

Trophic Position of the Species and Site Trophic State Affect Diet Niche and Individual Specialization: From Apex Predator to Herbivore


 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Stable Isotope Analysis
2.3. Data Processing and Statistics
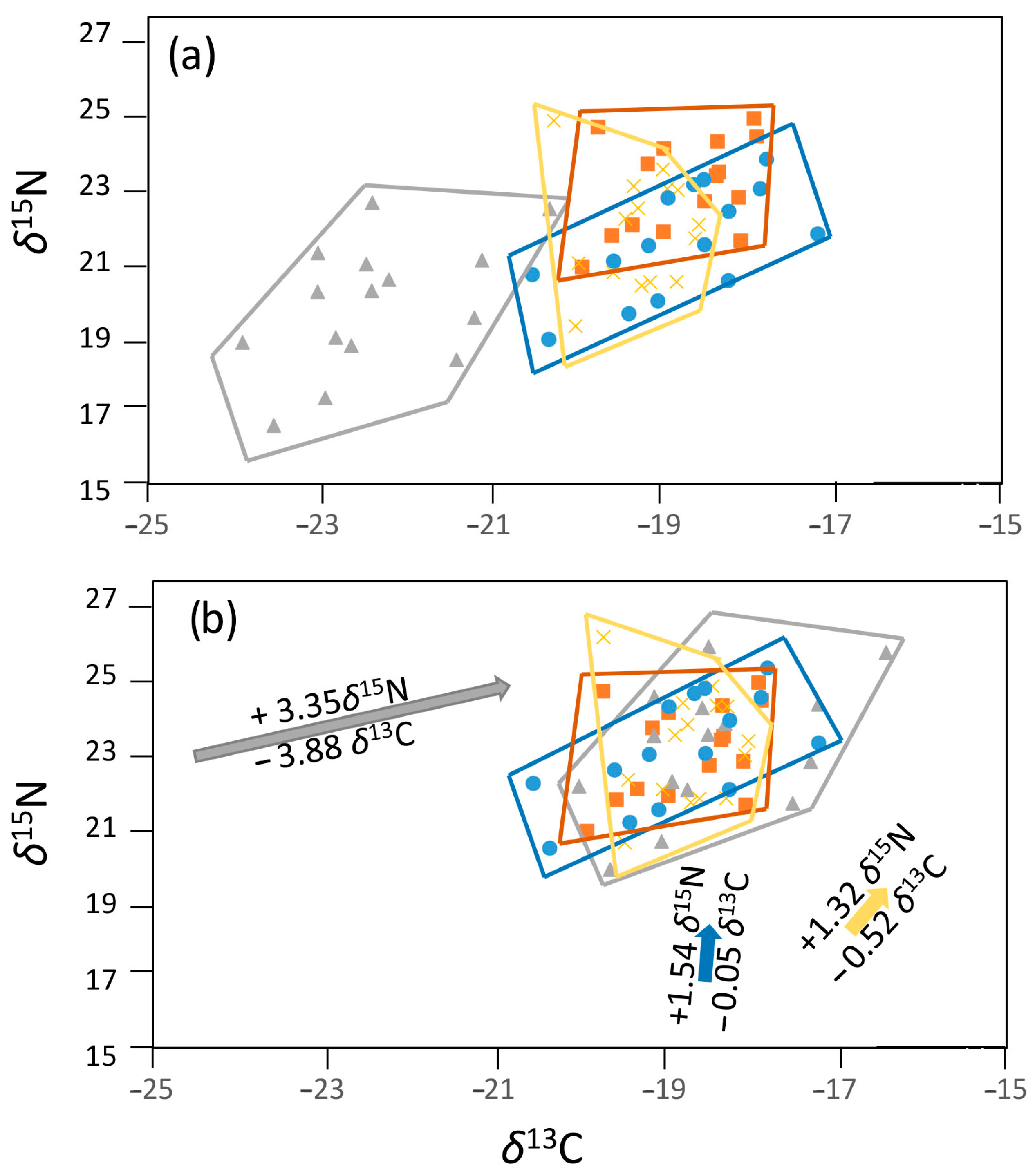
2.3.1. Data Centering
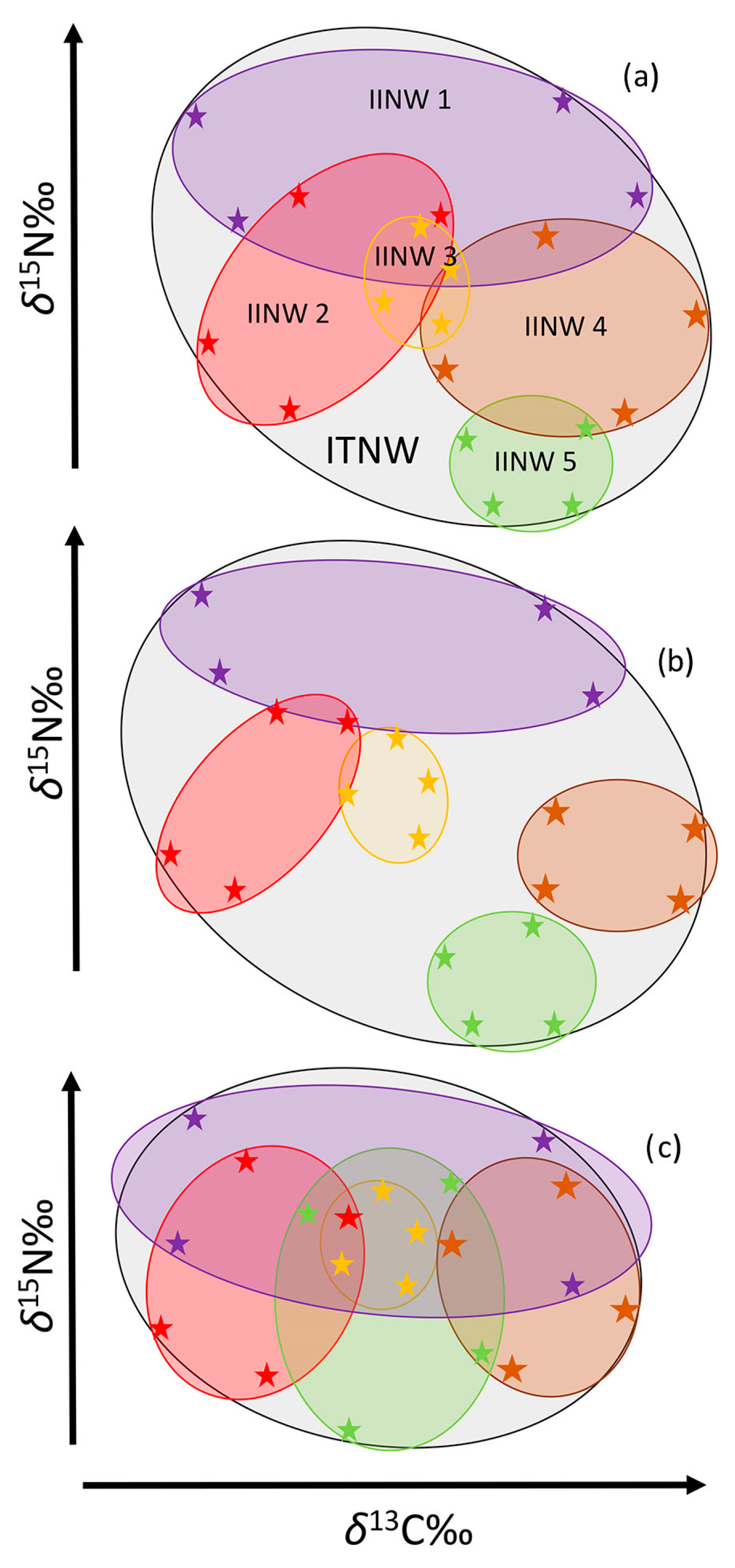
2.3.2. Isotopic Baseline Corrections and Subsequent Determination of ITNW and IINW
2.3.3. Degree of Individual Specialization (IIS)
3. Results
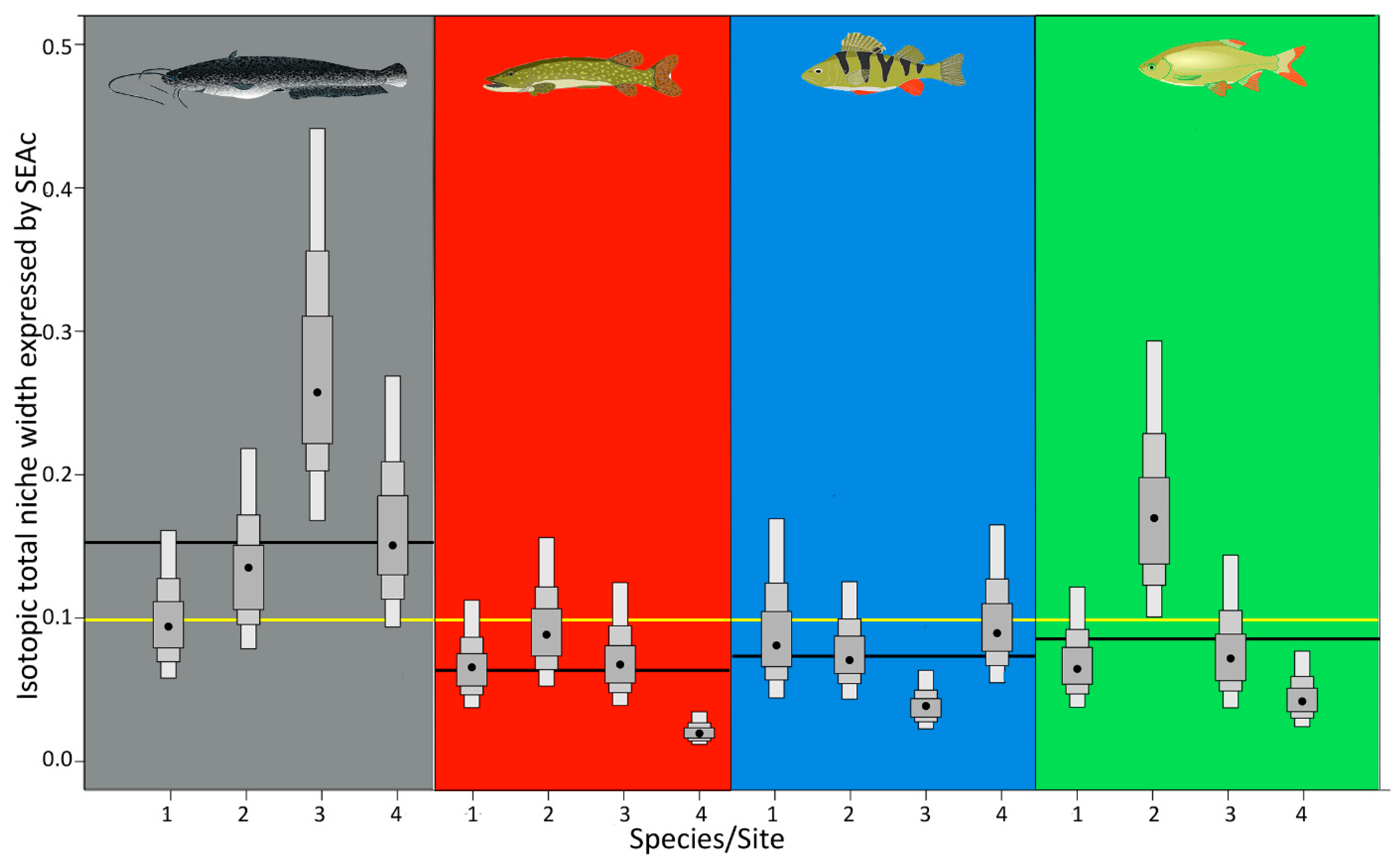
3.1. TNW in Relation to Site Trophic State
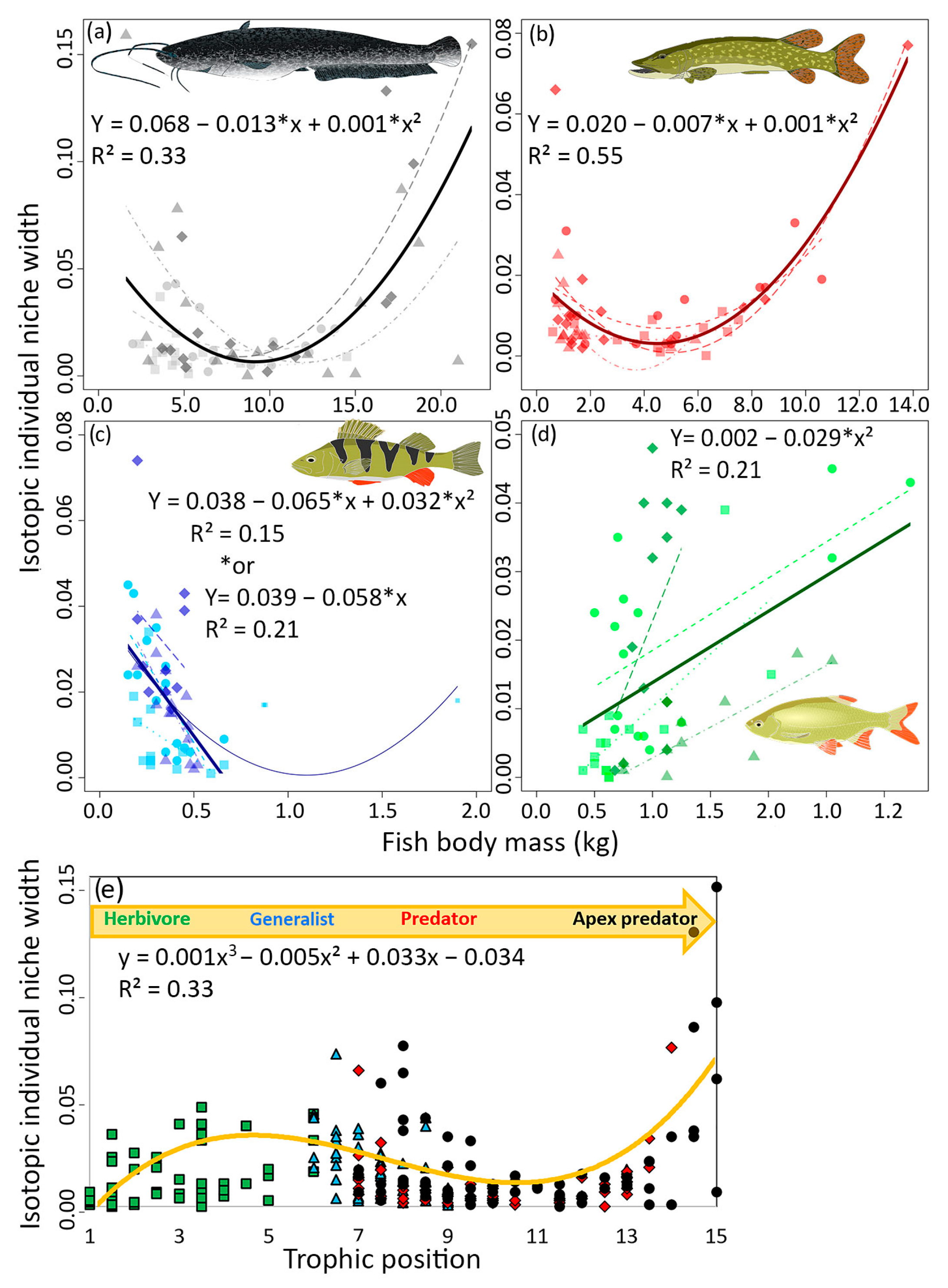
3.2. IINW in Relation to Body Mass
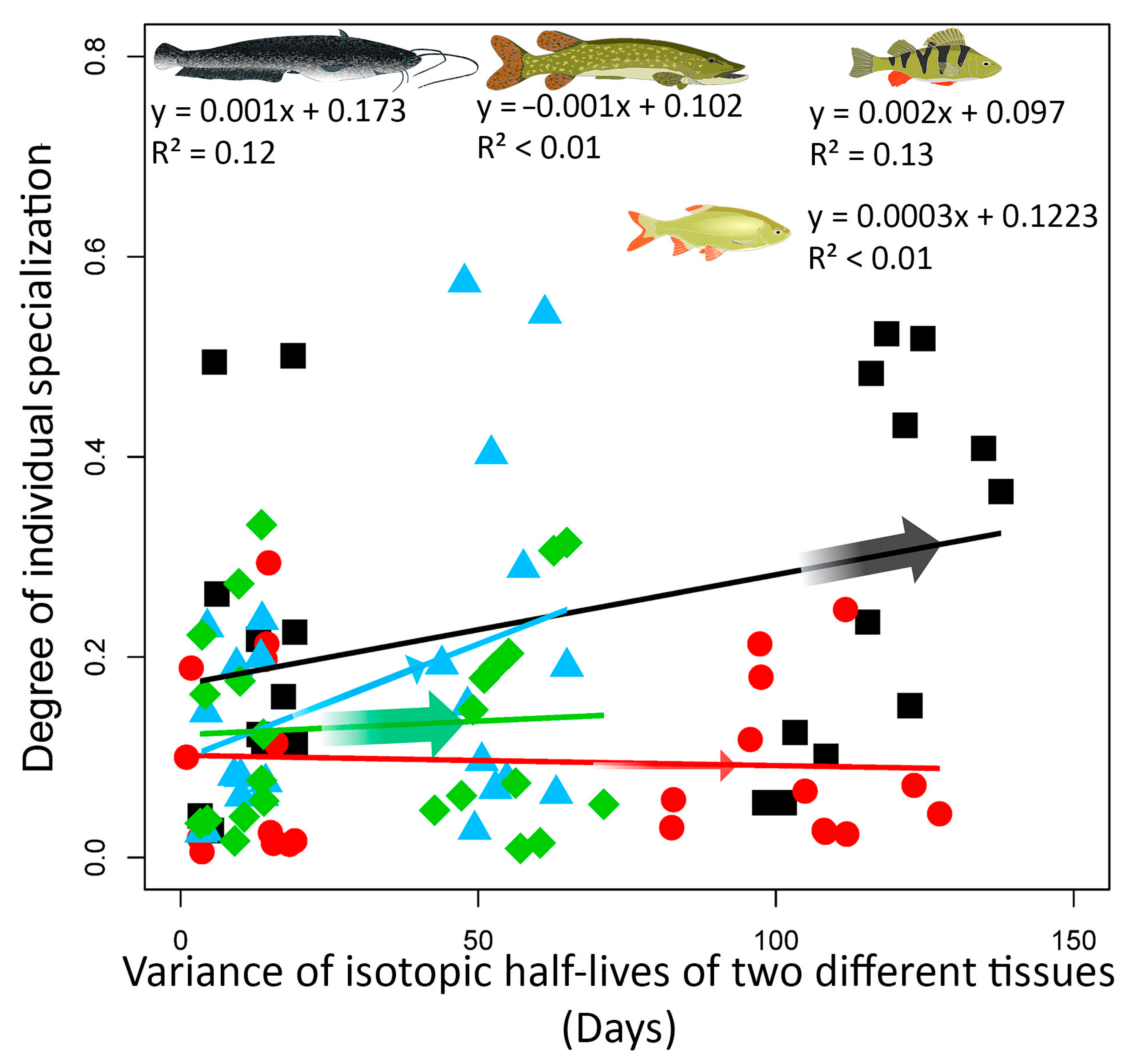
3.3. IIS in Relation to Species Position in the Food Web and Site Trophic State
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Owen-Smith, N.; Fryxell, J.M.; Merrill, E.H. Foraging theory upscaled: The behavioural ecology of herbivore movement. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2267–2278. [Google Scholar]
- DeSantis, L.R.G.; Pardi, M.I.; Du, A.; Greshko, M.A.; Yann, L.T.; Hulbert, R.C.; Louys, J. Global long-term stability of individual dietary specialization in herbivorous mammals. Proc. R. Soc. B 2022, 289, 20211839. [Google Scholar] [CrossRef]
- Van Valen, L. Morphological Variation and Width of Ecological Niche. Am. Nat. 1965, 99, 377–389. [Google Scholar] [CrossRef]
- Urton, E.J.M.; Hobson, K.A. Intrapopulation variation in gray wolf isotope (δ15 N and δ13C) profiles: Implications for the ecology of individuals. Oecologia 2005, 145, 317–326. [Google Scholar]
- Woo, K.J.; Elliott, K.H.; Davidson, M.; Gaston, A.J.; Davoren, G.K. Individual specialization in diet by a generalist marine predator reflects specialization in foraging behaviour. J. Anim. Ecol. 2008, 77, 1082–1091. [Google Scholar] [CrossRef]
