

Overnutrition and Lipotoxicity: Impaired Efferocytosis and Chronic Inflammation as Precursors to Multifaceted Disease Pathogenesis

Abstract
:Simple Summary
Abstract
1. Introduction
2. Overnutrition and Lipotoxicity
2.1. Excess Caloric Intake
2.2. Adipose Tissue Dysfunction
- Lipotoxicity in Other Tissues: Elevated levels of circulating free fatty acids can contribute to lipotoxic effects in various tissues like the liver, pancreas, and skeletal muscle (Figure 2). This lipotoxicity can impair cellular function and contribute to insulin resistance, inflammation, and other metabolic disturbances [35,36].
- Insulin Resistance: Excess fatty acids can disrupt insulin signaling, leading to insulin resistance. This condition impairs glucose uptake and metabolism, resulting in elevated blood sugar levels (hyperglycemia). Insulin resistance is a key factor in the development of metabolic disorders like type 2 diabetes, influenced by both overnutrition and lipotoxicity [37].
- Inflammation: Lipotoxicity is associated with an inflammatory response. Cytokines play a crucial role in inflammation, which is the body’s response to injury, infection, or other stimuli. They are small proteins secreted by various cells, including immune cells, and act as signaling molecules to regulate immune responses, inflammation, and other physiological processes [38]. In the context of inflammation, cytokines can be pro-inflammatory or anti-inflammatory, and their balance is essential for maintaining immune homeostasis. Pro-inflammatory cytokines promote inflammation by inducing vasodilation, increasing vascular permeability, recruiting immune cells to the site of injury or infection, and activating immune responses [39]. Examples include interleukin-1 (IL-1), interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), and interferon-gamma (IFN-γ) [40]. Pro-inflammatory cytokines play a critical role in the initial response to pathogens and tissue damage [39]. In contrast, anti-inflammatory cytokines help to resolve inflammation and maintain an immune balance by inhibiting pro-inflammatory responses and promoting tissue repair and regeneration. Examples include interleukin-10 (IL-10) and transforming growth factor-beta (TGF-β). Anti-inflammatory cytokines are essential for preventing excessive inflammation and tissue damage [41].
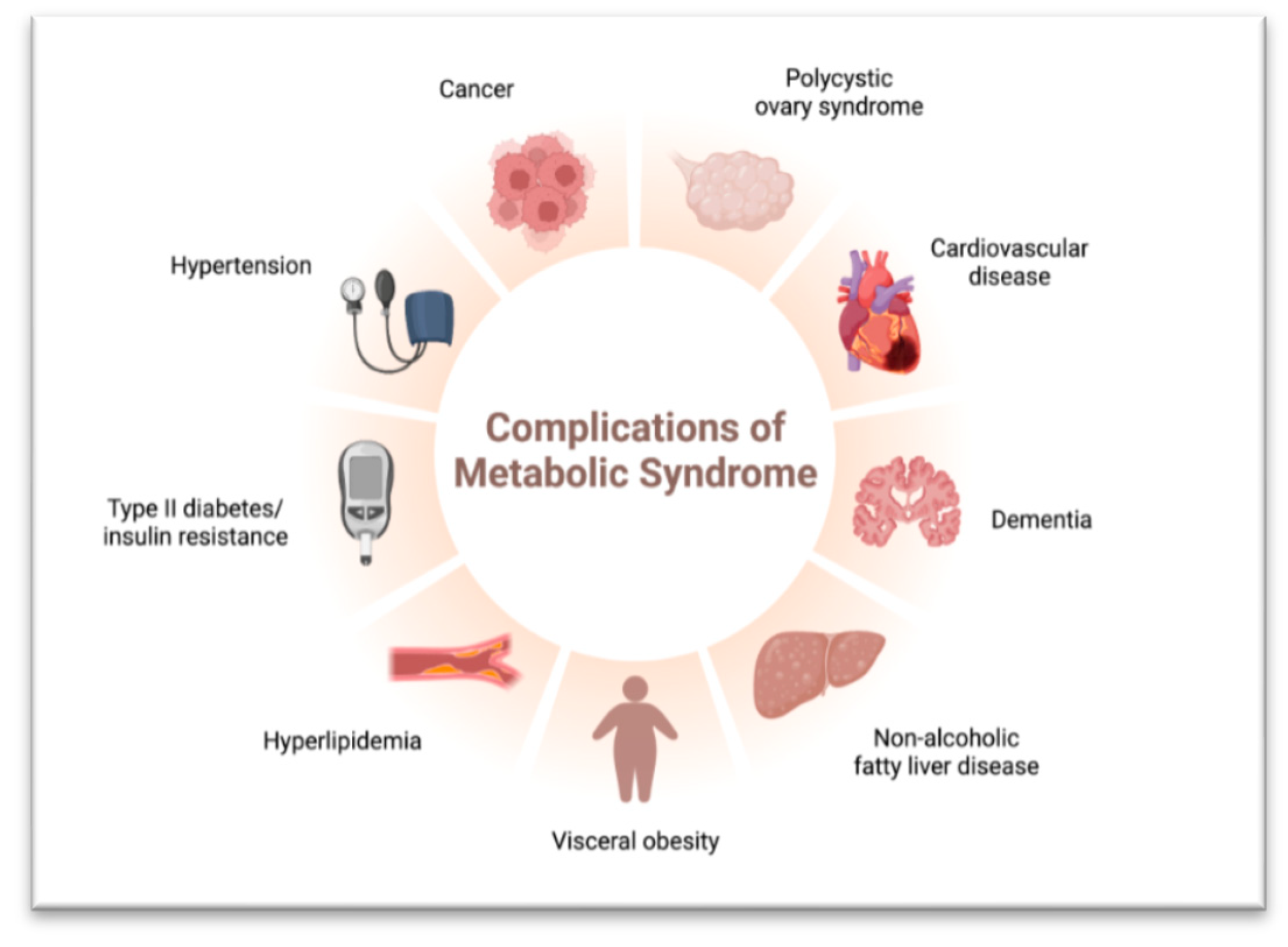
- Metabolic Syndrome: Overnutrition contributes to a cluster of conditions known as metabolic syndrome, including abdominal obesity, insulin resistance, high blood pressure, and dyslipidemia. Lipotoxicity exacerbates these conditions, leading to obesity, type 2 diabetes, and non-alcoholic fatty liver disease (NAFLD). Chronic inflammation further worsens metabolic dysfunction [55].
2.3. Impaired Efferocytosis/Oxidative Stress/Immune Response
3. Linking Impaired Efferocytosis to Chronic Inflammation
3.1. Accumulation of Apoptotic Cells
3.2. Secondary Necrosis
3.3. Activation of Inflammatory Pathways
4. Chronic Inflammation and Its Association with Diseases
- Enhancing Efferocytosis: Promoting efficient efferocytosis can help reduce inflammation and remove immunosuppressive apoptotic cells from the tumor microenvironment. Strategies to enhance efferocytosis include targeting efferocytosis receptors on phagocytes, modulating signaling pathways involved in efferocytosis, and promoting the resolution of inflammation [129].
- Immune Checkpoint Inhibition: Blocking immune checkpoint molecules, such as PD-1/PD-L1 and CTLA-4, can restore anti-tumor immune responses and enhance T cell-mediated tumor killing. Immune checkpoint inhibitors (ICIs) have shown efficacy in a variety of cancers by unleashing the immune system to recognize and attack tumor cells [131].
- Targeting Tumor-Associated Inflammation: Modulating the inflammatory microenvironment of tumors can enhance anti-tumor immunity and inhibit tumor growth. This can be achieved through targeting inflammatory mediators, such as cytokines and chemokines, or by repolarizing TAMs and MDSCs from immunosuppressive to anti-tumor phenotypes [132].
- Immunotherapy: Various immunotherapeutic approaches, including cancer vaccines, adoptive cell therapy (e.g., CAR-T cells), and cytokine therapy, aim to boost anti-tumor immune responses and overcome immune evasion mechanisms employed by tumors. These strategies harness the power of the immune system to recognize and eliminate tumor cells [133].Tumor immune escape is facilitated by chronic inflammation and impaired efferocytosis, which create an immunosuppressive microenvironment that is conducive to tumor growth [126]. Strategies aimed at restoring immunological responses, enhancing efferocytosis, and targeting tumor-associated inflammation can help to counteract tumor immune escape and improve the efficacy of cancer immunotherapy [134].
