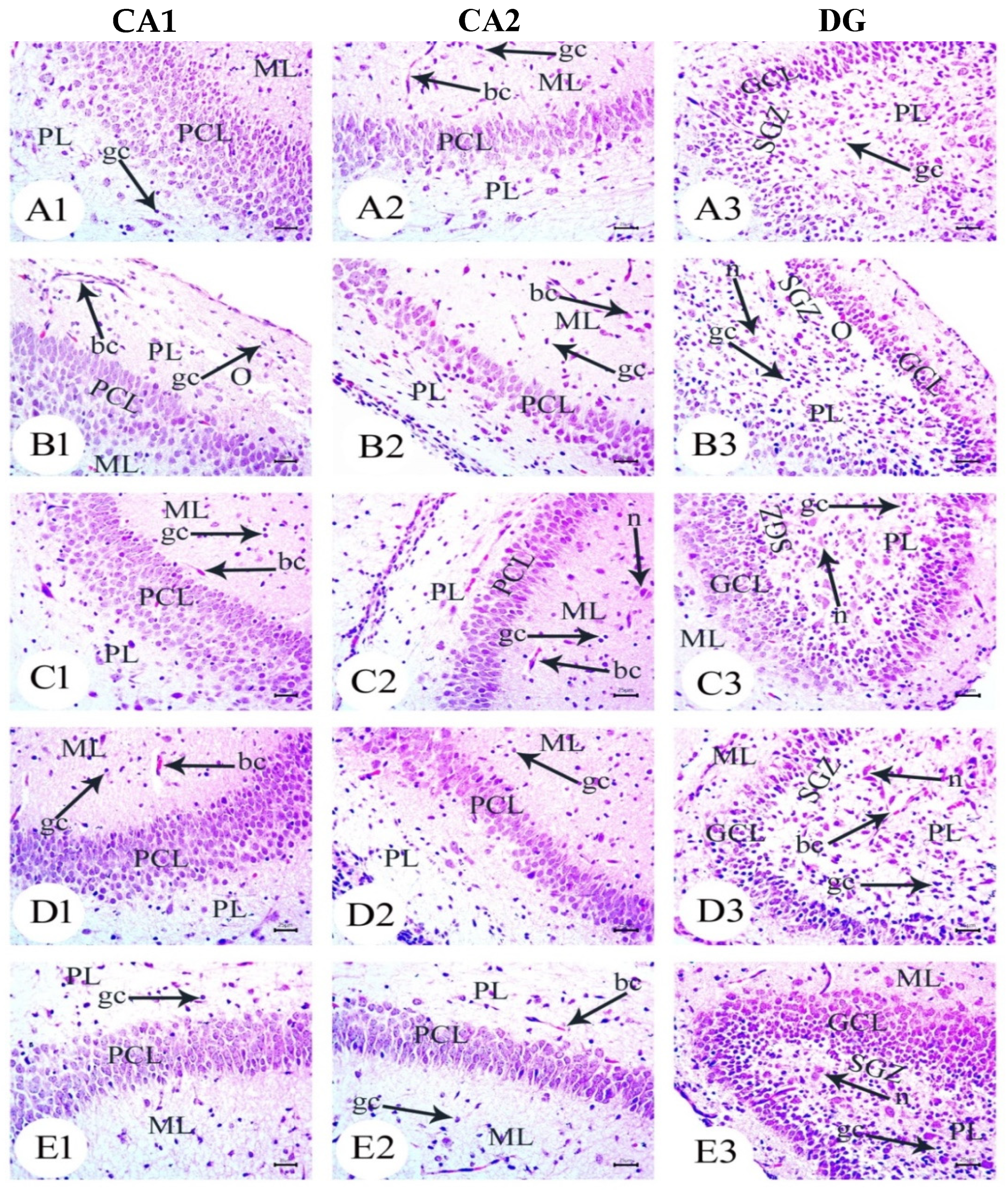
3.2. Aβ 25–35 i.c.v. Injection in Dams Disrupted Development in Offspring, an Effect Reversed by BM-MSC and GSI-953 Treatments
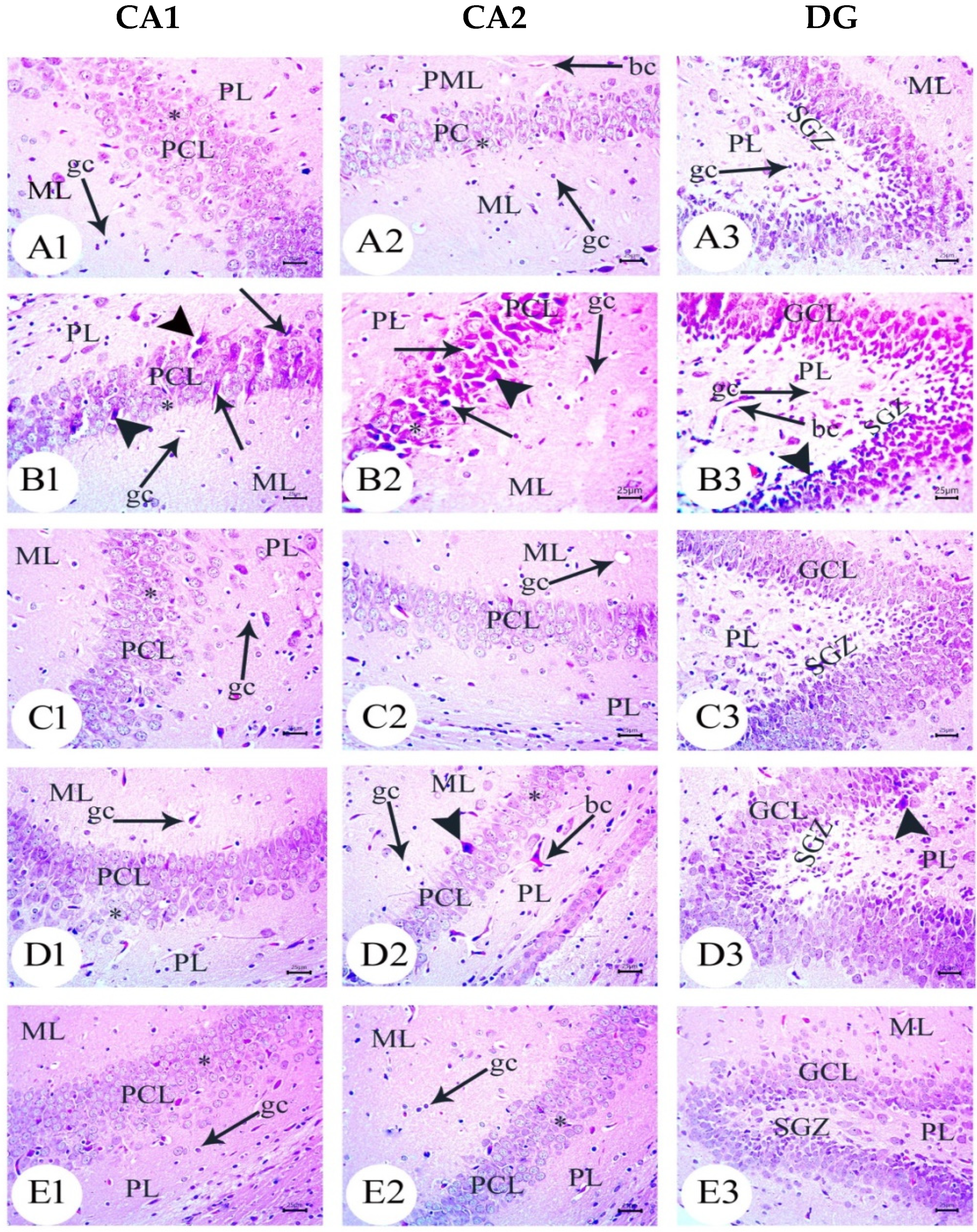
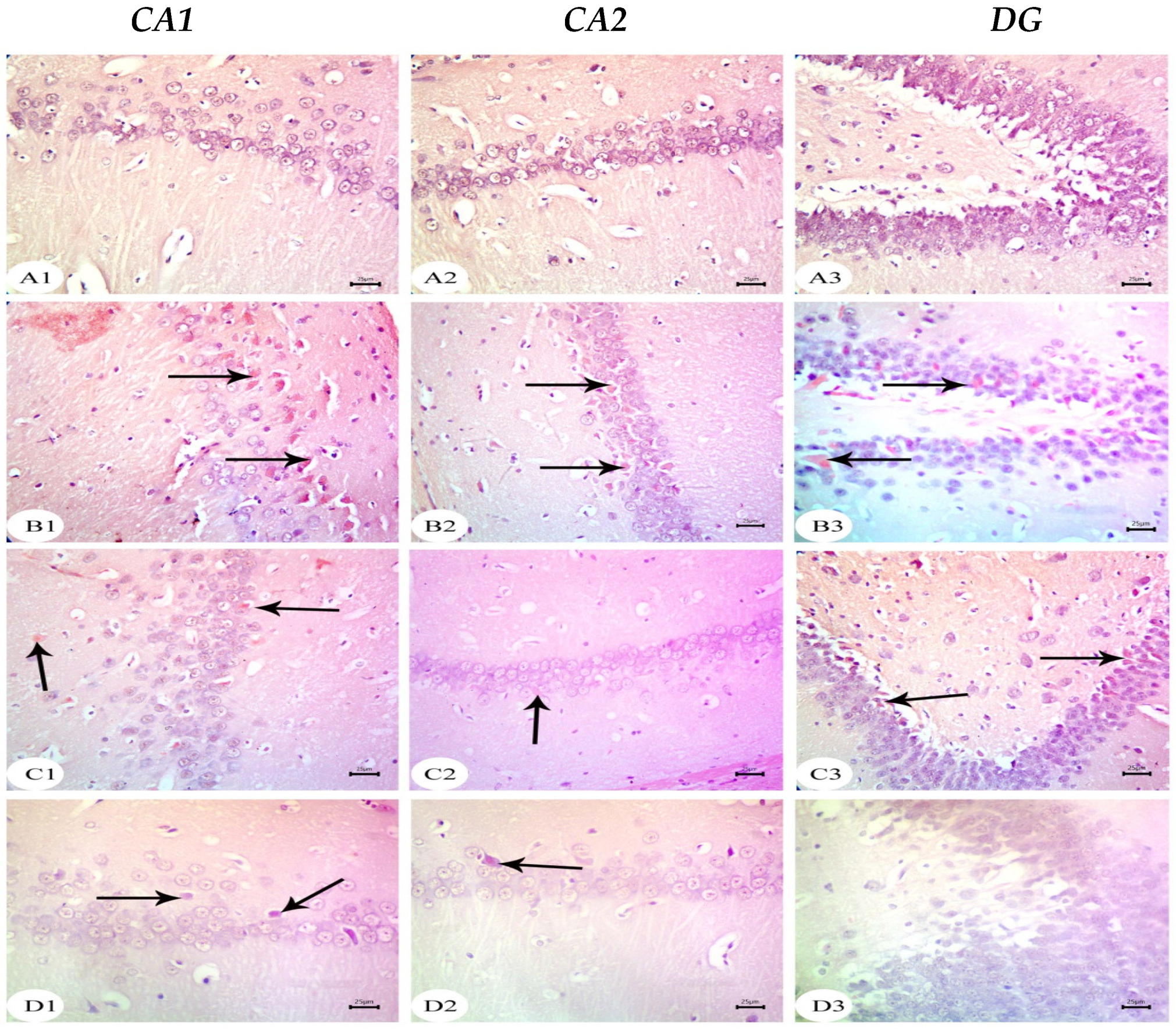
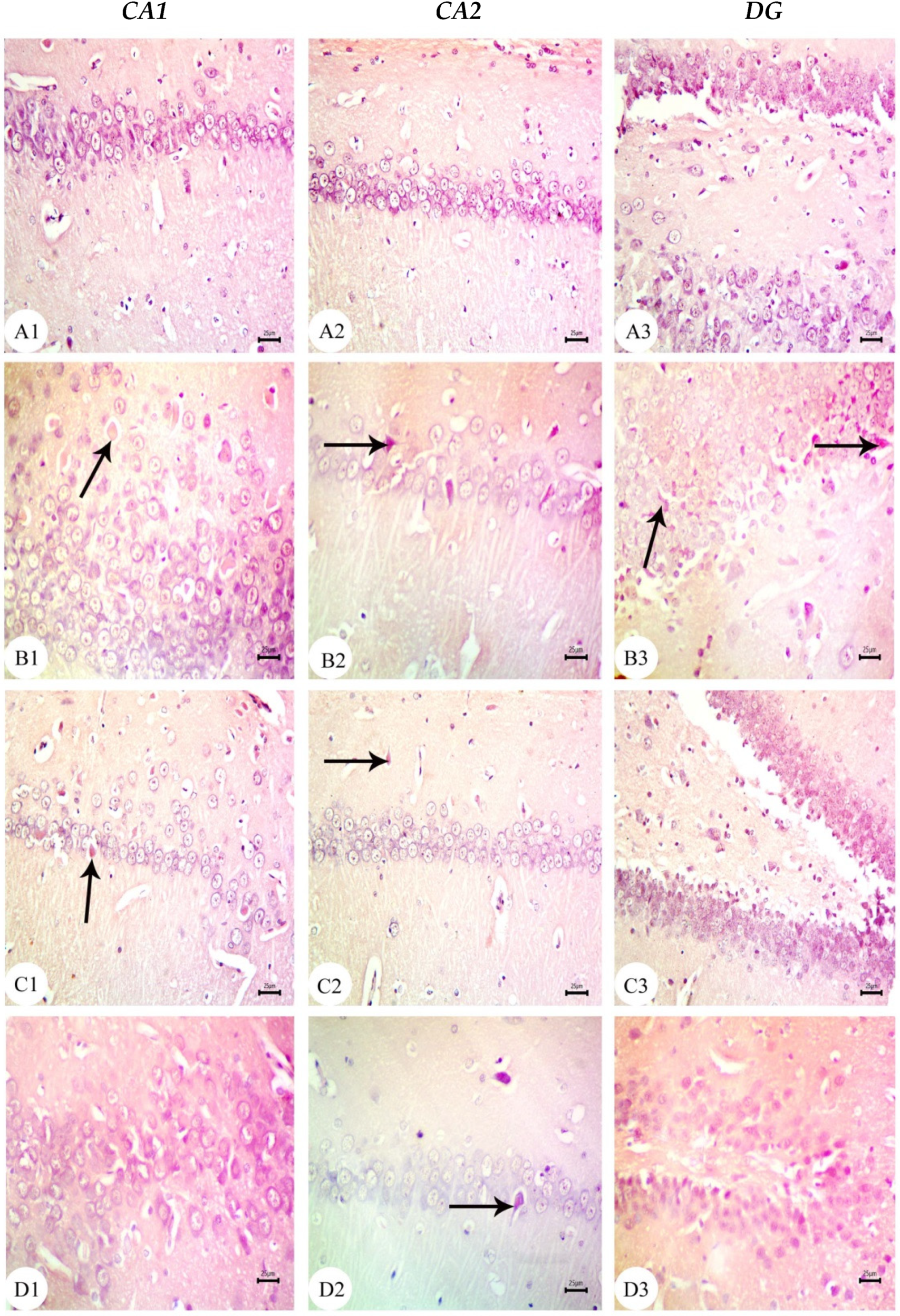
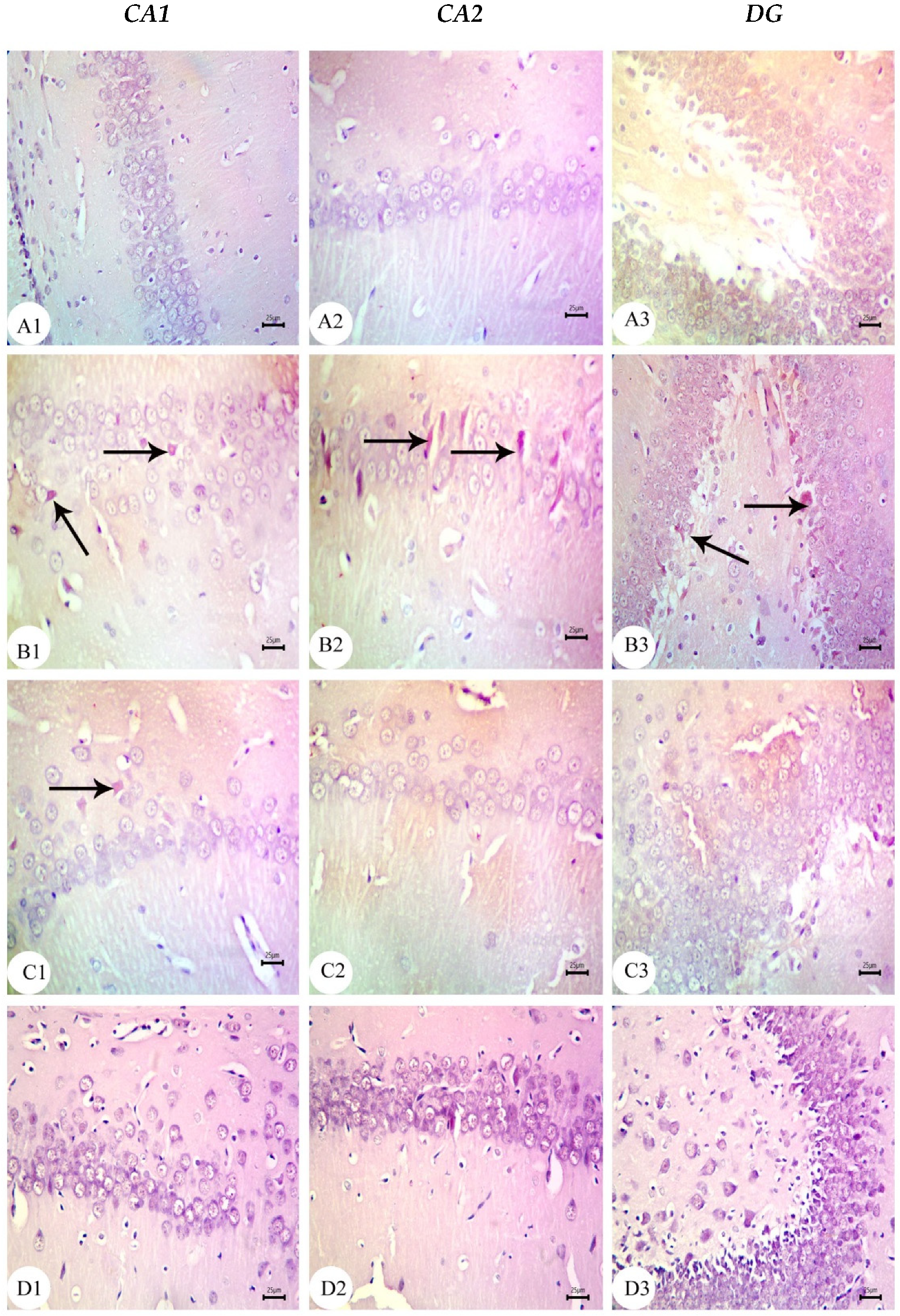
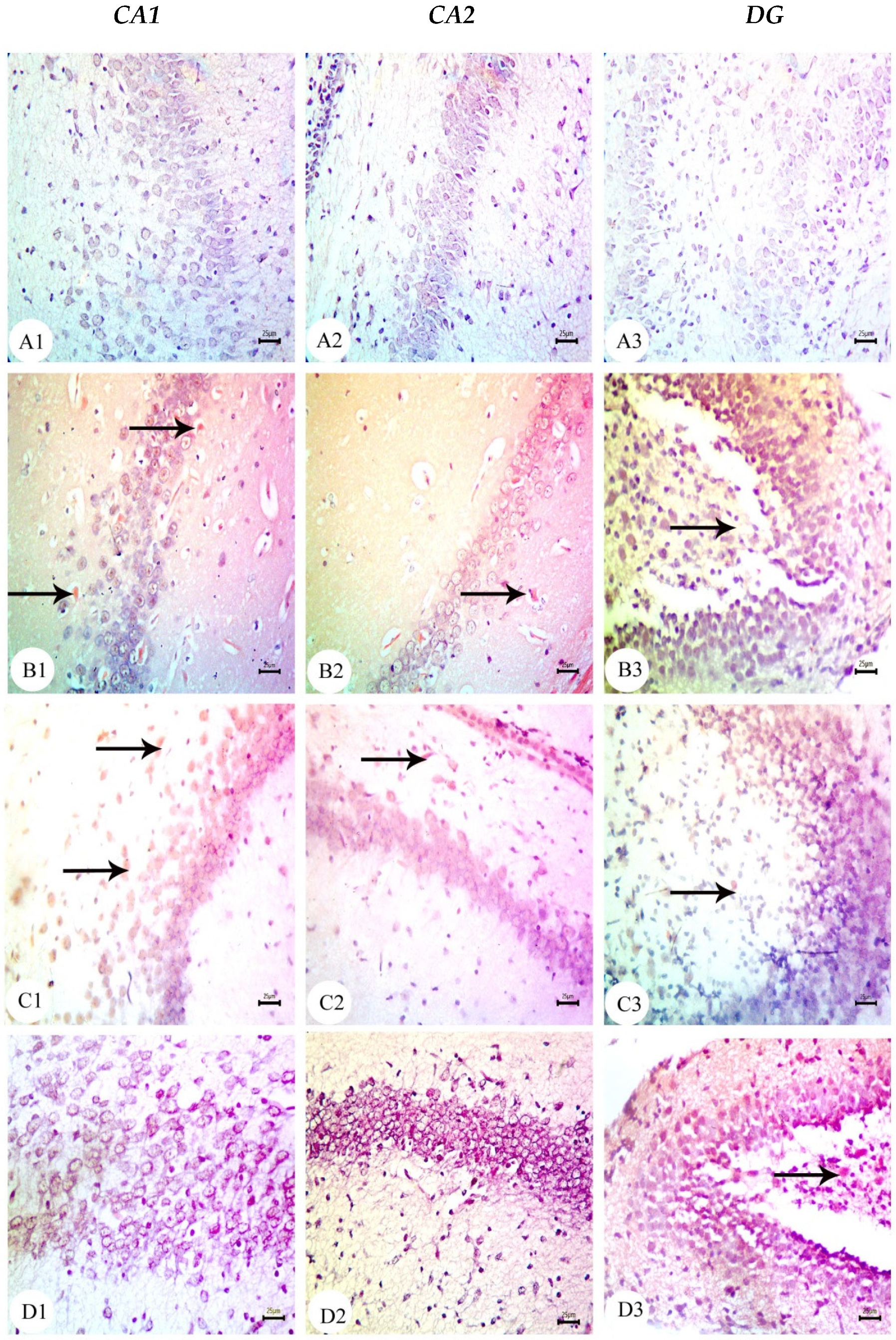
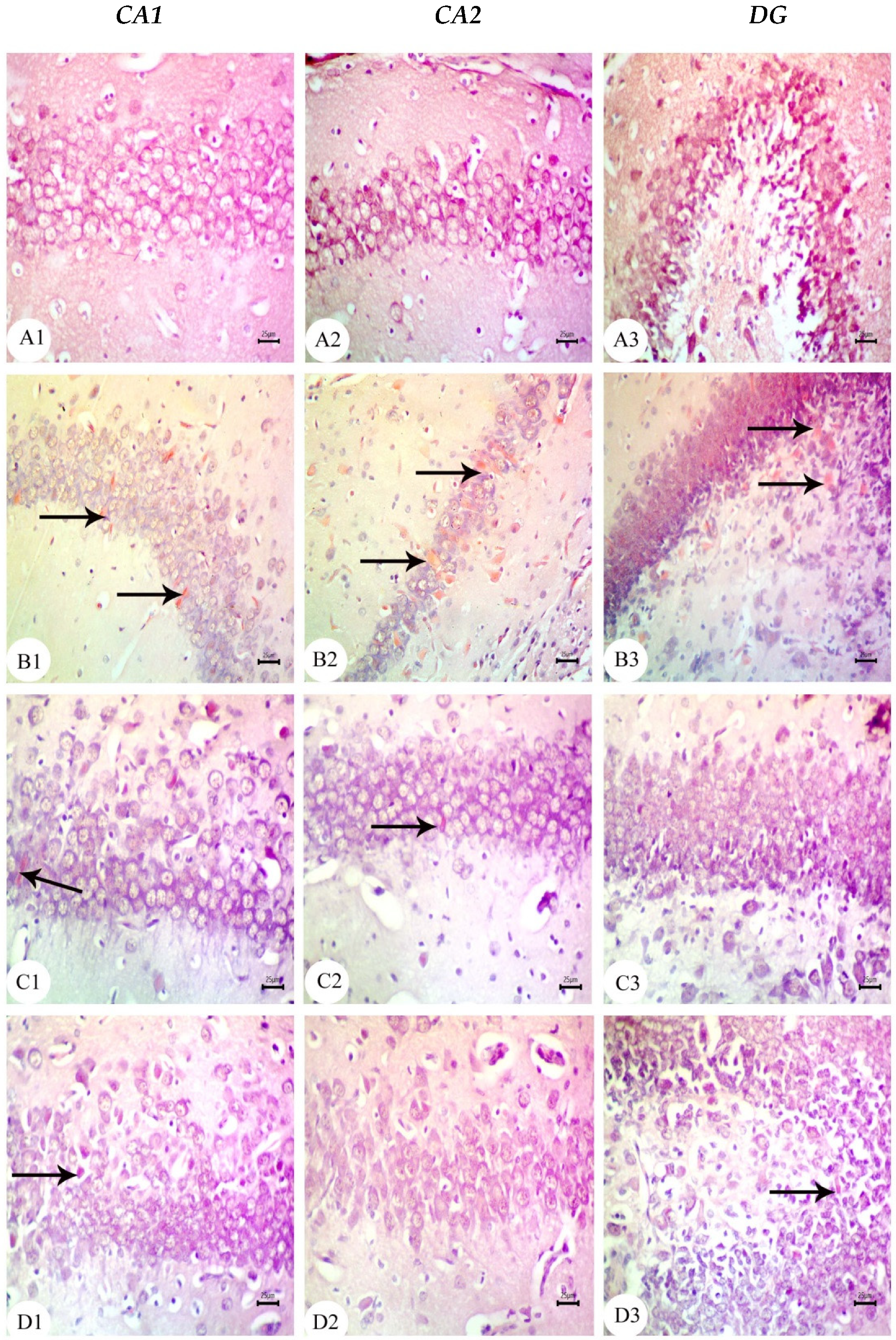
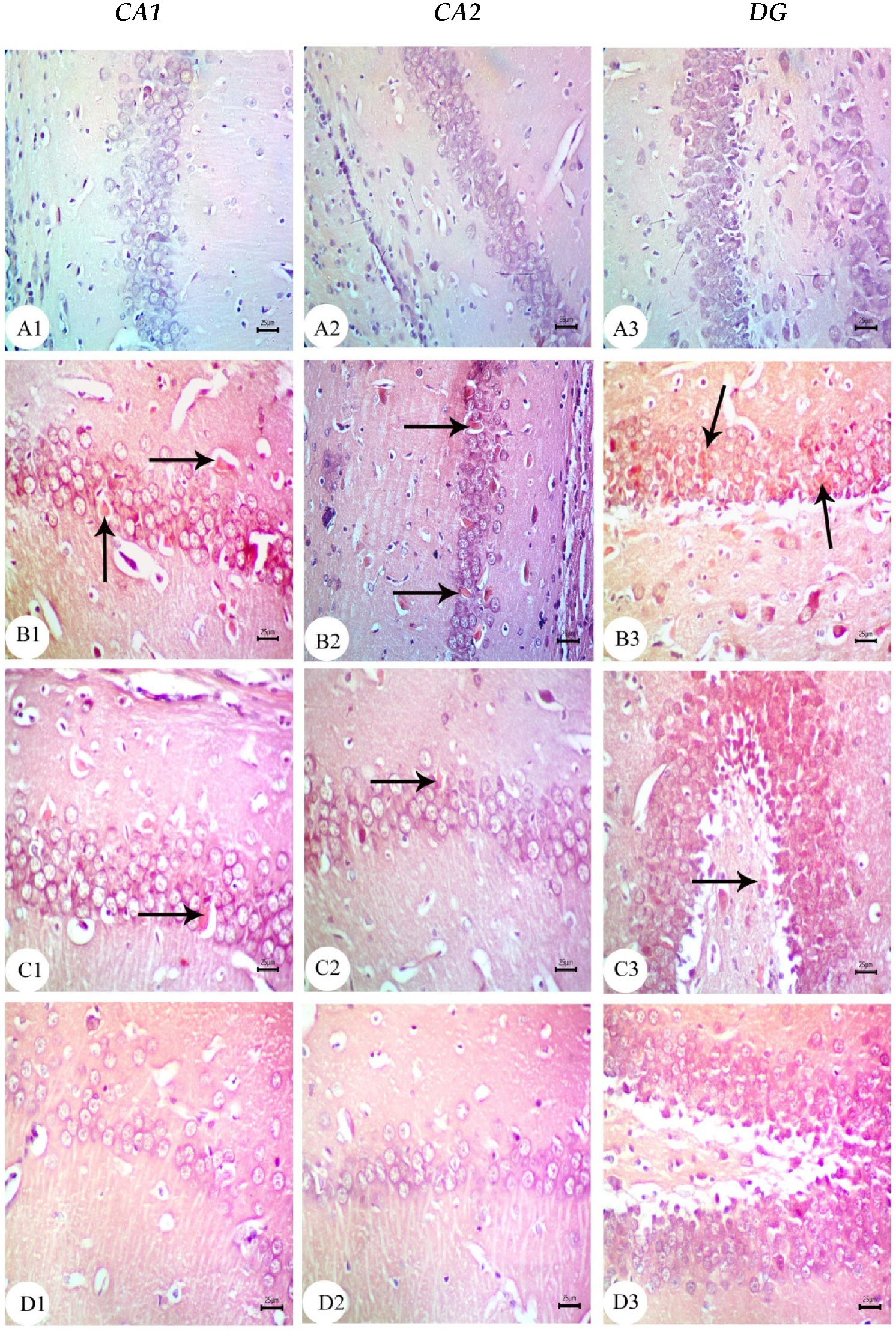
3.3. Amyloid Beta Deposition Confirmed by Congo Red Staining: Figure 6, Figure 7, Figure 8, Figure 9, Figure 10 and Figure 11






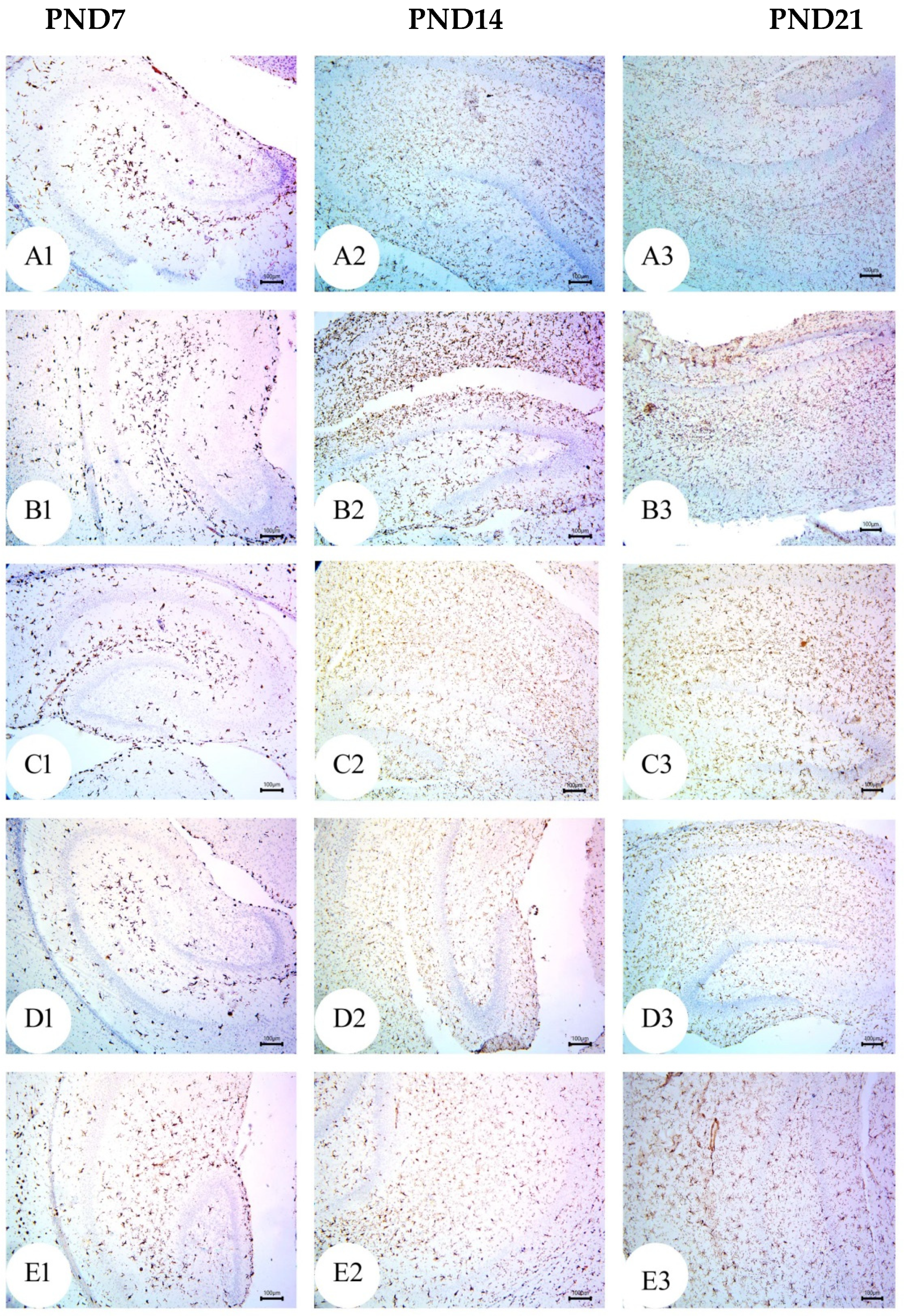
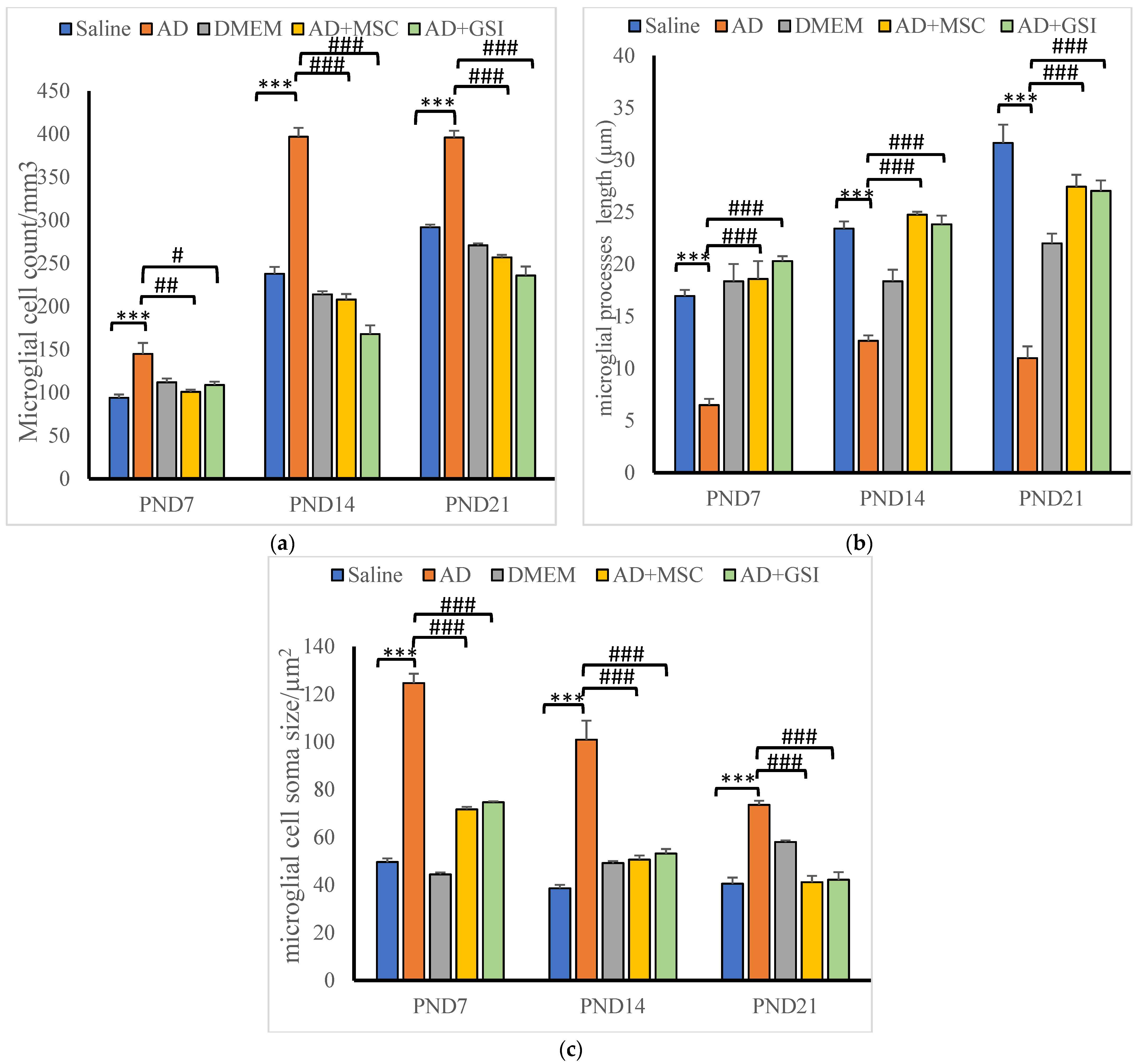
3.4. BM-MSC and GSI-953 Treatments Restored near Normal Microglial Cell Counts, Soma Size, and Dendrite Length in the Hippocampus of Offspring from Aβ 25–35-Injected Dams
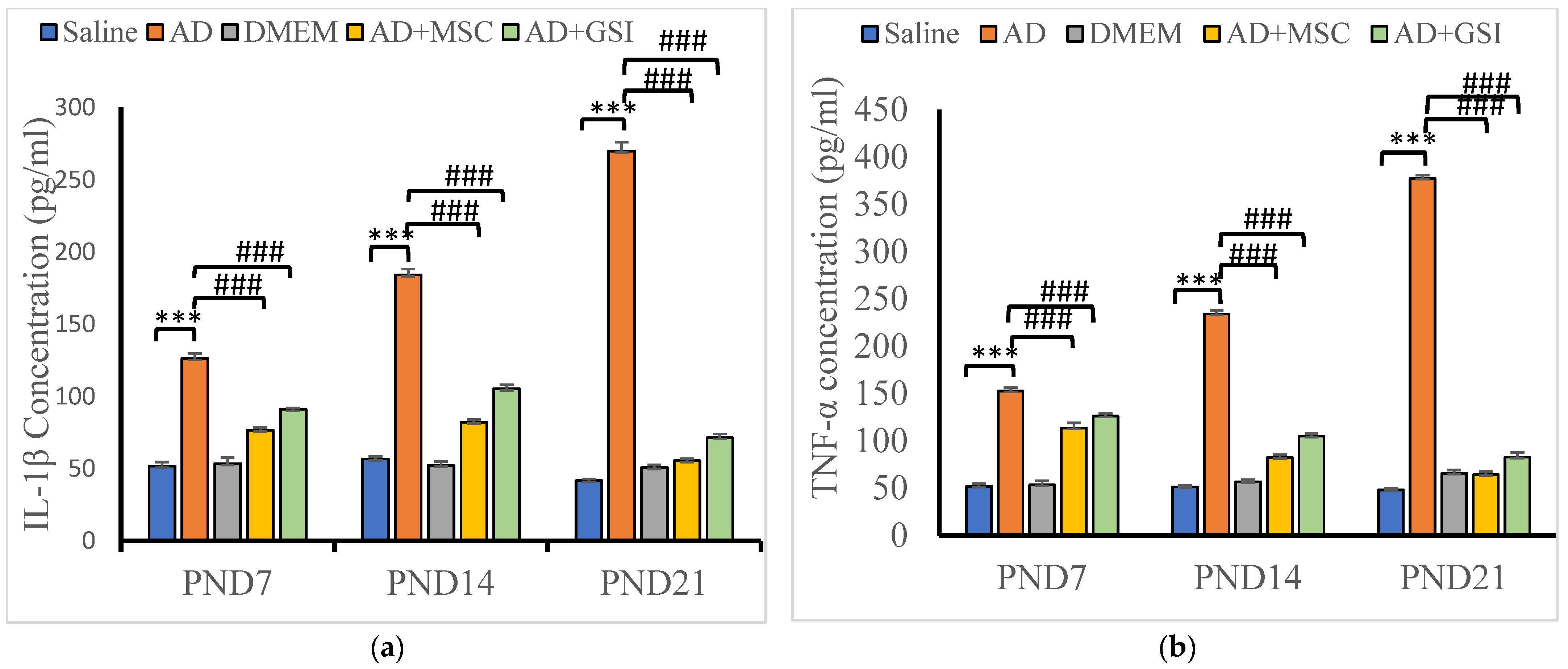
3.5. BM-MSC and GSI-953 Treatments Restored Normal Serum Levels of Proinflammatory, Prodegenerative, and Neuroprotective Factors in Offspring of Aβ 25–35-Injected Dams
3.5.1. Effects on Neuroinflammatory Cytokine Serum Levels
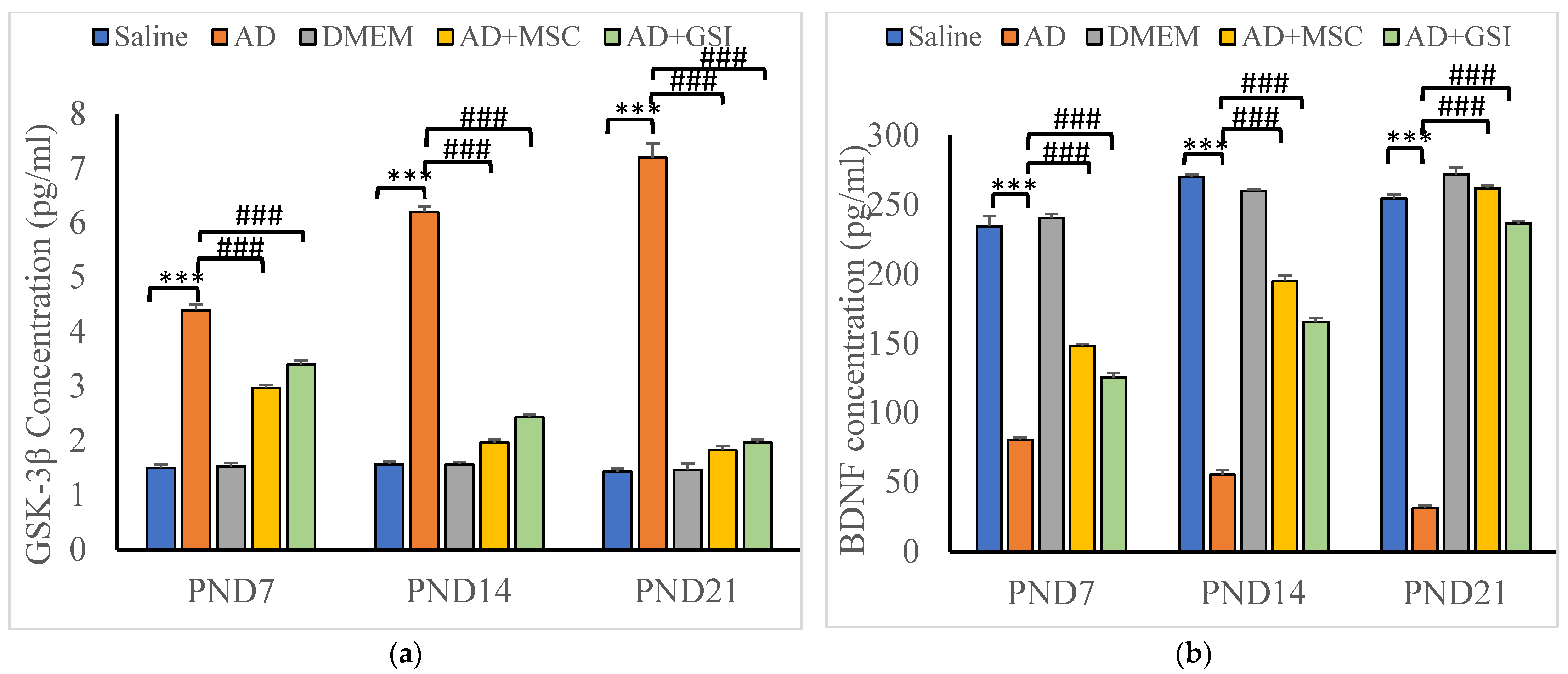
3.5.2. Effects on Glycogen Synthase Kinase-3β (GSK-3β) and BDNF Serum Levels
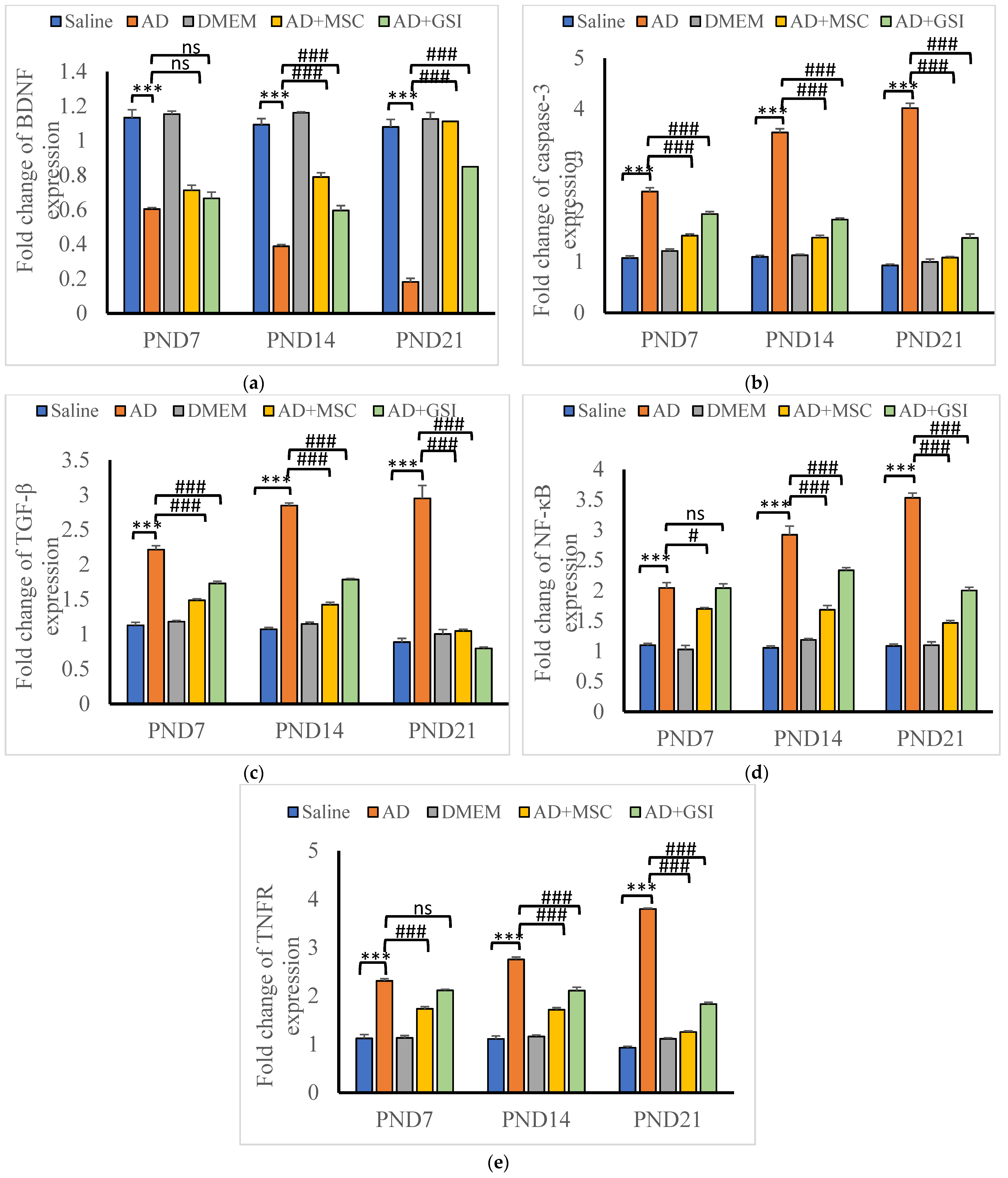
3.6. BM-MSC and GSI-953 Treatments Restore Normal Expression Levels of Neuroprotective and Proapoptotic Genes in the Hippocampus of Offspring from Aβ 25–35-Injected Dams
3.6.1. Effects on BDNF, Caspase-3, TGFβ, NF-κB, and TNFR Gene Expression in the Neonatal Hippocampus
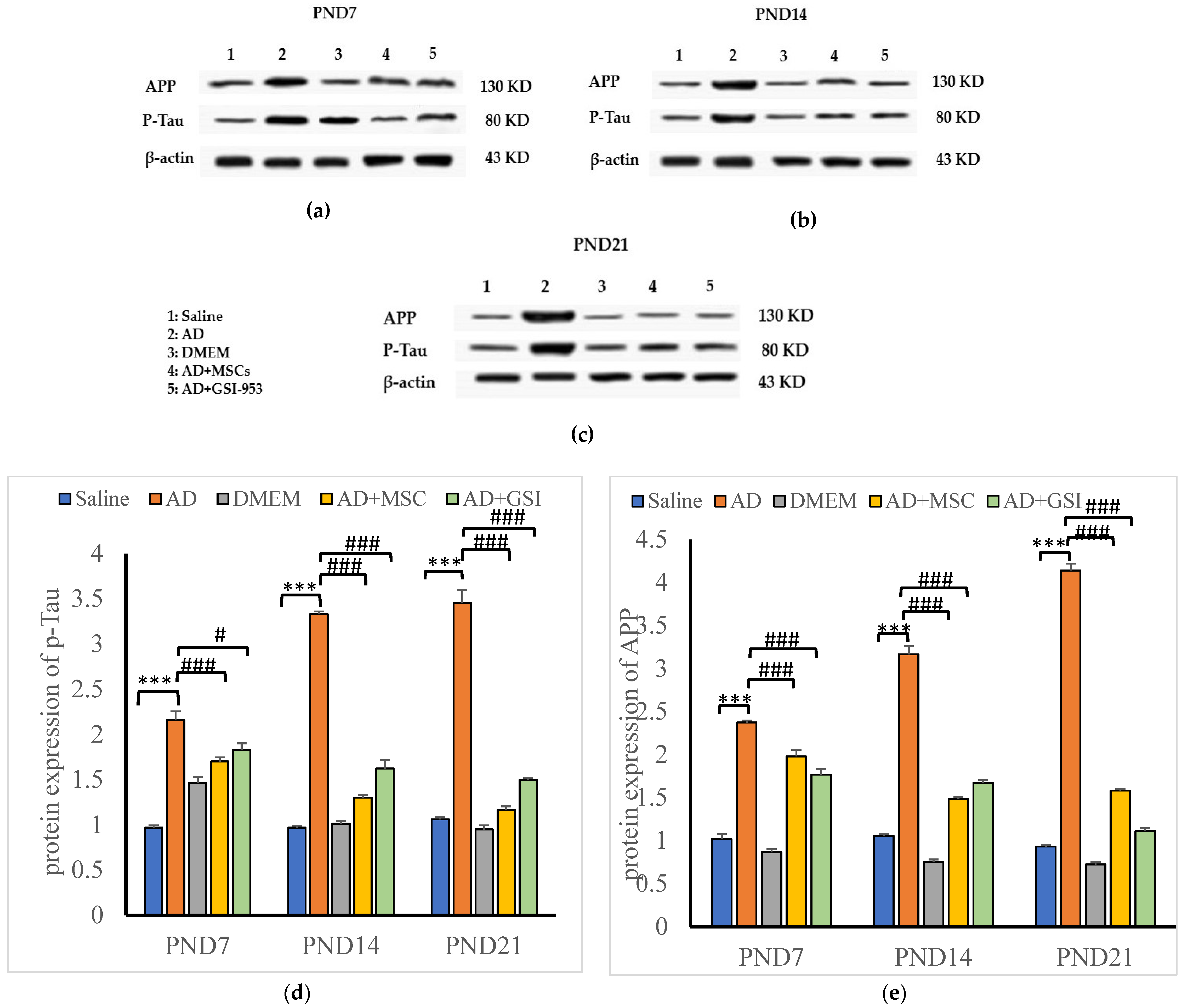
3.6.2. Effects on Phosphorylated Tau and APP Protein Levels in Neonatal Hippocampus
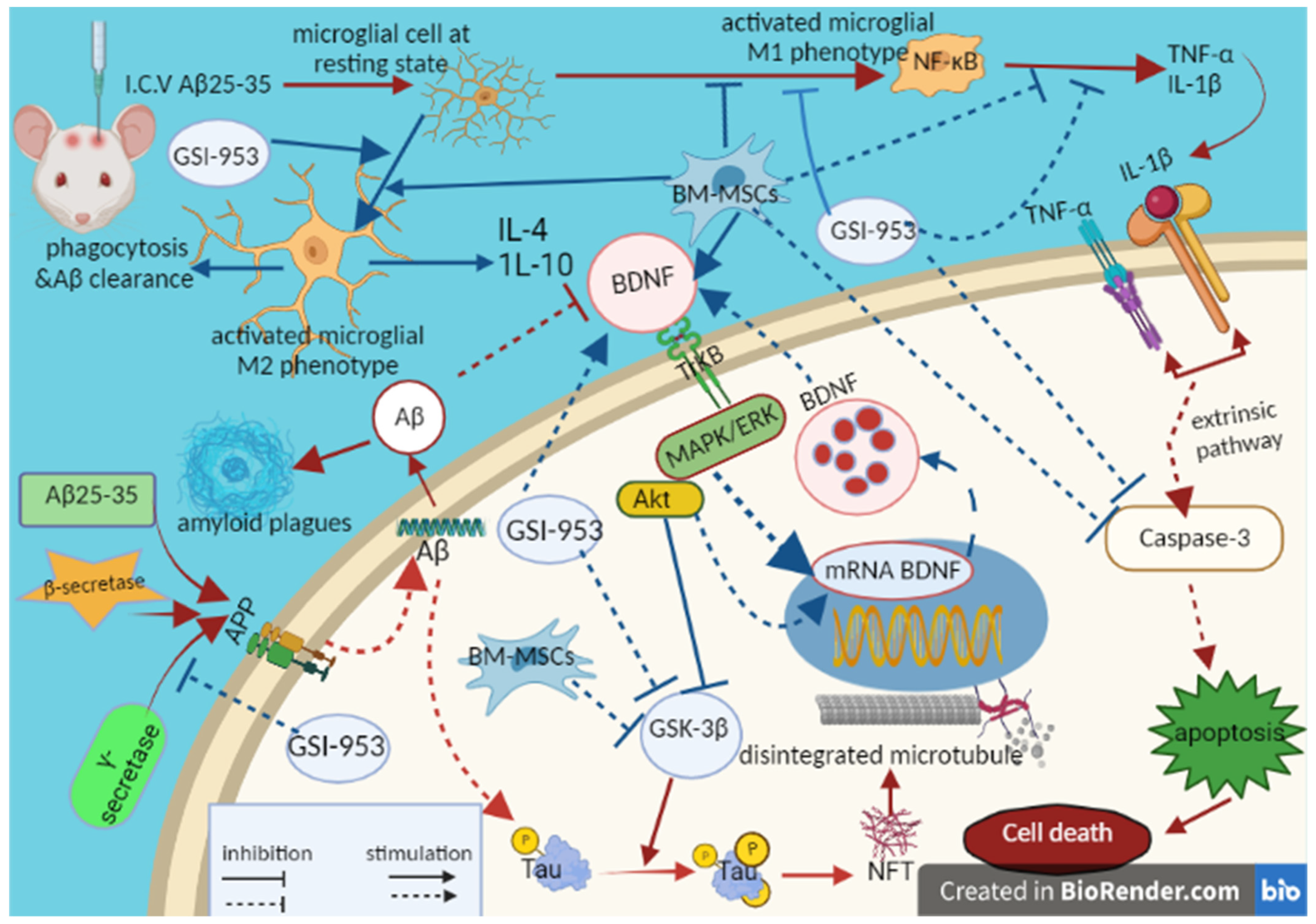
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neves, A.F.; Camargo, C.; Premer, C.; Hare, J.M.; Baumel, B.S.; Pinto, M. Intravenous administration of mesenchymal stem cells reduces tau phosphorylation and inflammation in the 3xtg-ad mouse model of alzheimer’s disease. Exp. Neurol. 2021, 341, 113706. [Google Scholar] [CrossRef] [PubMed]
- Brookmeyer, R.; Johnson, E.; Ziegler-Graham, K.; Arrighi, H.M. Forecasting the global burden of alzheimer’s disease. Alzheimer’s Dement. 2007, 3, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzheimer’s Association. Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2019, 15, 321–387. [Google Scholar] [CrossRef]
- Young, J.E.; Goldstein, L.S. Alzheimer’s disease in a dish: Promises and challenges of human stem cell models. Hum. Mol. Genet. 2012, 21, R82–R89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler-Graham, K.; Brookmeyer, R.; Johnson, E.; Arrighi, H.M. Worldwide variation in the doubling time of alzheimer’s disease incidence rates. Alzheimer’s Dement. 2008, 4, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Chakari-Khiavi, F.; Dolati, S.; Chakari-Khiavi, A.; Abbaszadeh, H.; Aghebati-Maleki, L.; Pourlak, T.; Mehdizadeh, A.; Yousefi, M. Prospects for the application of mesenchymal stem cells in alzheimer’s disease treatment. Life Sci. 2019, 231, 116564. [Google Scholar] [CrossRef]
