
BLTR1 Is Decreased in Steroid Resistant Pro-Inflammatory CD28nullCD8+ T Lymphocytes in Patients with COPD—The Spillover Hypothesis Explained?
 , ,
, ,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Patient and Control Groups
2.2. Leucocyte Counts
2.3. CD28± CD8± T, NKT-like Cell Subsets
2.4. BLTR1, IFNγ, and TNFα Intracellular Cytokine Production by CD28± CD8± T, NKT-like Cell Subsets Using Flow Cytometry
2.5. BLTR1 Expression in CD28+ and CD28null T Cells via Western Blot
2.6. Effect of Prednisolone on BLTR1, IFNγ and TNFα Expression by CD28± CD8+ T and NKT-like Cells
2.7. Migration of CD28± CD8+ T and NKT-like Cells
2.8. Statistical Analysis
3. Results
3.1. Percentages of Blood Lymphocyte Subsets
3.2. Percentages of BAL Lymphocyte Subsets
3.3. Percentages of Large Airway Brushing Intraepithelial Lymphocyte Subsets
3.4. Increased CD28nullCD8+ T and NKT-like Cells in COPD
3.5. Increased CD28± CD8± T and NKT-like Lymphocyte Subsets Producing IFNγ and TNFα Pro-Inflammatory Cytokines in COPD Patients
3.6. BLTR1 Expression by CD28± CD8± T and NKT-like Cells
3.7. Correlation between Small Airway CD28nullCD8+BLTR1-T Cells Producing IFNγ/TNFα and Patient FEV1
3.8. Effect of Prednisolone on BLTR1, IFNγ, and TNFα Expression by CD28nullCD8+ T and NKT-like Cells
3.9. Chemotaxis of CD28± CD8+ T and NKT-like Cells
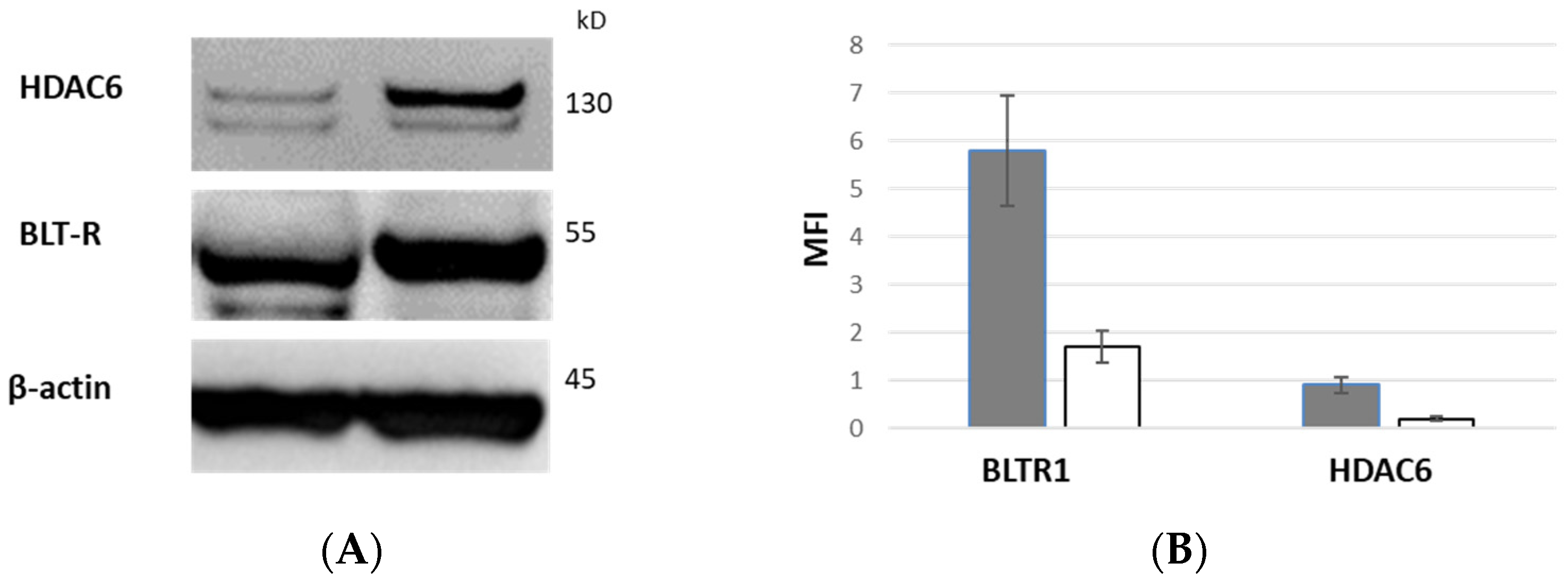
3.10. BLTR1 Expression of CD28+ and CD28null T Cells by Western Blot
3.11. HDAC6 Expression of CD28+ and CD28null T Cells by Western Blot
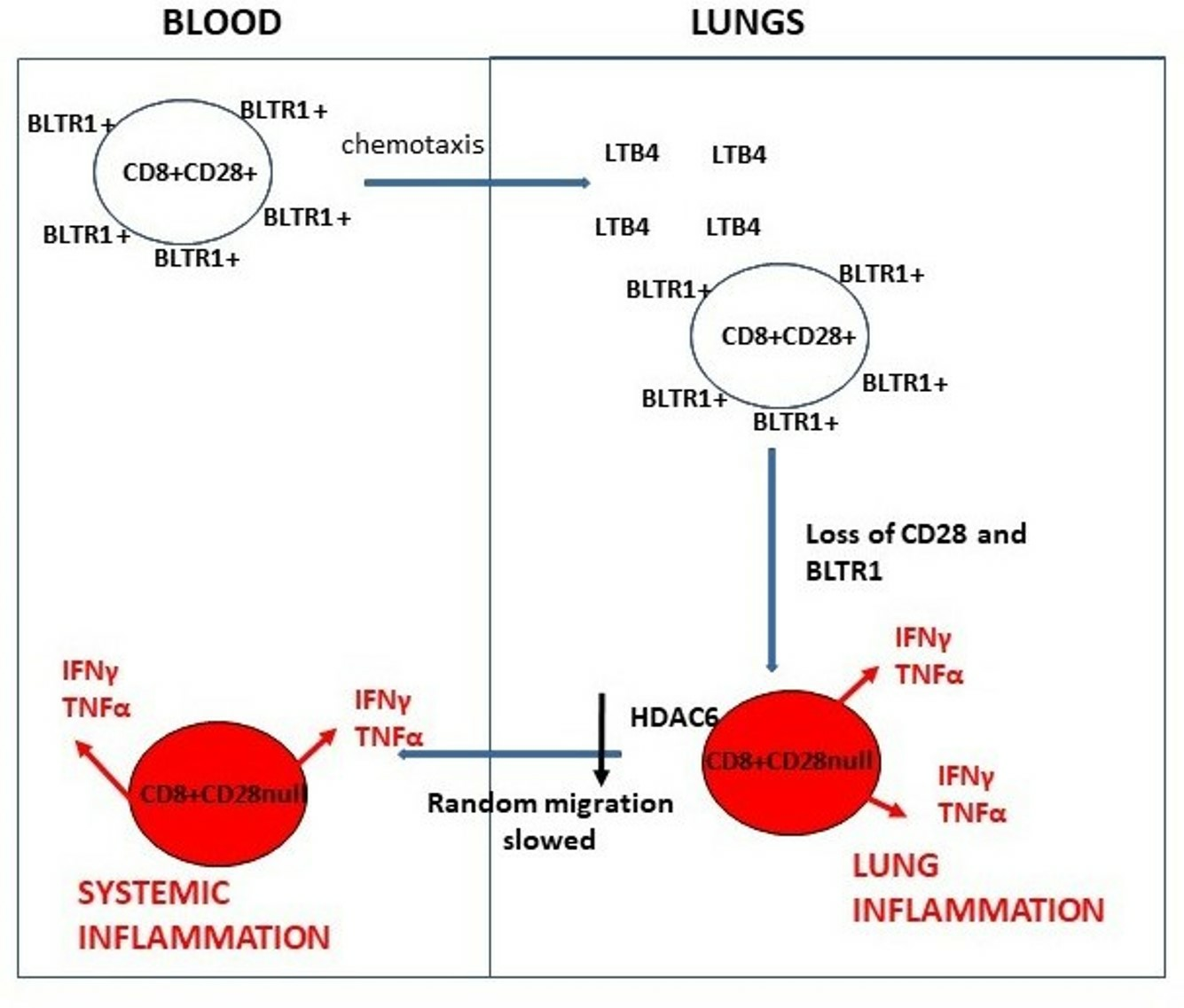
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Barnes, P.J.; Adcock, I.M. Glucocorticoid resistance in inflammatory diseases. Lancet 2009, 373, 1905–1917. [Google Scholar] [CrossRef]
- Barnes, P.J.; Celli, B.R. Systemic manifestations and comorbidities of COPD. Eur. Respir. J. 2009, 33, 1165–1185. [Google Scholar] [CrossRef]
- Wouters, E.F.M.; Reynaert Dentener, M.A.; Vernpy, J.H.J. Systemic and local inflammation in asthma and chronic obstructive pulmonary disease. Is there a connection? Proc. Am. Thorac. Soc. 2009, 6, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Hodge, G.; Nairn, J.; Holmes, M.; Reynolds, P.; Hodge, S. Increased intracellular Th1 pro-inflammatory cytokine production in peripheral blood, bronchoalveolar lavage and intraepithelieal T cells of COPD subjects. Clin. Exp. Immunol. 2007, 150, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Hodge, G.; Mukaro, V.; Reynolds, P.N.; Hodge, S. Role of increased CD8/CD28null T cells and alternative co-stimulatory molecules in chronic obstructive pulmonary disease. Clin. Exp. Immunol. 2011, 166, 94–102. [Google Scholar] [CrossRef]