- Severe Infectious Diseases: Compromised immune responses, particularly linked to impaired efferocytosis and chronic inflammation, heighten a patient’s susceptibility to severe infections [135]. Apoptotic cell debris in tissues fosters a pathogen-friendly environment when efferocytosis is hindered, leading to the accumulation of dead cells and potential pathogen reservoirs [136]. Chronic inflammation associated with impaired efferocytosis amplifies pro-inflammatory cytokine and chemokine production, weakening overall immune defenses and promoting pathogen survival [135]. Prolonged exposure to inflammation and apoptotic cell debris impairs immune cell function, hindering pathogen recognition and elimination [137]. Continuous exposure to apoptotic cell debris may induce immune tolerance, compromising the immune system’s ability to mount effective defenses against infections [138]. Some pathogens exploit this compromised immune response, utilizing apoptotic cell debris as a protective niche to evade immune surveillance, replicate, and cause secondary infections [139]. This scenario is prominent in conditions featuring chronic inflammation or impaired efferocytosis, such as autoimmune diseases or chronic inflammatory disorders, further perpetuating the cycle [75]. The failure to efficiently clear apoptotic cells contributes to prolonged and dysregulated inflammation, leading to tissue damage, disease progression, and associated symptoms [14]. Enhancing apoptotic cell clearance could offer novel therapeutic approaches for managing these conditions.
5. Therapeutic Implications
- Enhancement of Efferocytosis: Developing pharmacological agents that are capable of boosting efferocytosis represents a promising therapeutic avenue [141]. This strategy may entail medications designed to enhance the recognition and clearance of apoptotic cells by phagocytes. Exploring the targeted modulation of signaling pathways involved in efferocytosis offers potential for improving its efficiency, including interventions that regulate the interactions between phagocytes and apoptotic cells [142].
- Anti-Inflammatory Therapies/ Cytokine Modulation: Modulating the levels of pro-inflammatory cytokines released in response to impaired efferocytosis may be considered [143]. Anti-inflammatory therapies targeting cytokines such as interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α) could help dampen chronic inflammation [142,144]. Developing therapeutic strategies that promote the production of resolution-inducing mediators, which help to resolve inflammation, could be beneficial [145].
- Immunomodulation: Modulating the activity of immune cells involved in the inflammatory response, such as macrophages and T cells, may help restore the balance and prevent excessive inflammation [146]. In autoimmune diseases associated with impaired efferocytosis, therapies aimed at modulating the immune response and preventing the recognition of self-antigens may be explored [147].
- Lipid Mediators and DAMP Clearance: Specialized Pro-Resolving Lipid Mediators (SPMs) such as resolvins and lipoxins are lipid mediators that actively promote the resolution of inflammation [148]. Developing strategies to enhance the production or administration of SPMs could be a therapeutic approach. Addressing the clearance of damage-associated molecular patterns (DAMPs) released during impaired efferocytosis may be crucial. Strategies that can enhance the removal of these molecules could help to mitigate the inflammatory response [149].
- Personalized Medicine/Patient-Specific Approaches: Considering the heterogeneity of inflammatory disorders, personalized medicine approaches that consider individual variations in efferocytosis and immune responses may be explored [150]. Identifying biomarkers associated with impaired efferocytosis and heightened inflammatory responses in individual patients can help tailor treatment strategies [151]. Biomarkers may include circulating levels of apoptotic cells, phagocyte function markers, cytokine profiles, and genetic variants associated with efferocytosis and inflammation [152].
- Identifying Drug Targets: GWAS can pinpoint genetic variants linked to disease susceptibility or treatment response, informing the identification of potential drug targets for more targeted therapies [155].
- Predicting Drug Response: Genetic variations identified through GWAS can affect drug metabolism, pharmacodynamics, and adverse reactions, enabling clinicians to predict individual patients’ responses to drugs and tailor treatment regimens accordingly [156].
- Personalizing Drug Therapy: Pharmacogenomics utilizes GWAS findings to personalize drug therapy based on patients’ genetic profiles, minimizing adverse effects, and enhancing treatment outcomes by selecting the most suitable drugs and dosages [157].
- Optimizing Drug Dosing: GWAS uncovers genetic variants influencing drug pharmacokinetics and pharmacodynamics, informing optimal dosing strategies tailored to different patient populations to achieve therapeutic levels effectively [158].
- Drug Development and Precision Medicine: GWAS results guide drug development by highlighting genetic targets and patient subpopulations that are likely to benefit from novel therapies, aligning with the principles of precision medicine to deliver tailored healthcare interventions [159].
6. Mitigation
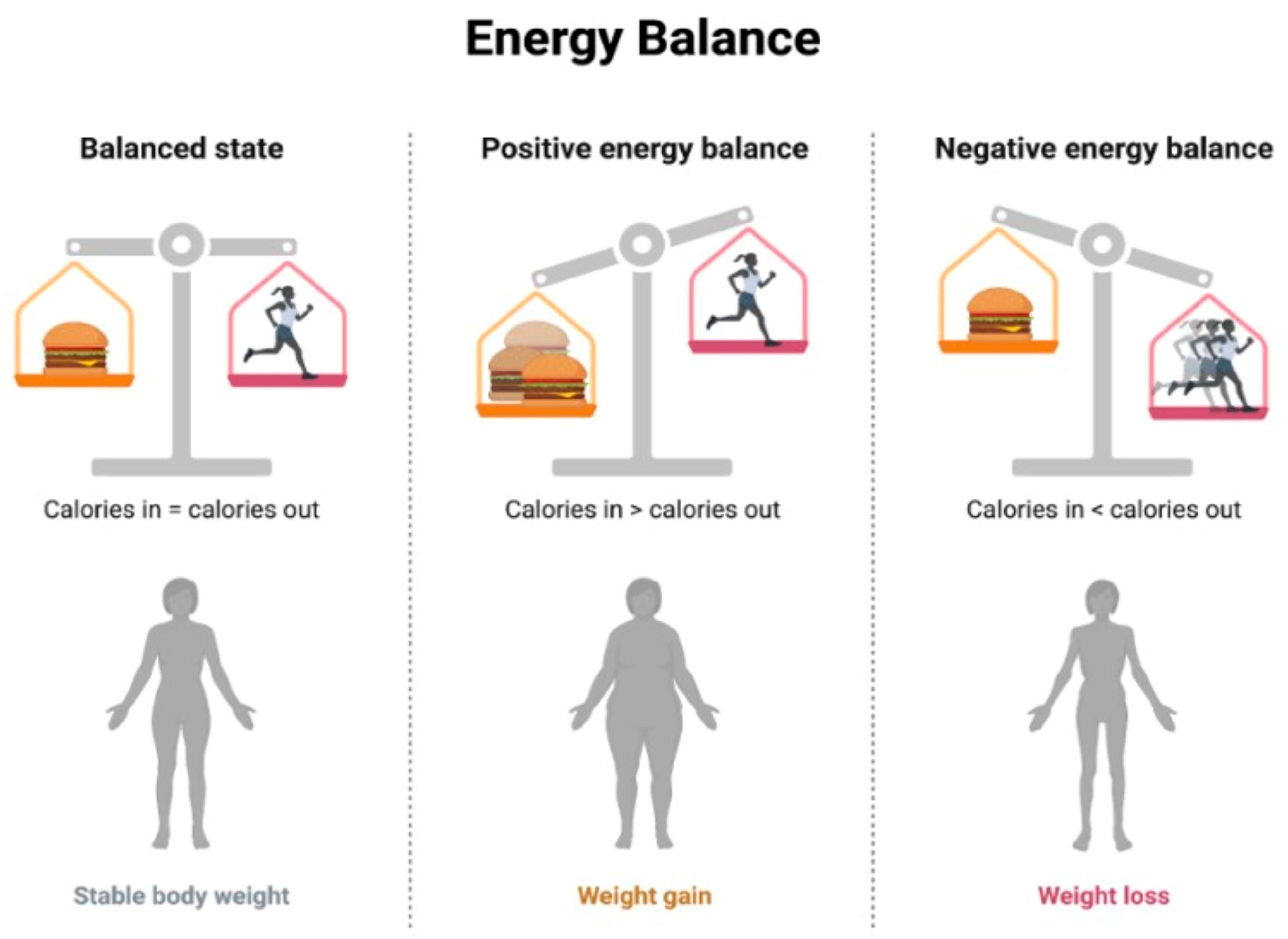
- Nutritional Counseling and Education: This involves working with healthcare professionals, including registered dietitians, to develop personalized dietary plans based on individual needs, preferences, and health conditions, implementing controlled caloric intake to achieve and maintain a healthy weight [167], and emphasizing a balanced distribution of macronutrients (carbohydrates, proteins, and fats) to meet nutritional needs without an excessive caloric intake [168]. The concept of energy balance refers to the relationship between energy intake (from food and beverages) and energy expenditure (through physical activity, metabolic processes, and other bodily functions) [169]. When the energy intake and energy expenditure are equal, there is said to be an energy balance. When the energy intake is greater than the energy expenditure, a positive energy balance occurs, resulting in weight gain over time. Conversely, when the energy expenditure is greater than the energy intake, a negative energy balance occurs, resulting in weight loss over time [170] (Figure 4).
- Regular Physical Activity: This involves incorporating regular physical activity tailored to individual fitness levels and health conditions and engaging in aerobic exercises to improve cardiovascular health and enhance metabolic function, including the use of resistance training to build muscle mass, which can contribute to improved insulin sensitivity [171].
- Weight Management In cases of severe obesity or metabolic disorders, medical supervision may be required, and interventions like bariatric surgery could be contemplated [172].
