

Rays in the Shadows: Batoid Diversity, Occurrence, and Conservation Status in Fiji


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
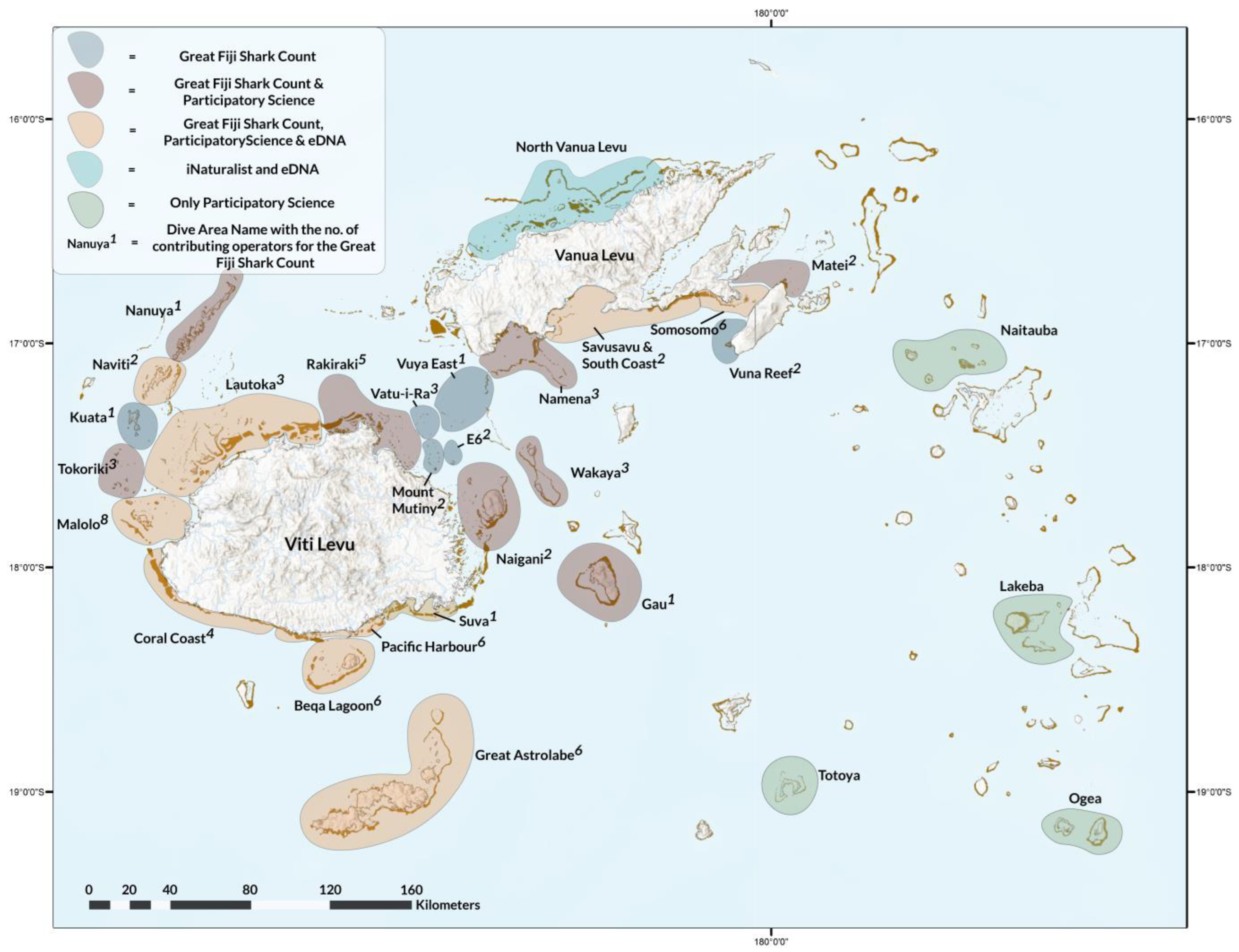
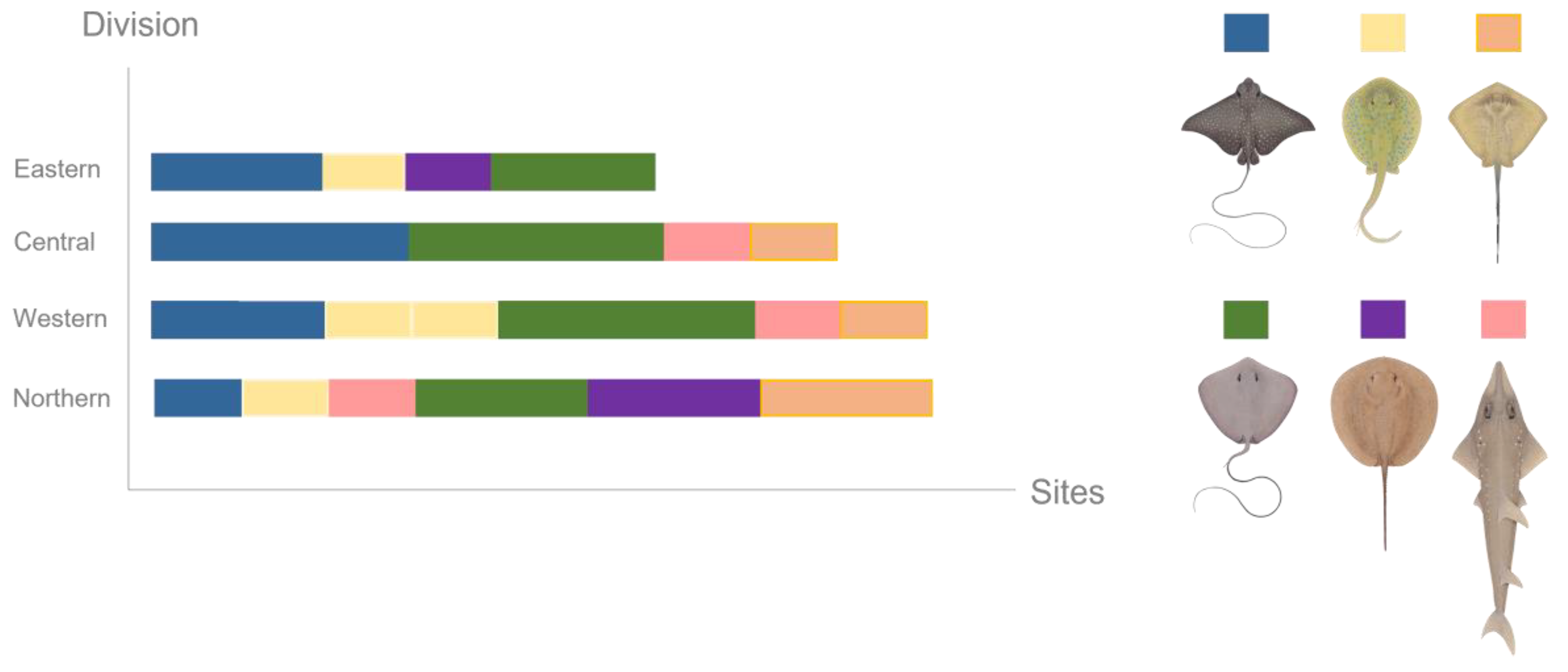
2.1. Study Area
2.2. Literature Review
2.3. Participatory Science Programs
2.3.1. GFSC
2.3.2. PA
2.3.3. MPF
2.3.4. iNaturalist
2.4. Environmental DNA Collection and Sample Processing
3. Results
3.1. Literature Review
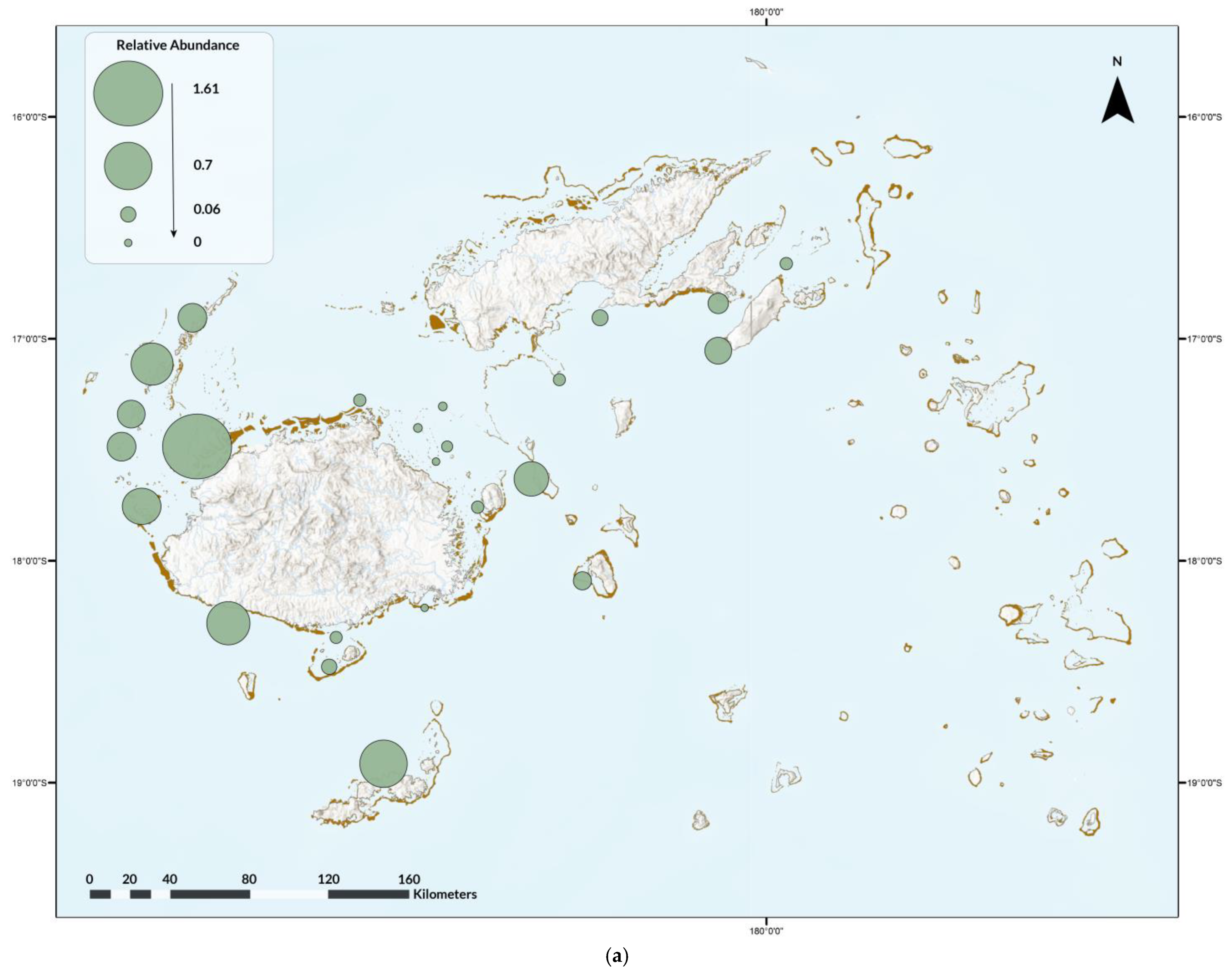
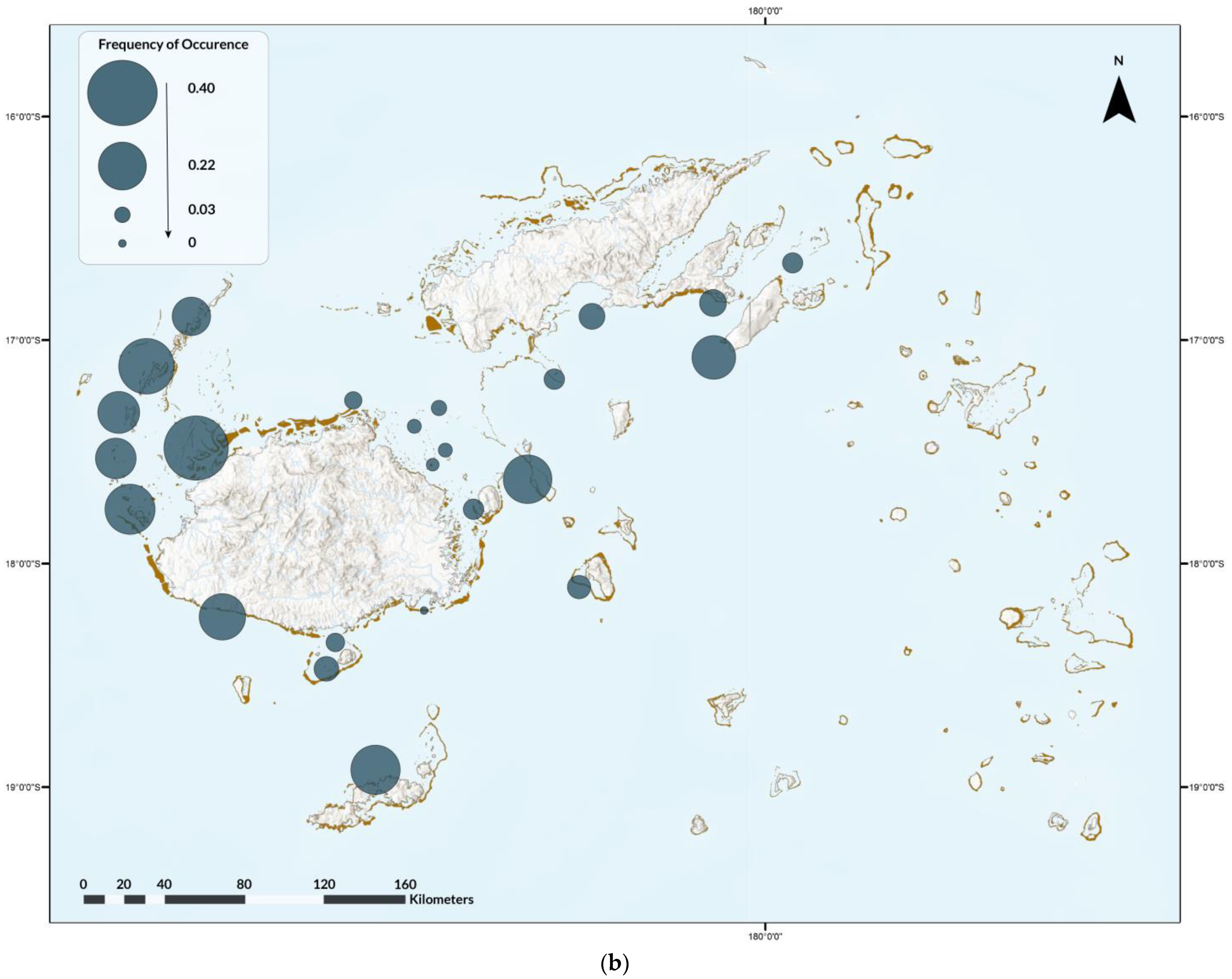
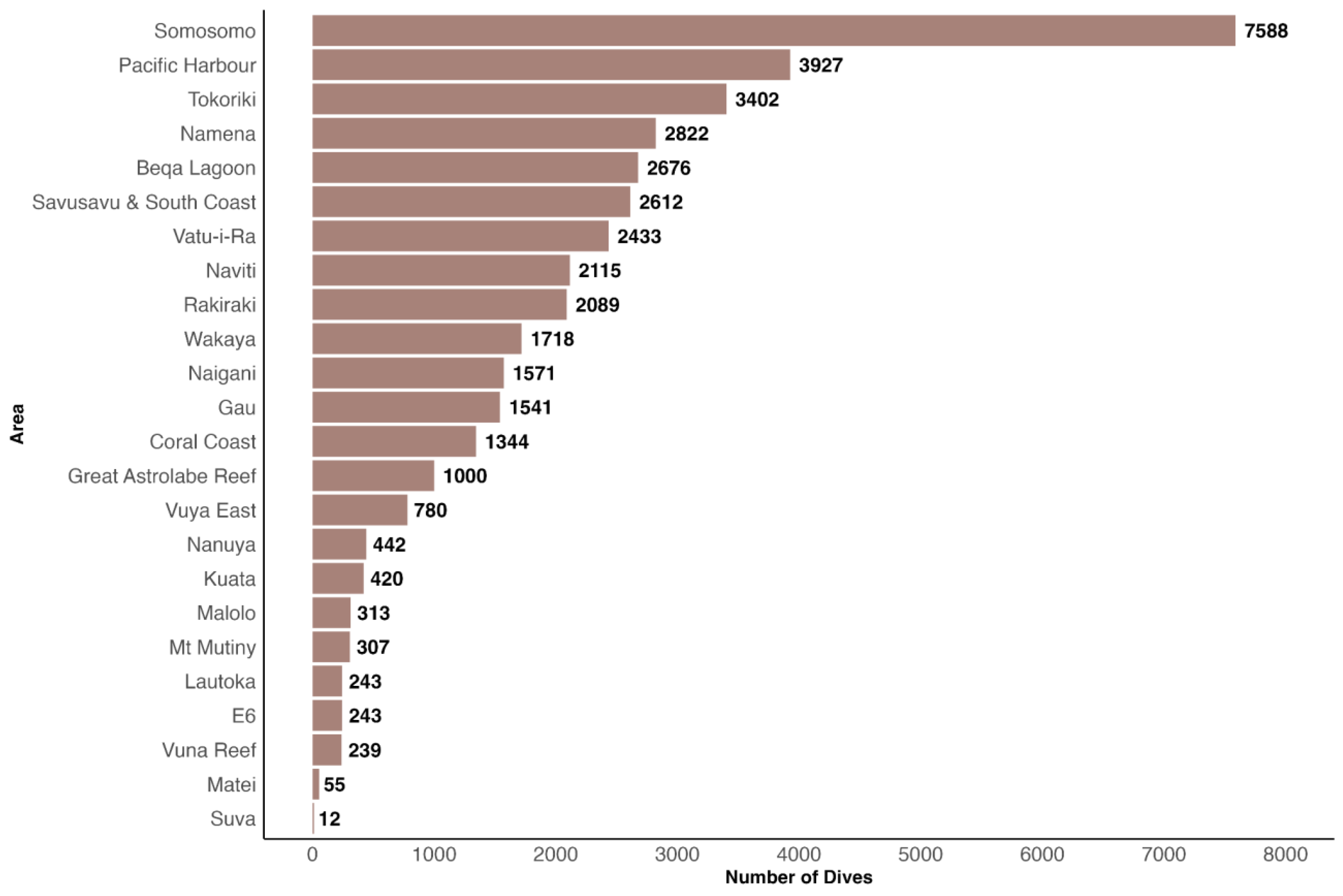
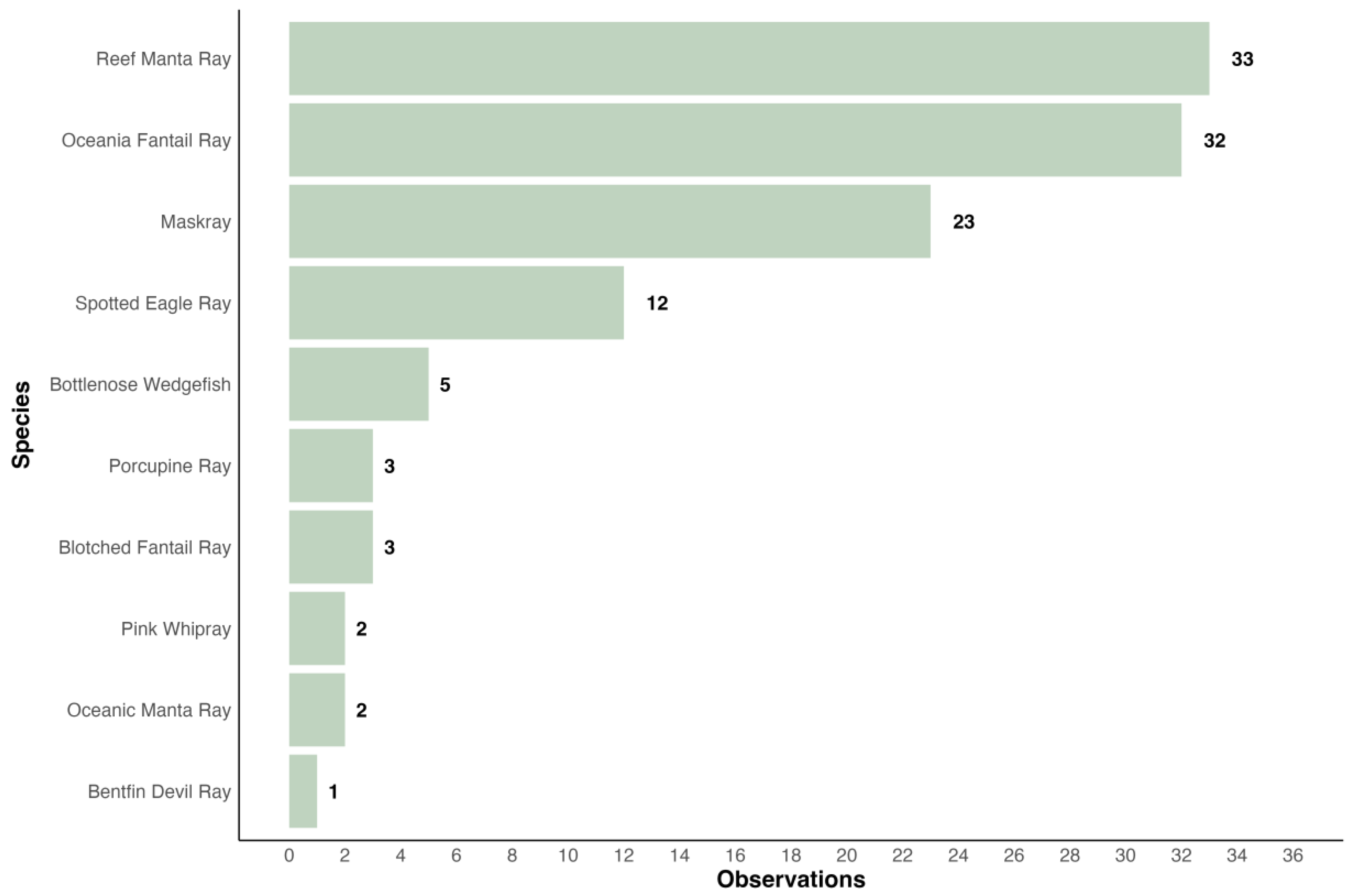
3.2. Participatory Science
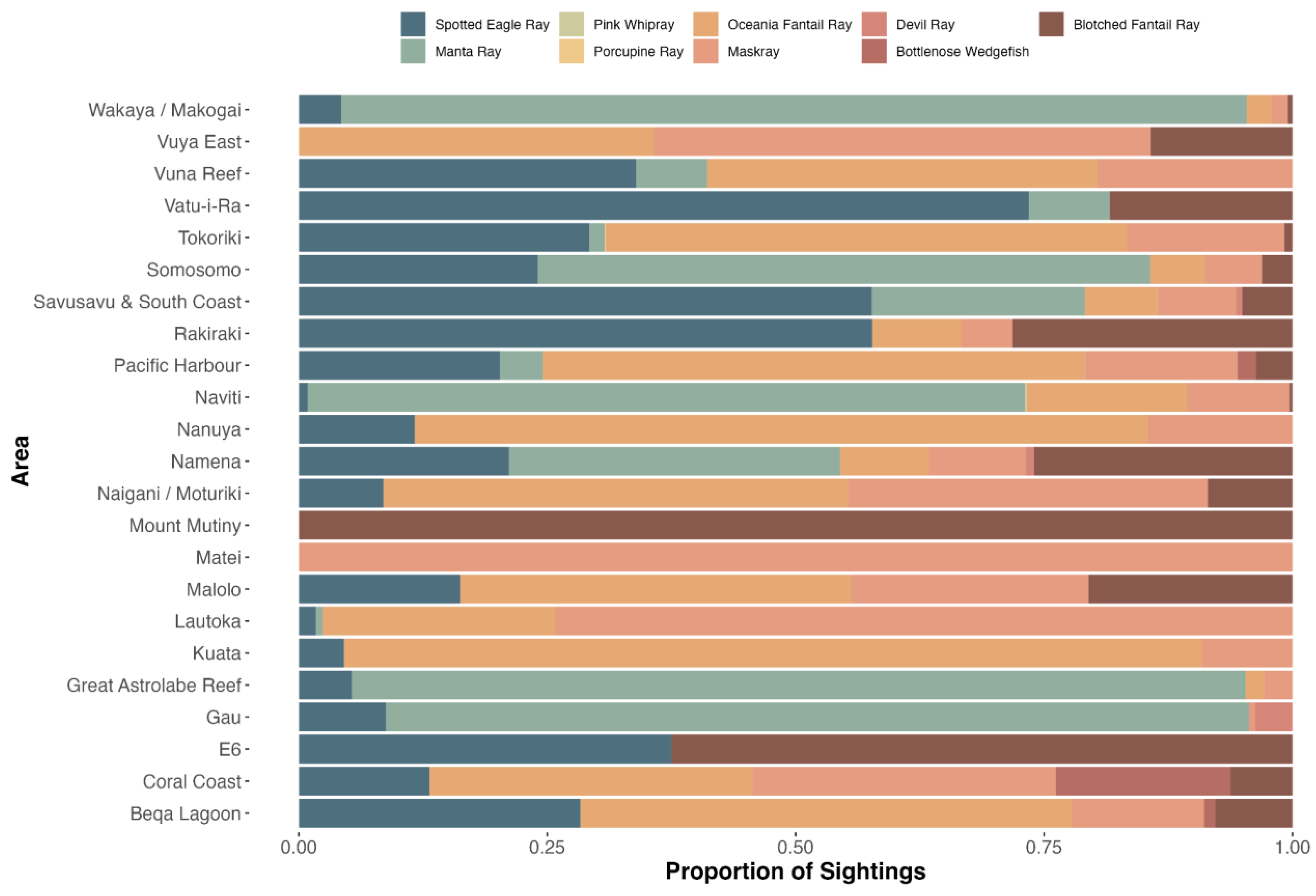
3.2.1. GFSC
3.2.2. Projects Abroad
3.2.3. Manta Project Fiji
3.2.4. iNaturalist
3.3. eDNA
4. Discussion
4.1. Literature Review
4.2. Participatory Science Programs
Species Verification
4.3. eDNA
4.4. Wedgefishes and Sawfishes
4.5. Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dulvy, N.K.; Pacoureau, N.; Rigby, C.L.; Pollom, R.A.; Jabado, R.W.; Ebert, D.A.; Finucci, B.; Pollock, C.M.; Cheok, J.; Derrick, D.H.; et al. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 2021, 31, 4773–4787.e8. [Google Scholar] [CrossRef]
- Worm, B.; Orofino, S.; Burns, E.S.; D’Costa, N.G.; Feitosa, L.M.; Palomares, M.L.D.; Schiller, L.; Bradley, D. Global shark fishing mortality still rising despite widespread regulatory change. Science 2024, 383, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Cardeñosa, D.; Shea, S.K.; Zhang, H.; Fischer, G.A.; Simpfendorfer, C.A.; Chapman, D.D. Two thirds of species in a global shark fin trade hub are threatened with extinction: Conservation potential of international trade regulations for coastal sharks. Conserv. Lett. 2022, 15, e12910. [Google Scholar] [CrossRef]
- Sherman, C.S.; Simpfendorfer, C.A.; Pacoureau, N.; Matsushiba, J.H.; Yan, H.F.; Walls, R.H.; Rigby, C.L.; VanderWright, W.J.; Jabado, R.W.; Pollom, R.A.; et al. Half a century of rising extinction risk of coral reef sharks and rays. Nat. Commun. 2023, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Pacoureau, N.; Rigby, C.L.; Kyne, P.M.; Sherley, R.B.; Winker, H.; Carlson, J.K.; Fordham, S.V.; Barreto, R.; Fernando, D.; Francis, M.P.; et al. Half a century of global decline in oceanic sharks and rays. Nature 2021, 589, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Simpfendorfer, C.A.; Heithaus, M.R.; Heupel, M.R.; MacNeil, M.A.; Meekan, M.; Harvey, E.; Sherman, C.S.; Currey-Randall, L.M.; Goetze, J.S.; Kiszka, J.J.; et al. Widespread diversity deficits of coral reef sharks and rays. Science 2023, 380, 1155–1160. [Google Scholar] [CrossRef]
