


Functional Cardiovascular Characterization of the Common Marmoset (Callithrix jacchus)
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Infant | Juvenile | Adolescent | Adult | Aged |
|---|---|---|---|---|---|
| Human [29,30] | <12 m | 1–12 y | 12–18 y | 18–65 y | >65 y |
| Marmoset [31] | <3 m | 3–5 m | 0.5–2 y | 2–8 y | >8 y |
| Pig [32] | <2.5 m | 2.5–4.5 m | 4.5–6.5 m | >6.5 m | |
| Mouse [33] | <3 w | 3–4 w | 1–3 m | 3–20 m | >20 m |
2. Materials and Methods
2.1. Ethics
2.2. Animals, Husbandry and Housing
2.3. Study Design
2.4. MRI
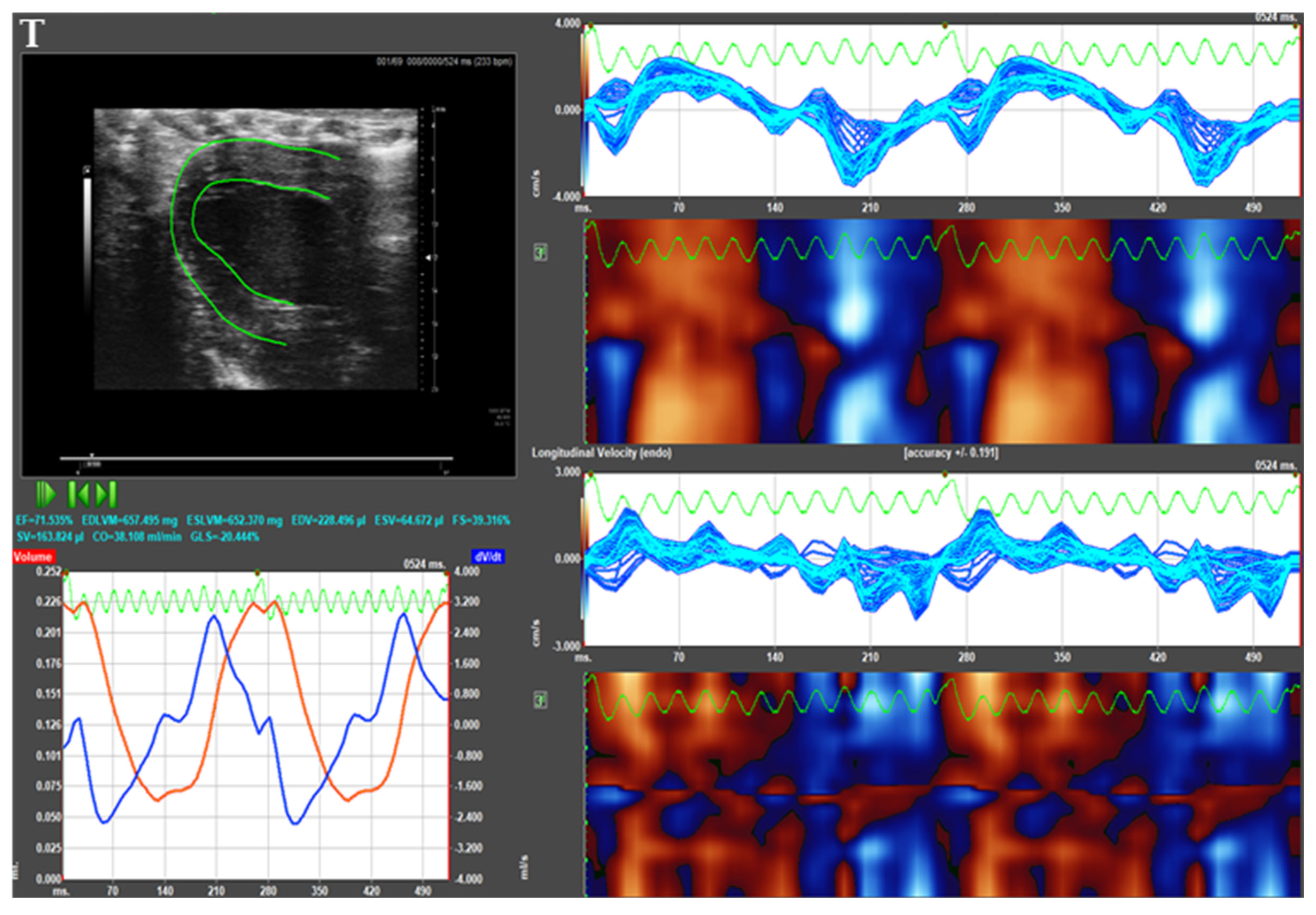
2.5. Echocardiography
2.6. PV Loop Measurement
2.6.1. Surgery for Hemodynamic Measurement Intervention
2.6.2. Hemodynamic Measurement
- (a)
- Blood and heart muscle resistivity were determined for each animal using a calibration probe (5.0 mm, Model FM-1287-IM, Transonic Europe B.V., Elsloo, The Netherlands). For measuring blood resistivity, ~0.2 mL blood was drawn from the femoral vein just before the intervention and added to a 1.5 mL Eppendorf tube. The calibration probe was immediately introduced to the blood to acquire values for resistivity. For determination of the heart muscle resistivity, the calibration catheter was positioned directly on the heart surface avoiding coronary arteries, before inserting the cannula as described above. All readouts were directly saved into the ADV500 system.
- (b)
- The pressure–volume catheter was presoaked in body-warm saline in a 1 mL syringe on the level of the animal’s heart 20 min prior to the measurement to adjust to 37 °C fluid. After zeroing pressure offset, the catheter was inserted into the ventricle lumen.
