
Allocation of Nutrients and Leaf Turnover Rate in Poplar under Ambient and Enriched Ozone Exposure and Soil Nutrient Manipulation




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Sampling and Measurements
2.3. Statistical Analyses
3. Results
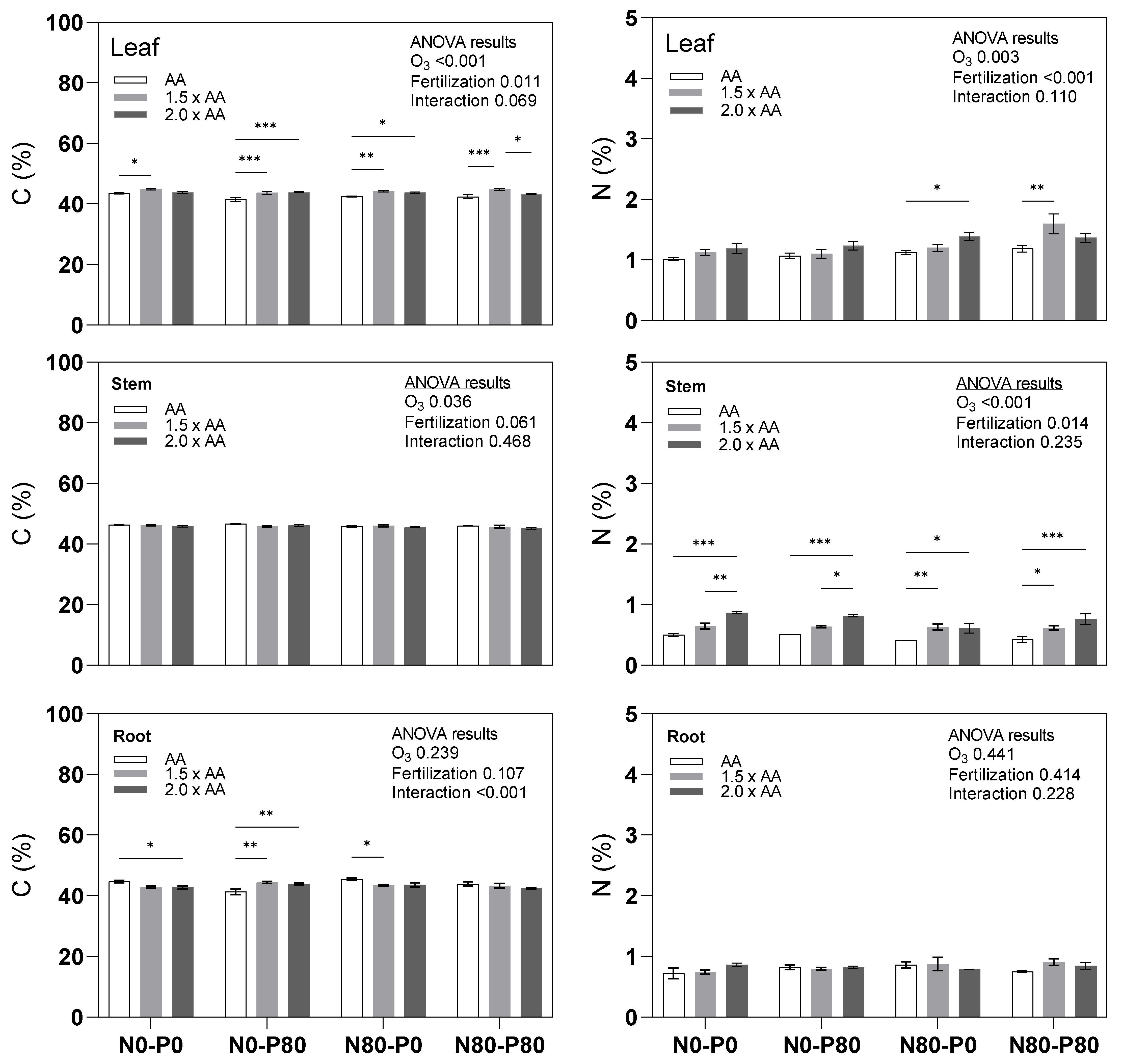
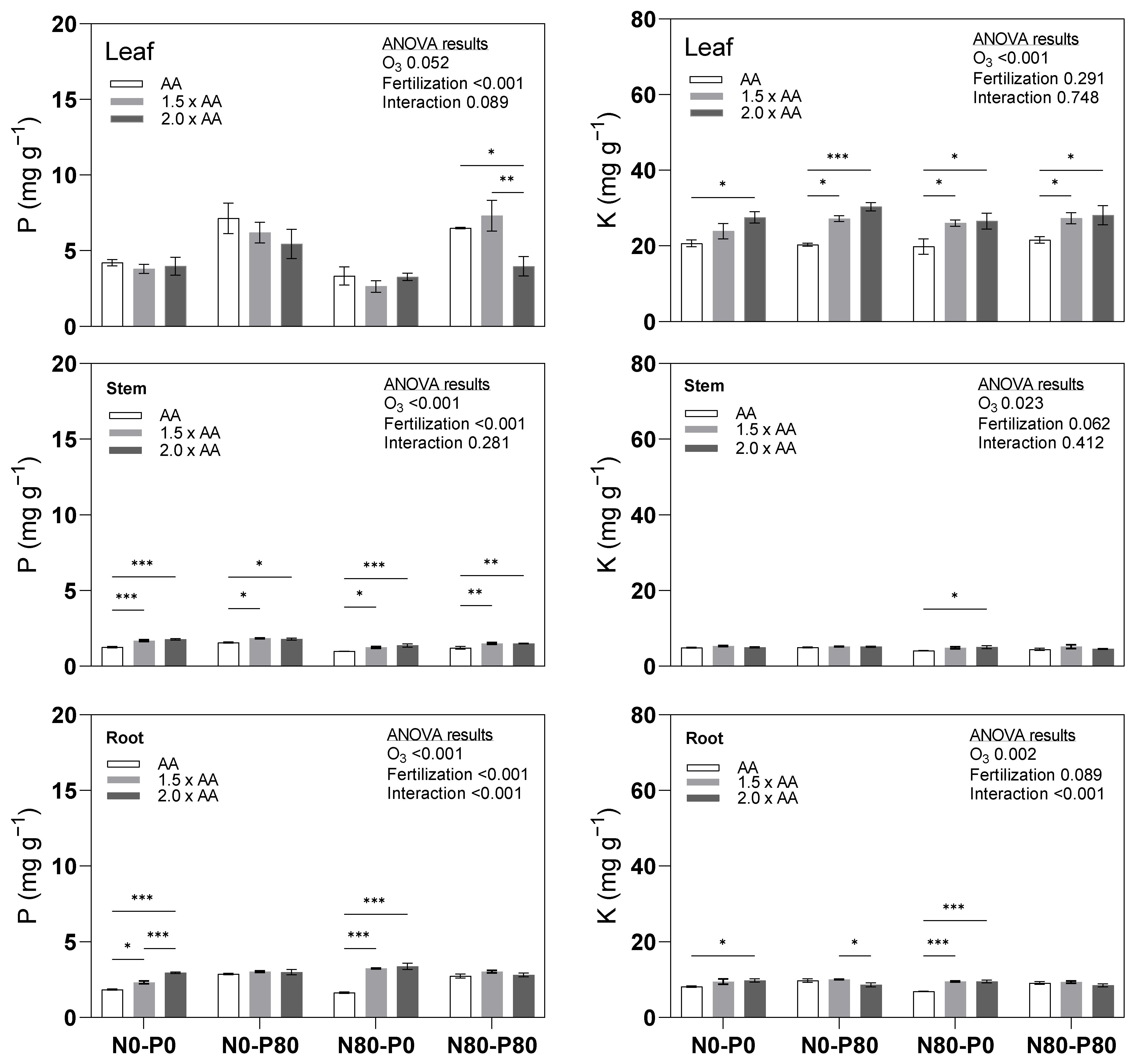
3.1. Primary Macronutrients (C, N, P, K)
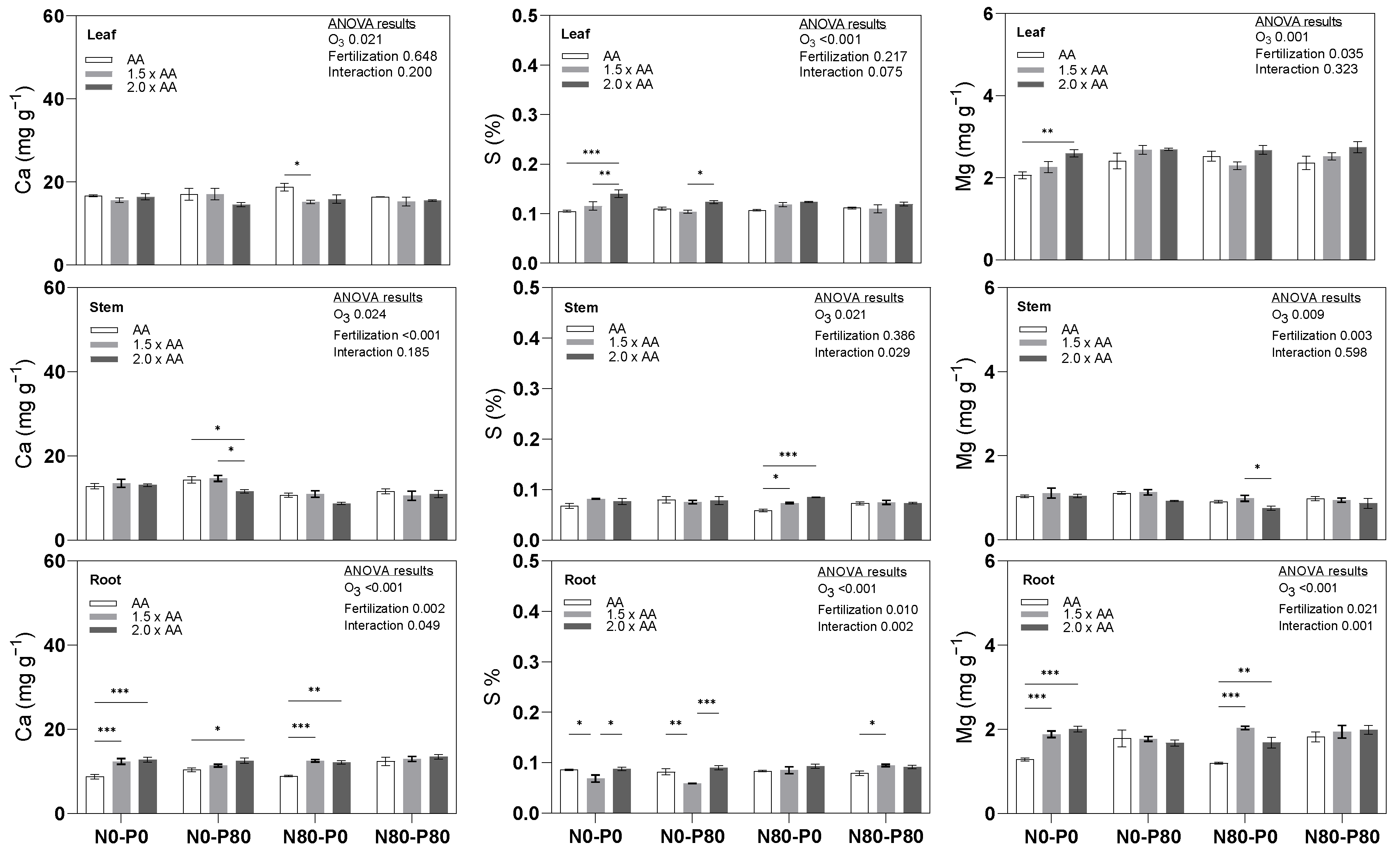
3.2. Secondary Macronutrients (Ca, S, Mg)
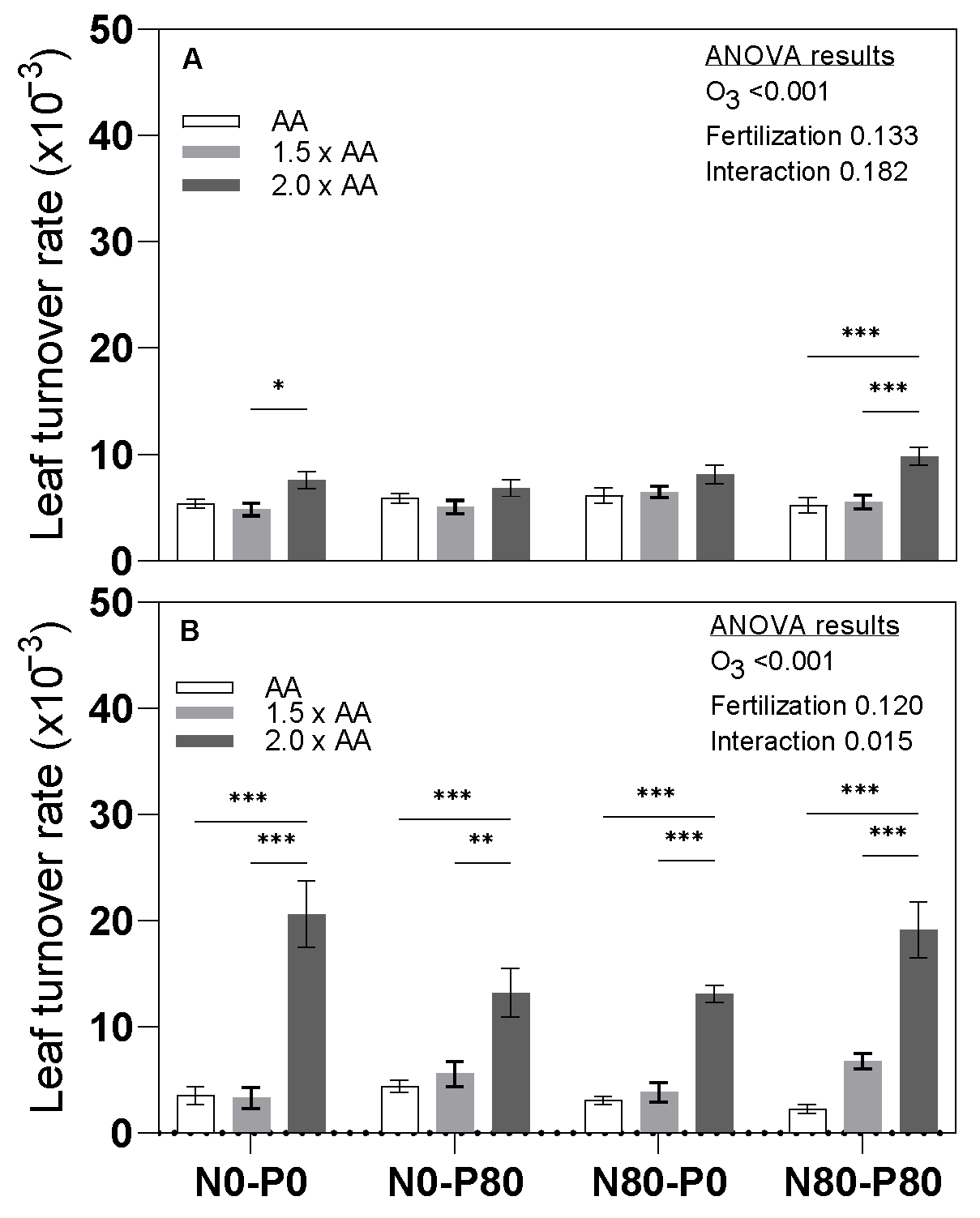
3.3. Leaf Turnover Rate
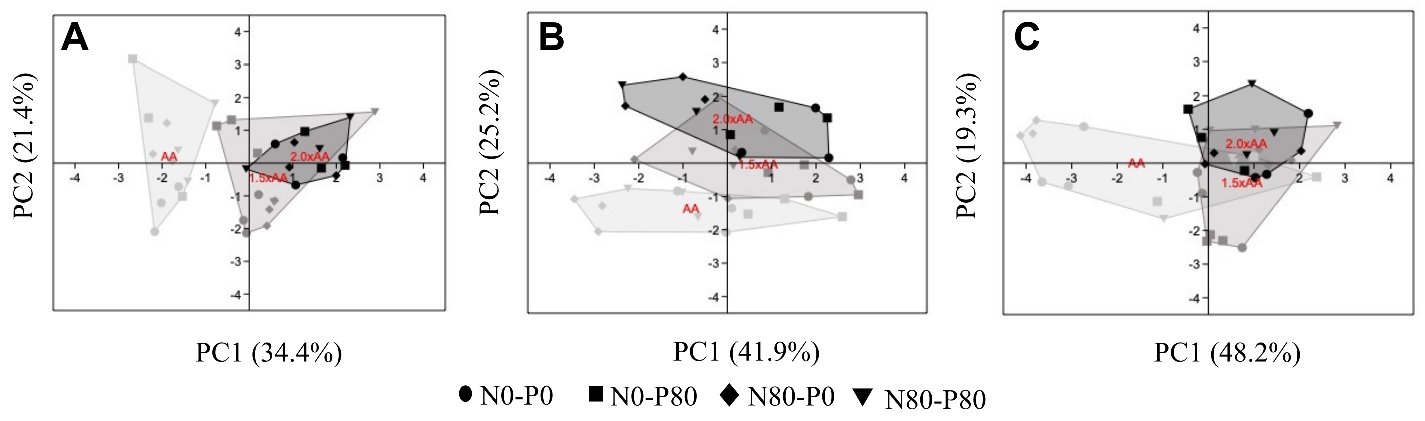
3.4. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ågren, G.I.; Weih, M. Multi-Dimensional Plant Element Stoichiometry—Looking beyond Carbon, Nitrogen, and Phosphorus. Front. Plant Sci. 2020, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Wang, H.; Sterner, R.W.; Elser, J.J. On the “Strict Homeostasis” Assumption in Ecological Stoichiometry. Ecol. Model. 2012, 243, 81–88. [Google Scholar] [CrossRef]
- Minden, V.; Kleyer, M. Internal and External Regulation of Plant Organ Stoichiometry. Plant Biol. 2014, 16, 897–907. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, N.; Liu, C.; Yang, H.; Li, M.; Yu, G.; Wilcox, K.; Yu, Q.; He, N. C:N:P Stoichiometry in China’s Forests: From Organs to Ecosystems. Funct. Ecol. 2018, 32, 50–60. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F. Global Assessment of Nitrogen Deposition Effects on Terrestrial Plant Diversity: A Synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed]
- Sicard, P.; Anav, A.; De Marco, A.; Paoletti, E. Projected Global Ground-Level Ozone Impacts on Vegetation under Different Emission and Climate Scenarios. Atmos. Chem. Phys. 2017, 17, 12177–12196. [Google Scholar] [CrossRef]