- Kyne, P.M.; Jabado, R.W.; Rigby, C.L.; Dharmadi; Gore, M.A.; Pollock, C.M.; Herman, K.B.; Cheok, J.; Ebert, D.A.; Simpfendorfer, C.A. The thin edge of the wedge: Extremely high extinction risk in wedgefishes and giant guitarfishes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 1337–1361. [Google Scholar] [CrossRef]
- Last, P.; Naylor, G.; Séret, B.; White, W.; de Carvalho, M.; Stehmann, M. Rays of the World; CSIRO Publishing: Clayton South, Australia, 2016. [Google Scholar]
- Bräutigam, A.; Callow, M.; Campbell, I. Global Priorities for Conserving Sharks and Rays: A 2015-2025 Strategy; Global Sharks and Rays Initiative: Cambridge, UK, 2016. [Google Scholar]
- Jorgensen, S.J.; Micheli, F.; White, T.D.; Van Houtan, K.S.; Alfaro-Shigueto, J.; Andrzejaczek, S.; Arnoldi, N.S.; Baum, J.K.; Block, B.; Britten, G.L.; et al. Emergent research and priorities for shark and ray conservation. Endanger. Species Res. 2022, 47, 171–203. [Google Scholar] [CrossRef]
- Naylor, G.J.; Caira, J.N.; Jensen, K.; Rosana, K.; White, W.T.; Last, P. A DNA sequence–based approach to the identification of shark and ray species and its implications for global elasmobranch diversity and parasitology. Bull. Am. Mus. Nat. Hist. 2012, 367, 1–262. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Fowler, S.L.; Musick, J.A.; Cavanagh, R.D.; Kyne, P.M.; Harrison, L.R.; Carlson, J.K.; Davidson, L.N.; Fordham, S.V.; Francis, M.P.; et al. Extinction risk and conservation of the world’s sharks and rays. elife 2014, 3, e00590. [Google Scholar] [CrossRef]
- Hirschfeld, M.; Dudgeon, C.; Sheaves, M.; Barnett, A. Barriers in a sea of elasmobranchs: From fishing for populations to testing hypotheses in population genetics. Glob. Ecol. Biogeogr. 2021, 30, 2147–2163. [Google Scholar] [CrossRef]
- Shiffman, D.S.; Elliott, J.N.; Macdonald, C.C.; Wester, J.N.; Polidoro, B.A.; Ferry, L.A. The next generation of conservation research and policy priorities for threatened and exploited chondrichthyan fishes in the United States: An expert solicitation approach. Conserv. Sci. Pract. 2022, 4, e12629. [Google Scholar] [CrossRef]
- Weisler, M.I.; McNiven, I.J. Four thousand years of western Torres Strait fishing in the Pacific-wide context. J. Archaeol. Sci. Rep. 2016, 7, 764–774. [Google Scholar] [CrossRef]
- WHITE, W.T. Atelomycterus erdmanni, a new species of catshark (Scyliorhinidae: Carcharhiniformes) from Indonesia. J. Ocean Sci. Found. 2015, 14, 14–27. [Google Scholar]
- White, W.T.; Appleyard, S.A.; Sabub, B.; Kyne, P.M.; Harris, M.; Lis, R.; Baje, L.; Usu, T.; Smart, J.J.; Corrigan, S.; et al. Rediscovery of the threatened river sharks, Glyphis garricki and G. glyphis, in Papua New Guinea. PLoS ONE 2015, 10, e0140075. [Google Scholar] [CrossRef]