2.6.3. Analysis of PV Loop Data
2.7. Statistics
3. Results
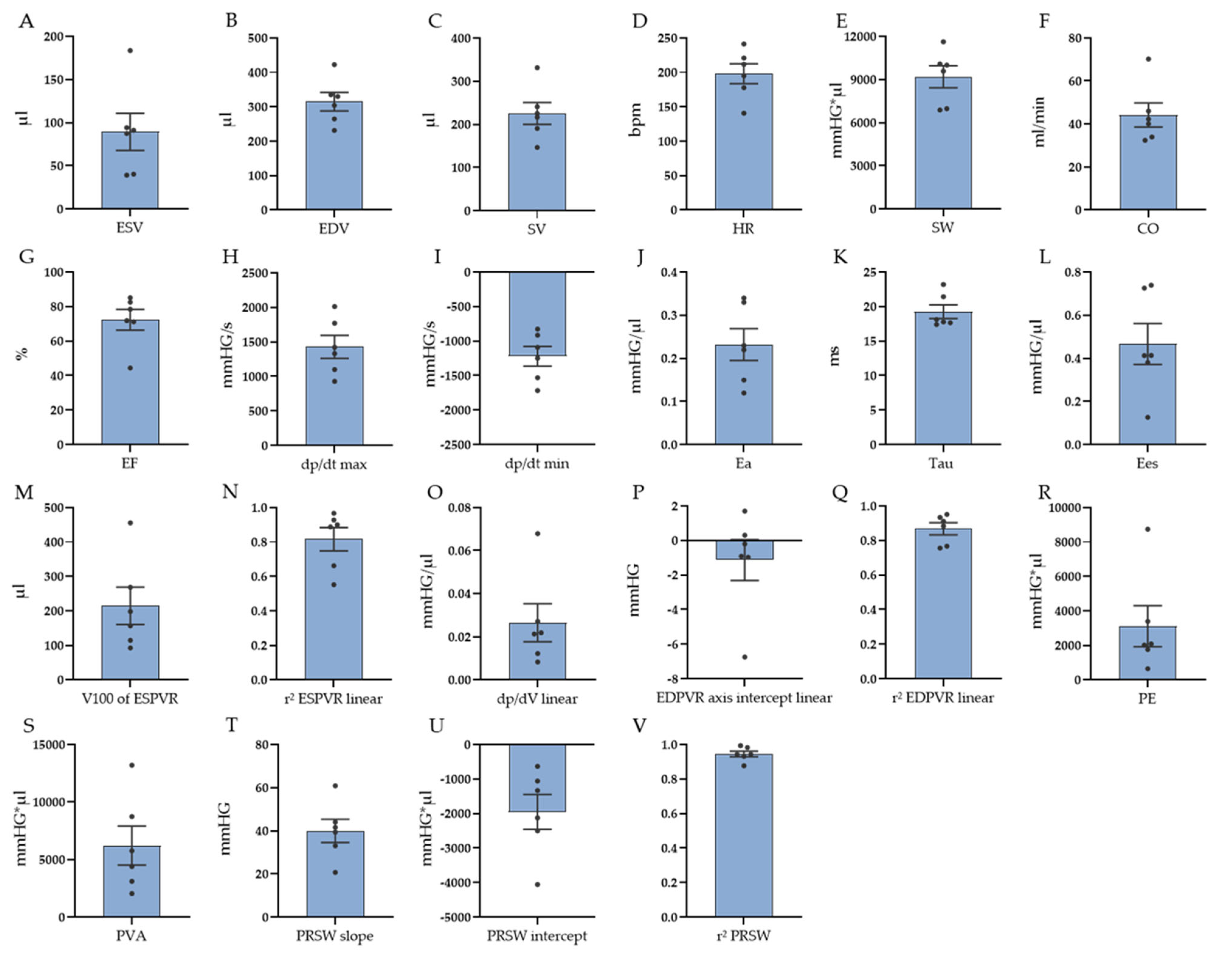
3.1. PV Loop Measurements
3.2. MRI Measurements
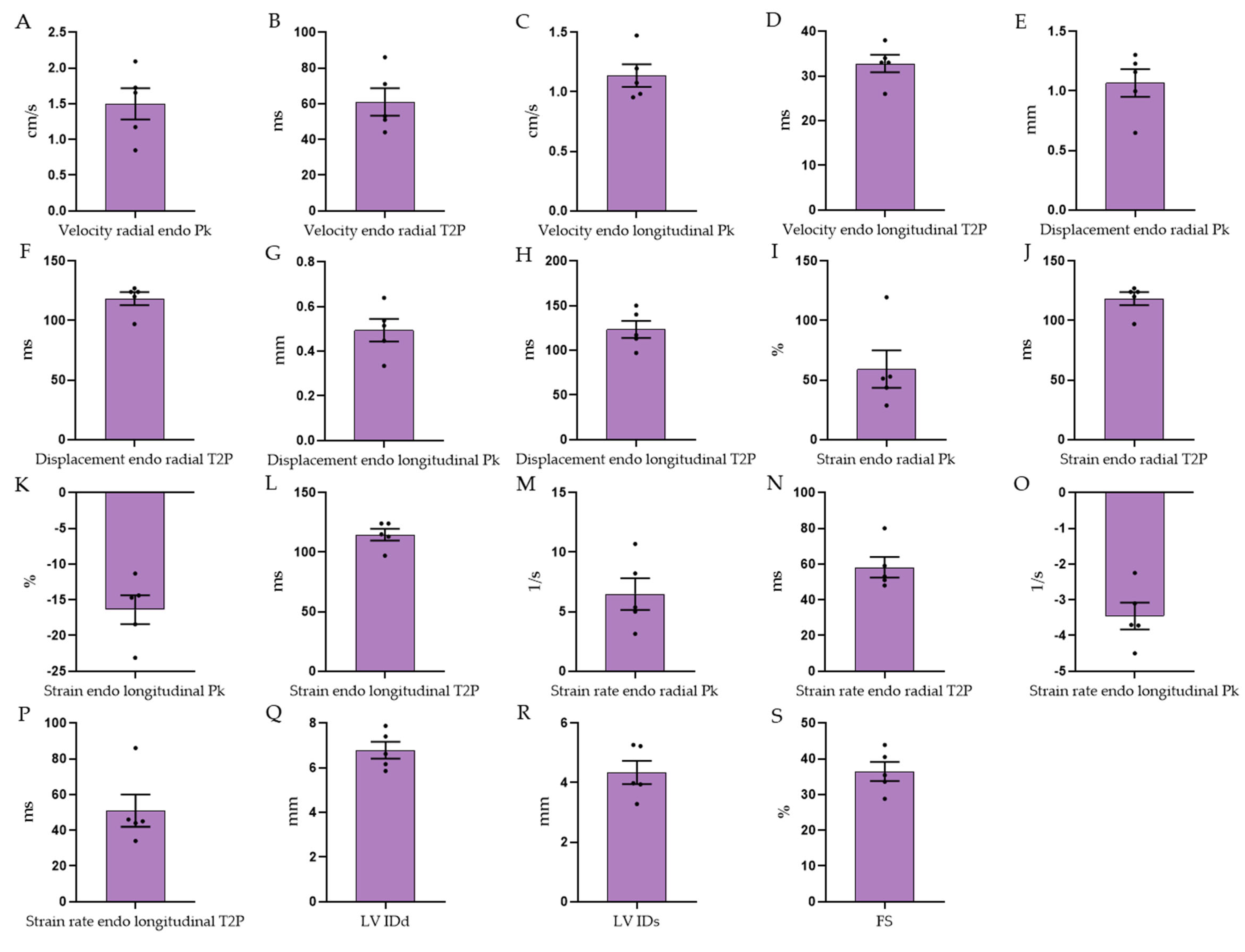
3.3. Echocardiographic Measurements
3.4. Comparison of PV Loops with MRI and Echocardiography
4. Discussion
4.1. Study Design
4.2. PV Loop Data in the Context of Reported Cardiovascular Data for the Common Marmoset
4.3. Comparison of PV Loop Measurements with Echocardiography and MRI
4.4. Considerations for PV Loop Measurements in Common Marmosets
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| a | constant of curvilinearity |
| b | constant of square regression |
| bpm | beats per minute |
| BW | body weight |
| CO | cardiac output |
| dP/dt max | maximum rate of pressure change |
| dP/dt min | minimum rate of diastolic pressure change |
| dp/dV | dp/dV = slope of EDPVR and index of stiffness |
| Ea | arterial elastance |
| EDPVR | end-diastolic pressure–volume relationship |
| EDV | end-diastolic volume |
| EF | ejection fraction |
| Ees | end-systolic elastance |
| ESPVR | end-systolic pressure–volume relationship |
| ESV | end-systolic volume |
| HR | heart rate |
| k2 | constant |
| mmHg | millimeter mercury |
| MRI | magnetic resonance imaging |
| PE | potential energy |
| PRSW | preload recruitable stroke work |
| PVA | pressure–volume area |
| PV loop | pressure–volume loop |
| r2 | coefficient of determination |
| SV | stroke volume |
| SW | stroke work |
| Tau | isovolumic relaxation constant |
Appendix A. Additional Tips and Tricks/Considerations for Performing Pressure–Volume Measurements in the Common Marmoset
References
- World Health Organization. Cardiovascular Diseases (CVDs) Fact Sheet, Letzte Aktualisierung 17 May 2017. Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 12 December 2020).
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Alonso, A.; Beaton, A.Z.; Bittencourt, M.S.; Boehme, A.K.; Buxton, A.E.; Carson, A.P.; Commodore-Mensah, Y.; et al. Heart Disease and Stroke Statistics-2022 Update: A Report From the American Heart Association. Circulation 2022, 145, e153–e639. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Hashim, M.J.; Mustafa, H.; Baniyas, M.Y.; Al Suwaidi, S.; AlKatheeri, R.; Alblooshi, F.M.K.; Almatrooshi, M.; Alzaabi, M.E.H.; Al Darmaki, R.S.; et al. Global Epidemiology of Ischemic Heart Disease: Results from the Global Burden of Disease Study. Cureus 2020, 12, e9349. [Google Scholar] [CrossRef]
- Richter, C.; Hinkel, R. Research(’s) Sweet Hearts: Experimental Biomedical Models of Diabetic Cardiomyopathy. Front. Cardiovasc. Med. 2021, 8, 703355. [Google Scholar] [CrossRef] [PubMed]
- Pilz, P.M.; Ward, J.E.; Chang, W.T.; Kiss, A.; Bateh, E.; Jha, A.; Fisch, S.; Podesser, B.K.; Liao, R. Large and Small Animal Models of Heart Failure with Reduced Ejection Fraction. Circ. Res. 2022, 130, 1888–1905. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.A.S.; Emter, C.A. Large Animal Models of Heart Failure: A Translational Bridge to Clinical Success. JACC. Basic Transl. Sci. 2020, 5, 840–856. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K. Marmoset models commonly used in biomedical research. Comp. Med. 2003, 53, 383–392. [Google Scholar] [PubMed]
- Carrion, J.R.; Patterson, J.L. An animal model that reflects human disease: The common marmoset (Callithrix jacchus). Curr. Opin. Virol. 2012, 2, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Okano, H.; Hikishima, K.; Iriki, A.; Sasaki, E. The common marmoset as a novel animal model system for biomedical and neuroscience research applications. Semin. Fetal Neonatal Med. 2012, 17, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Trennery, P.; Farningham, D.; Klapwijk, J. The selection of marmoset monkeys (Callithrix jacchus) in pharmaceutical toxicology. Lab. Anim. 2001, 35, 117–130. [Google Scholar] [CrossRef]
- Tardif, S.D.; Mansfield, K.G.; Ratnam, R.; Ross, C.N.; Ziegler, T.E. The marmoset as a model of aging and age-related diseases. ILAR J. 2011, 52, 54–65. [Google Scholar] [CrossRef]
- Petkov, S.; Dressel, R.; Rodriguez-Polo, I.; Behr, R. Controlling the Switch from Neurogenesis to Pluripotency during Marmoset Monkey Somatic Cell Reprogramming with Self-Replicating mRNAs and Small Molecules. Cells 2020, 9, 2422. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.M.; Ross, C.N.; Bliss-Moreau, E. Noninvasive cardiac psychophysiology as a tool for translational science with marmosets. Am. J. Primatol. 2019, 81, e23037. [Google Scholar] [CrossRef] [PubMed]