- Lipid-Lowering Medications: This involves administering statin medications to decrease elevated cholesterol levels and mitigate the risk of cardiovascular events, as well as using fibrates to target triglyceride levels and improve lipid profiles and incorporating omega-3 fatty acid supplements, which may have beneficial effects on lipid metabolism [173].
- Insulin Sensitizers: This involves administering metformin, an insulin-sensitizing medication commonly used in the management of type 2 diabetes. In certain cases, thiazolidinediones may be considered to improve insulin sensitivity [174].
- Anti-inflammatory Agents: This involves implementing anti-inflammatory lifestyle choices, including a diet rich in anti-inflammatory foods (e.g., fruits, vegetables, and fatty fish) and regular exercise. In some cases, medications with anti-inflammatory properties may be considered to address inflammation associated with lipotoxicity [175].
- Monitoring and Screening: Periodic health check-ups can be carried out to monitor metabolic parameters, lipid profiles, and other indicators of metabolic health. Screening for comorbidities can also help in identifying and managing comorbid conditions such as diabetes, hypertension, and cardiovascular diseases [176].
- Psychosocial Support: A comprehensive approach to combatting overnutrition includes behavioral counseling, lifestyle coaching, and public health initiatives [177]. Behavioral counseling provides crucial support and addresses emotional or psychological factors contributing to overnutrition. Lifestyle coaching guides individuals in adopting sustainable changes to their dietary and activity habits for long-term well-being [178].
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations
| DAMP | Damage-associated molecular patterns |
| GWAS | Genome-wide association studies |
| IBD | Inflammatory bowel disease |
| IL | Interleukin |
| HCC | Hepatocellular carcinoma |
| MHO | Metabolically Healthy Obesity |
| MUO | Metabolically Unhealthy Obesity |
| NAFLD | Non-Alcoholic fatty liver |
| NASH | Non-Alcoholic steatohepatitis |
| PAMP | Pathogen associated molecular patterns |
| RA | Rheumatoid arthritis |
| ROS | Reactive oxygen species |
| SLE | Systemic lupus erythematosus |
| SPM | Specialized pro-resolving lipid mediators |
| TNFα | Tumor necrosis factor-α |
References
- Cavaliere, G.; Cimmino, F.; Trinchese, G.; Catapano, A.; Petrella, L.; D’Angelo, M.; Lucchin, L.; Mollica, M.P. From Obesity-Induced Low-Grade Inflammation to Lipotoxicity and Mitochondrial Dysfunction: Altered Multi-Crosstalk between Adipose Tissue and Metabolically Active Organs. Antioxidants 2023, 12, 1172. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125 (Suppl. S2), S3–S23. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.A.; Punt, J.; Stranford, S.A. Chapter 1: Overview of the Immune System. In Kuby Immunology, 7th ed.; W.H. Freeman: New York, NY, USA, 2012; p. 2. ISBN 13-978-1429202114. [Google Scholar]
- de Nava, A.S.L.; Raja, A. Physiology, Metabolism. In StatPearls [Internet]; [Updated 12 September 2022]; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Lipke, K.; Kubis-Kubiak, A.; Piwowar, A. Molecular Mechanism of Lipotoxicity as an Interesting Aspect in the Development of Pathological States-Current View of Knowledge. Cells 2022, 11, 844. [Google Scholar] [CrossRef] [PubMed]
- Oyesola, O.O.; Souza, C.O.S.; Loke, P. The Influence of Genetic and Environmental Factors and Their Interactions on Immune Response to Helminth Infections. Front. Immunol. 2022, 13, 869163. [Google Scholar] [CrossRef] [PubMed]
- Unger, R.H. Minireview: Weapons of lean body mass destruction: The role of ectopic lipids in the metabolic syndrome. Endocrinology 2003, 144, 5159–5165. [Google Scholar] [CrossRef]
- Romieu, I.; Dossus, L.; Barquera, S.; Blottière, H.M.; Franks, P.W.; Gunter, M.; Hwalla, N.; Hursting, S.D.; Leitzmann, M.; Margetts, B.; et al. Energy balance and obesity: What are the main drivers? Cancer Causes Control CCC 2017, 28, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Popkin, B.M.; Ng, S.W. The nutrition transition to a stage of high obesity and noncommunicable disease prevalence dominated by ultra-processed foods is not inevitable. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2022, 23, e13366. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, J.E. Lipotoxicity: When tissues overeat. Curr. Opin. Lipidol. 2003, 14, 281–287. [Google Scholar] [CrossRef]
- Monga, S.P.; Behari, J. Chapter 20—Molecular Basis of Liver Disease. In Molecular Pathology, 2nd ed.; Coleman, W.B., Tsongalis, G.J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 417–456. ISBN 9780128027615. [Google Scholar]
- Grundy, S.M. Adipose tissue and metabolic syndrome: Too much, too little or neither. Eur. J. Clin. Investig. 2015, 45, 1209–1217. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Effect of High-Fat Diets on Oxidative Stress, Cellular Inflammatory Response and Cognitive Function. Nutrients 2019, 11, 2579. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.; Mohiuddin, S.S. Biochemistry, Lipolysis. In StatPearls [Internet]; [Updated 17 July 2023]; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK560564 (accessed on 15 January 2024).
- Estadella, D.; da Penha Oller do Nascimento, C.M.; Oyama, L.M.; Ribeiro, E.B.; Dâmaso, A.R.; de Piano, A. Lipotoxicity: Effects of dietary saturated and transfatty acids. Mediat. Inflamm. 2013, 2013, 137579. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Poti, J.M.; Braga, B.; Qin, B. Ultra-processed Food Intake and Obesity: What Really Matters for Health-Processing or Nutrient Content? Curr. Obes. Rep. 2017, 6, 420–431. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, A.; Jacka, F.N.; Quirk, S.E.; Cocker, F.; Taylor, C.B.; Oldenburg, B.; Berk, M. A shared framework for the common mental disorders and Non-Communicable Disease: Key considerations for disease prevention and control. BMC Psychiatry 2015, 15, 15. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.M.; Gamage, E.; Du, S.; Ashtree, D.N.; McGuinness, A.J.; Gauci, S.; Baker, P.; Lawrence, M.; Rebholz, C.M.; Srour, B.; et al. Ultra-processed food exposure and adverse health outcomes: Umbrella review of epidemiological meta-analyses. BMJ 2024, 384, e077310. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.D.; Ayuketah, A.; Brychta, R.; Cai, H.; Cassimatis, T.; Chen, K.Y.; Chung, S.T.; Costa, E.; Courville, A.; Darcey, V.; et al. Ultra-Processed Diets Cause Excess Calorie Intake and Weight Gain: An Inpatient Randomized Controlled Trial of Ad Libitum Food Intake. Cell Metab. 2019, 30, 67–77.e3. [Google Scholar] [CrossRef] [PubMed]
