
Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
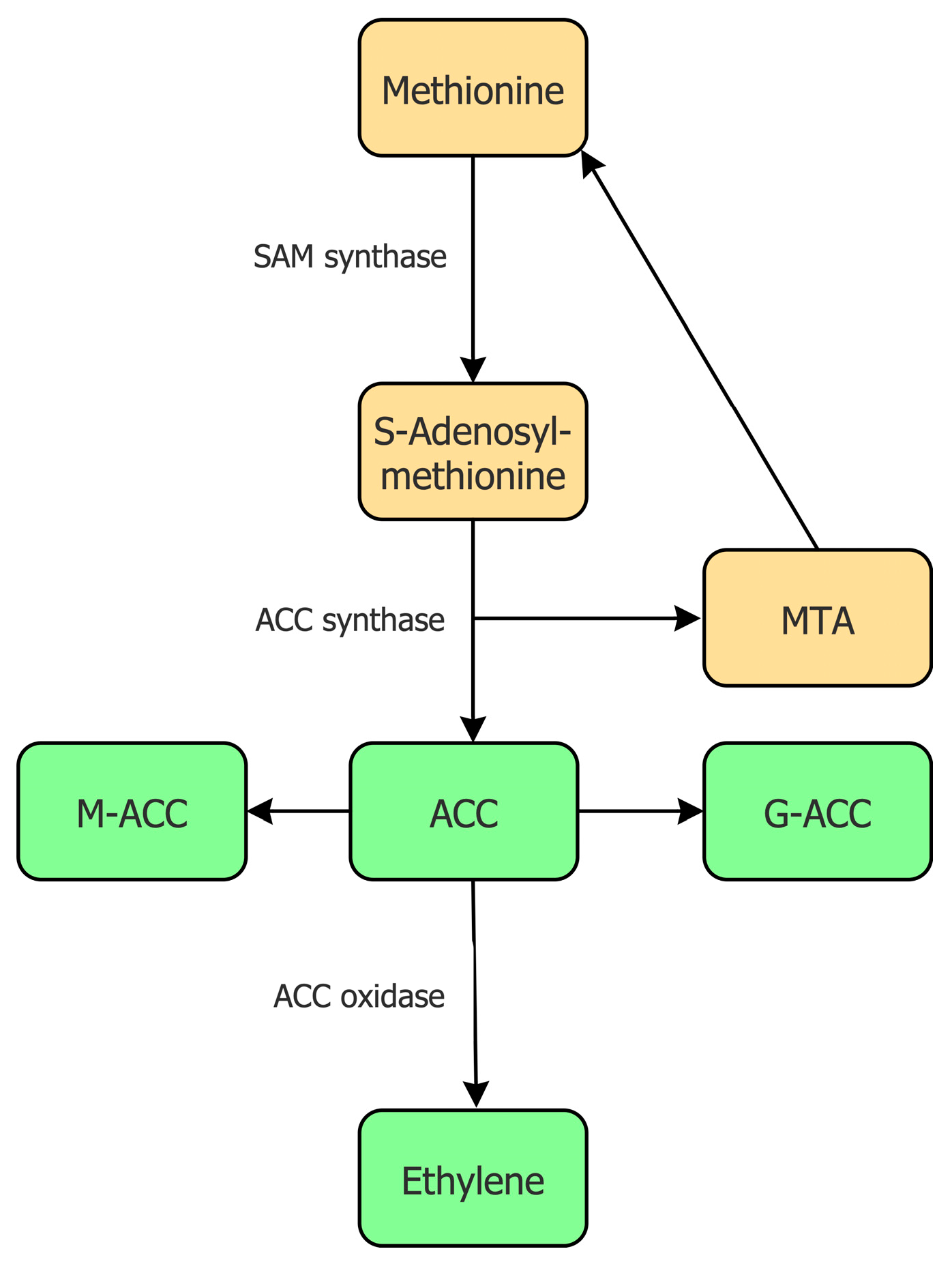
2. Ethylene and ACC Biosynthesis in Plants
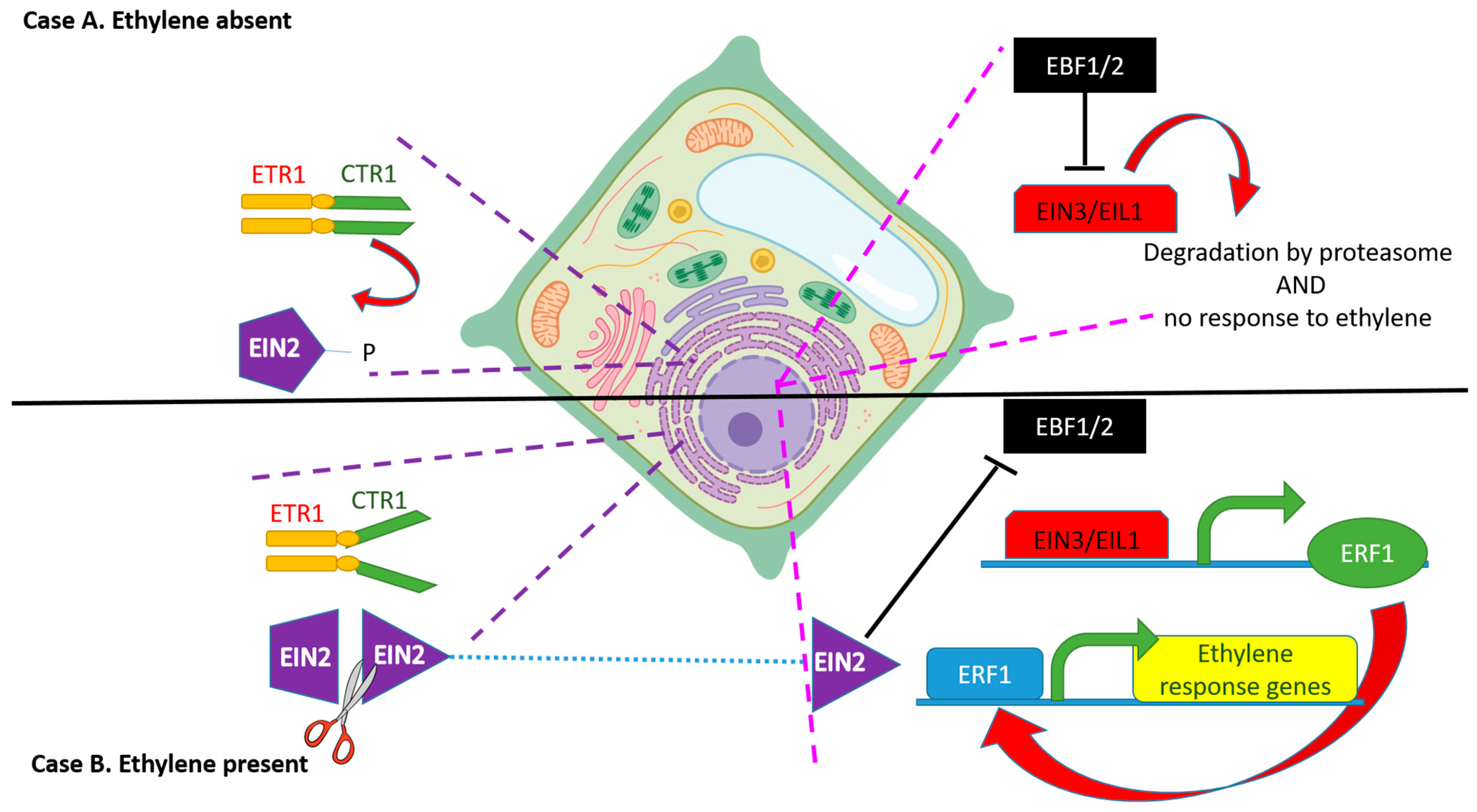
3. Ethylene as a Signaling Molecule
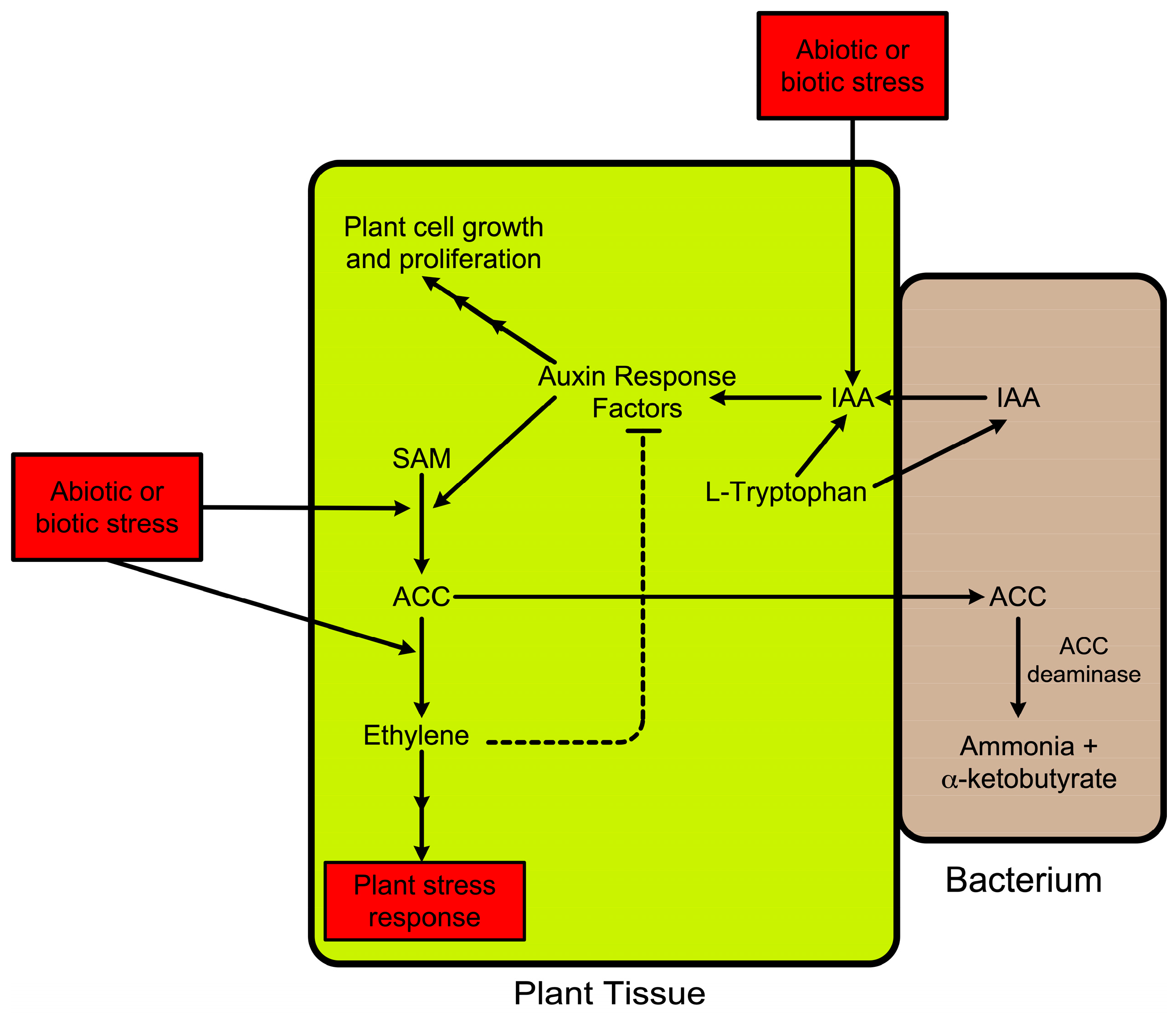
4. ACC as a Signaling Molecule
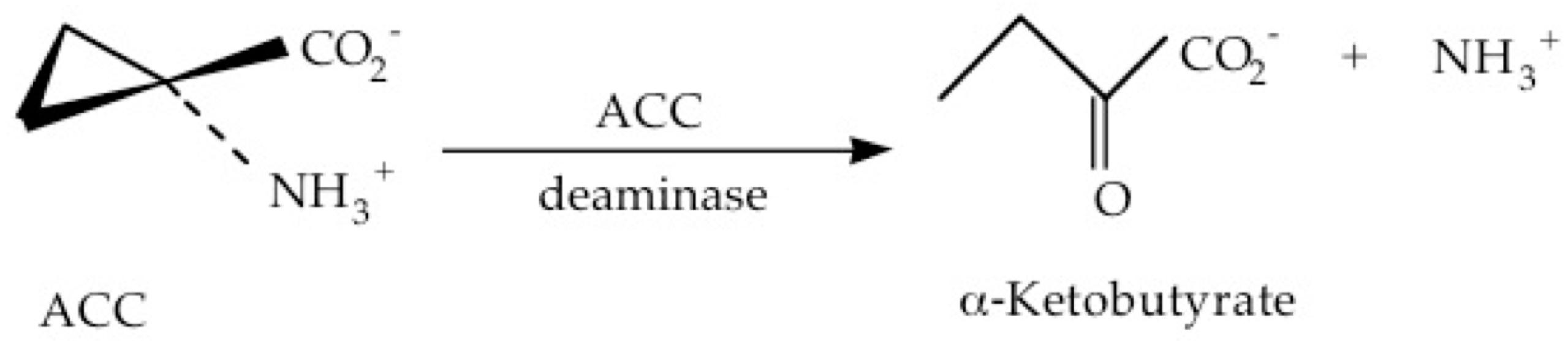
5. ACC Deaminase
6. D-Amino Acids in Plants
7. D-Cysteine Desulfhydrase
8. Plant Evolution
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Orozco-Mosqueda, M.C.; Santoyo, X.; Glick, B.R. Recent advances in the bacterial phytohormone modulation of plant growth. Plants 2023, 12, 606. [Google Scholar] [CrossRef]
- Weyers, J.D.B.; Patterson, N.W. Plant hormones and the control of physiological processes. N. Phytol. 2001, 152, 375–407. [Google Scholar] [CrossRef]
- Santaner, A.; Calderon-Villalobos, L.I.A.; Estelle, M. Plant hormones are versatile regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Sabagh, A.E.L.; Islam, M.S.; Hossain, A.; Iqbal, M.A.; Mubeen, M.; Waleed, M.; Reginato, M.; Battaglia, M.; Ahmed, S.; Rehman, A.; et al. Phytohormones as growth regulators during abiotic stress tolerance in plants. Front. Agron. 2022, 4, 765068. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, Q.; Wang, B.; Yuan, F. Roles of phytohormones and their signaling pathways in leaf development and stress responses. J. Agric. Food Chem. 2021, 69, 3566–3584. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stress. Plant Grow. Regulat. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Abeles, F.; Morgan, P.W.; Saltveit, M.E., Jr. Ethylene in Plant Biology, 2nd ed.; Academic Press: San Diego, CA, USA, 1992. [Google Scholar]
- Glick, B.R. Beneficial Plant-Bacterial Interactions, 2nd ed.; Springer: Heidelberg/Berlin, Germany, 2020; 383p. [Google Scholar]
- Mattoo, A.K.; Suttle, J.C. The Plant Hormone Ethylene; CRC Press, Inc.: Boca Raton, FL, USA, 1991; 347p. [Google Scholar]
- Chang, C.; Qamp, A. How do plants respond to ethylene and what is its importance? BMC Biol. 2016, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.B. Ethylene-promoted elongation: An adaptation to submergence stress. Ann. Bot. 2008, 101, 229–248. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Tivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, F.X.; McConkey, B.J.; Glick, B.R. New insights into ACC deaminase phylogeny, evolution, and evolutionary significance. PLoS ONE 2014, 9, e99168. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef] [PubMed]
- Yoon, G.M.; Kieber, J.J. 14-3-3 regulates 1-aminocyclopropane-1-carboxylate synthase protein turnover in Arabidopsis. Plant Cell 2013, 25, 1016–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorovic, B.; Glick, B.R. The interconversion of ACC deaminase and D-cysteine desulfhydrase by directed mutagenesis. Planta 2008, 229, 193–205. [Google Scholar] [CrossRef]
- Pattyn, J.; Vaughn-Hirsch, J.; Van de Poel, B. The regulation of ethylene biosynthesis: A complex multilevel control circuitry. N. Phytol. 2021, 229, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Fluhr, R.; Matoo, A.K.; Dilley, D.R. Ethylene—Biosynthesis and perception. Crit. Rev. Plant Sci. 2008, 15, 479–523. [Google Scholar]
- Freebairn, H.T.; Buddenhagen, I.W. Ethylene production by Pseudomonas solanacearum. Nature 1964, 202, 313–314. [Google Scholar] [CrossRef]
- Eckert, C.; Xu, W.; Xiong, W.; Lynch, S.; Ungerer, J.; Tao, L.; Gill, R.; Maness, P.-C.; Yu, J. Ethylene-forming enzyme and bioethylene production. Biotechnol. Biofuels 2014, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, B.; Li, C.; Tarczynski, M.C. High free-methionine and decreased lignin content result from a mutation in the Arabidopsis S-adenosyl-L-methionine synthetase 3 gene. Plant J. 2002, 29, 371–380. [Google Scholar] [CrossRef]
- Zarembinski, T.I.; Theologis, A. Ethylene biosynthesis and action: A case of conservation. Plant Mol. Biol. 1994, 26, 1579–1597. [Google Scholar] [CrossRef]
- Eun, H.-D.; Ali, S.; Jung, H.; Kim, K.; Kim, W.-C. Profiling of ACC synthase gene (ACS11) expression in Arabidopsis induced by abiotic stresses. Appl. Biol. Chem. 2019, 62, 42. [Google Scholar] [CrossRef] [Green Version]
- Harpaz-Saad, S.; Yoon, G.M.; Mattoo, A.; Kieber, J.J. The formation of ACC and competition between polyamides and ethylene for SAM. Ann. Plant Rev. 2012, 44, 53–81. [Google Scholar]
- Polko, J.K.; Kieber, J.J. 1-Aminocyclopropane 1-carboxylic acid and its emerging role as an ethylene-dependent growth regulator. Front. Plant Sci. 2019, 10, 1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kende, H. Enzymes of ethylene biosynthesis. Plant Physiol. 1989, 91, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, A.J.; Lycett, G.W.; Grierson, D. Antisense gene that inhibits synthesis of the hormone ethylene in transgenic plants. Nature 1990, 346, 284–287. [Google Scholar] [CrossRef]