- Tardif, S.D.; Power, M.L.; Ross, C.N.; Rutherford, J.N.; Layne-Colon, D.G.; Paulik, M.A. Characterization of obese phenotypes in a small nonhuman primate, the common marmoset (Callithrix jacchus). Obesity 2009, 17, 1499–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, C.N.; Davis, K.; Dobek, G.; Tardif, S.D. Aging Phenotypes of Common Marmosets (Callithrix jacchus). J. Aging Res. 2012, 2012, 567143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, E.; Suemizu, H.; Shimada, A.; Hanazawa, K.; Oiwa, R.; Kamioka, M.; Tomioka, I.; Sotomaru, Y.; Hirakawa, R.; Eto, T.; et al. Generation of transgenic non-human primates with germline transmission. Nature 2009, 459, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Mietsch, M.; Paqué, K.; Drummer, C.; Stahl-Hennig, C.; Roshani, B. The aging common marmoset’s immune system: From junior to senior. Am. J. Primatol. 2020, 82, e23128. [Google Scholar] [CrossRef] [PubMed]
- Mietsch, M.; Baldauf, K.; Reitemeier, S.; Suchowski, M.; Schoon, H.A.; Einspanier, A. Blood pressure as prognostic marker for body condition, cardiovascular, and metabolic diseases in the common marmoset (Callithrix jacchus). J. Med. Primatol. 2016, 45, 126–138. [Google Scholar] [CrossRef]
- Falcão, B.M.R.; de Souza, J.G.; de La Salles, A.Y.F.; Batista, L.N.; Rocha, E.F.; Diniz, J.; Fernandes, A.R.F.; Dos Santos, J.R.S.; de Menezes, D.J.A.; Medeiros, G.X. Heart anatomy and topography of the common marmoset (Callithrix jacchus Linnaeus, 1758). J. Med. Primatol. 2020, 49, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Senos, R.; Gomes Benedicto, H.; Macedo Del Rio do Valle, C.; Del Rio do Valle, R.; Nayudu, P.L.; Roberto-Rodrigues, M.; Bombonato, P.P. Collagen quantification in the ventricular walls of the heart of the common marmoset (Callithrix jacchus). Anat. Rec. 2021, 304, 1275–1279. [Google Scholar] [CrossRef]
- Senos, R.; Benedicto, H.G.; del Rio do Valle, C.M.; del Rio do Valle, R.; Nayudu, P.L.; Kfoury Junior, J.R.; Bombonato, P.P. Gross morphometry of the heart of the Common marmoset. Folia Morphol. 2014, 73, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Moussavi, A.; Mietsch, M.; Drummer, C.; Behr, R.; Mylius, J.; Boretius, S. Cardiac MRI in common marmosets revealing age-dependency of cardiac function. Sci. Rep. 2020, 10, 10221. [Google Scholar] [CrossRef]
- Townsend, D. Measuring Pressure Volume Loops in the Mouse. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, N.; Abdou, H.; Edwards, J.; Elansary, N.N.; Poe, K.; Richmond, M.J.; Madurska, M.J.; Rasmussen, T.E.; Morrison, J.J. Measuring Cardiac Output in a Swine Model. J. Vis. Exp. 2021, e62333. [Google Scholar] [CrossRef]
- Kelly, K.M.; Tarwater, P.M.; Karper, J.M.; Bedja, D.; Queen, S.E.; Tunin, R.S.; Adams, R.J.; Kass, D.A.; Mankowski, J.L. Diastolic dysfunction is associated with myocardial viral load in simian immunodeficiency virus-infected macaques. Aids 2012, 26, 815–823. [Google Scholar] [CrossRef]
- Pinsky, M.R.; Perlini, S.; Solda, P.L.; Pantaleo, P.; Calciati, A.; Bernardi, L. Dynamic right and left ventricular interactions in the rabbit: Simultaneous measurement of ventricular pressure-volume loops. J. Crit. Care 1996, 11, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Lips, D.J.; van der Nagel, T.; Steendijk, P.; Palmen, M.; Janssen, B.J.; van Dantzig, J.M.; de Windt, L.J.; Doevendans, P.A. Left ventricular pressure-volume measurements in mice: Comparison of closed-chest versus open-chest approach. Basic Res. Cardiol. 2004, 99, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Van Hout, G.P.; Jansen of Lorkeers, S.J.; Gho, J.M.; Doevendans, P.A.; van Solinge, W.W.; Pasterkamp, G.; Chamuleau, S.A.; Hoefer, I.E. Admittance-based pressure-volume loops versus gold standard cardiac magnetic resonance imaging in a porcine model of myocardial infarction. Physiol. Rep. 2014, 2, e00287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasundaram, P.; Avulakunta, I.D. Human Growth and Development. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- NIH. NIH Style Guide: Age. Available online: https://www.nih.gov/nih-style-guide/age (accessed on 8 May 2023).
- Abbott, D.H.; Barnett, D.K.; Colman, R.J.; Yamamoto, M.E.; Schultz-Darken, N.J. Aspects of common marmoset basic biology and life history important for biomedical research. Comp. Med. 2003, 53, 339–350. [Google Scholar] [PubMed]