- Brunnschweiler, J.M. The Shark Reef Marine Reserve: A marine tourism project in Fiji involving local communities. J. Sustain. Tour. 2010, 18, 29–42. [Google Scholar] [CrossRef]
- Ferrier-Watson, S. Swimming with Gods: Exploring the Nation, Oceans, and Myths of Fiji; Collin College: McKinney, TX, USA, 2015. [Google Scholar]
- Glaus, K.B.; Adrian-Kalchhauser, I.; Burkhardt-Holm, P.; White, W.T.; Brunnschweiler, J.M. Characteristics of the shark fisheries of Fiji. Sci. Rep. 2015, 5, 17556. [Google Scholar] [CrossRef]
- Abrantes, K.G.; Brunnschweiler, J.M.; Barnett, A. You are what you eat: Examining the effects of provisioning tourism on shark diets. Biol. Conserv. 2018, 224, 300–308. [Google Scholar] [CrossRef]
- Vierus, T.; Gehrig, S.; Brunnschweiler, J.M.; Glaus, K.; Zimmer, M.; Marie, A.D.; Rico, C. Discovery of a multispecies shark aggregation and parturition area in the Ba Estuary, Fiji Islands. Ecol. Evol. 2018, 8, 7079–7093. [Google Scholar] [CrossRef]
- Brunnschweiler, J.M.; Marosi, N.D. Two years of impairment: Plastic packing strap on a bull shark (Carcharhinus leucas) in Fiji. Pac. Conserv. Biol. 2019, 26, 208–209. [Google Scholar] [CrossRef]
- Glaus, K.B.; Appleyard, S.A.; Stockwell, B.; Brunnschweiler, J.M.; Shivji, M.; Clua, E.; Marie, A.D.; Rico, C. Insights into Insular Isolation of the Bull Shark, Carcharhinus leucas (Müller and Henle, 1839), in Fijian Waters. Front. Mar. Sci. 2020, 7, 586015. [Google Scholar] [CrossRef]
- Bouveroux, T.; Loiseau, N.; Barnett, A.; Marosi, N.D.; Brunnschweiler, J.M. Companions and casual acquaintances: The nature of associations among bull sharks at a shark feeding site in Fiji. Front. Mar. Sci. 2021, 8, 678074. [Google Scholar] [CrossRef]
- Glaus, K.; Genter, F.; Brunnschweiler, J.M. Red eyes in juvenile bull sharks (Carcharhinus leucas) from Fiji. Pac. Conserv. Biol. 2023, 30, PC2300. [Google Scholar] [CrossRef]
- Séret, B.; Last, P.R. New deep water skates of the genus Notoraja Ishiyama, 1958 (Rajoidei, Arhynchobatidae) from the southwest Pacific. Zoosystema 2012, 34, 319–341. [Google Scholar] [CrossRef]
- Murphy, S.E.; Campbell, I.; Drew, J.A. Examination of tourists’ willingness to pay under different conservation scenarios; Evidence from reef manta ray snorkeling in Fiji. PLoS ONE 2018, 13, e0198279. [Google Scholar] [CrossRef]
- Gordon, L.; Vierus, T. First photographic evidence of oceanic manta rays (Mobula birostris) at two locations in the Fiji islands. PeerJ 2022, 10, e13883. [Google Scholar] [CrossRef]