- Roberts, P.; Stewart, B.A. Defining the ‘generalist specialist’ niche for Pleistocene Homo sapiens. Nat. Hum. Behav. 2018, 2, 542–550. [Google Scholar] [CrossRef]
- Quevedo, M.; Svanbäck, R.; Eklöv, P. Intrapopulation niche partitioning in a generalist predator limits food web connectivity. Ecology 2009, 90, 2263–2274. [Google Scholar] [CrossRef]
- Matich, P.; Heithaus, M.R.; Layman, C.A. Contrasting patterns of individual specialization and trophic coupling in two marine apex predators. J. Anim. Ecol. 2011, 80, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Vejřík, L.; Vejříková, I.; Blabolil, P.; Eloranta, A.P.; Kočvara, L.; Peterka, J.; Sajdlová, Z.; Chung, S.H.T.; Šmejkal, M.; Kiljunen, M.; et al. European catfish (Silurus glanis) as a freshwater apex predator drives ecosystem via its diet adaptability. Sci. Rep. 2017, 7, 5970. [Google Scholar] [CrossRef] [Green Version]
- Kapuscinski, K.L.; Farrell, J.M.; Wilkinson, M.A. Feeding patterns and population structure of an invasive cyprinid, the rudd Scardinius erythrophthalmus (Cypriniformes, Cyprinidae), in Buffalo Harbor (Lake Erie) and the upper Niagara River. Hydrobiologia 2012, 693, 169–181. [Google Scholar]
- Vejříková, I.; Vejřík, L.; Syväranta, J.; Kiljunen, M.; Čech, M.; Blabolil, P.; Vašek, M.; Sajdlová, Z.; Chung, S.H.T.; Šmejkal, M.; et al. Distribution of herbivorous fish is frozen by low temperature. Sci. Rep. 2016, 6, 39600. [Google Scholar] [CrossRef] [Green Version]
- Bolnick, D.I.; Ingram, T.; Stutz, W.E.; Snowberg, L.; Lau, O.L.; Paull, J. Ecological release from interspecific competition leads to decoupled changes in population and individual niche width. Proc. R. Soc. B Biol. Sci. 2010, 277, 1789–1797. [Google Scholar] [CrossRef]