- Stanhope, K.L. Sugar consumption, metabolic disease and obesity: The state of the controversy. Crit. Rev. Clin. Lab. Sci. 2016, 53, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Juul, F.; Vaidean, G.; Parekh, N. Ultra-processed Foods and Cardiovascular Diseases: Potential Mechanisms of Action. Adv. Nutr. 2021, 12, 1673–1680. [Google Scholar] [CrossRef]
- Elizabeth, L.; Machado, P.; Zinöcker, M.; Baker, P.; Lawrence, M. Ultra-Processed Foods and Health Outcomes: A Narrative Review. Nutrients 2020, 12, 1955. [Google Scholar] [CrossRef]
- Calcaterra, V.; Cena, H.; Rossi, V.; Santero, S.; Bianchi, A.; Zuccotti, G. Ultra-Processed Food, Reward System and Childhood Obesity. Children 2023, 10, 804. [Google Scholar] [CrossRef] [PubMed]
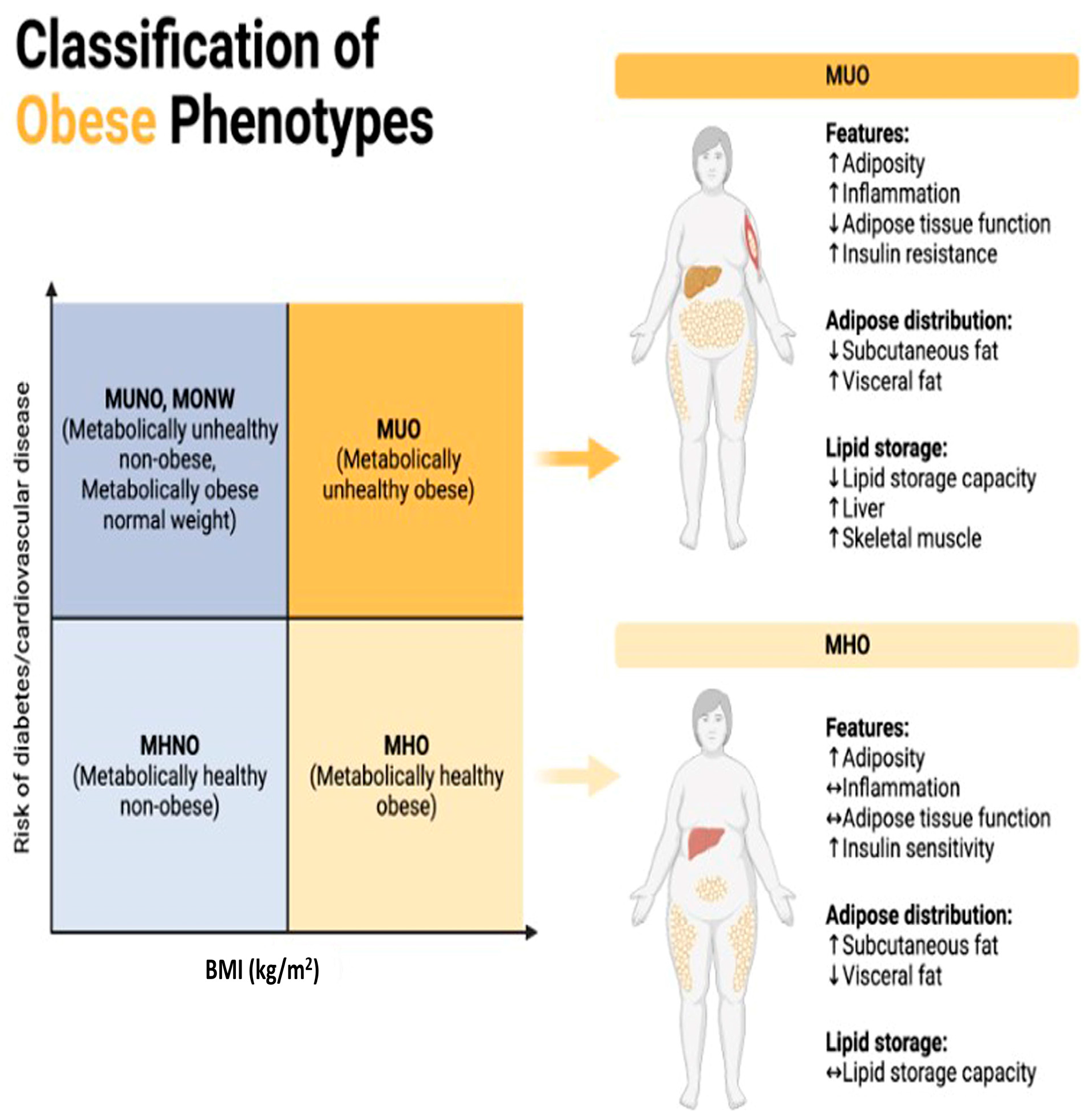
- Pluta, W.; Dudzińska, W.; Lubkowska, A. Metabolic Obesity in People with Normal Body Weight (MONW)-Review of Diagnostic Criteria. Int. J. Environ. Res. Public Health 2022, 19, 624. [Google Scholar] [CrossRef]
- Blaton, V. How is the Metabolic Syndrome Related to the Dyslipidemia? EJIFCC 2007, 18, 15–22. [Google Scholar] [CrossRef]
- Mathew, H.; Farr, O.M.; Mantzoros, C.S. Metabolic health and weight: Understanding metabolically unhealthy normal weight or metabolically healthy obese patients. Metab. Clin. Exp. 2016, 65, 73–80. [Google Scholar] [CrossRef]
- Yang, H.K.; Han, K.; Kwon, H.S.; Park, Y.M.; Cho, J.H.; Yoon, K.H.; Kang, M.I.; Cha, B.Y.; Lee, S.H. Obesity, metabolic health, and mortality in adults: A nationwide population-based study in Korea. Sci. Rep. 2016, 6, 30329. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.M.; Dillon, C.; Harrington, J.M.; McCarthy, V.J.; Kearney, P.M.; Fitzgerald, A.P.; Perry, I.J. Defining metabolically healthy obesity: Role of dietary and lifestyle factors. PLoS ONE 2013, 8, e76188. [Google Scholar] [CrossRef]
- Corrales, P.; Vidal-Puig, A.; Medina-Gómez, G. Obesity and pregnancy, the perfect metabolic storm. Eur. J. Clin. Nutr. 2021, 75, 1723–1734. [Google Scholar] [CrossRef] [PubMed]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution 2021, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adipose tissue in control of metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11, 1290. [Google Scholar] [CrossRef]
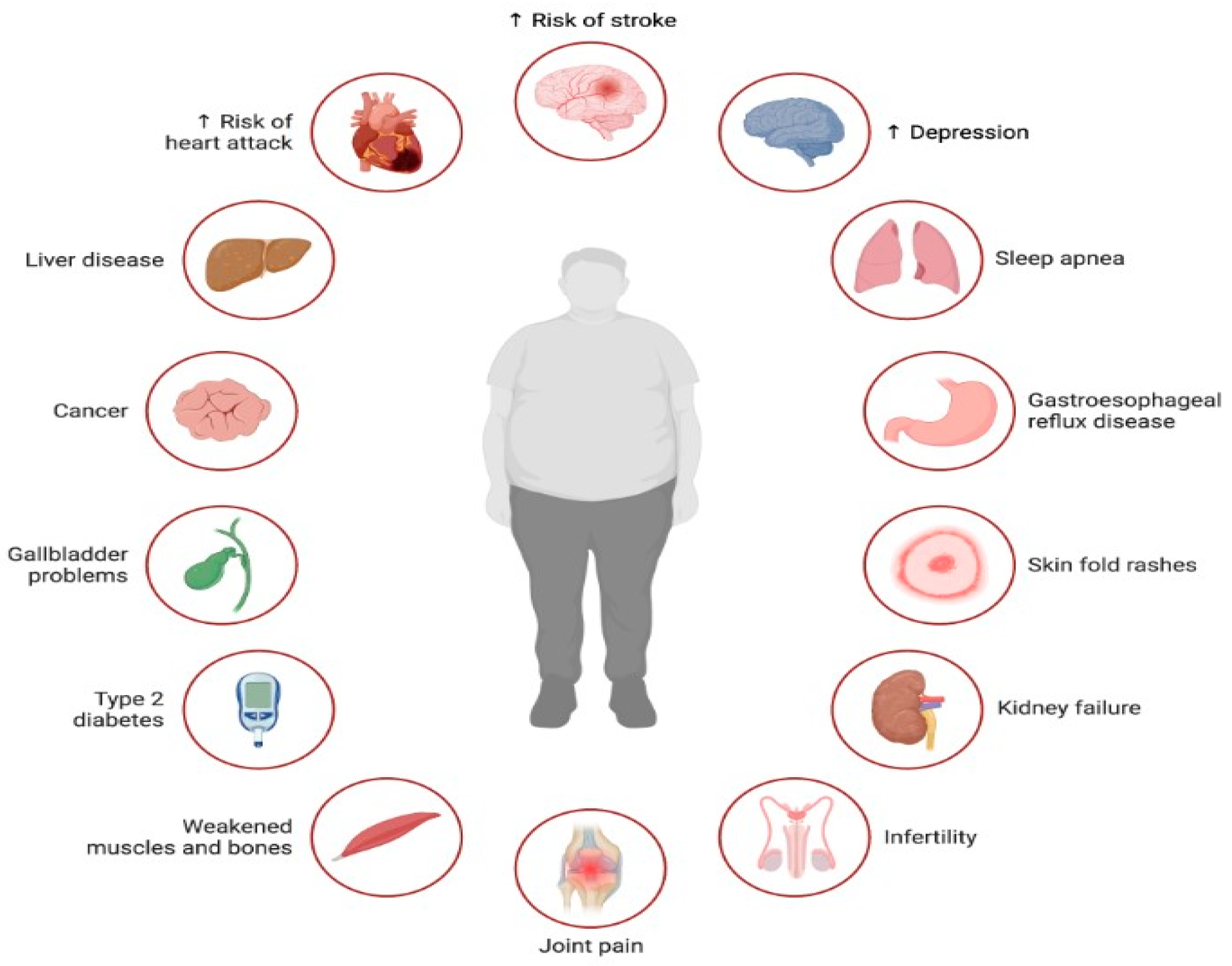
- Djalalinia, S.; Qorbani, M.; Peykari, N.; Kelishadi, R. Health impacts of Obesity. Pak. J. Med. Sci. 2015, 31, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, F.Q. Body mass index: Obesity, BMI, and health: A critical review. Nutr. Today 2015, 50, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Wondmkun, Y.T. Obesity, Insulin Resistance, and Type 2 Diabetes: Associations and Therapeutic Implications. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 3611–3616. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Al-Qahtani, A.A.; Alhamlan, F.S.; Al-Qahtani, A.A. Pro-Inflammatory and Anti-Inflammatory Interleukins in Infectious Diseases: A Comprehensive Review. Trop. Med. Infect. Dis. 2024, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Enayati, S.; Seifirad, S.; Amiri, P.; Abolhalaj, M.; Mohammad-Amoli, M. Interleukin-1 beta, interferon-gamma, and tumor necrosis factor-alpha gene expression in peripheral blood mononuclear cells of patients with coronary artery disease. ARYA Atheroscler. 2015, 11, 267–274. [Google Scholar]
- Steen, E.H.; Wang, X.; Balaji, S.; Butte, M.J.; Bollyky, P.L.; Keswani, S.G. The Role of the Anti-Inflammatory Cytokine Interleukin-10 in Tissue Fibrosis. Adv. Wound Care 2020, 9, 184–198. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Pu, F.; Chen, F.; Zhang, Z.; Shi, D.; Zhong, B.; Lv, X.; Tucker, A.B.; Fan, J.; Li, A.J.; Qin, K.; et al. Ferroptosis as a novel form of regulated cell death: Implications in the pathogenesis, oncometabolism and treatment of human cancer. Genes Dis. 2020, 9, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Jia, J.; Zheng, J.; Zhou, Y.; Jia, D.; Wang, J. Recent Progress of Ferroptosis in Lung Diseases. Front. Cell Dev. Biol. 2021, 9, 789517. [Google Scholar] [CrossRef]
- Zhou, Y.; Lin, W.; Rao, T.; Zheng, J.; Zhang, T.; Zhang, M.; Lin, Z. Ferroptosis and Its Potential Role in the Nervous System Diseases. J. Inflamm. Res. 2022, 15, 1555–1574. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhou, X.; Xie, F.; Zhang, L.; Yan, H.; Huang, J.; Zhang, C.; Zhou, F.; Chen, J.; Zhang, L. Ferroptosis in cancer and cancer immunotherapy. Cancer Commun. 2022, 42, 88–116. [Google Scholar] [CrossRef] [PubMed]
- Jurcau, A.; Ardelean, A.I. Oxidative Stress in Ischemia/Reperfusion Injuries following Acute Ischemic Stroke. Biomedicines 2022, 10, 574. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Sadoshima, J. Molecular mechanisms of mitochondrial autophagy/mitophagy in the heart. Circ. Res. 2015, 116, 1477–1490. [Google Scholar] [CrossRef]
- Li, A.; Gao, M.; Liu, B.; Qin, Y.; Chen, L.; Liu, H.; Wu, H.; Gong, G. Mitochondrial autophagy: Molecular mechanisms and implications for cardiovascular disease. Cell Death Dis. 2022, 13, 444. [Google Scholar] [CrossRef] [PubMed]
- Jaishy, B.; Abel, E.D. Lipids, lysosomes, and autophagy. J. Lipid Res. 2016, 57, 1619–1635. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, T.; Wu, J.M.F.; Schulze, P.C. Mitochondrial Homeostasis Mediates Lipotoxicity in the Failing Myocardium. Int. J. Mol. Sci. 2021, 22, 1498. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Makkar, R.; Anwer, M.K.; Hassani, R.; Khuwaja, G.; Khalid, A.; Mohan, S.; Alhazmi, H.A.; Sachdeva, M.; Rachamalla, M. Mitochondrial Dysfunction: A Cellular and Molecular Hub in Pathology of Metabolic Diseases and Infection. J. Clin. Med. 2023, 12, 2882. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, J.; Kang, R.; Tang, D. Interplay Between Lipid Metabolism and Autophagy. Front. Cell Dev. Biol. 2020, 8, 431. [Google Scholar] [CrossRef] [PubMed]