- Hind-Ozan, E.J.; Pecl, G.T.; Ward-Paige, C.A. Communication and trust-building with the broader public through coastal and marine citizen science. In Citizen Science for Coastal and Marine Conservation; Routledge: Oxfordshire, UK, 2017; pp. 261–278. [Google Scholar]
- Ward-Paige, C.A.; Sykes, H.; Osgood, G.; Brunnschweiler, J. Community-driven shark monitoring for informed decision making: A case study from Fiji. Pac. Conserv. Biol. 2022, 29, 402–418. [Google Scholar] [CrossRef]
- Ward-Paige, C.A. The role of the tourism industry. In Sharks: Conservation, Governance and Management; Routledge: Oxfordshire, UK, 2014; pp. 157–175. [Google Scholar]
- Cisneros-Montemayor, A.M.; Townsel, A.; Gonzales, C.M.; Haas, A.R.; Navarro-Holm, E.E.; Salorio-Zuñiga, T.; Johnson, A.F. Nature-based marine tourism in the Gulf of California and Baja California Peninsula: Economic benefits and key species. Nat. Resour. Forum 2020, 44, 111–128. [Google Scholar] [CrossRef]
- Mustika, P.L.K.; Ichsan, M.; Booth, H. The economic value of shark and ray tourism in Indonesia and its role in delivering conservation outcomes. Front. Mar. Sci. 2020, 7, 261. [Google Scholar] [CrossRef]
- Gray, C.; Peters-Burton, E.; Smith, C.; Parsons, E. Basking shark tourism in Donegal, Ireland—A case-study of public interest and support for shark conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 537–550. [Google Scholar] [CrossRef]
- Marie, A.D.; Miller, C.; Cawich, C.; Piovano, S.; Rico, C. Fisheries-independent surveys identify critical habitats for young scalloped hammerhead sharks (Sphyrna lewini) in the Rewa Delta, Fiji. Sci. Rep. 2017, 7, 17273. [Google Scholar] [CrossRef] [PubMed]
- Cardeñosa, D.; Glaus, K.B.; Brunnschweiler, J.M. Occurrence of juvenile bull sharks (Carcharhinus leucas) in the Navua River in Fiji. Mar. Freshw. Res. 2016, 68, 592–597. [Google Scholar] [CrossRef]
- Glaus, K.B.; Brunnschweiler, J.M.; Piovano, S.; Mescam, G.; Genter, F.; Fluekiger, P.; Rico, C. Essential waters: Young bull sharks in Fiji’s largest riverine system. Ecol. Evol. 2019, 9, 7574–7585. [Google Scholar] [CrossRef]
- Matheson, C.A. iNaturalist. Ref. Rev. 2014, 28, 36–38. [Google Scholar]
- Mesaglio, T.; Callaghan, C.T. An overview of the history, current contributions and future outlook of iNaturalist in Australia. Wildl. Res. 2021, 48, 289–303. [Google Scholar] [CrossRef]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Polanco Fernández, A.; Marques, V.; Fopp, F.; Juhel, J.B.; Borrero-Pérez, G.H.; Cheutin, M.C.; Dejean, T.; González Corredor, J.D.; Acosta-Chaparro, A.; Hocdé, R.; et al. Comparing environmental DNA metabarcoding and underwater visual census to monitor tropical reef fishes. Environ. DNA 2021, 3, 142–156. [Google Scholar] [CrossRef]