- Houben, M.; Van de Poel, B. 1-Aminocyclopropane-1-carboxylic acid oxidase (ACO): The enzyme that makes the plant hormone ethylene. Front. Plant Sci. 2019, 10, 695. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.S.; Blume, B.; Bouzayen, M.; Cooper, W.; Hamilton, A.J.; Grierson, D. Differential expression of the 1-aminocyclopropane-1-carboxylate oxidase gene family of tomato. Plant J. 1996, 9, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Peiser, G.; Yang, S.F. Evidence for 1-(malonylamino)cyclopropane-1-carboxylic acid being the major conjugate of aminocyclopropane-1-carboxylic acid in tomato fruit. Plant Physiol. 1998, 116, 1527–1532. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.F. The biosynthesis and metabolism of 1-(malonylamino) cyclopropane-1-carboxylic acid in relation to ethylene production. In Conjugated Plant Hormones: Structure, Metabolism, and Function; Schreiber, K., Schütte, H.R., Sembdner, G., Eds.; VEB Deutscher Verlag der Wissenschaaften: Berlin, Germany, 1987; pp. 92–101. [Google Scholar]
- Martin, M.N.; Cohen, J.D.; Saftner, R.A. A new 1-aminocyclopropane-1-carboxylic acid conjugating activity in tomato fruit. Plant Physiol. 1995, 109, 917–926. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.F.; Hoffman, N.E. Ethylene biosynthesis and its regulation in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Bakshi, A.; Shemansky, J.M.; Chang, C.; Binder, B.M. History of research on the plant hormone ethylene. J. Plant Growth Regul. 2015, 34, 809–827. [Google Scholar] [CrossRef]
- Fahnestock, G.W. Memoranda of the effects of carbureted hydrogen gas upon a collection of exotic plants. Proc. Acad. Nat. Sci. Phil. 1858, 9–10, 118–134. [Google Scholar]
- Neljubow, D. Uber die horizontale nutation der stengel von Pisum sativum und einiger anderen pflanzen. Beih. Bot. Zentralb. 1901, 10, 128–139. [Google Scholar]
- Bleecker, A.B. Ethylene perception and signaling: An evolutionary perspective. Trends Plant Sci. 1999, 4, 269–274. [Google Scholar] [CrossRef]
- Mount, S.M.; Chang, C. Evidence for a plastid origin of plant ethylene receptor gene. Plant Physiol. 2002, 130, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, F.I.; Esch, J.J.; Hall, A.E.; Binder, B.M.; Schaller, G.E.; Bleecker, A.B. A copper cofactor for the ethylene receptor ETR1 from Arabidopsis. Science 1999, 283, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Sisler, E.C. Measurement of ethylene binding in plant tissue. Plant Physiol. 1979, 64, 538–542. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.E.; Bengochea, T.; Cairns, A.J.; Dodds, J.H.; Hall, M.A. Studies on ethylene binding by cell-free preparations from cotyledons of Phaseolus vulgaris L.: Subcellular localization. Plant Cell Environ. 1982, 5, 101–107. [Google Scholar]
- Evans, D.E.; Dodds, J.H.; Lloyd, P.C.; Apgwynn, I.; Hall, M.A. A study of the subcellular localization of an ethylene binding site in developing cotyledons of Phaseolus vulgaris L. by high resolution autoradiography. Planta 1982, 154, 48–52. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, R.C.; Rodriguez, F.I.; Esch, J.J.; Binder, B.M.; O’Donnell, P.; Klee, H.J.; Bleecker, A.B. Ethylene-binding activity, gene expression levels, and receptor system output for ethylene receptor family members from Arabidopsis and tomato. Plant J. 2005, 41, 651–659. [Google Scholar] [CrossRef]
- Hall, A.E.; Findell, J.L.; Schaller, G.E.; Sisler, E.C.; Bleecker, A.B. Ethylene perception by the ERS1 protein in Arabidopsis. Plant Physiol. 2000, 123, 1449–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.; Liu, Q.; Wen, C.-K. Receptor signal output mediated by the ETR1 N terminus is primarily subfamily I receptor dependent. Plant Physiol. 2006, 142, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Grefen, C.; Städele, K.; Růzicka, K.; Obrdlik, P.; Harter, K.; Horák, J. Subcellular localization and in vivo interactions of the Arabidopsis thaliana ethylene receptor family members. Mol. Plant 2008, 1, 308–320. [Google Scholar] [CrossRef]
- Gao, Z.; Wen, C.-K.; Binder, B.M.; Chen, Y.-F.; Chang, J.; Chiang, Y.-H.; Kerris, R.J.; Chang, C.; Schaller, G.E. Heteromeric interactions among ethylene receptors mediate signaling in Arabidopsis. J. Biol. Chem. 2008, 283, 23801–23810. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.S.; Burden, L.M.; Hurley, J.H. Structure of the GAF domain, a ubiquitous signaling motif and a new class of cyclic GMP receptor. EMBO J. 2000, 19, 5288–5299. [Google Scholar] [CrossRef] [Green Version]
- Binder, B.M.; Rodriguez, F.I.; Bleecker, A.B. The copper transporter RAN1 is essential for biogenesis of ethylene receptors in Arabidopsis. J. Biol. Chem. 2010, 285, 37263–37270. [Google Scholar] [CrossRef] [Green Version]