- Namkoong, S.; Cho, C.S.; Semple, I.; Lee, J.H. Autophagy Dysregulation and Obesity-Associated Pathologies. Mol. Cells 2018, 41, 3–10. [Google Scholar] [CrossRef]
- Cornier, M.A.; Dabelea, D.; Hernandez, T.L.; Lindstrom, R.C.; Steig, A.J.; Stob, N.R.; Van Pelt, R.E.; Wang, H.; Eckel, R.H. The metabolic syndrome. Endocr. Rev. 2008, 29, 777–822. [Google Scholar] [CrossRef] [PubMed]
- National Heart, Lung and Blood Institute. Overweight and Obesity. Available online: http://www.nhlbi.nih.gov/health/health-topics/topics/obe/risks (accessed on 20 March 2024).
- Hassamal, S. Chronic stress, neuroinflammation, and depression: An overview of pathophysiological mechanisms and emerging anti-inflammatories. Front. Psychiatry 2023, 14, 1130989. [Google Scholar] [CrossRef] [PubMed]
- Jehan, S.; Zizi, F.; Pandi-Perumal, S.R.; Wall, S.; Auguste, E.; Myers, A.K.; Jean-Louis, G.; McFarlane, S.I. Obstructive Sleep Apnea and Obesity: Implications for Public Health. Sleep Med. Disord. Int. J. 2017, 1, 00019. [Google Scholar]
- Bendarska-Czerwińska, A.; Zmarzły, N.; Morawiec, E.; Panfil, A.; Bryś, K.; Czarniecka, J.; Ostenda, A.; Dziobek, K.; Sagan, D.; Boroń, D.; et al. Endocrine disorders and fertility and pregnancy: An update. Front. Endocrinol. 2023, 13, 970439. [Google Scholar] [CrossRef]
- Elliott, M.R.; Koster, K.M.; Murphy, P.S. Efferocytosis Signaling in the Regulation of Macrophage Inflammatory Responses. J. Immunol. 2017, 198, 1387–1394. [Google Scholar] [CrossRef]
- Xu, S.; Xi, J.; Wu, T.; Wang, Z. The Role of Adipocyte Endoplasmic Reticulum Stress in Obese Adipose Tissue Dysfunction: A Review. Int. J. Gen. Med. 2023, 16, 4405–4418. [Google Scholar] [CrossRef]
- Pouwels, S.; Sakran, N.; Graham, Y.; Leal, A.; Pintar, T.; Yang, W.; Kassir, R.; Singhal, R.; Mahawar, K.; Ramnarain, D. Non-alcoholic fatty liver disease (NAFLD): A review of pathophysiology, clinical management and effects of weight loss. BMC Endocr. Disord. 2022, 22, 63. [Google Scholar] [CrossRef] [PubMed]
- Daher, D.; Dahan, K.S.E.; Singal, A.G. Non-alcoholic fatty liver disease-related hepatocellular carcinoma. J. Liver Cancer 2023, 23, 127–142. [Google Scholar] [CrossRef]
- Juanola, O.; Martínez-López, S.; Francés, R.; Gómez-Hurtado, I. Non-Alcoholic Fatty Liver Disease: Metabolic, Genetic, Epigenetic and Environmental Risk Factors. Int. J. Environ. Res. Public Health 2021, 18, 5227. [Google Scholar] [CrossRef]
- Ahmed, I.A.; Mikail, M.A.; Mustafa, M.R.; Ibrahim, M.; Othman, R. Lifestyle interventions for non-alcoholic fatty liver disease. Saudi J. Biol. Sci. 2019, 26, 1519–1524. [Google Scholar] [CrossRef]
- Elliott, M.R.; Ravichandran, K.S. Clearance of apoptotic cells: Implications in health and disease. J. Cell Biol. 2010, 189, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S.; Hanayama, R.; Kawane, K. Autoimmunity and the clearance of dead cells. Cell 2010, 140, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Gardai, S.J.; Bratton, D.L.; Ogden, C.A.; Henson, P.M. Recognition ligands on apoptotic cells: A perspective. J. Leukoc. Biol. 2006, 79, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Hart, S.P.; Dransfield, I.; Rossi, A.G. Phagocytosis of apoptotic cells. Methods 2008, 44, 280–285. [Google Scholar] [CrossRef]
- Lemke, G.; Burstyn-Cohen, T. TAM receptors and the clearance of apoptotic cells. Ann. N. Y. Acad. Sci. 2010, 1209, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Henson, P.M.; Hume, D.A. Apoptotic cell removal in development and tissue homeostasis. Trends Immunol. 2006, 27, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Penberthy, K.K.; Ravichandran, K.S. Apoptotic cell recognition receptors and scavenger receptors. Immunol. Rev. 2016, 269, 44–59. [Google Scholar] [CrossRef] [PubMed]
- Erwig, L.P.; Henson, P.M. Immunological consequences of apoptotic cell phagocytosis. Am. J. Pathol. 2007, 171, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, S.; Grinstein, S. Phagocytosis and innate immunity. Curr. Opin. Immunol. 2002, 14, 136–145. [Google Scholar] [CrossRef]
- Ge, Y.; Huang, M.; Yao, Y.M. Efferocytosis and Its Role in Inflammatory Disorders. Front. Cell Dev. Biol. 2022, 10, 839248. [Google Scholar] [CrossRef]
- Parameswaran, N.; Patial, S. Tumor necrosis factor-α signaling in macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.D.; Devitt, A. The macrophage and the apoptotic cell: An innate immune interaction viewed simplistically? Immunology 2004, 113, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lazarov, T.; Juarez-Carreño, S.; Cox, N.; Geissmann, F. Physiology and diseases of tissue-resident macrophages. Nature 2023, 618, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.A.; Devitt, A.; Johnson, J.R. Macrophages: The Good, the Bad, and the Gluttony. Front. Immunol. 2021, 12, 708186. [Google Scholar] [CrossRef]
- Gui, Y.; Zheng, H.; Cao, R.Y. Foam Cells in Atherosclerosis: Novel Insights into Its Origins, Consequences, and Molecular Mechanisms. Front. Cardiovasc. Med. 2022, 9, 845942. [Google Scholar] [CrossRef]
- Basak, U.; Sarkar, T.; Mukherjee, S.; Chakraborty, S.; Dutta, A.; Dutta, S.; Nayak, D.; Kaushik, S.; Das, T.; Sa, G. Tumor-associated macrophages: An effective player of the tumor microenvironment. Front. Immunol. 2023, 14, 1295257. [Google Scholar] [CrossRef]
- Liu, T.; Liu, F.; Peng, L.W.; Chang, L.; Jiang, Y.M. The Peritoneal Macrophages in Inflammatory Diseases and Abdominal Cancers. Oncol. Res. 2018, 26, 817–826. [Google Scholar] [CrossRef]
- Finicelli, M.; Digilio, F.A.; Galderisi, U.; Peluso, G. The Emerging Role of Macrophages in Chronic Obstructive Pulmonary Disease: The Potential Impact of Oxidative Stress and Extracellular Vesicle on Macrophage Polarization and Function. Antioxidants 2022, 11, 464. [Google Scholar] [CrossRef]
- Loh, W.; Vermeren, S. Anti-Inflammatory Neutrophil Functions in the Resolution of Inflammation and Tissue Repair. Cells 2022, 11, 4076. [Google Scholar] [CrossRef]
- Ilias, G.; Konstantinos, P.; Nektarios, T. The Role of Mitophagy in Innate Immunity. Front. Immunol. 2018, 9, 1664–3224. [Google Scholar]