- Vander Zanden, H.B.; Bjorndal, K.A.; Reich, K.J.; Bolten, A.B. Individual specialists in a generalist population: Results from a long-term stable isotope series. Biol. Lett. 2010, 6, 711–714. [Google Scholar]
- Pyke, G.H.; Pulliam, H.R.; Charnov, E.L. Optimal Foraging: A Selective Review of Theory and Tests. Q. Rev. Biol. 1977, 52, 137–154. [Google Scholar]
- Barabás, G.; D’Andrea, R. The effect of intraspecific variation and heritability on community pattern and robustness. Ecol. Lett. 2016, 19, 977–986. [Google Scholar] [CrossRef]
- Araújo, M.S.; Bolnick, D.I.; Layman, C.A. The ecological causes of individual specialisation. Ecol. Lett. 2011, 14, 948–958. [Google Scholar] [CrossRef]
- Belovsky, G.E.; Jordan, P.A. The time-energy budget of a moose. Theor. Popul. Biol. 1978, 14, 76–104. [Google Scholar] [CrossRef]
- Schoener, T.W. Theory of feeding strategies. Annu. Rev. Ecol. Evol. Syst. 1971, 2, 369–404. [Google Scholar] [CrossRef] [Green Version]
- Sol, D.; Elie, M.; Marcoux, M.; Chrostovsky, E.; Porcher, C.; Lefebvre, L. Ecological mechanisms of a resource polymorphism in Zenaida Doves of Barbados. Ecology 2005, 86, 2397–2407. [Google Scholar] [CrossRef] [Green Version]
- Stephens, D.W.; Krebs, J.R. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Araújo, M.S.; Gonzaga, M.O. Individual specialisation in the hunting wasp Trypoxylon (Trypargilum) albonigrum (Hymenoptera, Crabronidae). Behav. Ecol. Sociobiol. 2007, 61, 1855–1863. [Google Scholar] [CrossRef]
- Svanbäck, R.; Persson, L. Individual diet specialisation, niche width and population dynamics: Implications for trophic polymorphisms. J. Anim. Ecol. 2004, 73, 973–982. [Google Scholar]