- Schott-Verdugo, S.; Müller, L.; Classen, E.; Gohlke, H.; Groth, G. Structural model of the ETR1 Ethylene Receptor Transmembrane Sensor Domain. Sci. Rep. 2019, 9, 8869. [Google Scholar] [CrossRef] [Green Version]
- Azhar, B.J.; Abbas, S.; Aman, S.; Yamburenko, M.V.; Chen, W.; Müller, L.; Uzun, B.; Jewell, D.A.; Dong, J.; Shakeel, S.N.; et al. Basis for high-affinity ethylene binding by the ethylene receptor ETR1 of Arabidopsis. Proc. Natl. Acad. Sci. USA 2023, 120, e2215195120. [Google Scholar] [CrossRef]
- Matoo, K.; Baker, J.E.; Moline, H.E. Induction by Copper Ions of Ethylene Production in Spirodela oligorrhiza: Evidence for a Pathway Independent of 1-Aminocyclopropane-1-carboxylic Acid. J. Plant Physiol. 1983, 123, 193–202. [Google Scholar] [CrossRef]
- Roman, G.; Lubarsky, B.; Kieber, J.J.; Rothenberg, M.; Ecker, J.R. Genetic analysis of ethylene signal transduction in Arabidopsis thaliana: Five novel mutant loci integrated into a stress response pathway. Genetics 1995, 139, 1393–1409. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.L.; Larsen, P.B.; Wang, X.; Chang, C. Association of the Arabidopsis CTR1 Raf-like kinase with the ETR1 and ERS ethylene receptors. Proc. Natl. Acad. Sci. USA 1998, 95, 5401–5406. [Google Scholar] [CrossRef]
- Cancel, J.D.; Larsen, P.B. Loss-of-function mutations in the ethylene receptor ETR1 cause enhanced sensitivity and exaggerated response to ethylene in Arabidopsis. Plant Physiol. 2002, 129, 1557–1567. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Chen, Y.F.; Randlett, M.D.; Zhao, X.C.; Findell, J.L.; Kieber, J.J.; Schaller, G.E. Localization of the Raf-like kinase CTR1 to the endoplasmic reticulum of Arabidopsis through participation in ethylene receptor signaling complexes. J. Biol. Chem. 2003, 278, 34725–34732. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Li, H.; Hutchison, C.E.; Laskey, J.; Kieber, J.J. Biochemical and functional analysis of CTR1, a protein kinase that negatively regulates ethylene signaling in Arabidopsis. Plant J. 2003, 33, 221–233. [Google Scholar] [CrossRef]
- Alonso, J.M.; Hirayama, T.; Roman, G.; Nourizadeh, S.; Ecker, J.R. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 1999, 284, 2148–2152. [Google Scholar] [CrossRef]
- Bisson, M.M.; Bleckmann, A.; Allekotte, S.; Groth, G. EIN2, the central regulator of ethylene signaling, is localized at the ER membrane where it interacts with the ethylene receptor ETR1. Biochem. J. 2009, 23, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Ju, C.; Van de Poel, B.; Cooper, E.D.; Thierer, J.H.; Gibbons, T.R.; Delwiche, C.F.; Chang, C. Conservation of ethylene as a plant hormone over 450 million years of evolution. Nat. Plants 2015, 1, 14004. [Google Scholar] [CrossRef]
- Qiao, H.; Shen, Z.; Huang, S.S.; Schmitz, R.J.; Urich, M.A.; Briggs, S.P.; Ecker, J.R. Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas. Science 2012, 338, 390–393. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ma, M.; Feng, Y.; Li, H.; Wang, Y.; Ma, Y.; Li, M.; An, F.; Guo, H. EIN2-directed translational regulation of ethylene signaling in Arabidopsis. Cell 2015, 163, 670–683. [Google Scholar] [CrossRef] [Green Version]
- Dolgikh, V.A.; Pukhovaya, E.M.; Zemlyanskaya, E.V. Shaping ethylene response: The role of EIN3/EIL1 Transcription Factors. Front. Plant Sci. 2019, 10, 1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.L.; Rahman, A.; Baskin, T.I.; Kieber, J.J. Two leucine-rich repeat receptor kinases mediate signaling linking cell wall biosynthesis and ACC synthase in Arabidopsis. Plant Cell 2008, 20, 3065–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, D.L.; Edmond, C.; Harrington, J.L.; Nuhse, T.S. Cell wall integrity controls root elongation via a general 1-aminocyclopropane-1-carboxylic acid-dependent, ethylene-independent pathway. Plant Physiol. 2011, 156, 596–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderstraeten, L.; Depaepe, T.; Bertrand, S.; Van Der Straeten, D. The ethylene precursor ACC affects early vegetative development independently of ethylene signaling. Front. Plant Sci. 2019, 10, 1591. [Google Scholar] [CrossRef] [Green Version]
- Van de Poel, B.; Van Der Straeten, D. 1-Aminocyclopropane-1-carboxylic acid (ACC) in plants: More than just the precursor of ethylene! Front. Plant Sci. 2014, 5, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, W.; Kao, Y.-T.; Michard, E.; Simon, A.A.; Li, D.; Wudick, M.M.; Lizzio, M.A.; Feijo, J.A.; Chang, C. Ethylene-independent signalling by the ethylene precursor ACC in Arabidopsis ovular pollen tube attraction. Nat. Common. 2020, 11, 4082. [Google Scholar] [CrossRef] [PubMed]