- Elser, J.; Bennett, E. A Broken Biogeochemical Cycle. Nature 2011, 478, 29–31. [Google Scholar] [CrossRef]
- van Vuuren, D.P.; Bouwman, L.F.; Smith, S.J.; Dentener, F. Global Projections for Anthropogenic Reactive Nitrogen Emissions to the Atmosphere: An Assessment of Scenarios in the Scientific Literature. Curr. Opin. Environ. Sustain. 2011, 3, 359–369. [Google Scholar] [CrossRef]
- Fenn, M.E.; Baron, J.S.; Allen, E.B.; Rueth, H.M.; Nydick, K.R.; Geiser, L.; Bowman, W.D.; Sickman, J.O.; Meixner, T.; Johnson, D.W. Ecological Effects of Nitrogen Deposition in the Western United States. BioScience 2003, 53, 404–420. [Google Scholar] [CrossRef]
- Peng, Y.; Chen, G.; Chen, G.; Li, S.; Peng, T.; Qiu, X.; Luo, J.; Yang, S.; Hu, T.; Hu, H. Soil Biochemical Responses to Nitrogen Addition in a Secondary Evergreen Broad-Leaved Forest Ecosystem. Sci. Rep. 2017, 7, 2783. [Google Scholar] [CrossRef]
- Ollinger, S.V.; Aber, J.D.; Reich, P.B.; Freuder, R.J. Interactive Effects of Nitrogen Deposition, Tropospheric Ozone, Elevated CO2 and Land Use History on the Carbon Dynamics of Northern Hardwood Forests. Glob. Chang. Biol. 2002, 8, 545–562. [Google Scholar] [CrossRef]
- Zhang, L.; Hoshika, Y.; Carrari, E.; Badea, O.; Paoletti, E. Ozone Risk Assessment Is Affected by Nutrient Availability: Evidence from a Simulation Experiment under Free Air Controlled Exposure (FACE). Environ. Pollut. 