- Marchi, S.; Guilbaud, E.; Tait, S.W.; Yamazaki, T.; Galluzzi, L. Mitochondrial control of inflammation. Nat. Rev. Immunol. 2023, 23, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Chen, G.; Chen, Q. Crosstalk between mitochondrial biogenesis and mitophagy to maintain mitochondrial homeostasis. J. Biomed. Sci. 2023, 30, 86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Y.; Ding, J.; Liu, P. Efferocytosis in multisystem diseases (Review). Mol. Med. Rep. 2022, 25, 13. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Cheng, X.; Li, F.; Guan, Z.; Xu, J.; Wu, D.; Gao, Y.; Zhan, X.; Wang, P.; Zhou, H.; et al. Defective efferocytosis by aged macrophages promotes STING signaling mediated inflammatory liver injury. Cell Death Discov. 2023, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Abdal Dayem, A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.M.; Choi, H.Y.; Cho, S.G. The Role of Reactive Oxygen Species (ROS) in the Biological Activities of Metallic Nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef]
- Milkovic, L.; Cipak Gasparovic, A.; Cindric, M.; Mouthuy, P.A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef]
- Chae, S.Y.; Kim, Y.; Park, C.W. Oxidative Stress Induced by Lipotoxicity and Renal Hypoxia in Diabetic Kidney Disease and Possible Therapeutic Interventions: Targeting the Lipid Metabolism and Hypoxia. Antioxidants 2023, 12, 2083. [Google Scholar] [CrossRef]
- Römer, A.; Linn, T.; Petry, S.F. Lipotoxic Impairment of Mitochondrial Function in β-Cells: A Review. Antioxidants 2021, 10, 293. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Afzal, S.; Abdul Manap, A.S.; Attiq, A.; Albokhadaim, I.; Kandeel, M.; Alhojaily, S.M. From imbalance to impairment: The central role of reactive oxygen species in oxidative stress-induced disorders and therapeutic exploration. Front. Pharmacol. 2023, 14, 1269581. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, M.S.; Ruiz, J.; Watts, J.L. Polyunsaturated Fatty Acids Drive Lipid Peroxidation during Ferroptosis. Cells 2023, 12, 804. [Google Scholar] [CrossRef] [PubMed]
- Checa, J.; Aran, J.M. Reactive Oxygen Species: Drivers of Physiological and Pathological Processes. J. Inflamm. Res. 2020, 13, 1057–1073. [Google Scholar] [CrossRef] [PubMed]
- Ertunc, M.E.; Hotamisligil, G.S. Lipid signaling and lipotoxicity in metaflammation: Indications for metabolic disease pathogenesis and treatment. J. Lipid Res. 2016, 57, 2099–2114. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Kaufman, R.J. The role of ER stress in lipid metabolism and lipotoxicity. J. Lipid Res. 2016, 57, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Hauck, A.K.; Bernlohr, D.A. Oxidative stress and lipotoxicity. J. Lipid Res. 2016, 57, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Monks, J.; Rosner, D.; Jon Geske, F.; Lehman, L.; Hanson, L.; Neville, M.C.; Fadok, V.A. Epithelial Cells as Phagocytes: Apoptotic Epithelial Cells Are Engulfed by Mammary Alveolar Epithelial Cells and Repress Inflammatory Mediator Release. Cell Death Differ. 2005, 12, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Moon, B.; Yang, S.; Moon, H.; Lee, J.; Park, D. After cell death: The molecular machinery of efferocytosis. Exp. Mol. Med. 2023, 55, 1644–1651. [Google Scholar] [CrossRef]
- Henein, M.Y.; Vancheri, S.; Longo, G.; Vancheri, F. The Role of Inflammation in Cardiovascular Disease. Int. J. Mol. Sci. 2022, 23, 12906. [Google Scholar] [CrossRef]
- Parolini, C. The Role of Marine n-3 Polyunsaturated Fatty Acids in Inflammatory-Based Disease: The Case of Rheumatoid Arthritis. Mar. Drugs 2023, 22, 17. [Google Scholar] [CrossRef] [PubMed]
- Amor, S.; Puentes, F.; Baker, D.; van der Valk, P. Inflammation in neurodegenerative diseases. Immunology 2010, 129, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Sachet, M.; Liang, Y.Y.; Oehler, R. The immune response to secondary necrotic cells. Apoptosis Int. J. Program. Cell Death 2017, 22, 1189–1204. [Google Scholar] [CrossRef] [PubMed]
- Arandjelovic, S.; Ravichandran, K.S. Phagocytosis of apoptotic cells in homeostasis. Nat. Immunol. 2015, 16, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhao, M.; Jia, S. Macrophage: Key player in the pathogenesis of autoimmune diseases. Front. Immunol. 2023, 14, 1080310. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, H.; Tang, Y.; Yao, P. Potential Mechanisms and Effects of Efferocytosis in Atherosclerosis. Front. Endocrinol. 2021, 11, 585285. [Google Scholar] [CrossRef] [PubMed]
- Akata, K.; van Eeden, S.F. Lung Macrophage Functional Properties in Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2020, 21, 853. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Matsuoka, S.; Yamazaki, M.; Shibata, T.; Nirei, K.; Takahashi, H.; Kaneko, T.; Fujisawa, M.; Higuchi, T.; Nakamura, H.; et al. Apoptosis and non-alcoholic fatty liver diseases. World J. Gastroenterol. 2018, 24, 2661–2672. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, W.; Wu, T.; Wang, H.; Mao, J.; Liu, J.; Zhou, Z.; Lin, X.; Yan, H.; Wang, Q. Efferocytosis in the Central Nervous System. Front. Cell Dev. Biol. 2021, 9, 773344. [Google Scholar] [CrossRef] [PubMed]
- Granger, D.N.; Senchenkova, E. Angiogenesis. In Inflammation and the Microcirculation; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010; Chapter 6. Available online: https://www.ncbi.nlm.nih.gov/books/NBK53373 (accessed on 15 January 2024).
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef]
- Yang, A.; Wu, Y.; Yu, G.; Wang, H. Role of specialized pro-resolving lipid mediators in pulmonary inflammation diseases: Mechanisms and development. Respir. Res. 2021, 22, 204. [Google Scholar] [CrossRef]
- Bennett, J.M.; Reeves, G.; Billman, G.E.; Sturmberg, J.P. Inflammation-Nature’s Way to Efficiently Respond to All Types of Challenges: Implications for Understanding and Managing “the Epidemic” of Chronic Diseases. Front. Med. 2018, 5, 316. [Google Scholar] [CrossRef] [PubMed]
- Pahwa, R.; Goyal, A.; Jialal, I. Chronic Inflammation. In StatPearls [Internet]; [Updated 7 August 2023]; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK493173 (accessed on 15 January 2024).