- Tsuchisaka, A.; Yu, G.; Jin, H.; Alonso, J.M.; Ecker, J.R.; Zhang, X.; Gao, S.; Theologis, A. A combinatorial interplay among the 1-aminocyclopropane-1-carboxylate isoforms regulates ethylene biosynthesis in Arabidopsis thaliana. Genetics 2009, 183, 979–1003. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Zhang, X.; Zhang, G.; Wen, Y.; Liang, G.; Chen, X. Aminocyclopropane-1-carboxylic acid is a key regulator of guard mother cell terminal division in Arabidopsis thaliana. J. Exp. Bot. 2018, 70, 897–908. [Google Scholar] [CrossRef] [Green Version]
- Tsolakidou, M.-D.; Pantelides, L.S.; Tzima, A.K.; Kang, S.; Paplomatas, E.J.; Tsaltas, D. Disruption and overexpression of the gene encoding acc (1-aminocyclopropane-1-carboxylic acid) deaminase in soil-borne fungal pathogen Verticillium dahlia revealed the role of ACC as a potential regulator of virulence and plant defense. Mol. Plant Microbe Interact. 2019, 32, 639–653. [Google Scholar] [CrossRef]
- Bradford, K.J.; Yang, S.F. Xylem transport of 1-aminocyclopropane-1-carboxylic acid, an ethylene precursor, in waterlogged tomato plants. Plant Physiol. 1980, 65, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Else, M.A.; Jackson, M.B. Transport of 1-aminocyclopropane-1-carboxylic acid (ACC) in the transpiration stream of tomato (Lycopersicon esculentum) in relation to foliar ethylene production and petiole epinasty. Aust. J. Plant Physiol. 1998, 25, 453–458. [Google Scholar] [CrossRef]
- Chen, Y.; Bonkowski, M.; Shen, Y.; Griffiths, B.S.; Jiang, Y.; Wang, X.; Sun, B. Root ethylene mediates rhizosphere microbial community reconstruction when chemically detecting cyanide produced by neighbouring plants. Microbiome 2020, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, F.; Rossi, M.; Glick, B. Ethylene and 1-aminocyclopropane-1-carboxylate (ACC) in plant-bacterial interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Mou, W.; Van de Poel, B.; Chang, C. Something old, something new: Conservation of the ethylene precursor 1-amino-cyclopropane-1-carboxylic acid as a signaling molecule. Curr. Opin. Plant Biol. 2022, 65, 102116. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.-J.; Chen, H.-W.; Ma, B.; Zhang, W.-K.; Chen, S.Y.; Zhang, J.-S. The role of ethylene in plants under salinity stress. Front. Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef] [Green Version]
- Stearns, J.C.; Glick, B.R. Transgenic plants with altered ethylene biosynthesis or perception. Biotechnol. Adv. 2003, 21, 193–210. [Google Scholar] [CrossRef]
- Timmusk, S.; Paalme, V.; Pavlicek, T.; Bergquist, J.; Vangala, A.; Danilas, T.; Nevo, E. Bacterial distribution in the rhizosphere of wild barley under contrasting microclimates. PLoS ONE 2011, 6, 17968. [Google Scholar] [CrossRef]
- Ekimova, G.A.; Fedorov, D.N.; Tani, A.; Doronina, N.V.; Trotsenko, Y.A. Distribution of 1-aminocyclopropane-1-carboxylate deaminase and d-cysteine desulfhydrase genes among type species of the genus Methylobacterium. Antonie Leeuwenhoek 2018, 111, 1723–1734. [Google Scholar] [CrossRef]
- Reed, M.L.E.; Glick, B.R. Applications of Plant Growth-Promoting Bacteria for Plant and Soil Systems; Gupta, V.K., Schmoll, M., Maki, M., Tuohy, M., Mazutti, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Grichko, V.P.; Glick, B.R. Amelioration of flooding stress by ACC deaminase-containing plant growth-promoting bacteria. Plant Physiol. Biochem. 2001, 39, 11–17. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M. The use of ACC deaminase-containing plant growth-promoting bacteria to protect plants against the deleterious effects of ethylene. In Plant Surface Microbiology; Springer: Berlin/Heidelberg, Germany, 2004; pp. 133–144. [Google Scholar]
- Glick, B.R. Bacterial ACC deaminase and the alleviation of plant stress. Adv. Appl. Microbiol. 2004, 56, 291–312. [Google Scholar]
- Glick, B.R. Modulation of plant ethylene levels by the enzyme ACC deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Saleh-Lakha, S.; Hontzeas, N.; Glick, B.R. Physiological and genetic effects of bacterial ACC deaminase on plants. In Microbial Biotechnology in Agriculture and Aquaculture; Science Publishers: Enfield, UK, 2006; Volume II, pp. 199–224. [Google Scholar]
- Gerhardt, K.; Greenberg, B.M.; Glick, B.R. The role of ACC deaminase in facilitating the phytoremediation of organics, metals and salt. Curr. Trends Microbiol. 2007, 2, 61–73. [Google Scholar]
- Cheng, Z.; Park, E.; Glick, B.R. 1-Aminocyclopropane-1-carboxylate (ACC) deaminase from Pseudomonas putida UW4 facilitates the growth of canola in the presence of salt. Can. J. Microbiol. 2007, 53, 912–918. [Google Scholar] [CrossRef]
- Hao, Y.; Charles, T.C.; Glick, B.R. ACC deaminase from plant growth promoting bacteria affects crown gall development. Can. J. Microbiol. 2007, 53, 1291–1299. [Google Scholar] [CrossRef]
- Gamalero, E.; Berta, G.; Massa, N.; Glick, B.R.; Lingua, G. 2008. Synergistic interactions between the ACC deaminase-producing bacterium Pseudomonas putida UW4 and the AM fungus Gigaspora rosea positively affect cucumber plant growth. FEMS Microbiol. Ecol. 2008, 64, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Charles, T.C.; Glick, B.R. ACC deaminase increases A. tumefaciens-mediated transformation frequency of commercial canola cultivars. FEMS Microbiol. Lett. 2010, 307, 185–190. [Google Scholar] [CrossRef]
