
Impact of Polyethylene Terephthalate Microplastics on Drosophila melanogaster Biological Profiles and Heat Shock Protein Levels



Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods and Materials
2.1. Formation of Polyethylene Terephthalate (PET) Microplastics
2.2. Culturing of Model Organisms for Examination of the Health Effects of PET MP
2.3. Drosophila Treatment Protocol Using PET MP Exposure
2.4. PET MP Internalization in Drosophila
2.5. Determination of Cellular Toxicity in the Gut Region (Trypan Blue Staining)
2.6. Preparation of Homogenates
2.7. Total Protein Content, Cu-Zn Superoxide Dismutase (SOD) Activity, Catalase (CAT) Activity, and Lipid Peroxidation
2.8. Dye Exclusion Test of the Ovaries and Testes of Drosophila to Determine Reproductive Toxicity
2.9. Fertility, Fecundity, and Reproductive Performance
2.10. Qualitative RT-PCR Analysis of the Stress Genes hsp83, hs70, hsp60, and hsp26
2.11. cDNA Synthesis
2.12. Polymerase Chain Reaction (PCR)
3. Statistical Analyses
4. Results
4.1. PET MP Internalization in Drosophila
4.2. Trypan Blue Assay
4.3. Total Protein Concentration, Cu-Zn SOD Activity, CAT Activity, and MDA Content
4.4. Cytotoxicity in Reproductive Organs
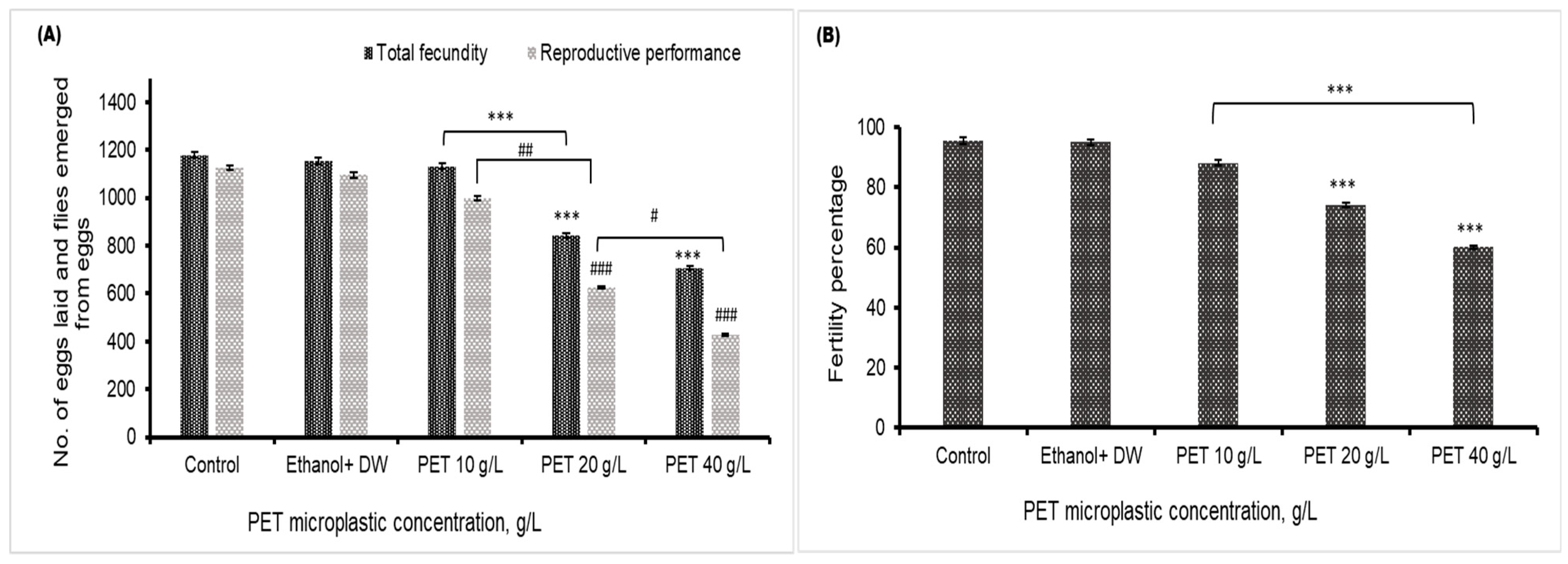
4.5. Fecundity, Fertility, and Reproductive Performance
4.6. RT-PCR Analysis
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akhbarizadeh, R.; Dobaradaran, S.; Nabipour, I.; Tajbakhsh, S.; Darabi, A.H.; Spitz, J. Abundance, composition, and potential intake of microplastics in canned fish. Mar. Pollut. Bull. 2020, 160, 111633. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Jia, Z. Recent insights into uptake, toxicity, and molecular targets of microplastics and nanoplastics relevant to human health impacts. Iscience 2023, 26, 106061. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, S.; Alqahtani, S.; Saquib, Q.; Mohiddin, F. Toxicological impact of microplastics and nanoplastics on humans: Understanding the mechanistic aspect of the interaction. Front. Toxicol. 2023, 5, 1193386. [Google Scholar] [CrossRef] [PubMed]
- Parenti, C.C.; Binelli, A.; Caccia, S.; Della Torre, C.; Magni, S.; Pirovano, G.; Casartelli, M. Ingestion and effects of polystyrene nanoparticles in the silkworm Bombyx mori. Chemosphere 2020, 257, 127203. [Google Scholar] [CrossRef] [PubMed]
- Matthews, S.; Xu, E.G.; Dumont, E.R.; Meola, V.; Pikuda, O.; Cheong, R.S.; Tufenkji, N. Polystyrene microand nanoplastics affect locomotion and daily activity of Drosophila melanogaster. Environ. Sci. Nano 2021, 8, 110–121. [Google Scholar] [CrossRef]