- Wilson, J.B.; Pauling, G.E.; McEwen, B.J.; Smart, N.; Carman, P.S.; Dick, C.P. A descriptive study of the frequency and characteristics of proliferative enteropathy in swine in Ontario by analyzing routine animal health surveillance data. Can. Vet. J. 1999, 40, 713–717. [Google Scholar]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Heiberg, E.; Sjögren, J.; Ugander, M.; Carlsson, M.; Engblom, H.; Arheden, H. Design and validation of Segment—Freely available software for cardiovascular image analysis. BMC Med. Imaging 2010, 10, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, R.M.; Bierig, M.; Devereux, R.B.; Flachskampf, F.A.; Foster, E.; Pellikka, P.A.; Picard, M.H.; Roman, M.J.; Seward, J.; Shanewise, J.S.; et al. Recommendations for chamber quantification: A report from the American Society of Echocardiography’s Guidelines and Standards Committee and the Chamber Quantification Writing Group, developed in conjunction with the European Association of Echocardiography, a branch of the European Society of Cardiology. J Am Soc. Echocardiogr. 2005, 18, 1440–1463. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.S.; Hartig, R.; Basso, M.A.; Jarrett, W.; Kastner, S.; Poirier, C. International primate neuroscience research regulation, public engagement and transparency opportunities. Neuroimage 2021, 229, 117700. [Google Scholar] [CrossRef]
- Suzuki, M.; Ogata, M.; Murata, Y.; Suzuki, S. Presence of the annular ridge and location of the coronary sinus ostium as morphological features ofthe right atrium and interatrial septum in healthy common marmosets (Callithrix jacchus). Exp. Anim. 2020, 69, 295–305. [Google Scholar] [CrossRef]
- Jia, T.; Wang, C.; Han, Z.; Wang, X.; Ding, M.; Wang, Q. Experimental Rodent Models of Cardiovascular Diseases. Front. Cardiovasc. Med. 2020, 7, 588075. [Google Scholar] [CrossRef]
- Neumann, B.; Shi, T.; Gan, L.L.; Klippert, A.; Daskalaki, M.; Stolte-Leeb, N.; Stahl-Hennig, C. Comprehensive panel of cross-reacting monoclonal antibodies for analysis of different immune cells and their distribution in the common marmoset (Callithrix jacchus). J. Med. Primatol. 2016, 45, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Burity, C.H.; Mandarim-de-Lacerda, C.A.; Pissinatti, A. Stereology of the myocardium in two species of Callithrix (Callitrichidae, primates). Ann. Anat. 1996, 178, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Kurata, Y.; Makinodan, F.; Kidachi, F.; Yokoyama, M.; Wako, Y.; Yamagishi, Y.; Katsuta, O.; Takechi, M.; Tsuchitani, M. Spontaneous lesions detected in the common cotton-eared marmosets (Callithrix jacchus). J. Vet. Med. Sci. 1996, 58, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Chamanza, R.; Parry, N.M.; Rogerson, P.; Nicol, J.R.; Bradley, A.E. Spontaneous lesions of the cardiovascular system in purpose-bred laboratory nonhuman primates. Toxicol. Pathol. 2006, 34, 357–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, C.N.; Adams, J.; Gonzalez, O.; Dick, E.; Giavedoni, L.; Hodara, V.L.; Phillips, K.; Rigodanzo, A.D.; Kasinath, B.; Tardif, S.D. Cross-sectional comparison of health-span phenotypes in young versus geriatric marmosets. Am. J. Primatol. 2019, 81, e22952. [Google Scholar] [CrossRef]
- Ohta, H.; Komatsu, T.; Muta, K.; Koizumi, M.; Iguchi, Y.; Okano, H.J. Marmoset angiography just by percutaneous puncture of the caudal ventral artery. PLoS ONE 2021, 16, e0250576. [Google Scholar] [CrossRef] [PubMed]
- McLennan, P.L.; Barnden, L.R.; Bridle, T.M.; Abeywardena, M.Y.; Charnock, J.S. Dietary fat modulation of left ventricular ejection fraction in the marmoset due to enhanced filling. Cardiovasc. Res. 1992, 26, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Hanton, G. Echocardiography in marmosets: A non-invasive method for the assessment of cardiovascular toxicology. In Novel Approaches towards Primate Toxicology, 1st ed.; Weinbauer, G.F., Vogel, F., Eds.; Waxmann: Münster, Germany, 2006; pp. 681–696. [Google Scholar]
- Locquet, L.; Houdellier, B.; Broeckx, B.J.G.; Bouts, T.; Liekens, V.; Saunders, J.H.; Smets, P. Transthoracic echocardiography and cardiac biomarkers in healthy captive male and female squirrel monkeys (Saimiri spp.). BMC Vet. Res. 2020, 16, 217. [Google Scholar] [CrossRef] [PubMed]
- Huss, M.K.; Ikeno, F.; Buckmaster, C.L.; Albertelli, M.A. Echocardiographic and electrocardiographic characteristics of male and female squirrel monkeys (Saimiri spp.). J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 25–28. [Google Scholar] [PubMed]