2018, 238, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; Van Der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J. Human-Induced Nitrogen–Phosphorus Imbalances Alter Natural and Managed Ecosystems across the Globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, F.J.; Cole, M.A. Cycles of Soils: Carbon, Nitrogen, Phosphorus, Sulfur, Micronutrients; John Wiley & Sons: Hoboken, NJ, USA, 1999; ISBN 0-471-32071-4. [Google Scholar]
- Wittig, V.E.; Ainsworth, E.A.; Naidu, S.L.; Karnosky, D.F.; Long, S.P. Quantifying the Impact of Current and Future Tropospheric Ozone on Tree Biomass, Growth, Physiology and Biochemistry: A Quantitative Meta-analysis. Glob. Chang. Biol. 2009, 15, 396–424. [Google Scholar] [CrossRef]
- Shang, B.; Feng, Z.; Li, P.; Calatayud, V. Elevated Ozone Affects C, N and P Ecological Stoichiometry and Nutrient Resorption of Two Poplar Clones. Environ. Pollut. 2018, 234, 136–144. [Google Scholar] [CrossRef]
- Agathokleous, E.; Kitao, M.; Qingnan, C.; Saitanis, C.J.; Paoletti, E.; Manning, W.J.; Watanabe, T.; Koike, T. Effects of Ozone (O3) and Ethylenediurea (EDU) on the Ecological Stoichiometry of a Willow Grown in a Free-Air Exposure System. Environ. Pollut. 2018, 238, 663–676. [Google Scholar] [CrossRef]
- Valkama, E.; Koricheva, J.; Oksanen, E. Effects of Elevated O3, Alone and in Combination with Elevated CO2, on Tree Leaf Chemistry and Insect Herbivore Performance: A Meta-analysis. Glob. Chang. Biol. 2007, 13, 184–201. [Google Scholar] [CrossRef]
- Shi, C.; Watanabe, T.; Koike, T. Leaf Stoichiometry of Deciduous Tree Species in Different Soils Exposed to Free-Air O3 Enrichment over Two Growing Seasons. Environ. Exp. Bot. 2017, 138, 148–163. [Google Scholar] [CrossRef]