- Liu, X.Y.; Yang, L.P.; Zhao, L. Stem cell therapy for Alzheimer’s disease. World J. Stem Cells 2020, 12, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Kent, S.A.; Spires-Jones, T.L.; Durrant, C.S. The physiological roles of tau and aβ: Implications for alzheimer’s disease pathology and therapeutics. Acta Neuropathol. 2020, 140, 417–447. [Google Scholar] [CrossRef]
- Scearce-Levie, K.; Sanchez, P.E.; Lewcock, J.W. Leveraging preclinical models for the development of alzheimer disease therapeutics. Nat. Rev. Drug Disc. 2020, 19, 447–462. [Google Scholar] [CrossRef]
- Park, H.S.; Pang, Q.Q.; Kim, Y.S.; Kim, J.H.; Cho, E.J. Neuroprotective effect of membrane-free stem cell extract against amyloid beta 25–35-induced neurotoxicity in sh-sy5y cells. Appl. Sci. 2021, 11, 2219. [Google Scholar] [CrossRef]
- Cai, Z.; Zhao, B.; Ratka, A. Oxidative stress and β-amyloid protein in alzheimer’s disease. Neuromol. Med. 2011, 13, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Stakos, D.A.; Stamatelopoulos, K.; Bampatsias, D.; Sachse, M.; Zormpas, E.; Vlachogiannis, N.I.; Tual-Chalot, S.; Stellos, K. The Alzheimer’s disease amyloid-beta hypothesis in cardiovascular aging and disease: JACC focus seminar. J. Am. Coll. Cardiol. 2020, 75, 952–967. [Google Scholar] [CrossRef] [PubMed]
- Si, Z.; Wang, X. Stem cell therapies in Alzheimer’s disease: Applications for disease modeling. J. Pharmacol. Exp. Ther. 2021, 377, 207–217. [Google Scholar] [CrossRef]
- Wei, W.; Wang, Y.; Liu, Y.; Dai, C.-L.; Tung, Y.-C.; Liu, F.; Iqbal, K. Prenatal to early postnatal neurotrophic treatment prevents alzheimer-like behavior and pathology in mice. Alzheimer’s Res. Ther. 2020, 12, 102. [Google Scholar] [CrossRef]
- Querfurth, H.W.; LaFerla, F.M. Mechanisms of disease. N. Eng. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Dash, P.K.; Moore, A.N.; Orsi, S.A. Blockade of γ-secretase activity within the hippocampus enhances long-term memory. Biochem. Biophys. Res. Commun. 2005, 338, 777–782. [Google Scholar] [CrossRef]
- Wen, J.; Liu, D.; Zhao, L. Small molecules targeting γ-secretase and their potential biological applications. Eur. J. Med. Chem. 2022, 232, 114169. [Google Scholar] [CrossRef] [PubMed]
- Martone, R.L.; Zhou, H.; Atchison, K.; Comery, T.; Xu, J.Z.; Huang, X.; Gong, X.; Jin, M.; Kreft, A.; Harrison, B. Begacestat (gsi-953): A novel, selective thiophene sulfonamide inhibitor of amyloid precursor protein γ-secretase for the treatment of alzheimer’s disease. J. Pharmacol. Exp. Ther. 2009, 331, 598–608. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, R.; Van Den Daele, J.; Fattorelli, N.; Wolfs, L.; Balusu, S.; Burton, O.; Liston, A.; Sierksma, A.; Fourne, Y.; Poovathingal, S. Stem-cell-derived human microglia transplanted in mouse brain to study human disease. Nat. Neurosc. 2019, 22, 2111–2116. [Google Scholar] [CrossRef]