- Riegler, J.; Cheung, K.K.; Man, Y.F.; Cleary, J.O.; Price, A.N.; Lythgoe, M.F. Comparison of segmentation methods for MRI measurement of cardiac function in rats. J. Magn. Reson. Imaging 2010, 32, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Stuckey, D.J.; Carr, C.A.; Tyler, D.J.; Clarke, K. Cine-MRI versus two-dimensional echocardiography to measure in vivo left ventricular function in rat heart. NMR Biomed. 2008, 21, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Quill, G.M.; Gilbert, K.; Wang, V.Y.; Houle, H.C.; Legget, M.E.; Ruygrok, P.N.; Doughty, R.N.; Pedrosa, J.; D’Hooge, J.; et al. Systematic Comparison of Left Ventricular Geometry Between 3D-Echocardiography and Cardiac Magnetic Resonance Imaging. Front. Cardiovasc. Med. 2021, 8, 728205. [Google Scholar] [CrossRef]
- Raake, P.W.J.; Barthelmes, J.; Krautz, B.; Buss, S.; Huditz, R.; Schlegel, P.; Weber, C.; Stangassinger, M.; Haberkorn, U.; Katus, H.A.; et al. Comprehensive cardiac phenotyping in large animals: Comparison of pressure-volume analysis and cardiac magnetic resonance imaging in pig post-myocardial infarction systolic heart failure. Int. J. Cardiovasc. Imaging 2019, 35, 1691–1699. [Google Scholar] [CrossRef]
- Chen, C.H.; Nevo, E.; Fetics, B.; Nakayama, M.; Pak, P.H.; Maughan, W.L.; Kass, D.A. Comparison of continuous left ventricular volumes by transthoracic two-dimensional digital echo quantification with simultaneous conductance catheter measurements in patients with cardiac diseases. Am. J. Cardiol. 1997, 80, 756–761. [Google Scholar] [CrossRef]
- Herberg, U.; Linden, K.; Dewald, O.; Gatzweiler, E.; Seehase, M.; Duerr, G.D.; Dörner, J.; Kleppe, S.; Ladage, D.; Breuer, J. 3D Real-Time Echocardiography Combined with Mini Pressure Wire Generate Reliable Pressure-Volume Loops in Small Hearts. PLoS ONE 2016, 11, e0165397. [Google Scholar] [CrossRef] [Green Version]
- Grune, J.; Blumrich, A.; Brix, S.; Jeuthe, S.; Drescher, C.; Grune, T.; Foryst-Ludwig, A.; Messroghli, D.; Kuebler, W.M.; Ott, C.; et al. Evaluation of a commercial multi-dimensional echocardiography technique for ventricular volumetry in small animals. Cardiovasc. Ultrasound 2018, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- Russo, I.; Micotti, E.; Fumagalli, F.; Magnoli, M.; Ristagno, G.; Latini, R.; Staszewsky, L. A novel echocardiographic method closely agrees with cardiac magnetic resonance in the assessment of left ventricular function in infarcted mice. Sci. Rep. 2019, 9, 3580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabmaier, U.; Theiss, H.D.; Keithahn, A.; Kreiner, J.; Brenner, C.; Huber, B.; von der Helm, C.; Gross, L.; Klingel, K.; Franz, W.M.; et al. The role of 1.5 tesla MRI and anesthetic regimen concerning cardiac analysis in mice with cardiomyopathy. PLoS ONE 2014, 9, e94615. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, C.; Molojavyi, A.; Flögel, U.; Merx, M.W.; Ding, Z.; Schrader, J. Direct comparison of magnetic resonance imaging and conductance microcatheter in the evaluation of left ventricular function in mice. Basic Res. Cardiol. 2006, 101, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Phoon, C.K.L.; Turnbull, D.H. Cardiovascular Imaging in Mice. Curr. Protoc. Mouse Biol. 2016, 6, 15–38. [Google Scholar] [CrossRef] [Green Version]
- Yasmin, F.; Shah, S.M.I.; Naeem, A.; Shujauddin, S.M.; Jabeen, A.; Kazmi, S.; Siddiqui, S.A.; Kumar, P.; Salman, S.; Hassan, S.A.; et al. Artificial intelligence in the diagnosis and detection of heart failure: The past, present, and future. Rev. Cardiovasc. Med. 2021, 22, 1095–1113. [Google Scholar] [CrossRef]
- Goodroe, A.; Fitz, C.; Bakker, J. Current Topics in Marmoset Anesthesia and Analgesia. ILAR J. 2021, 61, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K.E.; Austad, S.N. The development of small primate models for aging research. ILAR J. 2011, 52, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattison, J.A.; Vaughan, K.L. An overview of nonhuman primates in aging research. Exp. Gerontol. 2017, 94, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Abbott, D.H.; Hearn, J.P. Physical, hormonal and behavioural aspects of sexual development in the marmoset monkey, Callithrix jacchus. J. Reprod. Fertil. 1978, 53, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Tardif, S.D.; Smucny, D.A.; Abbott, D.H.; Mansfield, K.; Schultz-Darken, N.; Yamamoto, M.E. Reproduction in captive common marmosets (Callithrix jacchus). Comp. Med. 2003, 53, 364–368. [Google Scholar] [PubMed]