- Svanbäck, R.; Eklöv, P.; Fransson, R.; Holmgren, K. Intraspecific competition drives multiple species resource polymorphism in fish communities. Oikos 2008, 117, 114–124. [Google Scholar] [CrossRef]
- Knudsen, R.; Amundsen, P.A.; Primicerio, R.; Klemetsen, A.; Sorensen, P. Contrasting niche-based variation in trophic morphology within Arctic charr populations. Evol. Ecol. Res. 2007, 9, 1005–1021. [Google Scholar]
- Bolnick, D.I.; Amarasekare, P.; Araújo, M.S.; Bürger, R.; Levine, J.; Novak, M.; Rudolf, V.H.; Schreiber, S.J.; Urban, M.C.; Vasseur, D.A.; et al. Why intraspecific trait variation matters in community ecology. Trends Ecol. Evol. 2011, 26, 183–192. [Google Scholar]
- Prati, S.; Henriksen, E.H.; Smalås, A.; Knudsen, R.; Klemetsen, A.; Sánchez-Hernández, J.; Amundsen, P.A. The effect of inter- and intraspecific competition on individual and population niche widths: A four-decade study on two interacting salmonids. Oikos 2021, 130, 1679–1691. [Google Scholar]
- Eklöv, P.; Svanbäck, R. Predation risk influences adaptive morphological variation in fish populations. Am. Nat. 2006, 167, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, P.C.; Stefan, H.G.; Pereira, D.L. Coldwater fish oxythermal habitat in Minnesota lakes: Influence of total phosphorus, July air temperature, and relative depth. Can. J. Fish. Aquat. 2010, 67, 2002–2013. [Google Scholar]
- North, R.P.; North, R.L.; Livingstone, D.M.; Koster, O.; Kipfer, R. Long-term changes in hypoxia and soluble reactive phosphorus in the hypolimnion of a large temperate lake: Consequences of a climate regime shift. Glob. Chang. Biol. 2014, 20, 811–823. [Google Scholar]
- Kerches-Rogeri, N.; Niebuhr, B.B.; Muylaert, R.L.; Mello, M.A.L. Individual specialization in the use of space by frugivorous bats. J. Anim. Ecol. 2020, 89, 2584–2595. [Google Scholar] [CrossRef]