- Lee, C.; Willerth, S.M.; Nygaard, H.B. The use of patient-derived induced pluripotent stem cells for alzheimer’s disease modeling. Prog. Neurobiol. 2020, 192, 101804. [Google Scholar] [CrossRef]
- Xu, X.; Gu, D.; Xu, B.; Yang, C.; Wang, L. Circular rna circ_0005835 promotes promoted neural stem cells proliferation and differentiate to neuron and inhibits inflammatory cytokines levels through mir-576-3p in alzheimer’s disease. Environ. Sci. Pollut. Res. 2022, 29, 35934–35943. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Lu, P. Introduction for stem cell–based therapy for neurodegenerative diseases. In Stem Cell-Based Therapy for Neurodegenerative Diseases; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–8. [Google Scholar]
- Feng, Z.; Gao, F. Stem cell challenges in the treatment of neurodegenerative disease. CNS Neurosci. Ther. 2012, 18, 142–148. [Google Scholar] [CrossRef]
- Yang, H.; Xie, Z.H.; Wei, L.F.; Yang, H.N.; Yang, S.N.; Zhu, Z.Y.; Wang, P.; Zhao, C.P.; Bi, J.Z. Human umbilical cord mesenchymal stem cell-derived neuron-like cells rescue memory deficits and reduce amyloid-beta deposition in an aβpp/ps1 transgenic mouse model. Stem Cell Res. Ther. 2013, 4, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penney, J.; Ralvenius, W.T.; Tsai, L.-H. Modeling alzheimer’s disease with ipsc-derived brain cells. Mol. Psychiatry 2020, 25, 148–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodbury, D.; Reynolds, K.; Black, I.B. Adult bone marrow stromal stem cells express germline, ectodermal, endodermal, and mesodermal genes prior to neurogenesis. J. Neurosci. Res. 2002, 69, 908–917. [Google Scholar] [CrossRef]
- Qin, C.; Li, Y.; Wang, K. Functional mechanism of bone marrow-derived mesenchymal stem cells in the treatment of animal models with alzheimer’s disease: Inhibition of neuroinflammation. J. Inflamm. Res. 2021, 14, 4761. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.J.; Xiong, C.; Benzinger, T.L.; Fagan, A.M.; Goate, A.; Fox, N.C.; Marcus, D.S.; Cairns, N.J.; Xie, X.; Blazey, T.M. Clinical and biomarker changes in dominantly inherited alzheimer’s disease. N. Engl. J. Med. 2012, 367, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Lee, D.K.; Chung, B.-R.; Kim, H.V.; Kim, Y. Intracerebroventricular injection of amyloid-β peptides in normal mice to acutely induce alzheimer-like cognitive deficits. J. Vis. Exp. 2016, 109, e53308. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, R.H.; Galaly, S.R.; Moustafa, N.; Ahmed, R.R.; Ali, T.M.; Elesawy, B.H.; Ahmed, O.M.; Abdul-Hamid, M. Curcumin and mesenchymal stem cells ameliorate ankle, testis, and ovary deleterious histological changes in arthritic rats via suppression of oxidative stress and inflammation. Stem Cells Int. 2021, 2021, 3516834. [Google Scholar] [CrossRef]