- Zhuang, M.; Li, Y.; Guo, Z.; Li, Y.; Pan, W.; Chen, S. Elevated CO2 and O3 Levels Influence the Uptake and Leaf Concentration of Mineral N, P, K in Phyllostachys Edulis (Carrière) J. Houz. and Oligostachyum Lubricum (Wen) King f. Forests 2018, 9, 195. [Google Scholar]
- Cao, J.; Shang, H.; Chen, Z.; Tian, Y.; Yu, H. Effects of Elevated Ozone on Stoichiometry and Nutrient Pools of Phoebe Bournei (Hemsl.) Yang and Phoebe Zhennan S. Lee et FN Wei Seedlings in Subtropical China. Forests 2016, 7, 78. [Google Scholar]
- Kinose, Y.; Fukamachi, Y.; Okabe, S.; Hiroshima, H.; Watanabe, M.; Izuta, T. Nutrient Supply to Soil Offsets the Ozone-Induced Growth Reduction in Fagus Crenata Seedlings. Trees 2017, 31, 259–272. [Google Scholar] [CrossRef]
- You, C.; Peng, C.; Xu, Z.; Liu, Y.; Zhang, L.; Yin, R.; Liu, L.; Li, H.; Wang, L.; Liu, S. Nitrogen Addition Mediates the Response of Foliar Stoichiometry to Phosphorus Addition: A Meta-Analysis. Ecol. Process. 2021, 10, 58. [Google Scholar] [CrossRef]
- Tian, D.; Reich, P.B.; Chen, H.Y.; Xiang, Y.; Luo, Y.; Shen, Y.; Meng, C.; Han, W.; Niu, S. Global Changes Alter Plant Multi-element Stoichiometric Coupling. New Phytol. 2019, 221, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Bongers, F.; Popma, J. Leaf Dynamics of Seedlings of Rain Forest Species in Relation to Canopy Gaps. Oecologia 1990, 82, 122–127. [Google Scholar] [CrossRef]
- Aerts, R. The Effect of Increased Nutrient Availability on Leaf Turnover and Aboveground Productivity of Two Evergreen Ericaceous Shrubs. Oecologia 1989, 78, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Christersson, L. Wood Production Potential in Poplar Plantations in Sweden. Biomass Bioenergy 2010, 34, 1289–1299. [Google Scholar] [CrossRef]