- Sheppard, C.E.; Inger, R.; McDonald, R.A.; Barker, S.; Jackson, A.L.; Thompson, F.J.; Vitikainen, E.I.; Cant, M.A.; Marshall, H.H. Intragroup competition predicts individual foraging specialisation in a group-living mammal. Ecol. Lett. 2018, 21, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Robbins, C.T.; Felicetti, L.A.; Sponheimer, M. The effect of dietary protein quality on nitrogen isotope discrimination in mammals and birds. Oecologia 2005, 144, 534–540. [Google Scholar] [PubMed]
- Hette-Tronquart, N. Isotopic niche is not equal to trophic niche. Ecol. Lett. 2019, 22, 1987–1989. [Google Scholar] [CrossRef] [PubMed]
- Vejřík, L.; Vejříková, I.; Kočvara, L.; Blabolil, P.; Peterka, J.; Sajdlová, Z.; Jůza, T.; Šmejkal, M.; Kolařík, T.; Bartoň, D.; et al. The pros and cons of the invasive freshwater apex predator, European catfish Silurus glanis, and powerful angling technique for its population control. J. Environ. Manag. 2019, 241, 374–382. [Google Scholar]
- Bartoň, D.; Brabec, M.; Sajdlová, Z.; Souza, A.; Duras, J.; Kortan, D.; Blabolil, P.; Vejřík, L.; Kubečka, J.; Šmejkal, M. Hydropeaking causes spatial shifts in a reproducing rheophilic fish. Sci. Total Environ. 2022, 806, 150649. [Google Scholar] [CrossRef] [PubMed]
- Vejříková, I.; Eloranta, A.P.; Vejřík, L.; Šmejkal, M.; Čech, M.; Sajdlová, Z.; Frouzova, J.; Kiljunen, M.; Peterka, J. Macrophytes shape trophic niche variation among generalist fishes. PLoS ONE 2017, 12, e0177114. [Google Scholar] [CrossRef]
- Stephen, M.T.; Crowther, T.W. Predicting rates of isotopic turnover across the animal kingdom: A synthesis of existing data. J. Anim. Ecol. 2015, 84, 861–870. [Google Scholar]