- Martin, C.J.; Peters, K.N.; Behar, S.M. Macrophages clean up: Efferocytosis and microbial control. Curr. Opin. Microbiol. 2014, 17, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kojima, Y.; Weissman, I.L.; Leeper, N.J. The Role of Efferocytosis in Atherosclerosis. Circulation 2017, 135, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Cuda, C.M.; Pope, R.M.; Perlman, H. The inflammatory role of phagocyte apoptotic pathways in rheumatic diseases. Nat. Rev. Rheumatol. 2016, 12, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Kim, S.K.; Cho, S.W. The Evasion Mechanisms of Cancer Immunity and Drug Intervention in the Tumor Microenvironment. Front. Pharmacol. 2022, 13, 868695. [Google Scholar] [CrossRef]
- Vaught, D.B.; Stanford, J.C.; Cook, R.S. Efferocytosis creates a tumor microenvironment supportive of tumor survival and metastasis. Cancer Cell Microenviron. 2015, 2, e666. [Google Scholar] [CrossRef]
- Multhoff, G.; Molls, M.; Radons, J. Chronic inflammation in cancer development. Front. Immunol. 2012, 2, 98. [Google Scholar] [CrossRef]
- Zhao, Y.; Du, J.; Shen, X. Targeting myeloid-derived suppressor cells in tumor immunotherapy: Current, future and beyond. Front. Immunol. 2023, 14, 1157537. [Google Scholar] [CrossRef] [PubMed]
- Werfel, T.A.; Cook, R.S. Efferocytosis in the tumor microenvironment. Semin. Immunopathol. 2018, 40, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Reiser, J.; Banerjee, A. Effector, Memory, and Dysfunctional CD8+ T Cell Fates in the Antitumor Immune Response. J. Immunol. Res. 2016, 2016, 8941260. [Google Scholar] [CrossRef] [PubMed]
- Shiravand, Y.; Khodadadi, F.; Kashani, S.M.A.; Hosseini-Fard, S.R.; Hosseini, S.; Sadeghirad, H.; Ladwa, R.; O’Byrne, K.; Kulasinghe, A. Immune Checkpoint Inhibitors in Cancer Therapy. Curr. Oncol. 2022, 29, 3044–3060. [Google Scholar] [CrossRef]
- Belgiovine, C.; Digifico, E.; Anfray, C.; Ummarino, A.; Torres Andón, F. Targeting Tumor-Associated Macrophages in Anti-Cancer Therapies: Convincing the Traitors to Do the Right Thing. J. Clin. Med. 2020, 9, 3226. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Che, X.; Wang, X.; Ma, C.; Wu, G. Tumor Vaccines: Unleashing the Power of the Immune System to Fight Cancer. Pharmaceuticals 2023, 16, 1384. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Cui, J. Development of Immunotherapy Strategies Targeting Tumor Microenvironment Is Fiercely Ongoing. Front. Immunol. 2022, 13, 890166. [Google Scholar] [CrossRef] [PubMed]
- Doran, A.C.; Yurdagul, A., Jr.; Tabas, I. Efferocytosis in health and disease. Nat. Rev. Immunol. 2020, 20, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Heit, B. Cellular Responses to the Efferocytosis of Apoptotic Cells. Front. Immunol. 2021, 12, 631714. [Google Scholar] [CrossRef]
- Westman, J.; Grinstein, S.; Marques, P.E. Phagocytosis of Necrotic Debris at Sites of Injury and Inflammation. Front. Immunol. 2020, 10, 3030. [Google Scholar] [CrossRef]
- Nedeva, C. Inflammation and Cell Death of the Innate and Adaptive Immune System during Sepsis. Biomolecules 2021, 11, 1011. [Google Scholar] [CrossRef]
- Mak, T.W.; Saunders, M.E. Immunity to Pathogens. Immune Response 2006, 641–694. [Google Scholar] [CrossRef]
- Mehrotra, P.; Ravichandran, K.S. Drugging the efferocytosis process: Concepts and opportunities. Nat. Rev. Drug Discov. 2022, 21, 601–620. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Tosello-Trampont, A.C.; Elliott, M.R.; Lu, M.; Haney, L.B.; Ma, Z.; Klibanov, A.L.; Mandell, J.W.; Ravichandran, K.S. BAI1 is an engulfment receptor for apoptotic cells upstream of the ELMO/Dock180/Rac module. Nature 2007, 450, 430–434. [Google Scholar] [CrossRef]
- Ipseiz, N.; Uderhardt, S.; Scholtysek, C.; Steffen, M.; Schabbauer, G.; Bozec, A.; Schett, G.; Krönke, G. The nuclear receptor Nr4a1 mediates anti-inflammatory effects of apoptotic cells. J. Immunol. 2014, 192, 4852–4858. [Google Scholar] [CrossRef] [PubMed]
- Korns, D.; Frasch, S.C.; Fernandez-Boyanapalli, R.; Henson, P.M.; Bratton, D.L. Modulation of macrophage efferocytosis in inflammation. Front. Immunol. 2011, 2, 57. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Sugimoto, M.A.; Sousa, L.P.; Pinho, V.; Perretti, M.; Teixeira, M.M. Resolution of Inflammation: What Controls Its Onset? Front. Immunol. 2016, 7, 160. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yao, Z.; Xue, L.; Wang, D.; Tan, Z. The role of immune cells in modulating chronic inflammation and osteonecrosis. Front. Immunol. 2022, 13, 1064245. [Google Scholar] [CrossRef]
- Abdolmaleki, F.; Farahani, N.; Gheibi Hayat, S.M.; Pirro, M.; Bianconi, V.; Barreto, G.E.; Sahebkar, A. The Role of Efferocytosis in Autoimmune Diseases. Front. Immunol. 2018, 9, 1645. [Google Scholar] [CrossRef]
- Kumar, V.; Yasmeen, N.; Chaudhary, A.A.; Alawam, A.S.; Al-Zharani, M.; Suliman Basher, N.; Harikrishnan, S.; Goud, M.D.; Pandey, A.; Lakhawat, S.S.; et al. Specialized pro-resolving lipid mediators regulate inflammatory macrophages: A paradigm shift from antibiotics to immunotherapy for mitigating COVID-19 pandemic. Front. Mol. Biosci. 2023, 10, 1104577. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Mukherjee, A.; Nongthomba, U. Before the “cytokine storm”: Boosting efferocytosis as an effective strategy against SARS-CoV-2 infection and associated complications. Cytokine Growth Factor Rev. 2022, 63, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Park, S.H. Personalized medicine in inflammatory bowel disease: Perspectives on Asia. J. Gastroenterol. Hepatol. 2022, 37, 1434–1445. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yao, Y.; Deng, Y.; Shao, A. Regulation of efferocytosis as a novel cancer therapy. Cell Commun. Signal. 2020, 18, 71. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Lei, Q.; Luo, X.; Yin, J.; Chen, S.; Hao, C.; Shiyu, L.; Ma, D. Advances in biological functions and applications of apoptotic vesicles. Cell Commun. Signal. 2023, 21, 260. [Google Scholar] [CrossRef]
- Chu, X.; Janssen, A.W.M.; Koenen, H.; Chang, L.; He, X.; Joosten, I.; Stienstra, R.; Kuijpers, Y.; Wijmenga, C.; Xu, C.J.; et al. A genome-wide functional genomics approach uncovers genetic determinants of immune phenotypes in type 1 diabetes. eLife 2022, 11, e73709. [Google Scholar] [CrossRef] [PubMed]
- Witte, J.S. Genome-wide association studies and beyond. Annu. Rev. Public Health 2010, 31, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Floris, M.; Olla, S.; Schlessinger, D.; Cucca, F. Genetic-Driven Druggable Target Identification and Validation. Trends Genet. TIG 2018, 34, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Madian, A.G.; Wheeler, H.E.; Jones, R.B.; Dolan, M.E. Relating human genetic variation to variation in drug responses. Trends Genet. TIG 2012, 28, 487–495. [Google Scholar] [CrossRef]
- Ni, X.; Zhang, W.; Huang, R.S. Pharmacogenomics discovery and implementation in genome-wide association studies era. Wiley Interdiscip. Rev. Syst. Biol. Med. 2013, 5, 1–9. [Google Scholar] [CrossRef]
- McInnes, G.; Yee, S.W.; Pershad, Y.; Altman, R.B. Genomewide Association Studies in Pharmacogenomics. Clin. Pharmacol. Ther. 2021, 110, 637–648. [Google Scholar] [CrossRef]
- Dugger, S.A.; Platt, A.; Goldstein, D.B. Drug development in the era of precision medicine. Nat. Rev. Drug Discov. 2018, 17, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Albert-Vega, C.; Tawfik, D.M.; Trouillet-Assant, S.; Vachot, L.; Mallet, F.; Textoris, J. Immune Functional Assays, From Custom to Standardized Tests for Precision Medicine. Front. Immunol. 2018, 9, 2367. [Google Scholar] [CrossRef] [PubMed]
- Bjornson, Z.B.; Nolan, G.P.; Fantl, W.J. Single-cell mass cytometry for analysis of immune system functional states. Curr. Opin. Immunol. 2013, 25, 484–494. [Google Scholar] [CrossRef]
- Lin, W.; Liu, S.; Huang, Z.; Li, H.; Lu, T.; Luo, Y.; Zhong, J.; Xu, Z.; Liu, Y.; Li, Y.; et al. Mass cytometry and single-cell RNA sequencing reveal immune cell characteristics of active and inactive phases of Crohn’s disease. Front. Med. 2023, 9, 1064106. [Google Scholar] [CrossRef] [PubMed]
- Soehnlein, O.; Libby, P. Targeting inflammation in atherosclerosis—From experimental insights to the clinic. Nat. Rev. Drug Discov. 2021, 20, 589–610. [Google Scholar] [CrossRef] [PubMed]
- Goetz, L.H.; Schork, N.J. Personalized medicine: Motivation, challenges, and progress. Fertil. Steril. 2018, 109, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Sugandh, F.; Chandio, M.; Raveena, F.; Kumar, L.; Karishma, F.; Khuwaja, S.; Memon, U.A.; Bai, K.; Kashif, M.; Varrassi, G.; et al. Advances in the Management of Diabetes Mellitus: A Focus on Personalized Medicine. Cureus 2023, 15, e43697. [Google Scholar] [CrossRef] [PubMed]
- Wadden, T.A.; Tronieri, J.S.; Butryn, M.L. Lifestyle modification approaches for the treatment of obesity in adults. Am. Psychol. 2020, 75, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (US) Committee on Dietary Guidelines Implementation; Thomas, P.R. (Eds.) Health-Care Professionals: Strategies and Actions for Implementation. In Improving America’s Diet and Health: From Recommendations to Action; National Academies Press (US): Washington, DC, USA, 1991; p. 7. Available online: https://www.ncbi.nlm.nih.gov/books/NBK235263/ (accessed on 3 January 2024).