- Stock, V.; Laurisch, C.; Franke, J.; Dönmez, M.H.; Voss, L.; Böhmert, L.; Sieg, H. Uptake and cellular effects of PE, PP, PET and PVC microplastic particles. Toxicol. Vitr. 2021, 70, 105021. [Google Scholar] [CrossRef] [PubMed]
- Schymanski, D.; Goldbeck, C.; Humpf, H.U.; Fürst, P. Analysis of microplastics in water by micro-Raman spectroscopy: Release of plastic particles from different packaging into mineral water. Water Res. 2018, 129, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Verla, A.W.; Enyoh, C.E.; Verla, E.N.; Nwarnorh, K.O. Microplastic–toxic chemical interaction: A review study on quantified levels, mechanism and implication. SN Appl. Sci. 2019, 1, 1400. [Google Scholar] [CrossRef]
- Harvey, F.; Watts, J. Microplastics found in human stools for the first time. Guard 2018, 22, 1–5. [Google Scholar]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Giorgini, E. Plasticenta: First evidence of microplastics in human placenta. Environ. Int. 2021, 146, 106274. [Google Scholar] [CrossRef]
- Zhong, L.; Wu, T.; Sun, H.J.; Ding, J.; Pang, J.W.; Zhang, L.; Yang, S.S. Recent advances towards micro (nano) plastics research in wetland ecosystems: A systematic review on sources, removal, and ecological impacts. J. Hazard. Mater. 2023, 452, 131341. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Lee, I.S.; Jantrapirom, S.; Suda, K.; Yoshida, H. Drosophila models to study causative genes for human rare intractable neurological diseases. Exp. Cell Res. 2021, 403, 112584. [Google Scholar] [CrossRef] [PubMed]
- Saibil, H. Chaperone machines for protein folding, unfolding and disaggregation. Nat. Rev. Mol. Cell Biol. 2013, 14, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, L.; Bolaender, A.; J Patel, H.; Taldone, T. Heat shock protein (HSP) drug discovery and development: Targeting heat shock proteins in disease. Curr. Top. Med. Chem. 2016, 16, 2753–2764. [Google Scholar] [CrossRef] [PubMed]
- Van Noort, J.M.; Bsibsi, M.; Nacken, P.; Gerritsen, W.H.; Amor, S. The link between small heat shock proteins and the immune system. Int. J. Biochem. Cell Biol. 2012, 44, 1670–1679. [Google Scholar] [CrossRef]
- Mudiam, M.K.R.; Singh, M.P.; Chowdhuri, D.K.; Murthy, R.C. Quantitative evaluation of benzene, toluene, and xylene in the larvae of Drosophila melanogaster by solid-phase microextraction/gas chromatography/mass spectrometry for potential use in toxicological studies. J. AOAC Int. 2010, 93, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
- El Kholy, S.; Wang, K.; El-Seedi, H.R.; Al Naggar, Y. Dopamine modulates Drosophila gut physiology, providing new insights for future gastrointestinal pharmacotherapy. Biology 2021, 10, 983. [Google Scholar] [CrossRef] [PubMed]