- Toklikishvili, N.; Dandurishvili, N.; Tediashvili, M.; Giorgobiani, N.; Szegedi, E.; Glick, B.R.; Vainstein, A.; Chernin, L. Inhibitory effect of ACC deaminase-producing bacteria on crown gall formation in tomato plants infected by Agrobacterium tumefaciens or A. vitis. Plant Pathol. 2010, 59, 1023–1030. [Google Scholar] [CrossRef]
- Hao, Y.; Charles, T.C.; Glick, B.R. An ACC deaminase containing A. tumefaciens strain D3 shows biocontrol activity to crown gall disease. Can. J. Microbiol. 2011, 57, 278–286. [Google Scholar] [CrossRef]
- Siddikee, M.A.; Glick, B.R.; Chauhan, P.S.; Yim, W.-J.; Sa, T. Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L.) by regulating stress ethylene synthesis with halotolerant bacteria containing ACC deaminase activity. Plant Physiol. Biochem. 2011, 49, 427–434. [Google Scholar] [CrossRef]
- Chernin, L.; Glick, B.R. The use of ACC deaminase to increase the tolerance of plants to various phytopathogens. In Bacteria in Agrobiology: Stress Management; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 279–299. [Google Scholar]
- Ali, S.; Charles, T.C.; Glick, B.R. Delay of carnation flower senescence by bacterial endophytes expressing ACC deaminase. J. Appl. Microbiol. 2012, 113, 1139–1144. [Google Scholar] [CrossRef]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2013, 366, 93–105. [Google Scholar] [CrossRef]
- Kong, Z.; Glick, B.R.; Duan, J.; Ding, S.; Tian, J.; McConkey, B.; Wei, G. Effects of 1-aminocyclopropane-1-carboxylate (ACC) deaminase-overproducing Sinorhizobium meliloti on plant growth and copper tolerance of Medicago lupulina. Plant Soil 2015, 391, 383–398. [Google Scholar] [CrossRef]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase: A weapon to “stress ethylene” produced in plants. Front. Microbiol. 2015, 6, 937. [Google Scholar] [PubMed] [Green Version]
- Mendoza-Hernández, J.C.; Perea-Vélez, Y.S.; Arriola-Morales, J.; Martínez-Simón, S.M.; Pérez-Osorio, G. Assessing the effects of heavy metals in ACC deaminase and IAA production on plant growth-promoting bacteria. Microbiol. Res. 2016, 188–189, 53–61. [Google Scholar]
- Gamalero, E.; Marzachì, C.; Galetto, L.; Veratti, F.; Massa, N.; Bona, E.; Novello, G.; Glick, B.R.; Ali, S.; Cantamessa, S.; et al. An ACC deaminase-expressing endophyte increases plant tolerance to Flavescence dorée phytoplasma infection. Plant Biosys. 2016, 151, 331–340. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Role of ACC deaminase in stress control of leguminous plants. In Plant Growth Promoting Actinobacteria; Subramanian, G., Arumugam, S., Rajendran, V., Eds.; Springer Science: Singapore, 2016; pp. 179–192. [Google Scholar]
- Nascimento, F.X.; Tavares, M.J.; Franck, J.; Ali, S.; Glick, B.R.; Rossi, M.J. ACC deaminase plays a major role in Pseudomonas fluorescens YsS6 ability to promote the nodulation of Alpha- and Betaproteobacteria rhizobial strains. Arch. Microbiol. 2019, 201, 817–822. [Google Scholar] [CrossRef]
- Gupta, S.; Pandey, S. ACC deaminase producing bacteria with multifarious plant growth promoting traits alleviates salinity stress in French bean (Phaseolus vulgaris) plants. Front. Microbiol. 2019, 10, 1506. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Mosqueda, M.C.; Duan, J.; DiBernardo, M.; Zetter, E.; Campos-García, J.; Glick, B.R.; Santoyo, G. The production of ACC deaminase and trehalose by the plant growth promoting bacterium Pseudomonas sp. UW4 synergistically protect tomato plants against salt stress. Front. Microbiol. 2019, 10, 1392. [Google Scholar] [CrossRef] [Green Version]
- Chandra, D.; Srivastava, R.; Glick, B.R.; Sharma, A.K. Rhizobacteria producing ACC deaminase mitigate water-stress response in finger millet (Eleusine coracana (L.) Gaertn.). 3 Biotech 2020, 10, 65. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.C.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Paço, A.; da-Silva, J.R.; Denise Pereira Torres, D.; Glick, B.R.; Brígido, C. Exogenous ACC deaminase is key to improving the performance of pasture legume-rhizobial symbioses in the presence of a high manganese concentration. Plants 2020, 9, 1630. [Google Scholar] [CrossRef]
- Duan, B.; Li, L.; Chen, G.; Su-Zhou, C.; Li, Y.; Merkeryan, H.; Liu, W.; Liu, X. 1-Aminocyclopropane-1-carboxylate deaminase producing plant growth-promoting rhizobacteria improve drought stress tolerance in grapevine (Vitis vinifera L.). Front. Plant Sci. 2021, 12, 706990. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Urón, P.; Glick, B.R.; Giachini, A.; Rossi, M.J. Genomic analysis of the ACC deaminase-producing Pseudomonas thivervalensis SC5 reveals its multifaceted roles in soil and beneficial interactions with plants. Front. Microbiol. 2021, 12, 752288. [Google Scholar] [CrossRef]