- Vašek, M.; Vejřík, L.; Vejříková, I.; Šmejkal, M.; Baran, R.; Muška, M.; Kubečka, J.; Peterka, J. Development of non-lethal monitoring of stable isotopes in asp (Leuciscus aspius): A comparison of muscle, fin and scale tissues. Hydrobiology 2017, 785, 327–335. [Google Scholar]
- France, R.L. Critical examination of stable isotope analysis as a means for tracing carbon pathways in stream ecosystems. Can. J. Fish. Aquat. 1995, 52, 651–656. [Google Scholar]
- Anderson, C.; Cabana, G. Estimating the trophic position of aquatic consumers in river food webs using stable nitrogen isotopes. J. N. Am. Benthol. Soc. 2007, 26, 273–285. [Google Scholar]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar]
- Vander Zanden, M.J.; Shuter, B.J.; Lester, N.; Rasmussen, J.B. Patterns of food chain length in lakes: A stable isotope study. Am. Nat. 1999, 154, 406–416. [Google Scholar] [PubMed]
- Olsson, K.; Stenroth, P.; Nyström, P.; Granéli, W. Invasions and niche width: Does niche width of an introduced crayfish differ from a native crayfish? Freshw. Biol. 2009, 54, 1731–1740. [Google Scholar]
- Layman, C.A.; Arrington, D.A.; Montaña, C.G.; Post, D.M. Can stable isotope ratios provide quantitative measures of trophic diversity within food webs? Ecology 2007, 88, 42–48. [Google Scholar]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER – Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar]
- Bolnick, D.I.; Svanbäck, R.; Fordyce, J.A.; Yang, L.H.; Davis, J.M.; Hulsey, C.D.; Forister, M.L. The Ecology of Individuals: Incidence and Implications of Individual Specialization. Am. Nat. 2003, 161, 1–28. [Google Scholar]
- Bolnick, D.I. Ind Spec1; Center for Population Biology Store Hall University of California: Davis, CA, USA, 2002. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 7 August 2023).