- Hodge, G.; Hodge, S. Steroid resistant CD8+CD28nullNKT-like pro-inflammatory cytotoxic cells in chronic obstructive pulmonary disease. Front. Immunol. 2016, 7, 617. [Google Scholar] [CrossRef]
- Goodarzi, K.; Goodarzi, M.; Tager, A.M.; Luster, A.D.; Andrian, U.H. Leukotriene B4 and BLTR1 control cytotoxic effector T cell recruitment to inflamed tissues. Nat. Immunol. 2003, 4, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Montrushi, P.; Kharitonov, S.A.; Ciabattoni, G.; Barnes, P.J. Exhaled leukotrienes and prostaglandins in COPD. Thorax 2003, 58, 585–588. [Google Scholar]
- Marian, E.; Baraldo, S.; Visentin, A.; Papi, A.; Saetta, M.; Fabbri, L.; Maestrelli, P. Upregulated membrane nuclear leukotriene B4 receptors in COPD. Chest 2006, 129, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Hodge, G.; Jersmann, H.; Tran, H.B.; Asare, P.F.; Jayapal, M.; Reynolds, P.N.; Holmes, M.; Hodge, S. COPD is associated with increased pro-inflammatory CD28nullCD8+ T and NKT-like cells in the small airways. Clin. Exp. Immunol. 2022, 207, 351–359. [Google Scholar] [CrossRef]
- Global Initiative for Chronic Obstructive Lung Disease. Global Strategy for the Diagnosis, Management and Prevention of Chronic Obstructive Pulmonary Disease. 2011. Available online: www.goldcopd.org/ (accessed on 7 July 2023).
- Cabrero, J.V.; Serrador, J.M.; Barreiro, O.; Mittelbrunn, M.; Naranjo-Suárez, S.; Martín-Cófreces, N.; Vicente-Manzanares, M.; Mazitschek, R.; Bradner, J.E.; Jesús Ávila, J.; et al. Lymphocyte chemotaxis Is regulated by histonedeacetylase 6, independently of its deacetylase activity. Mol. Biol. Cell 2006, 17, 3435–3445. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Chronic obstructive pulmonary disease: Effects beyond the lungs. PLoS Med. 2010, 7, e1000220. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rio, F.; Mirravittles, M.; Soriano, J.B.; Munoz, L.; Duran-Tauleria, E.; Sanchez, G.; Sobradillo, V.; Ancochea, J.; EPI-SCAN Steering Committee. Systemic inflammation in chronic obstructive pulmonary disease: A population-based study. Respir. Res. 2010, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Saetta, M.; Stefano, A.D.; Turato, G.; Facchini, F.M.; Corbino, L.; Mapp, C.E.; Maestrelli, P.; Ciaccia, A.; Fabbri, L.M. CD8+ T-lymphocytes in peripheral airways of smokers with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1998, 157, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Cosio, M.G.; Guerassimov, A. Chronic obstructive pulmonary disease. Inflammation of the small airways and lung parenchyma. Am. J. Respir. Crit. Care Med. 1999, 160, s21–s25. [Google Scholar] [CrossRef]
- Bryl, E.; Vallejo, A.N.; Weynard, C.M.; Goronzy, J.J. Down-regulation of CD28 expression by TNFalpha. J. Immunol. 2001, 167, 3231–3238. [Google Scholar] [CrossRef] [PubMed]
- Tager, A.M.; Bradley, S.K.; Medoff, B.D.; Islam, S.A.; Bercury, S.D.; Friedrich, E.B.; Carafone, S.D.; Gerszten, R.E.; Luster, A.D. Leukotriene B4 receptor BLT1 mediates early effector T cell recruitment. Nat. Immunol. 2003, 4, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Borrill, Z.L.; Singh, R.D. Exhaled breath condensate biomarkers. . Eur Respir. J 2008, 32, 472–486. [Google Scholar] [CrossRef]
- Onishi, H.O.; Miyakara, N.; Dakhama, A.; Takeda, K.; Mathis, S.; Haribabu, B.; Gelfand, E.W. Corticosteroids enhance CD8+ T cell–mediated airway hyperresponsiveness and allergic inflammation by upregulating leukotriene B4 receptor 1. J. Allergy Clin. Immunol. 2008, 121, 864–871. [Google Scholar] [CrossRef]
- Stephen Kershaw, S.; Morgan, D.J.; Boyd, J.; Spiller, D.G.; Kitchen, G.; Zindy, E.; Iqbal, M.; Rattray, M.; Sanderson, C.M.; Brass, A.; et al. Glucocorticoids rapidly inhibit cell migration through a novel, non-transcriptional HDAC6 pathway. J. Cell Sci. 2020, 133, jcs242842. [Google Scholar] [CrossRef]
- Teo, F.H.; de Oliveira, R.T.; Mamoni, R.L.; Ferreira, M.C.; Nadruz, W.; Coelho, O.R.; Fernandes, J.; Blotta, M. Characterisation of CD4+CD28null T cells in patients witjh coronary artery disease and individuals with risk factors for atherosclerosis. Cell Immunol. 2013, 281, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Thewissen, M.; Somers, V.; Hellings, N.; Fraussen, J.; Damoiseaux, J.; Stinissen, P. CD4+CD28null T cells in autoimmune disease: Pathogenic features and decreased susceptibility to immunoregulation. J. Immunol. 2007, 179, 6514–6523. [Google Scholar] [CrossRef] [PubMed]