- Ahmed, O.M.; Saleh, A.S.; Ahmed, E.A.; Ghoneim, M.M.; Ebrahim, H.A.; Abdelgawad, M.A.; Abdel-Gabbar, M. Efficiency of Bone Marrow-Derived Mesenchymal Stem Cells and Hesperetin in the Treatment of Streptozotocin-Induced Type 1 Diabetes in Wistar Rats. Pharmaceuticals 2023, 16, 859. [Google Scholar] [CrossRef]
- Gaber, A.; Elbakry, A.M.; Aljarari, R.M.; Jaber, F.A.; Khadrawy, Y.A.; Sabry, D.; Abo-ELeneen, R.E.; Ahmed, O.M. Bone Marrow-Derived Mesenchymal Stem Cells and γ-Secretase Inhibitor Treatments Suppress Amyloid-β25–35-Induced Cognitive Impairment in Rat Dams and Cortical Degeneration in Offspring. Stem Cells Int. 2023, 2023, 2690949. [Google Scholar] [CrossRef]
- Sayed, H.M.; Awaad, A.S.; Abdel Rahman, F.E.Z.S.; Al-Dossari, M.; El-Gawaad, A.; Ahmed, O.M. Combinatory Effect and Modes of Action of Chrysin and Bone Marrow-Derived Mesenchymal Stem Cells on Streptozotocin/Nicotinamide-Induced Diabetic Rats. Pharmaceuticals 2023, 16, 34. [Google Scholar] [CrossRef]
- Sumathi, T.; Asha, D.; Nagarajan, G.; Sreenivas, A.; Nivedha, R. L-theanine alleviates the neuropathological changes induced by pcb (aroclor 1254) via inhibiting upregulation of inflammatory cytokines and oxidative stress in rat brain. Environ. Toxicol. Pharmacol. 2016, 42, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Nalivaeva, N.N.; Belyaev, N.D.; Lewis, D.I.; Pickles, A.R.; Makova, N.Z.; Bagrova, D.I.; Dubrovskaya, N.M.; Plesneva, S.A.; Zhuravin, I.A.; Turner, A.J. Effect of sodium valproate administration on brain neprilysin expression and memory in rats. J. Mol. Neurosci. 2012, 46, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Botton, P.H.; Costa, M.S.; Ardais, A.P.; Mioranzza, S.; Souza, D.O.; da Rocha, J.B.T.; Porciúncula, L.O. Caffeine prevents disruption of memory consolidation in the inhibitory avoidance and novel object recognition tasks by scopolamine in adult mice. Behav. Brain Res. 2010, 214, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Hammond, R.S.; Tull, L.E.; Stackman, R.W. On the delay-dependent involvement of the hippocampus in object recognition memory. Neurobiol. Learn. Mem. 2004, 82, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Iulita, M.F.; Allard, S.; Richter, L.; Munter, L.-M.; Ducatenzeiler, A.; Weise, C.; Carmo, S.D.; Klein, W.L.; Multhaup, G.; Cuello, A.C. Intracellular aβ pathology and early cognitive impairments in a transgenic rat overexpressing human amyloid precursor protein: A multidimensional study. Acta Neuropathol. Commun. 2014, 2, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amirazodi, F.; Mehrabi, A.; Amirazodi, M.; Parsania, S.; Rajizadeh, M.A.; Esmaeilpour, K. The combination effects of resveratrol and swimming HIIT exercise on novel object recognition and open-field tasks in aged rats. Exp. Aging Res. 2020, 46, 336–358. [Google Scholar] [CrossRef]