- Pollastrini, M.; Desotgiu, R.; Camin, F.; Ziller, L.; Gerosa, G.; Marzuoli, R.; Bussotti, F. Severe Drought Events Increase the Sensitivity to Ozone on Poplar Clones. Environ. Exp. Bot. 2014, 100, 94–104. [Google Scholar] [CrossRef]
- Hoshika, Y.; Carrari, E.; Zhang, L.; Carriero, G.; Pignatelli, S.; Fasano, G.; Materassi, A.; Paoletti, E. Testing a Ratio of Photosynthesis to O3 Uptake as an Index for Assessing O3-Induced Foliar Visible Injury in Poplar Trees. Environ. Sci. Pollut. Res. 2018, 25, 8113–8124. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, E.; Materassi, A.; Fasano, G.; Hoshika, Y.; Carriero, G.; Silaghi, D.; Badea, O. A New-Generation 3D Ozone FACE (Free Air Controlled Exposure). Sci. Total Environ. 2017, 575, 1407–1414. [Google Scholar] [CrossRef]
- Thomas, R.; Lewis, J.; Strain, B. Effects of Leaf Nutrient Status on Photosynthetic Capacity in Loblolly Pine (Pinus taeda L.) Seedlings Grown in Elevated Atmospheric CO2. Tree Physiol. 1994, 14, 947–960. [Google Scholar]
- Lewis, J.; Strain, B. The Role of Mycorrhizas in the Response of Pinus Taeda Seedlings to Elevated CO2. New Phytol. 1996, 133, 431–443. [Google Scholar] [CrossRef]
- Mao, Q.; Watanabe, M.; Makoto, K.; Kita, K.; Koike, T. High Nitrogen Deposition May Enhance Growth of a New Hybrid Larch F1 Growing at Two Phosphorus Levels. Landsc. Ecol. Eng. 2014, 10, 1–8. [Google Scholar] [CrossRef]
- Nomura, M.; Hatada, A.; Itioka, T. Correlation between the Leaf Turnover Rate and Anti-Herbivore Defence Strategy (Balance between Ant and Non-Ant Defences) amongst Ten Species of Macaranga (Euphorbiaceae). Plant Ecol. 2011, 212, 143–155. [Google Scholar] [CrossRef]