- Fasth, A.E.; Snir, O.; Johansson, A.A.; Nordmark, B.; Rahbar, A.; Klint, E.; Björkström, N.K.; Ulfgren, A.K.; van Vollenhoven, R.F.; Malmström, V.; et al. Skewed distribution of pro-inflammatory CD4+CD28null T cells in rheumatoid arthritis. Arthritis Res. Ther. 2007, 9, R87. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Fukunaga, K.; Ikeuchi, H.; Hamikozuru, K.; Hida, N.; Ohda, Y.; Iimuro, M.; Yoshida, K.; Kikuyama, R.; Kato, K.; et al. The CD4+CD28null and the regulatory CD4+CD25high T-cell phenotypes in patients with ulcerative colitis during active and quiescent disease following colectomy. Cytokine 2011, 56, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, A.N. CD28 extinction in human T cells: Altered functions and the program of T-cell senescence. Immunol. Rev. 2005, 205, 158–169. [Google Scholar] [CrossRef]
- Yao, H.; Rahman, I. Role of histone deacetylase 2 in epigenetics and cellular senescence: Implications in lung inflammaging and COPD. Am. J. Physiol. Lung Mol. Physiol. 2012, 303, 557–566. [Google Scholar] [CrossRef]
- Ohnishi, H.; Miyahara, N.; Dakhama, A.; Takeda, K.; Mathis, S.; Haribabu, B.; Gelfand, E.W. Role of leukotriene B4 in allergic diseases. Allergology 2008, 57, 291–298. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects | Control | COPD |
|---|---|---|
| Number | 10 | 10 |
| Age (years) | 55 (43–69) # | 56 (36–67) |
| FEV1% pred | 103.6 (94.8–102.2) | 51 (42–80) * |
| FEV1% FVC | 69 (58–71) | 50 (34–64) * |
| Male/female | 5/5 | 5/5 |
| COPD | T Cells | NKT-like Cells | ||
| CD28nullCD8+ | CD28nullCD4+ | CD28nullCD8+ | CD28nullCD4+ | |
| BLOOD | * 54 (36–64) # | 7 (3–11) | * 60 (34–71) | 5 (3–9) |
| BAL | * 61 (41–74) | 6 (3–12) | * 65 (46–90) | 8 (3–10) |
| LA | * 70 (51–91) | 7 (2–12) | * 72 (40–76) | 7 (2–12) |
| SA | * 87 (71–98) @ | 10 (2–16) | * 89 (74–98) | 10 (2–14) |
| Control | T Cells | NKT-like Cells | ||
| CD28nullCD8+ | CD28nullCD4+ | CD28nullCD8+ | CD28nullCD4+ | |
| BLOOD | 33 (18–40) | 5 (2–10) | 34 (21–42) | 4 (2–7) |
| BAL | 32 (13–44) | 7 (1–14) | 35 (19–53) | 5 (2–8) |
| LA | 30 (10–41) | 8 (3–14) | 35 (14–49) | 8 (3–14) |
| SA | 37 (20–55) | 12 (1–21) | 44 (23–58) | 11 (3–15) |
| COPD | T Cells | NKT-like Cells | ||
| CD28nullCD8+ | CD28nullCD8+ | |||
| IFNγ | TNFα | IFNγ | TNFα | |
| BLOOD | * 54 (37–65) # | * 56 (43–67) | * 62 (34–69) | * 65 (39–75) |
| BAL | * 64 (41–78) | * 61 (41–72) | * 66 (46–88) | * 71 (43–76) |
| LA | * 71 (51–88) | * 65 (52–86) | * 72 (42–82) | * 79 (50–81) |