- Bockhorst, K.; Narayana, P.; Liu, R.; Ahobila-Vijjula, P.; Ramu, J.; Kamel, M.; Wosik, J.; Bockhorst, T.; Hahn, K.; Hasan, K. Early postnatal development of rat brain: In vivo diffusion tensor imaging. J. Neurosci. Res. 2008, 86, 1520–1528. [Google Scholar] [CrossRef] [Green Version]
- Jahn, H. Memory loss in alzheimer’s disease. Dialogues Clin. Neurosci. 2022, 15, 445–454. [Google Scholar] [CrossRef]
- Cao, B.; Zeng, M.; Zhang, Q.; Zhang, B.; Cao, Y.; Wu, Y.; Feng, W.; Zheng, X. Amentoflavone ameliorates memory deficits and abnormal autophagy in aβ25−35-induced mice by mtor signaling. Neurochem. Res. 2021, 46, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Zussy, C.; Brureau, A.; Keller, E.; Marchal, S.; Blayo, C.; Delair, B.; Ixart, G.; Maurice, T.; Givalois, L. Alzheimer’s disease related markers, cellular toxicity and behavioral deficits induced six weeks after oligomeric amyloid-β peptide injection in rats. PLoS ONE 2013, 8, e53117. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lu, F.; Jia, X.; Yan, Q.; Zhang, X.; Mu, P. Amyloid-β (25–35) regulates neuronal damage and memory loss via sirt1/nrf2 in the cortex of mice. J. Chem. Neuroanat. 2021, 114, 101945. [Google Scholar] [CrossRef] [PubMed]
- Kälin, A.M.; Park, M.T.; Chakravarty, M.M.; Lerch, J.P.; Michels, L.; Schroeder, C.; Broicher, S.D.; Kollias, S.; Nitsch, R.M.; Gietl, A.F. Subcortical shape changes, hippocampal atrophy and cortical thinning in future alzheimer’s disease patients. Front. Aging Neurosci. 2017, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Yang, X.; Han, Z.-P.; Qu, F.-F.; Shao, L.; Shi, Y.-F. Mesenchymal stem cells: A new trend for cell therapy. Acta Pharmacol. Sin. 2013, 34, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Glenn, J.D.; Whartenby, K.A. Mesenchymal stem cells: Emerging mechanisms of immunomodulation and therapy. World J. Stem Cells 2014, 6, 526. [Google Scholar] [CrossRef] [PubMed]
- Mandal, B.; Niyogi, S.; Sarkar, K.; Das, R. Structure based virtual screening of rivastigmine derivatives as cholinesterase inhibitors. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Schlachetzki, J.; Hull, M. Microglial activation in alzheimer’s disease. Curr. Alzheimer Res. 2009, 6, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Beja-Glasser, V.F.; Nfonoyim, B.M.; Frouin, A.; Li, S.; Ramakrishnan, S.; Merry, K.M.; Shi, Q.; Rosenthal, A.; Barres, B.A. Complement and microglia mediate early synapse loss in alzheimer mouse models. Science 2016, 352, 712–716. [Google Scholar] [CrossRef] [Green Version]
- Harach, T.; Jammes, F.; Muller, C.; Duthilleul, N.; Cheatham, V.; Zufferey, V.; Cheatham, D.; Lukasheva, Y.A.; Lasser, T.; Bolmont, T. Administrations of human adult ischemia-tolerant mesenchymal stem cells and factors reduce amyloid beta pathology in a mouse model of alzheimer’s disease. Neurobiol. Aging 2017, 51, 83–96. [Google Scholar] [CrossRef]