- United Nations Department of Economic and Social Affairs. Voluntary Commitments for the implementation of Sustainable Development Goal 14. In Proceedings of the 2017 United Nations Ocean Conference, New York, NY, USA, 5–9 June 2017. [Google Scholar]
- Lee, S.; Lewis, A.; Gillett, R.; Fox, M.; Tuqiri, N.; Sadovy, Y.; Batibasaga, A.; Lalavanua, W.; Lovell, E. Fiji Fishery Resource Profiles. Information for Management on 44 of the Most Important Species Groups; Gillett, Preston and Associates and the Wildlife Conservation Society: Suva, Fiji, 2018. [Google Scholar]
- Seeto, J.; Baldwin, W.J. A Checklist of the Fishes of Fiji and a Bibliography of Fijian Fish; Technical Report 1/2010; Division of Marine Studies, School of Islands and Oceans, Faculty of Science, The University of the South Pacific: Suva, Fiji, 2010. [Google Scholar]
- Gillett, R.; Lewis, A.; Cartwright, I. Coastal Fisheries in Fiji; Resources, Issues, & Enhancing Role of the Fisheries Department: Suva, Fiji, 2014. [Google Scholar]
- Thaman, R.; Fong, T.; Balawa, A. Qio Kei Na Vai-Sharks and Rays of Fiji; Institute of Applied Sciences, The University of the South Pacific: Suva, Fiji, 2020. [Google Scholar]
- Ostertagova, E.; Ostertag, O.; Kováč, J. Methodology and application of the Kruskal-Wallis test. Appl. Mech. Mater. 2014, 611, 115–120. [Google Scholar] [CrossRef]
- Longhurst, R. Semi-structured interviews and focus groups. Key Methods Geogr. 2003, 3, 143–156. [Google Scholar]
- Chapuis, S.M. Using semi-directive interview: An analysis of the power and social responsibility of researchers. In Proceedings of the 14th European Conference on Research Methodology for Business and Management Studies, Valletta, Malta, 11–12 June 2015; pp. 282–289. [Google Scholar]
- Singh, Y.N. Kava: An overview. J. Ethnopharmacol. 1992, 37, 13–45. [Google Scholar] [CrossRef]
- Piovano, S.; Basciano, G.; Swimmer, Y.; Giacoma, C. Evaluation of a bycatch reduction technology by fishermen: A case study from Sicily. Mar. Policy 2012, 36, 272–277. [Google Scholar] [CrossRef]
- Harris, P.; Brock, C.; McInnes, E.; Neill, B.; Diamond, A.; Carter, J.; Camaitoga, U.; Krishna, M.; Giannakis, E.; Harris, P.; et al. In Fiji. In Children’s Multilingual Literacy: Fostering Childhood Literacy in Home and Community Settings; Springer: Berlin/Heidelberg, Germany, 2020; pp. 11–39. [Google Scholar]
- Huntington, H.P. Observations on the utility of the semi-directive interview for documenting traditional ecological knowledge. ARCTIC 1998, 51, 237–242. [Google Scholar] [CrossRef]
- Stevens, G.; Fernando, D.; Di Sciara, G.N. Guide to the Manta and Devil Rays of the World; Princeton University Press: Princeton, NJ, USA, 2018; Volume 17. [Google Scholar]