| SA | * 88 (71–98) @ | * 93 (76–97) @ | * 89 (72–99) @ | * 90 (71–96) @ |
| Control | T Cells | NKT-like Cells | ||
| CD28nullCD8+ | CD28nullCD8+ | |||
| IFNγ | TNFα | IFNγ | TNFα | |
| BLOOD | 32 (18–40) | 34 (23–41) | 34 (22–41) | 33 (14–46) |
| BAL | 34 (12–44) | 32 (13–35) | 37 (17–51) | 34 (12–44) |
| LA | 30 (14–41) | 38 (12–39) | 31 (15–44) | 38 (13–55) |
| SA | 39 (20–56) | 29 (10–42) | 41 (26–65) | 32 (10–44) |
| COPD | BLTR+T Cells | BLTR+NKT-like Cells | ||
| CD28+CD8+ | CD28nullCD8+ | CD28+CD8+ | CD28nullCD8+ | |
| BLOOD | 56 (37–69) # | * 6 (4–9) | 60 (38–70) | * 3 (1–4) |
| BAL | 52 (35–66) | * 7 (3–10) | 65 (41–77) | * 5 (2–7) |
| LA | 50 (36–61) | * 8 (4–11) | 72 (40–78) | * 6 (4–10) |
| SA | 47 (34–63) | * 3 (2–5) | 58 (38–72) | * 2 (1–4) |
| Control | BLTR1+T Cells | BLTR1+NKT-like Cells | ||
| CD28+CD8+ | CD28nullCD8+ | CD28+CD8+ | CD28nullCD8+ | |
| BLOOD | 62 (38–73) | * 10 (2–10) | 62 (37–75) | * 9 (5–13) |
| BAL | 59 (38–71) | * 11 (1–14) | 61 (39–73) | * 11 (6–15) |
| LA | 60 (39–72) | * 13 (3–14) | 67 (42–78) | * 12 (7–15) |
| SA | 55 (37–68) | * 10 (1–12) | 57 (37–70) | * 11 (6–14) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hodge, G.; Jersmann, H.; Holmes, M.; Asare, P.; Roscioli, E.; Reynolds, P.N.; Hodge, S. BLTR1 Is Decreased in Steroid Resistant Pro-Inflammatory CD28nullCD8+ T Lymphocytes in Patients with COPD—The Spillover Hypothesis Explained? Biology 2023, 12, 1261. https://doi.org/10.3390/biology12091261
Hodge G, Jersmann H, Holmes M, Asare P, Roscioli E, Reynolds PN, Hodge S. BLTR1 Is Decreased in Steroid Resistant Pro-Inflammatory CD28nullCD8+ T Lymphocytes in Patients with COPD—The Spillover Hypothesis Explained? Biology. 2023; 12(9):1261. https://doi.org/10.3390/biology12091261
Chicago/Turabian StyleHodge, Greg, Hubertus Jersmann, Mark Holmes, Patrick Asare, Eugene Roscioli, Paul N. Reynolds, and Sandra Hodge. 2023. "BLTR1 Is Decreased in Steroid Resistant Pro-Inflammatory CD28nullCD8+ T Lymphocytes in Patients with COPD—The Spillover Hypothesis Explained?" Biology 12, no. 9: 1261. https://doi.org/10.3390/biology12091261
APA StyleHodge, G., Jersmann, H., Holmes, M., Asare, P., Roscioli, E., Reynolds, P. N., & Hodge, S. (2023). BLTR1 Is Decreased in Steroid Resistant Pro-Inflammatory CD28nullCD8+ T Lymphocytes in Patients with COPD—The Spillover Hypothesis Explained? Biology, 12(9), 1261. https://doi.org/10.3390/biology12091261