- Kim, K.-S.; Kim, H.S.; Park, J.-M.; Kim, H.W.; Park, M.-K.; Lee, H.-S.; Lim, D.S.; Lee, T.H.; Chopp, M.; Moon, J. Long-term immunomodulatory effect of amniotic stem cells in an alzheimer’s disease model. Neurobiol. Aging 2013, 34, 2408–2420. [Google Scholar] [CrossRef] [PubMed]
- Lykhmus, O.; Koval, L.; Voytenko, L.; Uspenska, K.; Komisarenko, S.; Deryabina, O.; Shuvalova, N.; Kordium, V.; Ustymenko, A.; Kyryk, V. Intravenously injected mesenchymal stem cells penetrate the brain and treat inflammation-induced brain damage and memory impairment in mice. Front. Pharmacol. 2019, 10, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchenko, M.; Poltavtseva, R.; Bobkova, N.; Vel’meshev, D.; Nesterova, I.; Samokhin, A.; Sukhikh, G. Localization and differentiation pattern of transplanted human multipotent mesenchymal stromal cells in the brain of bulbectomized mice. Bull. Exp. Biol. Med. 2014, 158, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Mayer, S.C.; Kreft, A.F.; Harrison, B.; Abou-Gharbia, M.; Antane, M.; Aschmies, S.; Atchison, K.; Chlenov, M.; Cole, D.C.; Comery, T.; et al. Discovery of begacestat, a Notch-1-sparing g-secretase inhibitor for the treatment of Alzheimer’s disease. J. Med. Chem. 2008, 51, 7348–7351. [Google Scholar] [CrossRef]
- Pooler, A.M.; Noble, W.; Hanger, D.P. A role for tau at the synapse in alzheimer’s disease pathogenesis. Neuropharmacology 2014, 76, 1–8. [Google Scholar] [CrossRef]
- Petry, F.d.S.; Coelho, B.P.; Gaelzer, M.M.; Kreutz, F.; Guma, F.T.C.R.; Salbego, C.G.; Trindade, V.M.T. Genistein protects against amyloid-beta-induced toxicity in sh-sy5y cells by regulation of akt and tau phosphorylation. Phytother. Res. 2020, 34, 796–807. [Google Scholar] [CrossRef]
- Hoppe, J.B.; Coradini, K.; Frozza, R.L.; Oliveira, C.M.; Meneghetti, A.B.; Bernardi, A.; Pires, E.S.; Beck, R.C.; Salbego, C.G. Free and nanoencapsulated curcumin suppress β-amyloid-induced cognitive impairments in rats: Involvement of bdnf and akt/gsk-3β signaling pathway. Neurobiol. Learn. Mem. 2013, 106, 134–144. [Google Scholar] [CrossRef]
- dos Santos Petry, F.; Hoppe, J.B.; Klein, C.P.; Santos, B.G.D.; Hözer, R.M.; Bifi, F.; Matté, C.; Salbego, C.G.; Trindade, V.M.T. Genistein attenuates amyloid-beta-induced cognitive impairment in rats by modulation of hippocampal synaptotoxicity and hyperphosphorylation of tau. J. Nutr. Biochem. 2021, 87, 108525. [Google Scholar] [CrossRef]
- Naseri, N.N.; Wang, H.; Guo, J.; Sharma, M.; Luo, W. The complexity of tau in alzheimer’s disease. Neurosci. Lett. 2019, 705, 183–194. [Google Scholar] [CrossRef]