- Benirschke, K.; Anderson, J.M.; Brownhill, L.E. Marrow Chimerism in Marmosets. Science 1962, 138, 513–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benirschke, K.; Brownhill, L.E. Further observations on marrow chimerism in marmosets. Cytogenetics 1962, 1, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.O.M.; Armada, J.L.A.; Verona, C.E.S.; Heliodoro, G.; Nogueira, D.M. Cytogenetics and Molecular Genetic Analysis of Chimerism in Marmosets (Callithrix: Primates). An. Acad. Bras. Cienc. 2017, 89, 2793–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenough, T.C.; Carville, A.; Coderre, J.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Mansfield, K. Pneumonitis and multi-organ system disease in common marmosets (Callithrix jacchus) infected with the severe acute respiratory syndrome-associated coronavirus. Am. J. Pathol. 2005, 167, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Charnock, J.S.; McLennan, P.L.; McIntosh, G.H.; Barnden, L.R.; Buttfield, I.H. Radionuclide angiographic study of the influence of dietary lipid supplements on cardiac function in the marmoset (Callithrix jacchus). Cardiovasc. Res. 1987, 21, 369–376. [Google Scholar] [CrossRef]
- McMurchie, E.J.; Patten, G.S.; McLennan, P.L.; Charnock, J.S.; Nestel, P.J. The influence of dietary lipid supplementation on cardiac beta-adrenergic receptor adenylate cyclase activity in the marmoset monkey. Biochim. Biophys. Acta 1988, 937, 347–358. [Google Scholar] [CrossRef]
- McMurchie, E.J.; Patten, G.S.; McLennan, P.L.; Charnock, J.S. A comparison of the properties of the cardiac beta-adrenergic receptor adenylyl cyclase system in the rat and the marmoset monkey. Comp. Biochem. Physiol. B 1987, 88, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Sobin, S.S.; Bernick, S.; Ballard, K.W. Histochemical characterization of the aging microvasculature in the human and other mammalian and non-mammalian vertebrates by the periodic acid-Schiff reaction. Mech. Ageing Dev. 1992, 63, 183–192. [Google Scholar] [CrossRef]
- Geula, C.; Nagykery, N.; Wu, C.K. Amyloid-beta deposits in the cerebral cortex of the aged common marmoset (Callithrix jacchus): Incidence and chemical composition. Acta Neuropathol. 2002, 103, 48–58. [Google Scholar] [CrossRef]
- Berkovitz, B.K.; Pacy, J. Age changes in the cells of the intra-articular disc of the temporomandibular joints of rats and marmosets. Arch. Oral Biol. 2000, 45, 987–995. [Google Scholar] [CrossRef]
- Harada, T.; Tokuriki, M.; Tanioka, Y. Age-related changes in the brainstem auditory evoked potentials of the marmoset. Heart Res. 1999, 128, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Black, A.; Lane, M.A. Nonhuman primate models of skeletal and reproductive aging. Gerontology 2002, 48, 72–80. [Google Scholar] [CrossRef]
- Ross, C.N.; Salmon, A.B. Aging research using the common marmoset: Focus on aging interventions. Nutr. Healthy Aging 2019, 5, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Crook, D.; Weisgraber, K.H.; Boyles, J.K.; Mahley, R.W. Isolation and characterization of plasma lipoproteins of common marmoset monkey. Comparison of effects of control and atherogenic diets. Arteriosclerosis 1990, 10, 633–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbey, M.; Clifton, P.M.; McMurchie, E.J.; McIntosh, G.H.; Nestel, P.J. Effect of a high fat/cholesterol diet with or without eicosapentaenoic acid on plasma lipids, lipoproteins and lipid transfer protein activity in the marmoset. Atherosclerosis 1990, 81, 163–174. [Google Scholar] [CrossRef]
- Lima, V.L.; Sena, V.L.; Stewart, B.; Owen, J.S.; Dolphin, P.J. An evaluation of the marmoset Callithrix jacchus (sagüi) as an experimental model for the dyslipoproteinemia of human Schistosomiasis mansoni. Biochim. Biophys. Acta 1998, 1393, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Michel, J.B.; Wood, J.; Hofbauer, K.; Corvol, P.; Menard, J. Blood pressure effects of renin inhibition by human renin antiserum in normotensive marmosets. Am. J. Physiol. 1984, 246, F309–F316. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, M.; Kim, S.; Ikemoto, F.; Murakami, K.; Yamamoto, K. Fate of recombinant human renin administered exogenously to anesthetized monkeys. Hypertension 1988, 12, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummer, C.; Vogt, E.J.; Heistermann, M.; Roshani, B.; Becker, T.; Mätz-Rensing, K.; Kues, W.A.; Kügler, S.; Behr, R. Generation and Breeding of EGFP-Transgenic Marmoset Monkeys: Cell Chimerism and Implications for Disease Modeling. Cells 2021, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Preuss, T.M. Critique of Pure Marmoset. Brain Behav. Evol. 2019, 93, 92–107. [Google Scholar] [CrossRef]