- Cools, N.; De Vos, B. Sampling and Analysis of Soil. Manual Part X. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE, ICP Forests: Hamburg, Germany, 2010; Volume 208, pp. 536–542. [Google Scholar]
- Podda, A.; Pisuttu, C.; Hoshika, Y.; Pellegrini, E.; Carrari, E.; Lorenzini, G.; Nali, C.; Cotrozzi, L.; Zhang, L.; Baraldi, R. Can Nutrient Fertilization Mitigate the Effects of Ozone Exposure on an Ozone-Sensitive Poplar Clone? Sci. Total Environ. 2019, 657, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Shang, B.; Li, Z.; Yuan, X.; Xu, Y.; Feng, Z. Effects of Elevated Ozone on the Uptake and Allocation of Macronutrients in Poplar Saplings Above- and Belowground. Sci. Total Environ. 2022, 851, 158044. [Google Scholar] [CrossRef] [PubMed]
- Lazar, T.; Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Wiley: Hoboken, NJ, USA, 2003. [Google Scholar]
- Hoshika, Y.; Cotrozzi, L.; Gavrichkova, O.; Nali, C.; Pellegrini, E.; Scartazza, A.; Paoletti, E. Functional Responses of Two Mediterranean Pine Species in an Ozone Free-Air Controlled Exposure (FACE) Experiment. Tree Physiol. 2023, 43, 1548–1561. [Google Scholar] [CrossRef] [PubMed]
- Shang, B.; Xu, Y.; Dai, L.; Yuan, X.; Feng, Z. Elevated Ozone Reduced Leaf Nitrogen Allocation to Photosynthesis in Poplar. Sci. Total Environ. 2019, 657, 169–178. [Google Scholar] [CrossRef]
- Ping, Q.; Fang, C.; Yuan, X.; Agathokleous, E.; He, H.; Zheng, H.; Feng, Z. Nitrogen Addition Changed the Relationships of Fine Root Respiration and Biomass with Key Physiological Traits in Ozone-Stressed Poplars. Sci. Total Environ. 2023, 875, 162721. [Google Scholar] [CrossRef]