- Khan, S.S.; Bloom, G.S. Tau: The center of a signaling nexus in alzheimer’s disease. Front. Neurosci. 2016, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Coelho, B.P.; Gaelzer, M.M.; Petry, F.D.S.; Hoppe, J.B.; Trindade, V.M.T.; Salbego, C.G.; Guma, F.T. Dual effect of doxazosin: Anticancer activity on sh-sy5y neuroblastoma cells and neuroprotection on an in vitro model of alzheimer’s disease. Neuroscience 2019, 404, 314–325. [Google Scholar] [CrossRef]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A simple role for bdnf in learning and memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejeda, G.S.; Díaz-Guerra, M. Integral characterization of defective bdnf/trkb signalling in neurological and psychiatric disorders leads the way to new therapies. Int. J. Mol. Sci. 2017, 18, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-H.; Ko, Y.J.; Kim, J.-Y.; Sim, Y.-J. Treadmill running improves spatial learning memory through inactivation of nuclear factor kappa b/mitogen-activated protein kinase signaling pathway in amyloid-β-induced alzheimer disease rats. Int. Neur. J. 2021, 25, S35. [Google Scholar] [CrossRef]
- Yoon, Y.-S.; Wecker, A.; Heyd, L.; Park, J.-S.; Tkebuchava, T.; Kusano, K.; Hanley, A.; Scadova, H.; Qin, G.; Cha, D.H.; et al. Clonally expanded novel multipotent stem cells from human bone marrow regenerate myocardium after myocardial infarction. J. Clin. Investig. 2005, 115, 326–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Kim, M.-J.; Kim, Y.J.; Lee, Y.-H.; Bae, D.; Kim, S.; Na, Y.; Yoon, H.-G. Selective pcaf inhibitor ameliorates cognitive and behavioral deficits by suppressing nf-κb-mediated neuroinflammation induced by aβ in a model of alzheimer’s disease. Int. J. Mol. Med. 2015, 35, 1109–1118. [Google Scholar] [CrossRef] [Green Version]
- Ling, I.; Golde, T.E.; Galasko, D.R.; Koo, E.H. Modulation of aβ42 in vivo by γ-secretase modulator in primates and humans. Alzheimer’s Res. Ther. 2015, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-H.; Chung, K.-S.; Jin, B.-R.; Cheon, S.-Y.; Nugroho, A.; Roh, S.-S.; An, H.-J. Anti-inflammatory effects of an ethanol extract of aster glehni via inhibition of nf-κb activation in mice with dss-induced colitis. Food Funct. 2017, 8, 2611–2620. [Google Scholar] [CrossRef]
- Qin, C.; Bai, L.; Li, Y.; Wang, K. The functional mechanism of bone marrow-derived mesenchymal stem cells in the treatment of animal models with alzheimer’s disease: Crosstalk between autophagy and apoptosis. Stem Cell Res. Ther. 2022, 13, 1–13. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, J.; Liu, C.; Song, C.; Li, P.; Yin, F.; Xiao, Y.; Jiang, W.; Zong, A.; Zhang, X. Protective effects of low molecular weight chondroitin sulfate on amyloid beta (aβ)-induced damage in vitro and in vivo. Neuroscience 2015, 305, 169–182. [Google Scholar] [CrossRef]