- Allouche, O.; Kalyuzhny, M.; Moreno-Rueda, G.; Pizarro, M.; Kadmon, K. Area–heterogeneity tradeoff and the diversity of ecological communities. Proc. Natl. Acad. Sci. USA 2012, 109, 17495–17500. [Google Scholar] [CrossRef] [PubMed]
- Forsman, A.; Tibblin, P.; Berggren, H.; Nordahl, O.; Koch-Smidth, P.; Larsson, P. Pike Esox lucius as an emerging model organism for studies in ecology and evolutionary biology: A review. J. Fish. Biol. 2015, 87, 472–479. [Google Scholar]
- Prugh, R.L.; Stoner, C.J.; Epps, C.W.; Bean, W.T.; Ripple, W.J.; Laliberte, A.S.; Brashares, J.S. The rise of the mesopredator. BioScience 2009, 59, 779–791. [Google Scholar]
- Ripple, W.J.; Estes, J.; Beschta, R.; Wilmers, C.; Ritchie, E.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 1241484. [Google Scholar]
- Kozlowski, J.; Uchmanski, J. Optimal individual growth and reproduction in perennial species with indeterminate growth. Evol. Ecol. 1987, 1, 214–230. [Google Scholar] [CrossRef]
- Brooker, R.M.; Munday, P.L.; Brandl, S.J.; Jones, G.P. Local extinction of a coral reef fish explained by inflexible prey choice. Coral Reefs 2014, 33, 891–896. [Google Scholar] [CrossRef]
- Orr, H.A.; Unckless, R.L. Population extinction and the genetics of adaptation. Am. Nat. 2008, 172, 160–169. [Google Scholar]
- Foote, A.D.; Newton, J.; Piertney, S.B.; Willerslev, E.; Gilbert, M.T.P. Ecological, morphological and genetic divergence of sympatric North Atlantic killer whale populations. Mol. Ecol. 2009, 18, 5207–5217. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.E.; Gilliam, J.F. The ontogenetic niche and species interactions in size-structured populations. Annu. Rev. Ecol. Evol. Syst. 1984, 15, 393–425. [Google Scholar] [CrossRef]
- Rosenblatt, A.E.; Nifong, J.C.; Heithaus, M.R.; Mazzotti, F.J.; Cherkiss, M.S.; Jeffery, B.M.; Elsey, R.M.; Decker, R.A.; Silliman, B.R.; Guillette, L.J. Factors affecting individual foraging specialization and temporal diet stability across the range of a large “generalist” apex predator. Oecologia 2015, 178, 5–16. [Google Scholar]
- Nurminen, L.; Horppila, J.; Lappalainen, J.; Malinen, T. Implications of rudd (Scardinius erythrophthalmus) herbivory on submerged macrophytes in a shallow eutrophic lake. Hydrobiology 2003, 506–509, 511–518. [Google Scholar] [CrossRef]
- Elton, C.S. Animal Ecology; Sidgewick & Jackson: London, UK, 1927. [Google Scholar]
- Matthews, B.; Marchinko, K.B.; Bolnick, D.I.; Mazumder, A. Specialisation of trophic position and habitat use by sticklebacks in an adaptive radiation. Ecology 2010, 91, 1025–1034. [Google Scholar]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef]
- Heady, W.N.; Moore, J.W. Tissue turnover and stable isotope clocks to quantify resource shifts in anadromous rainbow trout. Oecologia 2013, 172, 21–34. [Google Scholar]
- Wakefield, E.D.; Cleasby, I.R.; Bearhop, S.; Bodey, T.W.; Davies, R.D.; Miller, P.I.; Newton, J.; Votier, S.C.; Hamer, K.C. Long-term individual foraging site fidelity—Why some gannets don’t change their spots. Ecology 2015, 96, 3058–3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Říha, M.; Gjelland, K.O.; Děd, V.; Eloranta, A.P.; Rabaneda Bueno, R.; Baktoft, H.; Vejřík, L.; Vejříková, I.; Draštík, V.; Šmejkal, M.; et al. Contrasting structural complexity differentiate hunting strategy in an ambush apex predator. Sci. Rep. 2021, 11, 17472. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Area (ha) | Volume (106 m3) | Long-Term Flow (m3/s) | Detention Time (Days) | Max. Depth | Altitude (m a.s.l) | Trophic Status, TP (μg L−1) |
|---|---|---|---|---|---|---|---|
| 1. Most | 311 | 70 | 0.06 | Drainless | 75 | 199 | 2.5 |