- Leone, I.A. Response of Potassium-Deficient Tomato Plants to Atmospheric Ozone. Phytopathology 1976, 66, 734–736. [Google Scholar] [CrossRef]
- Pleijel, H.; Eriksen, A.B.; Danielsson, H.; Bondesson, N.; Selldén, G. Differential Ozone Sensitivity in an Old and a Modern Swedish Wheat Cultivar—Grain Yield and Quality, Leaf Chlorophyll and Stomatal Conductance. Environ. Exp. Bot. 2006, 56, 63–71. [Google Scholar] [CrossRef]
- Xu, Y.; Feng, Z.; Peng, J.; Tarvainen, L. Elevated Ozone Decreases the Activity of Rubisco in Poplar but Not Its Activation under Fluctuating Light. Tree Physiol. 2022, 42, 1762–1775. [Google Scholar] [CrossRef]
- Paoletti, E.; Nali, C.; Lorenzini, G. Photosynthetic Behaviour of Two Italian Clones of European Beech (Fagus sylvatica L.) Exposed to Ozone. Phyton 2002, 42, 149–155. [Google Scholar]
- Li, Q.; Gao, Y.; Yang, A. Sulfur Homeostasis in Plants. Int. J. Mol. Sci. 2020, 21, 8926. [Google Scholar] [CrossRef]
- Piikki, K.; Vorne, V.; Ojanperä, K.; Pleijel, H. Impact of Elevated O3 and CO2 Exposure on Potato (Solanum tuberosum L. Cv. Bintje) Tuber Macronutrients (N, P, K, Mg, Ca). Agric. Ecosyst. Environ. 2007, 118, 55–64. [Google Scholar]
- Tjoelker, M.; Luxmoore, R. Soil Nitrogen and Chronic Ozone Stress Influence Physiology, Growth and Nutrient Status of Pinus taeda L. and Liriodendron tulipifera L. Seedlings. New Phytol. 1991, 119, 69–81. [Google Scholar] [CrossRef]
- Oksanen, E.; Holopainen, T. Responses of Two Birch (Betula Pendula Roth) Clones to Different Ozone Profiles with Similar AOT40 Exposure. Atmos. Environ. 2001, 35, 5245–5254. [Google Scholar] [CrossRef]