- Wang, S.-P.; Wang, Z.-H.; Peng, D.-Y.; Li, S.-M.; Wang, H.; Wang, X.-H. Therapeutic effect of mesenchymal stem cells in rats with intracerebral hemorrhage: Reduced apoptosis and enhanced neuroprotection. Mol. Med. Rep. 2012, 6, 848–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Fukuda, S.; Liu, L.; Chitteti, B.R.; Pelus, L.M. Survivin is required for mouse and human bone marrow mesenchymal stromal cell function. Stem Cells 2018, 36, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamm, I.; Wang, Y.; Sausville, E.; Scudiero, D.A.; Vigna, N.; Oltersdorf, T.; Reed, J.C. Iap-family protein survivin inhibits caspase activity and apoptosis induced by fas (cd95), bax, caspases, and anticancer drugs. Cancer Res. 1998, 58, 5315–5320. [Google Scholar] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Purpose of Use | Number of Animals |
|---|---|---|
| Female | Saline-injected group | 10 |
| AD group | 10 | |
| AD + MSCs | 10 | |
| AD + GSI-953 | 10 | |
| DMEM group | 10 | |
| Male | Mating with female at arrange of two female with one male | 30 adult male rats |
| Forward Sequence | Reverse Sequence | |
|---|---|---|
| Caspase-3 | 5′-TGGTTCATCCAGTCGCTTTGT-3′ | 5′-CAAATTCTGTTGCCACCTTTCG-3′ |
| TNFR | 5′-GGGATTCAGCTCCTGTCAAA-3′ | 5′-ATGAACTCCTTCCAGCGTGT-3′ |
| TGF-β | 5′-GTCACTGGAGTTGTACGGCA-3′ | 5′-GGGCTGATCCCGTTGATTTC-3′ |
| BDNF | 5′-CCGGTATCCAAAGGCCAACT-3′ | 5′-CTGCAGCCTTCCTTGGTGTA-3′ |
| NF-κB | 5′-TTCCCT GAA GTG GAG CTA GGA-3′ | 5′-CATGTC GAG GAA GAC ACT GGA-3′ |
| β-actin | 5′-AGGCCC CTC TGA ACC CTA AG-3′ | 5′-GGA GCG CGT AAC CCT CATAG-3′ |
| Group | Novel Object Recognition Test | Y-Maze Test | |
|---|---|---|---|
| (Discrimination Index) | Time Percent for Novel Arm | Percent of Novel Arm Entries | |
| Saline | 0.38450 ± 0.139316 | 80.4667 ± 1.02783 | 44.5833 ± 2.451 |
| AD | −0.53200 ± 0.093088 * | 41.8667 ± 6.95608 * | 44.0500 ± 4.312 ns |
| F-value | 29.919 | 30.135 | 0.012 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaber, A.; Ahmed, O.M.; Khadrawy, Y.A.; Zoheir, K.M.A.; Abo-ELeneen, R.E.; Alblihed, M.A.; Elbakry, A.M. Mesenchymal Stem Cells and Begacestat Mitigate Amyloid-β 25–35-Induced Cognitive Decline in Rat Dams and Hippocampal Deteriorations in Offspring. Biology 2023, 12, 905. https://doi.org/10.3390/biology12070905
Gaber A, Ahmed OM, Khadrawy YA, Zoheir KMA, Abo-ELeneen RE, Alblihed MA, Elbakry AM. Mesenchymal Stem Cells and Begacestat Mitigate Amyloid-β 25–35-Induced Cognitive Decline in Rat Dams and Hippocampal Deteriorations in Offspring. Biology. 2023; 12(7):905. https://doi.org/10.3390/biology12070905
Chicago/Turabian StyleGaber, Asmaa, Osama M. Ahmed, Yasser A. Khadrawy, Khairy M. A. Zoheir, Rasha E. Abo-ELeneen, Mohamed A. Alblihed, and Ahlam M. Elbakry. 2023. "Mesenchymal Stem Cells and Begacestat Mitigate Amyloid-β 25–35-Induced Cognitive Decline in Rat Dams and Hippocampal Deteriorations in Offspring" Biology 12, no. 7: 905. https://doi.org/10.3390/biology12070905
APA StyleGaber, A., Ahmed, O. M., Khadrawy, Y. A., Zoheir, K. M. A., Abo-ELeneen, R. E., Alblihed, M. A., & Elbakry, A. M. (2023). Mesenchymal Stem Cells and Begacestat Mitigate Amyloid-β 25–35-Induced Cognitive Decline in Rat Dams and Hippocampal Deteriorations in Offspring. Biology, 12(7), 905. https://doi.org/10.3390/biology12070905