- Cena, H.; Calder, P.C. Defining a Healthy Diet: Evidence for The Role of Contemporary Dietary Patterns in Health and Disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef]
- Hill, J.O.; Wyatt, H.R.; Peters, J.C. The Importance of Energy Balance. Eur. Endocrinol. 2013, 9, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.O.; Wyatt, H.R.; Peters, J.C. Energy balance and obesity. Circulation 2012, 126, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.E.; Wang, J.; Sevick, M.A. Self-monitoring in weight loss: A systematic review of the literature. J. Am. Diet Assoc. 2011, 111, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Fried, M.; Yumuk, V.; Oppert, J.M.; Scopinaro, N.; Torres, A.J.; Weiner, R.; Yashkov, Y.; Frühbeck, G.; European Association for the Study of Obesity; International Federation for the Surgery of Obesity—European Chapter. Interdisciplinary European Guidelines on metabolic and bariatric surgery. Obes. Facts 2013, 6, 449–468. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, T.A.; Zimmerman, F.H. Fibrates in combination with statins in the management of dyslipidemia. J. Clin. Hypertens. 2006, 8, 35–43. [Google Scholar] [CrossRef]
- Eggleton, J.S.; Jialal, I. Thiazolidinediones. In StatPearls [Internet]; [Updated 20 February 2023]; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Giugliano, D.; Ceriello, A.; Esposito, K. The effects of diet on inflammation: Emphasis on the metabolic syndrome. J. Am. Coll. Cardiol. 2006, 48, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Al-Awad, F.A.; Almutairi, H.A.; Almutairi, S.A.; Alessa, O.S.; Alanazi, S.F.; Alzain, N.M.; Albakr, D.M.; Alzahrani, S.I. Adherence to the monitoring of metabolic syndrome in patients receiving antipsychotics in outpatient clinics in Saudi Arabia. J. Fam. Community Med. 2024, 31, 42–47. [Google Scholar] [CrossRef]
- Tronieri, J.S.; Alamuddin, N.; Wadden, T.A. Behavioral Approaches to Obesity Management; [Updated 13 October 2022]; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK278952/ (accessed on 1 March 2024).
- Rippe, J.M. Lifestyle Medicine: The Health Promoting Power of Daily Habits and Practices. Am. J. Lifestyle Med. 2018, 12, 499–512. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Approach | Description | Personalized Aspect |
|---|---|---|
| Pharmacological Modulation | Development of pharmacological agents targeting efferocytosis receptors or signaling pathways involved in the recognition and engulfment of apoptotic cells. | Tailoring pharmacological interventions based on individual patient characteristics, such as genetic variations affecting efferocytosis receptors or underlying diseases. |
| Immunomodulatory Therapy | Utilization of immunomodulatory agents, such as cytokines, growth factors, or monoclonal antibodies, to enhance efferocytosis activity of immune cells. | Customizing immunomodulatory therapies based on the immune profile and responsiveness of individual patients, including considerations for immune cell function and cytokine levels. |
| Stem Cell Therapy | Administration of stem cells, such as mesenchymal stem cells (MSCs), which possess immunomodulatory properties and can promote efferocytosis by enhancing phagocytic activity of macrophages. | Selecting optimal stem cell sources and dosages based on individual patient characteristics, including age, underlying diseases, and immune status. |
| Gene Therapy | Genetic manipulation of efferocytosis-related genes or pathways using gene editing technologies, viral vectors, or RNA interference to enhance phagocytic capacity of immune cells. | Targeting specific genetic mutations or polymorphisms associated with impaired efferocytosis in individual patients and designing personalized gene therapy approaches. |
| Nutritional Interventions | Dietary interventions targeting nutrient deficiencies or imbalances that may impair efferocytosis function, such as omega-3 fatty acids, antioxidants, or vitamins. | Designing personalized nutrition plans based on individual dietary habits, nutritional status, and metabolic needs to optimize efferocytosis efficiency and overall immune function. |
| Lifestyle Modifications | Adoption of lifestyle habits, such as regular physical activity, stress management, and smoking cessation, which can positively influence immune function and efferocytosis activity. | Tailoring lifestyle recommendations to accommodate individual preferences, capabilities, and socioeconomic factors. |
| Combination Therapies | Integration of multiple therapeutic modalities to synergistically enhance efferocytosis and overall immune function. | Developing personalized combination therapy regimens based on individual patient profiles, including disease severity, treatment response, and potential drug interactions. |
| Approach | Description |
|---|---|
| Dietary Modifications | Emphasize a balanced diet rich in fruits, vegetables, whole grains, lean proteins, and healthy fats. Reduce intake of processed foods, sugary beverages, saturated fats, and trans fats. Monitor portion sizes and caloric intake. |
| Regular Physical Activity | Engage in regular aerobic exercise, strength training, and flexibility exercises. Aim for at least 150 min of moderate-intensity or 75 min of vigorous-intensity exercise per week. Incorporate physical activity into daily routines. |
| Weight Management | Maintain a healthy body weight through a combination of diet, exercise, and lifestyle modifications. Set realistic weight loss goals and seek support from healthcare professionals or support groups if needed. |
| Pharmacotherapy | Consider pharmacological interventions, such as anti-obesity medications or lipid-lowering drugs, under the guidance of a healthcare provider. May be recommended for individuals with obesity or dyslipidemia. |
| Bariatric Surgery | Surgical procedures such as gastric bypass, sleeve gastrectomy, or gastric banding may be considered for individuals with severe obesity or obesity-related comorbidities who have not responded to other weight loss interventions. |
| Nutritional Supplements | Consider supplementation with vitamins, minerals, and omega-3 fatty acids to address nutrient deficiencies and support overall health. Consult with healthcare provider or registered dietitian for personalized guidance. |
| Lifestyle Modifications | Adopt healthy lifestyle habits such as getting adequate sleep, managing stress, and avoiding smoking and excessive alcohol consumption. Practice mindful eating, focusing on hunger cues, portion control, and enjoyment of food. |
| Medical Monitoring | Regularly monitor blood glucose levels, lipid profiles, blood pressure, and other metabolic parameters to assess health status and track progress. Schedule routine check-ups with healthcare providers for personalized guidance. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mann, V.; Sundaresan, A.; Shishodia, S. Overnutrition and Lipotoxicity: Impaired Efferocytosis and Chronic Inflammation as Precursors to Multifaceted Disease Pathogenesis. Biology 2024, 13, 241. https://doi.org/10.3390/biology13040241
Mann V, Sundaresan A, Shishodia S. Overnutrition and Lipotoxicity: Impaired Efferocytosis and Chronic Inflammation as Precursors to Multifaceted Disease Pathogenesis. Biology. 2024; 13(4):241. https://doi.org/10.3390/biology13040241
Chicago/Turabian StyleMann, Vivek, Alamelu Sundaresan, and Shishir Shishodia. 2024. "Overnutrition and Lipotoxicity: Impaired Efferocytosis and Chronic Inflammation as Precursors to Multifaceted Disease Pathogenesis" Biology 13, no. 4: 241. https://doi.org/10.3390/biology13040241
APA StyleMann, V., Sundaresan, A., & Shishodia, S. (2024). Overnutrition and Lipotoxicity: Impaired Efferocytosis and Chronic Inflammation as Precursors to Multifaceted Disease Pathogenesis. Biology, 13(4), 241. https://doi.org/10.3390/biology13040241