- Jacobson, C.B.; Pasternak, J.J.; Glick, B.R. Partial purification and characterization of ACC deaminase from the plant growth-promoting rhizobacterium Pseudomonas putida GR12-2. Can. J. Microbiol. 1994, 40, 1019–1025. [Google Scholar] [CrossRef]
- Honma, M. Chemically reactive sulfhydryl groups of 1-aminocyclopropane-1-carboxylate deaminase. Agric. Biol. Chem. 1985, 49, 567–571. [Google Scholar]
- Ali, S.; Glick, B.R. The biochemistry and molecular biology of the enzyme ACC deaminase. In Microbes: The Foundation Stone of the Biosphere; Hurst, C., Ed.; Springer: Cham, Switzerland, 2021; Chapter 19; pp. 365–390. [Google Scholar]
- Hontzeas, N.; Zoidakis, J.; Glick, B.R.; Abu-Omar, M.M. Expression and characterization of 1-aminocyclopropane-1-carboxylate deaminase from the rhizobacterium Pseudomonas putida UW4: A key enzyme in bacterial plant growth promotion. Biochim. Biophys. Acta 2004, 1703, 11–19. [Google Scholar] [CrossRef]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of plant growth by ACC deaminase-containing soil bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant-microbe interactions. Antonie Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Duca, D.R.; Glick, B.R. Indole-3-acetic acid biosynthesis and its regulation in plant-associated bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 8607–8619. [Google Scholar] [CrossRef]
- Stearns, J.C.; Woody, O.Z.; McConkey, B.J.; Glick, B.R. Effects of bacterial ACC deaminase on Brassica napus gene expression. Mol. Plant-Microbe Interact. 2012, 25, 668–676. [Google Scholar] [CrossRef] [Green Version]
- Peck, S.C.; Kende, H. A gene encoding 1-aminocyclopropane-1-carboxylate (ACC) synthase produces two transcripts: Elucidation of a conserved response. Plant J. 1998, 14, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Majda, M.; Robert, S. The role of auxin in cell wall expansion. Int. J. Mol. Sci. 2018, 9, 951. [Google Scholar] [CrossRef] [Green Version]
- Cronin, J.R.; Pizzarello, S. Enantiomeric excesses in meteoritic amino acids. Science 1997, 14, 951–955. [Google Scholar] [CrossRef]
- Aponte, J.C.; Elsila, J.E.; Hein, J.E.; Dworkin, J.P.; Glavin, D.P.; McLain, H.L.; Parker, E.T.; Cao, T.; Berger, E.L.; Burton, A.S. Analysis of amino acids, hydroxy acids, and amines in CR chondrites. Meteorit. Planet Sci. 2020, 55, 2422–2439. [Google Scholar] [CrossRef] [PubMed]
- Elsila, J.E.; Aponte, J.C.; Blackmond, D.G.; Burton, A.S.; Dworkin, J.P.; Glavin, D.P. Meteoritic amino acids: Diversity in compositions reflects parent body histories. ACS Cent. Sci. 2016, 22, 370–379. [Google Scholar] [CrossRef]
- Ollivaux, C.; Soyez, D.; Toullec, J.Y. Biogenesis of D-amino acid containing peptides/proteins: Where, when and how? J. Pept. Sci. 2014, 20, 595–612. [Google Scholar] [CrossRef] [Green Version]
- Veiga, P.; Piquet, S.; Maisons, A.; Furlan, S.; Courtin, P.; Chapot-Chartier, M.P.; Kulakauskas, S. Identification of an essential gene responsible for D-Asp incorporation in the Lactococcus lactis peptidoglycan crossbridge. Mol. Microbiol. 2006, 62, 1713–1724. [Google Scholar] [CrossRef]
- Bruckner, H.; Westhauser, T. Chromatographic determination of L- and d-amino acids in plants. Amino Acids 2003, 24, 43–55. [Google Scholar] [CrossRef]
- Michard, E.; Lima, P.T.; Borges, F.; Silva, A.C.; Portes, M.T.; Carvalho, J.E.; Gilliham, M.; Liu, L.H.; Obermeyer, G.; Feijo, J.A. Glutamate receptor-like genes form Ca 2+ channels in pollen tubes and are regulated by pistil-serine. Science 2011, 332, 434–437. [Google Scholar] [CrossRef] [Green Version]
- Vranova, V.; Zahradnickova, H.; Janous, D.; Skene, K.R.; Matharu, A.S.; Rejsek, K.; Formanek, P. The significance of d-amino acids in soil, fate and utilization by microbes and plants: Review and identification of knowledge gaps. Plant Soil 2012, 354, 21–39. [Google Scholar] [CrossRef]
- Gordes, D.; Kolukisaoglu, U.; Thurow, K. Uptake and conversion of d-amino acids in Arabidopsis thaliana. Amino Acids 2011, 40, 553–563. [Google Scholar] [CrossRef]
- Serralta-Interian, A.A.; Miranda-Ham, M.D.L.; Echevarría-Machado, I. Stimulation of root growth and enhanced nitrogenous metabolite content in habanero pepper (Capsicum chinense Jacq.) treated with a d-amino acid mixture. Theor. Exp. Plant Physiol. 2020, 32, 31–47. [Google Scholar] [CrossRef]
- Kolukisaoglu, Ü. D-amino acids in plants: Sources, metabolism, and functions. Int. J. Mol. Sci. 2020, 30, 5421. [Google Scholar] [CrossRef] [PubMed]