- Glaus, K.B.J.; Adrian-Kalchhauser, I.; Piovano, S.; Appleyard, S.A.; Brunnschweiler, J.M.; Rico, C. Fishing for profit or food? Socio-economic drivers and fishers’ attitudes towards sharks in Fiji. Mar. Policy 2019, 100, 249–257. [Google Scholar] [CrossRef]
- Juhel, J.-B.; Utama, R.S.; Marques, V.; Vimono, I.B.; Sugeha, H.Y.; Kadarusman, n.; Pouyaud, L.; Dejean, T.; Mouillot, D.; Hocdé, R. Accumulation curves of environmental DNA sequences predict coastal fish diversity in the coral triangle. Proc. R. Soc. B 2020, 287, 20200248. [Google Scholar] [CrossRef] [PubMed]
- Collins, R.A.; Bakker, J.; Wangensteen, O.S.; Soto, A.Z.; Corrigan, L.; Sims, D.W.; Genner, M.J.; Mariani, S. Non-specific amplification compromises environmental DNA metabarcoding with COI. Methods Ecol. Evol. 2019, 10, 1985–2001. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, J.; Yao, M. A comprehensive and comparative evaluation of primers for metabarcoding eDNA from fish. Methods Ecol. Evol. 2020, 11, 1609–1625. [Google Scholar] [CrossRef]
- Polanco, F.A.; Richards, E.; Flück, B.; Valentini, A.; Altermatt, F.; Brosse, S.; Walser, J.C.; Eme, D.; Marques, V.; Manel, S.; et al. Comparing the performance of 12S mitochondrial primers for fish environmental DNA across ecosystems. Environ. DNA 2021, 3, 1113–1127. [Google Scholar] [CrossRef]
- Mathon, L.; Marques, V.; Mouillot, D.; Albouy, C.; Andrello, M.; Baletaud, F.; Borrero-Pérez, G.H.; Dejean, T.; Edgar, G.J.; Grondin, J.; et al. Cross-ocean patterns and processes in fish biodiversity on coral reefs through the lens of eDNA metabarcoding. Proc. R. Soc. B 2022, 289, 20220162. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2012, 41, D36–D42. [Google Scholar] [CrossRef]
- Madden, T. The BLAST sequence analysis tool. In The NCBI Handbook; U.S. Library of Medicine: Bethesda, MD, USA, 2003. [Google Scholar]
- Lawson, J.M.; Fordham, S.V.; O’Malley, M.P.; Davidson, L.N.; Walls, R.H.; Heupel, M.R.; Stevens, G.; Fernando, D.; Budziak, A.; Simpfendorfer, C.A.; et al. Sympathy for the devil: A conservation strategy for devil and manta rays. PeerJ 2017, 5, e3027. [Google Scholar] [CrossRef]
- Piovano, S.; Gilman, E. Elasmobranch captures in the Fijian pelagic longline fishery. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 381–393. [Google Scholar] [CrossRef]
- Manjaji Matsumoto, B.M.; White, W.T.; Fahmi Ishihara, H.; Morgan, D.L. Urogymnus granulatus (Amended Version of 2016 Assessment). The IUCN Red List of Threatened Species 2020: e.T161431A177282313. Available online: https://www.iucnredlist.org/species/161431/177282313 (accessed on 26 July 2023).