| 2. Milada | 250 | 36 | 0.04 | 10,248 | 25 | 145 | 5 |
| 3. Žlutice | 161 | 16 | 1.24 | 146 | 23 | 508 | 25.8 |
| 4. Římov | 210 | 34 | 4.38 | 93 | 45 | 468 | 26.4 |
| Species | Site | N | Mass (kg) | ITNW (SEAc) | IINW (SEAc) | IIS |
|---|---|---|---|---|---|---|
| Catfish | 1 | 16 | 2.4–14.5 (5.1 ± 2.78) | 0.105 | 0.011 (±0.008) | 0.1902 (±0.151) |
| 2 | 16 | 2–13.8 (7.87 ± 3.56) | 0.141 | 0.017 (±0.012) | 0.1609 (±0.120) | |
| 3 | 18 | 1.6–21 (9.69 ± 5.84) | 0.296 | 0.033 (±0.041) | 0.6673 (±0.283) | |
| 4 | 15 | 3.7–21.8 (10.59 ± 5.90) | 0.173 | 0.041 (±0.048) | 0.6310 (±0.268) | |
| Pike | 1 | 12 | 0.6–7.5 (4.7 ± 2.13) | 0.105 | 0.006 (±0.003) | 0.0554 (±0.033) |
| 2 | 14 | 0.7–10.6 (5.06 ± 3.09) | 0.103 | 0.013 (±0.010) | 0.0314 (±0.024) | |
| 3 | 13 | 0.8–5.9 (1.85 ± 1.58) | 0.077 | 0.008 (±0.007) | 0.2649 (±0.227) | |
| 4 | 15 | 0.7–13.8 (3.4 ± 3.65) | 0.022 | 0.017 (±0.022) | 0.1511 (±0.156) | |
| Perch | 1 | 15 | 0.18–1.9 (0.51 ± 0.43) | 0.102 | 0.011 (±0.009) | 0.0771 (±0.058) |
| 2 | 15 | 0.15–0.66 (0.33 ± 0.13) | 0.078 | 0.021 (±0.013) | 0.2767 (±0.185) | |
| 3 | 16 | 0.2–0.52 (0.39 ± 0.09) | 0.042 | 0.016 (±0.010) | 0.1334 (±0.085) | |
| 4 | 11 | 0.2–0.45 (0.33 ± 0.09) | 0.102 | 0.032 (±0.016) | 0.442 (±0.170) | |
| Rudd | 1 | 15 | 0.16–0.81 (0.31 ± 0.18) | 0.071 | 0.007(±0.009) | 0.0789 (±0.111) |
| 2 | 15 | 0.2–1.29 (0.48 ± 0.33) | 0.187 | 0.021 (±0.013) | 0.3449 (±0.232) | |
| 3 | 11 | 0.3–1.02 (0.58 ± 0.23) | 0.082 | 0.007 (±0.006) | 0.0495 (±0.0439) | |
| 4 | 12 | 0.27–0.5 (0.40 ± 0.07) | 0.045 | 0.023 (±0.016) | 0.3245 (±0.240) |
| Species | ANOVA | Study Site Comparisons, Post-Hoc Tukey Test | |||||
|---|---|---|---|---|---|---|---|
| 1 vs. 2 | 1 vs. 3 | 1 vs. 4 | 2 vs. 3 | 2 vs. 4 | 3 vs. 4 | ||
| Catfish | (3.61) = 25.69, p < 0.001 | p > 0.05 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p > 0.05 |
| Pike | (3.50) = 10.85, p < 0.001 | p > 0.05 | p < 0.01 | p < 0.01 | p < 0.001 | p < 0.001 | p > 0.05 |
| Perch | (3.53) = 19.34, p < 0.001 | p < 0.001 | p > 0.05 | p < 0.001 | p < 0.05 | p < 0.05 | p < 0.001 |
| Rudd | (3.50) = 10.12, p < 0.001 | p < 0.001 | p > 0.05 | p < 0.01 | p < 0.001 | p > 0.05 | p < 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vejřík, L.; Vejříková, I.; Blabolil, P.; Sajdlová, Z.; Kočvara, L.; Kolařík, T.; Bartoň, D.; Jůza, T.; Šmejkal, M.; Peterka, J.; et al. Trophic Position of the Species and Site Trophic State Affect Diet Niche and Individual Specialization: From Apex Predator to Herbivore. Biology 2023, 12, 1113. https://doi.org/10.3390/biology12081113
Vejřík L, Vejříková I, Blabolil P, Sajdlová Z, Kočvara L, Kolařík T, Bartoň D, Jůza T, Šmejkal M, Peterka J, et al. Trophic Position of the Species and Site Trophic State Affect Diet Niche and Individual Specialization: From Apex Predator to Herbivore. Biology. 2023; 12(8):1113. https://doi.org/10.3390/biology12081113
Chicago/Turabian StyleVejřík, Lukáš, Ivana Vejříková, Petr Blabolil, Zuzana Sajdlová, Luboš Kočvara, Tomáš Kolařík, Daniel Bartoň, Tomáš Jůza, Marek Šmejkal, Jiří Peterka, and et al. 2023. "Trophic Position of the Species and Site Trophic State Affect Diet Niche and Individual Specialization: From Apex Predator to Herbivore" Biology 12, no. 8: 1113. https://doi.org/10.3390/biology12081113
APA StyleVejřík, L., Vejříková, I., Blabolil, P., Sajdlová, Z., Kočvara, L., Kolařík, T., Bartoň, D., Jůza, T., Šmejkal, M., Peterka, J., & Čech, M. (2023). Trophic Position of the Species and Site Trophic State Affect Diet Niche and Individual Specialization: From Apex Predator to Herbivore. Biology, 12(8), 1113. https://doi.org/10.3390/biology12081113