- Holtze, S.; Gorshkova, E.; Braude, S.; Cellerino, A.; Dammann, P.; Hildebrandt, T.B.; Hoeflich, A.; Hoffmann, S.; Koch, P.; Terzibasi Tozzini, E.; et al. Alternative Animal Models of Aging Research. Front. Mol. Biosci. 2021, 8, 660959. [Google Scholar] [CrossRef] [PubMed]
- Swindle, M.M.; Makin, A.; Herron, A.J.; Clubb, F.J., Jr.; Frazier, K.S. Swine as models in biomedical research and toxicology testing. Vet. Pathol. 2012, 49, 344–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaragoza, C.; Gomez-Guerrero, C.; Martin-Ventura, J.L.; Blanco-Colio, L.; Lavin, B.; Mallavia, B.; Tarin, C.; Mas, S.; Ortiz, A.; Egido, J. Animal models of cardiovascular diseases. J. Biomed. Biotechnol. 2011, 2011, 497841. [Google Scholar] [CrossRef] [PubMed]
- Hiroaki, O.; Kozue, M.; Hidetoshi, M. Large Animal Models in Cardiovascular Research. In Animal Models and Experimental Research in Medicine; Mahmut, K., Volkan, G., Abdulsamed, K., Eds.; IntechOpen: Rijeka, Croatia, 2022; p. Ch. 11. [Google Scholar]
- Kooij, V.; Venkatraman, V.; Tra, J.; Kirk, J.A.; Rowell, J.; Blice-Baum, A.; Cammarato, A.; Van Eyk, J.E. Sizing up models of heart failure: Proteomics from flies to humans. Proteom. Clin. Appl. 2014, 8, 653–664. [Google Scholar] [CrossRef] [Green Version]
- Chorro, F.J.; Such-Belenguer, L.; López-Merino, V. Animal Models of Cardiovascular Disease. Rev. Española Cardiol. 2009, 62, 69–84. [Google Scholar] [CrossRef]
- Barthélémy, I.; Hitte, C.; Tiret, L. The Dog Model in the Spotlight: Legacy of a Trustful Cooperation. J. Neuromuscul. Dis. 2019, 6, 421–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.D.; Zhai, W.; Yang, H.C.; Fan, R.X.; Cao, X.; Zhong, L.; Wang, L.; Liu, F.; Wu, H.; Cheng, L.G.; et al. The genomics of selection in dogs and the parallel evolution between dogs and humans. Nat. Commun. 2013, 4, 1860. [Google Scholar] [CrossRef] [Green Version]
- Hasenfuss, G. Animal models of human cardiovascular disease, heart failure and hypertrophy. Cardiovasc. Res. 1998, 39, 60–76. [Google Scholar] [CrossRef] [Green Version]
- Kaese, S.; Frommeyer, G.; Verheule, S.; van Loon, G.; Gehrmann, J.; Breithardt, G.; Eckardt, L. The ECG in cardiovascular-relevant animal models of electrophysiology. Herzschrittmacherther. Elektrophysiol. 2013, 24, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Lavine, S.J. Effect of changes in contractility on the index of myocardial performance in the dysfunctional left ventricle. Cardiovasc. Ultrasound 2006, 4, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loen, V.; Vos, M.A.; van der Heyden, M.A.G. The canine chronic atrioventricular block model in cardiovascular preclinical drug research. Br. J. Pharmacol. 2022, 179, 859–881. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.C.; Recchia, F. Canine Model of Pacing-Induced Heart Failure. Methods Mol. Biol. 2018, 1816, 309–325. [Google Scholar] [CrossRef]
- Duncker, D.J.; Bakkers, J.; Brundel, B.J.; Robbins, J.; Tardiff, J.C.; Carrier, L. Animal and in silico models for the study of sarcomeric cardiomyopathies. Cardiovasc. Res. 2015, 105, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Lowe, J.E.; Reimer, K.A.; Jennings, R.B. Experimental infarct size as a function of the amount of myocardium at risk. Am. J. Pathol. 1978, 90, 363–379. [Google Scholar]
- Khan, M.S.; Smego, D.; Ishidoya, Y.; Hirahara, A.M.; Offei, E.; Ruiz Castillo, M.S.; Gharbia, O.; Li, H.; Palatinus, J.A.; Krueger, L.; et al. A canine model of chronic ischemic heart failure. Am. J. Physiol. Heart Circ. Physiol. 2023, 324, H751–H761. [Google Scholar] [CrossRef] [PubMed]
- Chevènement, O.; Borenstein, N.; Kieval, R.; Fiette, L.; Aujard, F. Animal model considerations to evaluate prosthetic tricuspid valve implants. Ann. Anat. 2021, 234, 151625. [Google Scholar] [CrossRef] [PubMed]
- Milani-Nejad, N.; Janssen, P.M. Small and large animal models in cardiac contraction research: Advantages and disadvantages. Pharmacol. Ther. 2014, 141, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Hornyik, T.; Rieder, M.; Castiglione, A.; Major, P.; Baczko, I.; Brunner, M.; Koren, G.; Odening, K.E. Transgenic rabbit models for cardiac disease research. Br. J. Pharmacol. 2022, 179, 938–957. [Google Scholar] [CrossRef]
- Ginis, I.; Luo, Y.; Miura, T.; Thies, S.; Brandenberger, R.; Gerecht-Nir, S.; Amit, M.; Hoke, A.; Carpenter, M.K.; Itskovitz-Eldor, J.; et al. Differences between human and mouse embryonic stem cells. Dev. Biol. 2004, 269, 360–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emini Veseli, B.; Perrotta, P.; De Meyer, G.R.A.; Roth, L.; Van der Donckt, C.; Martinet, W.; De Meyer, G.R.Y. Animal models of atherosclerosis. Eur. J. Pharmacol. 2017, 816, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Bader, M. Rat Models of Cardiovascular Diseases. In Rat Genomics: Methods and Protocols; Anegon, I., Ed.; Humana Press: Totowa, NJ, USA, 2010; pp. 403–414. [Google Scholar] [CrossRef]