- Food and Agriculture Organization of the United Nations. The Living Marine Resources of the Western Central Pacific. Batoid Fishes, Chimeras and Bony Fishes Part 1 (Elopidae to Linophrynidae); Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; Available online: https://www.fao.org/publications/card/fr/c/b0b92ebe-a54a-527e-b89a-d539c925de18/ (accessed on 26 July 2023).
- Uluivuda, J. Endangered and Protected Species Act 2002. In Enacted by the Parliament of the Fiji Islands; Parliament of Fiji: Suva, Fiji, 2002; Volume 23. [Google Scholar]
- Thaman, R.; Fong, T.; Balawa, A. Finfishes of Vanua Navakavu; The University of the South Pacific: Suva, Fiji, 2008. [Google Scholar]
- Seeto, J.; South, G. Marine Resources Inventory of the Fiji Islands; The University of the South Pacific: Suva, Fiji, 2010. [Google Scholar]
- Mangubhai, S.; Sykes, H.; Lovell, E.; Brodie, G.; Jupiter, S.; Morris, C.; Lee, S.; Loganimoce, E.M.; Rashni, B.; Lal, R.; et al. Fiji: Coastal and marine ecosystems. In World Seas: An Environmental Evaluation; Elsevier: Amsterdam, The Netherlands, 2019; pp. 765–792. [Google Scholar]
- Andrew, P. Dreketi River and Estuary Shark and Ray Survey; WWF-Pacific: Suva, Fiji, 2020. [Google Scholar]
- Earle, J.L.; Whitton, R.K.; Greene, B.D.; Pyle, R.L. Report on the Fishes of Shark Reef Marine Reserve, Beqa Passage, Fiji, with a Qualitative Comparison of Fish Assemblages before and after the COVID-19 Global Pandemic; Bernice Pauahi Bishop Museum: Honolulu, HI, USA, 2023. [Google Scholar]
- JICA. Urotrygon Daviesi—Fiji or Tuvalu; JICA: Tokyo, Japan, 1987; p. 111.
- Duffy, C.; Seeto, J.; Trnski, T. Review of records of sawfish (Chondrichthyes: Pristidae) from Fiji, with deletion of Pristis zijsron Bleeker, 1851 and Pristis sp. from the fauna. Zootaxa 2011, 3115, 65–67. [Google Scholar] [CrossRef]
- Hylton, S.; White, W.; Chin, A. The sharks and rays of the Solomon Islands: A synthesis of their biological diversity, values and conservation status. Pac. Conserv. Biol. 2017, 23, 324–334. [Google Scholar] [CrossRef]
- Aalbersberg, W.G.; Avosa, M.; James, R.; Kaluwin, C.; Lokani, P.; Opu, J.; Siwatibau, S.; Tuiwawa, M.; Waqa-Sakiti, H.F.; Tordoff, A. Ecosystem Profile: East Melanesian Islands Biodiversity Hotspot; The University of the South Pacific: Suva, Fiji, 2012. [Google Scholar]
- Devloo-Delva, F.; Burridge, C.; Kyne, P.; Brunnschweiler, J.; Chapman, D.; Charvet, P.; Chen, X.; Cliff, G.; Daly, R.; Drymon, M.; et al. From rivers to ocean basins: The role of ocean barriers and philopatry in the genetic structuring of a cosmopolitan coastal predator. Ecol. Evol. 2023, 13, e9837. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.D.; Nuttall, M.; Hickerson, E.L.; Johnston, M.A. Important juvenile manta ray habitat at Flower Garden Banks National Marine Sanctuary in the northwestern Gulf of Mexico. Mar. Biol. 2018, 165, 111. [Google Scholar]
- White, W.T.; Corrigan, S.; Yang, L.; Henderson, A.C.; Bazinet, A.L.; Swofford, D.L.; Naylor, G.J. Phylogeny of the manta and devilrays (Chondrichthyes: Mobulidae), with an updated taxonomic arrangement for the family. Zool. J. Linn. Soc. 2018, 182, 50–75. [Google Scholar] [CrossRef]
- White, W.T.; Last, P.R.; Naylor, G.J.P.; Jensen, K.; Caira, J.N. Clarification of Aetobatus ocellatus (Kuhl, 1823) as a valid species, and a comparison with Aetobatus narinari (Euphrasen, 1790) (Rajiformes: Myliobatidae). Descriptions of new sharks and rays from Borneo. CSIRO Mar. Atmos. Res. Pap. 2010, 32, 141–164. [Google Scholar]
- White, W.T. A revised generic arrangement for the eagle ray family Myliobatidae, with definitions for the valid genera. Zootaxa 2014, 3860, 149–166. [Google Scholar] [CrossRef]
- White, W.T.; Naylor, G. Resurrection of the family Aetobatidae (Myliobatiformes) for the pelagic eagle rays, genus Aetobatus. Zootaxa 2016, 4139, 435–438. [Google Scholar] [CrossRef]
- Berthe, C.; Mourier, J.; Lecchini, D.; Rummer, J.L.; Sellos, D.Y.; Iglesias, S.P. DNA barcoding supports the presence of the cryptic ocellated eagle ray, Aetobatus ocellatus (Myliobatidae), in French Polynesia, South Pacific. Cybium 2016, 40, 181–184. [Google Scholar]
- Ratnasingham, S.; Hebert, P.D. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Schluessel, V.; Broderick, D.; Collin, S.; Ovenden, J. Evidence for extensive population structure in the white-spotted eagle ray within the Indo-Pacific inferred from mitochondrial gene sequences. J. Zool. 2010, 281, 46–55. [Google Scholar] [CrossRef]
- Kyne, P.M.; Finucci, B. Taeniura lessoni. The IUCN Red List of Threatened Species; International Union for Conservation of Nature: Gland, Switzerland, 2018; p. E.T116855477A116855482. [CrossRef]
- Last, P.R.; White, W.T.; Seret, B. Taxonomic status of maskrays of the Neotrygon kuhlii species complex (Myliobatoidei: Dasyatidae) with the description of three new species from the Indo-West Pacific. Zootaxa 2016, 4083, 533–561. [Google Scholar] [CrossRef] [PubMed]
- Puckridge, M.; Last, P.R.; White, W.T.; Andreakis, N. Phylogeography of the Indo-West Pacific maskrays (Dasyatidae, Neotrygon): A complex example of chondrichthyan radiation in the Cenozoic. Ecol. Evol. 2013, 3, 217–232. [Google Scholar] [CrossRef]