- Strauch, R.C.; Svedin, E.; Dilkes, B.; Chapple, C.; Li, X. Discovery of a novel amino acid racemase through exploration of natural variation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2015, 15, 11726–11731. [Google Scholar] [CrossRef] [PubMed]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 is a high-affinity transporter for cellular amino acid uptake in both root epidermis and leaf mesophyll. Plant Cell 2006, 18, 1931–1946. [Google Scholar] [CrossRef] [Green Version]
- Kolukisaoglu, Ü.; Suarez, J. D-Amino acids in plants: New insights and aspects, but also more open questions. In Amino Acid-New Insights and Roles in Plant and Animal; Asao, T., Asaduzzaman, M., Eds.; IntechOpen: London, UK, 2017; pp. 155–164. [Google Scholar]
- Lehmann, S.; Gumy, C.; Blatter, E.; Boeffel, S.; Fricke, W.; Rentsch, D. In planta function of compatible solute transporters of the AtProT family. J. Exp. Bot. 2011, 62, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Hener, C.; Hummel, S.; Suarez, J.; Stahl, M.; Kolukisaoglu, Ü. d-Amino Acids Are Exuded by Arabidopsis thaliana Roots to the Rhizosphere. Int. J. Mol. Sci. 2018, 19, 1109. [Google Scholar] [CrossRef] [Green Version]
- Suarez, J.; Hener, C.; Lehnhardt, V.A.; Hummel, S.; Stahl, M.; Kolukisaoglu, Ü. AtDAT1 is a key enzyme of D-Amino Acid stimulated ethylene production in Arabidopsis thaliana. Front. Plant Sci. 2019, 12, 1609. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.G.; Min, X.; Zhou, Z.H. Hydrogen sulfide: A signal molecule in plant cross-adaptation. Front. Plant Sci. 2016, 7, 1621. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Tanidokoro, K.; Shimizu, Y.; Kawarabayasi, Y.; Ohshima, T.; Sato, M.; Tadano, S.; Ishikawa, H.; Takio, S.; Takechi, K.; et al. Moss chloroplasts are surrounded by a peptidoglycan wall containing d-amino acids. Plant Cell. 2016, 28, 1521–1532. [Google Scholar] [CrossRef] [Green Version]
- Takano, H.; Takechi, K. Plastid peptidoglycan. Biochim. Biophys. Acta 2010, 1800, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, N.; Kudo, H.; Zhang, Z.; Li, J.; Wang, L.; Zhang, W.; Takechi, K.; Takano, H. Genes sufficient for synthesizing peptidoglycan are retained in Gymnosperm genomes, and MurE from Larix gmelinii can rescue the albino phenotype of Arabidopsis MurE mutation. Plant Cell Physiol. 2017, 1, 587–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemenschneider, A.; Wegele, R.; Schmidt, A.; Papenbrock, J. Isolation and characterization of a D-cysteine desulfhydrase protein from Arabidopsis thaliana. FEBS J. 2005, 272, 1291–1304. [Google Scholar] [CrossRef]
- Bharath, S.R.; Bisht, S.; Harijan, R.K.; Svithri, H.S.; Murthy, M.R.N. Structural and mutational studies on substrate specificity and catalysis of Salmonella typhimurium D-cysteine desulfhydrase. PLoS ONE 2012, 7, e36267. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A. A cysteine desulfhydrase from spinach leaves specific for D-cysteine. Z. Pflanzenphysiol. 1982, 107, 301–312. [Google Scholar] [CrossRef]
- Schmidt, A.; Erdle, I. A cysteine desulfhydrase specific for D-cysteine from the green alga Chlorella fusca. Z. Naturforsch. C 1983, 38, 428–435. [Google Scholar] [CrossRef]
- Rennenberg, H. Cysteine desulfhydrase activity in cucurbit plants: Simulation by preincubation with L- and D-cysteine. Phytochemistry 1983, 22, 1557–1560. [Google Scholar] [CrossRef]
- Rennenberg, H.; Arabatzis, N.; Grundel, I. Cysteine desulphydrase activity in higher plants: Evidence for the action of L- and D-cysteine specific enzymes. Phytochemistry 1987, 26, 1583–1589. [Google Scholar] [CrossRef]
- Soutourina, J.; Blanquet, S.; Plateau, P. Role of D-cysteine desulfhydrase in the adaptation of Escherichia coli to D-cysteine. J. Biol. Chem. 2001, 276, 40864–40872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Jiang, W.; Cheng, Z.; Heikkila, J.J.; Glick, B.R. The complete genome sequence of the plant growth-promoting bacterium Pseudomonas putida UW4. PLoS ONE 2013, 8, e58640. [Google Scholar]
- Moreira, D.; Philippe, H. Sure facts and open questions about the origin and evolution of photosynthetic plastids. Res. Microbiol. 2001, 152, 771–780. [Google Scholar] [CrossRef]
- Chu, K.H.; Qi, J.; Yu, Z.G.; Anh, V. Origin and phylogeny of chloroplasts revealed by a simple correlation analysis of complete genomes. Mol. Biol. Evol. 2004, 21, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ezpeleta, N.; Brinkmann, H.; Burey, S.C.; Roure, B.; Burger, G.; Löffelhardt, W.; Bohnert, H.J.; Philippe, H.; Lang, B.F. Monophyly of primary photosynthetic eukaryotes: Green plants, red algae, and glaucophytes. Curr. Biol. 2005, 26, 1325–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhfel, B.R.; Gitzendanner, M.A.; Soltis, P.S.; Soltis, D.E.; Burleigh, J.G. From algae to angiosperms-inferring the phylogeny of green plants (Viridiplantae) from 360 plastid genomes. BMC Evol. Biol. 2014, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Gitzendanner, M.A.; Soltis, P.S.; Wong, G.K.-S.; Ruhfel, B.R.; Soltis, D.E. Plastid phylogenomic analysis of green plants: A billion years of evolutionary history. Am. J. Bot. 2018, 105, 291–301. [Google Scholar] [CrossRef]