- González, G.E.; Seropian, I.M.; Krieger, M.L.; Palleiro, J.; Lopez Verrilli, M.A.; Gironacci, M.M.; Cavallero, S.; Wilensky, L.; Tomasi, V.H.; Gelpi, R.J.; et al. Effect of early versus late AT(1) receptor blockade with losartan on postmyocardial infarction ventricular remodeling in rabbits. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H375–H386. [Google Scholar] [CrossRef] [Green Version]
- Shiomi, M.; Ito, T.; Yamada, S.; Kawashima, S.; Fan, J. Development of an animal model for spontaneous myocardial infarction (WHHLMI rabbit). Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1239–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chobanian, A.V.; Arquilla, E.R.; Clarkson, T.B.; Eder, H.A.; Howard, C.F., Jr.; Regan, T.J.; Williamson, J.R. Cardiovascular complications. Diabetes 1982, 31, 54–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calasans-Maia, M.D.; Monteiro, M.L.; Ascoli, F.O.; Granjeiro, J.M. The rabbit as an animal model for experimental surgery. Acta Cir. Bras. 2009, 24, 325–328. [Google Scholar] [CrossRef] [Green Version]







| Animal | Age (Months) | Body Weight (g) | Heart Weight (g) | Blood Resistivity (ρ) | Muscle Resistivity (ρ) | HR (Mean bpm) | O2 (Mean %) | RR (Mean/Min) | rT (Mean °C) |
|---|---|---|---|---|---|---|---|---|---|
| Animal 1 | 43 | 456 | 2.84 | 2.4 | 4.6 | 229 | 99 | 16 | 37.0 |
| Animal 2 | 43 | 449 | 3.13 | 2.7 | 4.6 | 142 | 99 | 16 | 35.0 |
| Animal 3 | 41 | 419 | 3.70 | 2.5 | 4.6 | 174 | 100 | 16 | 37.0 |
| Animal 4 | 34 | 473 | 3.46 | 2.5 | 4.6 | 200 | 98 | 16 | 36.0 |
| Animal 5 | 34 | 388 | 2.78 | 3.1 | 4.6 | 189 | 99 | 16 | 36.0 |
| Animal 6 | 33 | 423 | - | 2.7 | 4.7 | 174 | 96 | 16 | 35.0 |
| Mean ± SEM | 38 ± 2 | 435 ± 12.5 | 3.18 ± 0.2 | 2.65 ± 0.1 | 4.62 ± 0.0 | 184 ± 11.9 | 98 ± 0.5 | 16 ± 0.1 | 36.0 ± 0.4 |
| Parameter | Mean ± SEM (n = 6) | |
|---|---|---|
| Baseline data | ||
| HR (bpm) | 198 ± 14 | |
| ESV (µL) | 89 ± 21 | |
| EDV (µL) | 315 ± 27 | |
| SV (µL) | 226 ± 25 | |
| EF (%) | 72 ± 6 | |
| CO (ml/min) | 44 ± 6 | |
| SW (mmHG∗µL) | 9191 ± 768 | |
| dp/dt max (mmHG/s) | 1429 ± 166 | |
| dp/dt min (mmHG/s) | −1221 ± 142 | |
| Ea (mmHG/µL) | 0.23 ± 0.03 | |
| Tau (ms) | 19 ± 1.0 | |
| Contractility | ||
| ESPVR (linear) Ees (mmHG/µL) | 0.467 ± 0.095 | |
| ESPVR (linear) V100 (µL) | 214 ± 55 | |
| ESPVR (linear) r2 | 0.816 ± 0.069 | |
| ESPVR (quadratic) a | −0.012 ± 0.004 | |
| ESPVR (quadratic) b | 1.634 ± 0.494 | |
| ESPVR (quadratic) r2 | 0.857 ± 0.048 | |
| PRSW r2 | 0.945 ± 0.017 | |
| PRSW slope | 40 ± 5 | |
| PRSW axis intercept | −1949 ± 506 | |
| PE mmHG∗µL | 3112 ± 1181 | |
| PVA mmHG∗µL | 6217 ± 1689 | |
| Compliance | ||
| EDPVR (linear) dp/dV (mmHG/µL) | 0.027 ± 0.009 | |
| EDPVR (linear) axis intercept | −1.132 ± 1.189 | |
| EDPVR (linear) r2 | 0.867 ± 0.035 | |
| EDPVR (exponential) dp/dV (k1) (mmHG/µL) | 0.013 ± 0.004 | |
| EDPVR (exponential) k2 | 0.793 ± 0.292 | |
| EDPVR (exponential) r2 | 0.843 ± 0.039 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klösener, L.; Samolovac, S.; Barnekow, I.; König, J.; Moussavi, A.; Boretius, S.; Fuchs, D.; Haegens, A.; Hinkel, R.; Mietsch, M. Functional Cardiovascular Characterization of the Common Marmoset (Callithrix jacchus). Biology 2023, 12, 1123. https://doi.org/10.3390/biology12081123
Klösener L, Samolovac S, Barnekow I, König J, Moussavi A, Boretius S, Fuchs D, Haegens A, Hinkel R, Mietsch M. Functional Cardiovascular Characterization of the Common Marmoset (Callithrix jacchus). Biology. 2023; 12(8):1123. https://doi.org/10.3390/biology12081123
Chicago/Turabian StyleKlösener, Lina, Sabine Samolovac, Ina Barnekow, Jessica König, Amir Moussavi, Susann Boretius, Dieter Fuchs, Astrid Haegens, Rabea Hinkel, and Matthias Mietsch. 2023. "Functional Cardiovascular Characterization of the Common Marmoset (Callithrix jacchus)" Biology 12, no. 8: 1123. https://doi.org/10.3390/biology12081123
APA StyleKlösener, L., Samolovac, S., Barnekow, I., König, J., Moussavi, A., Boretius, S., Fuchs, D., Haegens, A., Hinkel, R., & Mietsch, M. (2023). Functional Cardiovascular Characterization of the Common Marmoset (Callithrix jacchus). Biology, 12(8), 1123. https://doi.org/10.3390/biology12081123