- Arlyza, I.S.; Shen, K.-N.; Durand, J.-D.; Borsa, P. Mitochondrial haplotypes indicate parapatric-like phylogeographic structure in blue-spotted maskray (Neotrygon kuhlii) from the Coral Triangle region. J. Hered. 2013, 104, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Stewart, K.A. Understanding the effects of biotic and abiotic factors on sources of aquatic environmental DNA. Biodivers. Conserv. 2019, 28, 983–1001. [Google Scholar] [CrossRef]
- Rourke, M.L.; Fowler, A.M.; Hughes, J.M.; Broadhurst, M.K.; DiBattista, J.D.; Fielder, S.; Wilkes Walburn, J.; Furlan, E.M. Environmental DNA (eDNA) as a tool for assessing fish biomass: A review of approaches and future considerations for resource surveys. Environ. DNA 2022, 4, 9–33. [Google Scholar] [CrossRef]
- Fernandez-Carvalho, J.; Imhoff, J.L.; Faria, V.V.; Carlson, J.K.; Burgess, G.H. Status and the potential for extinction of the largetooth sawfish Pristis pristis in the Atlantic Ocean. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 478–497. [Google Scholar] [CrossRef]
- Everett, B.; Cliff, G.; Dudley, S.; Wintner, S.; Van der Elst, R. Do sawfish Pristis spp. represent South Africa’s first local extirpation of marine elasmobranchs in the modern era? Afr. J. Mar. Sci. 2015, 37, 275–284. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Davidson, L.N.; Kyne, P.M.; Simpfendorfer, C.A.; Harrison, L.R.; Carlson, J.K.; Fordham, S.V. Ghosts of the coast: Global extinction risk and conservation of sawfishes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 134–153. [Google Scholar] [CrossRef]
- The Offshore Fisheries Management Act. 2012. Available online: https://www.laws.gov.fj/Acts/DisplayAct/2286 (accessed on 26 July 2023).
- Veitayaki, J.; Nakoro, A.D.; Sigarua, T.; Bulai, N. On cultural factors and marine managed areas in Fiji. In Pacific Island Heritage Archaeology, Identity and Community; ANU Press: Canberra, Australia, 2011; pp. 37–49. Available online: https://mcst-rmiusp.org/index.php/reference-library-main?task=download.send&id=763&catid=6&m=0 (accessed on 26 July 2023).
- Breckwoldt, A.; Seidel, H. The need to know what to manage—Community-based marine resource monitoring in Fiji. Curr. Opin. Environ. Sustain. 2012, 4, 331–337. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Family | Scientific Name | IUCN Status | CITES | CMS | Fiji EPS | Methodology Recorded | Previous Classification | ||
|---|---|---|---|---|---|---|---|---|---|---|
| - | - | Literature Review | Participatory Science | eDNA | ||||||
| Maskray | Dasyatidae | Neotrygon sp. | NA |  |  | | | Dasyatis kuhlii Amphotistius kuhlii | ||
| Porcupine ray | Dasyatidae | Urogymnus asperrimus | Vulnerable | | | | ||||
| Mangrove whipray | Dasyatidae | Urogymnus granulatus | Vulnerable | | Himantura granulatus | |||||
| Pink whipray | Dasyatidae | Pateobatis fai | Vulnerable | | | | Himantura fai | |||
| Oceania fantail ray | Dasyatidae | Taeniura lessoni | Data Deficient | | | | Taeniura lymma | |||
| Blotched fantail ray | Dasyatidae | Taeniurops meyeni | Least Concern | | | Taeniura meyeni | ||||
| Pelagic stingray | Dasyatidae | Pteroplatytrygon violacea | Least Concern | | Dasyatis violacea | |||||
| Broad cowtail ray | Dasyatidae | Pastinachus ater | Vulnerable | | ||||||
| Spotted eagle ray | Aetobatidae | Aetobatus ocellatus | Vulnerable | | | | Aetobatus narinari | |||
| Oceanic manta ray | Mobulidae | Mobula birostris | Endangered | Appendix II | Appendices I & II | | | | Manta birostris | |
| Reef manta ray | Mobulidae | Mobula alfredi | Vulnerable | Appendix II | Appendices I & II | | | | Manta alfredi | |
| Sicklefin devil ray | Mobulidae | Mobula tarapacana | Endangered | Appendix II | Appendices I & II | | | | ||
| Bentfin devil ray | Mobulidae | Mobula thurstoni | Endangered | Appendix II | Appendices I & II | | | | ||
| Spinetail devil ray | Mobulidae | Mobula mobular | Endangered | Appendix II | Appendices I & II | | | | ||
| Fijian velvet skate | Arhynchobatidae | Notoraja fijiensis | Least Concern | | | |||||
| Longlobe velvet skate | Arhynchobatidae | Notoraja longiventralis | Least Concern | | ||||||
| Bottlenose wedgefish | Rhinidae | Rhynchobatus australiae | Critically Endangered | Appendix II | Appendix II | | | | | Rhynchobatus djiddensis |
| Giant stingaree | Plesiobatidae | Plesiobatis daviesi | Least Concern | | ||||||
| Giant guitarfish | Glaucostegidae | Glaucostegus typus | Critically Endangered | Appendix II | | | Rhinobatus batillum | |||
| Sawfishes * | Pristidae | Pristis sp. | Critically Endangered | Appendix I | Appendices I & II | | | | Pristis zijsron Pristis microdon Pristis sp. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glaus, K.; Gordon, L.; Vierus, T.; Marosi, N.D.; Sykes, H. Rays in the Shadows: Batoid Diversity, Occurrence, and Conservation Status in Fiji. Biology 2024, 13, 73. https://doi.org/10.3390/biology13020073
Glaus K, Gordon L, Vierus T, Marosi ND, Sykes H. Rays in the Shadows: Batoid Diversity, Occurrence, and Conservation Status in Fiji. Biology. 2024; 13(2):73. https://doi.org/10.3390/biology13020073
Chicago/Turabian StyleGlaus, Kerstin, Luke Gordon, Tom Vierus, Natasha D. Marosi, and Helen Sykes. 2024. "Rays in the Shadows: Batoid Diversity, Occurrence, and Conservation Status in Fiji" Biology 13, no. 2: 73. https://doi.org/10.3390/biology13020073
APA StyleGlaus, K., Gordon, L., Vierus, T., Marosi, N. D., & Sykes, H. (2024). Rays in the Shadows: Batoid Diversity, Occurrence, and Conservation Status in Fiji. Biology, 13(2), 73. https://doi.org/10.3390/biology13020073