- Kitao, M.; Komatsu, M.; Yazaki, K.; Kitaoka, S.; Tobita, H. Growth Overcompensation against O3 Exposure in Two Japanese Oak Species, Quercus Mongolica Var. Crispula and Quercus Serrata, Grown under Elevated CO2. Environ. Pollut. 2015, 206, 133–141. [Google Scholar]
- Uddling, J.; Karlsson, P.E.; Glorvigen, A.; Selldén, G. Ozone Impairs Autumnal Resorption of Nitrogen from Birch (Betula pendula) Leaves, Causing an Increase in Whole-Tree Nitrogen Loss through Litter Fall. Tree Physiol. 2006, 26, 113–120. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paoletti, E.; Pagano, M.; Zhang, L.; Badea, O.; Hoshika, Y. Allocation of Nutrients and Leaf Turnover Rate in Poplar under Ambient and Enriched Ozone Exposure and Soil Nutrient Manipulation. Biology 2024, 13, 232. https://doi.org/10.3390/biology13040232
Paoletti E, Pagano M, Zhang L, Badea O, Hoshika Y. Allocation of Nutrients and Leaf Turnover Rate in Poplar under Ambient and Enriched Ozone Exposure and Soil Nutrient Manipulation. Biology. 2024; 13(4):232. https://doi.org/10.3390/biology13040232
Chicago/Turabian StylePaoletti, Elena, Mario Pagano, Lu Zhang, Ovidiu Badea, and Yasutomo Hoshika. 2024. "Allocation of Nutrients and Leaf Turnover Rate in Poplar under Ambient and Enriched Ozone Exposure and Soil Nutrient Manipulation" Biology 13, no. 4: 232. https://doi.org/10.3390/biology13040232
APA StylePaoletti, E., Pagano, M., Zhang, L., Badea, O., & Hoshika, Y. (2024). Allocation of Nutrients and Leaf Turnover Rate in Poplar under Ambient and Enriched Ozone Exposure and Soil Nutrient Manipulation. Biology, 13(4), 232. https://doi.org/10.3390/biology13040232