- Sato, N. Are Cyanobacteria an Ancestor of Chloroplasts or Just One of the Gene Donors for Plants and Algae? Genes 2021, 12, 823. [Google Scholar] [CrossRef]
- Burki, F.; Roger, A.J.; Brown, M.W.; Simpson, A.G.B. The new tree of Eukaryotes. Trends Ecol. Evol. 2020, 35, 43–55. [Google Scholar] [CrossRef] [Green Version]
- One Thousand Plant Transcriptomes Initiative. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, L.A.; McCourt, R.M. Green algae and the origin of land plants. Am. J. Bot. 2004, 91, 1535–1556. [Google Scholar] [CrossRef] [Green Version]
- Becker, B.; Marin, B. Streptophyte algae and the origin of embryophytes. Ann. Bot. 2009, 103, 999–1004. [Google Scholar] [CrossRef]
- Wickett, N.J.; Mirarab, S.; Nguyen, N.; Warnow, T.; Carpenter, E.; Matasci, N.; Ayyampalayam, S.; Barker, M.S.; Burleigh, J.G.; Gitzendanner, M.A.; et al. Phylotranscriptomic analysis of the origin and early diversification of land plants. Proc. Natl Acad. Sci. USA 2014, 111, E4859–E4868. [Google Scholar] [CrossRef]
- Lacey, R.F.; Binder, B.M. Ethylene regulates the physiology of the cyanobacterium Synechocystis sp. PCC 6803 via an ethylene receptor. Plant Physiol. 2016, 171, 2798–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narikawa, R.; Suzuki, F.; Yoshihara, S.; Higashi, K.; Watanabe, A.; Ikeuchi, M. Novel photosensory two-component system (PixA-NixB-NixC) involved in the regulation of positive and negative phototaxis of cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2011, 52, 2114–2224. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-Y.; Cho, H.S.; Cho, J.-I.; Jeon, J.-S.; Lagarias, J.C.; Park, Y.-I. Near-UV cyanobacteriochrome signaling system elicits negative phototaxis in the cyanobacterium Synechocystis sp. PCC 6803. Proc. Natl. Acad. Sci. USA 2011, 108, 10780–10785. [Google Scholar] [CrossRef]
- Ulijasz, A.T.; Cornilescu, G.; von Stetten, D.; Cornilescu, C.; Escobar, F.V.; Zhang, J.; Vierstra, R.D. Cyanochromes are blue/green light photoreversible photoreceptors defined by a stable double cysteine linkage to a phycoviolobilin-type chromophore. J. Biol. Chem. 2009, 284, 29757–29772. [Google Scholar] [CrossRef] [Green Version]
- Lacey, R.F.; Allen, C.J.; Bakshi, A.; Binder, B.M. Ethylene causes transcriptomic changes in Synechocystis during phototaxis. Plant Direct. 2018, 2, 1–16. [Google Scholar] [CrossRef]
- Wang, W.; Esch, J.J.; Shiu, S.H.; Agula, H.; Binder, B.M.; Chang, C.; Patterson, S.E.; Bleecker, A.B. Identification of important regions for ethylene binding and signaling in the transmembrane domain of the ETR1 ethylene receptor of Arabidopsis. Plant Cell 2006, 18, 3429–3442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Jimenez, P.; Robaina, R.R. Effects of ethylene on tetrasporogenesis in Pterocladiella capillacea (Rhodophyta). J. Phycol. 2012, 48, 710–715. [Google Scholar] [CrossRef]
- Maillard, P.; Thepenier, C.; Gudin, C. Determination of an ethylene biosynthesis pathway in the unicellular green-alga, Haematococcus pluvialis—Relationship between growth and ethylene production. J. Appl. Phycol. 1993, 5, 93–98. [Google Scholar] [CrossRef]
- Huang, T.-C.; Chow, T.-J. Ethylene production by blue-green algae. Bot. Bull. Acad. Sin. 1984, 25, 81–86. [Google Scholar]
- Chernys, J.; Kende, H. Ethylene biosynthesis in Regnellidium diphyllum and Marsilea quadrifolia. Planta 1996, 200, 113–118. [Google Scholar] [CrossRef]
- Osborne, D.J.; Walters, J.; Milborrow, B.V.; Norville, A.; Stange, L.M.C. Evidence for a non-ACC ethylene biosynthesis pathway in lower plants. Phytochemistry 1996, 42, 51–60. [Google Scholar] [CrossRef]
- Rohwer, F.; Bopp, M. Ethylene synthesis in moss protonema. J. Plant Physiol. 1985, 117, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Li, F.W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; de Vries, J.; Delaux, P.M.; Eily, A.; Koppers, N.; Kuo, L.Y.; Li, Z.; et al. Fern genomes elucidate land plant evolution and cyanobacterial symbioses. Nat. Plants 2018, 4, 460–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gamalero, E.; Lingua, G.; Glick, B.R. Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase. Biology 2023, 12, 1043. https://doi.org/10.3390/biology12081043
Gamalero E, Lingua G, Glick BR. Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase. Biology. 2023; 12(8):1043. https://doi.org/10.3390/biology12081043
Chicago/Turabian StyleGamalero, Elisa, Guido Lingua, and Bernard R. Glick. 2023. "Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase" Biology 12, no. 8: 1043. https://doi.org/10.3390/biology12081043
APA StyleGamalero, E., Lingua, G., & Glick, B. R. (2023). Ethylene, ACC, and the Plant Growth-Promoting Enzyme ACC Deaminase. Biology, 12(8), 1043. https://doi.org/10.3390/biology12081043