- Alaraby, M.; Annangi, B.; Marcos, R.; Hernández, A. Drosophila melanogaster as a suitable in vivo model to determine potential side effects of nanomaterials: A review. J. Toxicol. Environ. Health Part B 2016, 19, 65–104. [Google Scholar] [CrossRef] [PubMed]
- Kauts, S.; Mishra, Y.; Yousuf, S.; Bhardwaj, R.; Singh, S.K.; Alshabrmi, F.M.; Abdurahman, M.; Vamanu, E.; Singh, M.P. Toxicological Profile of Polyethylene Terephthalate (PET) Microplastic in Ingested Drosophila melanogaster (Oregon R+) and Its Adverse Effect on Behavior and Development. Toxics 2023, 11, 782. [Google Scholar] [CrossRef]
- Singh, M.P.; Ram, K.R.; Mishra, M.; Shrivastava, M.; Saxena, D.K.; Chowdhuri, D.K. Effects of coexposure of benzene, toluene and xylene to Drosophila melanogaster: Alteration in hsp70, hsp60, hsp83, hsp26, ROS generation and oxidative stress markers. Chemosphere 2010, 79, 577–587. [Google Scholar] [CrossRef]
- Singh, M.P.; Shabir, S.; Deopa, A.S.; Raina, S.R.; Bantun, F.; Jalal, N.A.; Vamanu, E. Synthesis of green engineered silver nanoparticles through Urtica dioica: An inhibition of microbes and alleviation of cellular and organismal toxicity in Drosophila melanogaster. Antibiotics 2022, 11, 1690. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Liang, B.; Zhang, D.; Li, Y.; Tang, H.; Zhong, L.; Xu, Y. Effects of PET MP on the physiology of Drosophila. Chemosphere 2021, 283, 131289. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Zhang, D.; Liu, X.; Xu, Y.; Tang, H.; Li, Y.; Shen, J. Sex-specific effects of PET-MPs on Drosophila lifespan. Arch. Insect Biochem. Physiol. 2022, 110, e21909. [Google Scholar] [CrossRef] [PubMed]
- Shruti, V.C.; Pérez-Guevara, F.; Roy, P.D.; Kutralam-Muniasamy, G. Analysing microplastics with Nile Red: Emerging trends, challenges, and prospects. J. Hazard. Mater. 2022, 423, 127171. [Google Scholar] [CrossRef] [PubMed]
- Villacorta, A.; Rubio, L.; Alaraby, M.; López-Mesas, M.; Fuentes-Cebrian, V.; Moriones, O.H.; Hernández, A. A new source of representative secondary PET nanoplastics. Obtention, characterization, and hazard evaluation. J. Hazard. Mater. 2022, 439, 129593. [Google Scholar] [CrossRef]
- Alaraby, M.; Villacorta, A.; Abass, D.; Hernández, A.; Marcos, R. The hazardous impact of true-to-life PET nanoplastics in Drosophila. Sci. Total Environ. 2023, 863, 160954. [Google Scholar] [CrossRef] [PubMed]
- Krebs, R.A.; Feder, M.E. Deleterious consequences of Hsp70 overexpression in Drosphilla melanogaster larvae. Cell Stress Chaperones 1997, 2, 60. [Google Scholar] [CrossRef] [PubMed]
- Shabir, S.S.; Sehgal, A.; Dutta, J.; Devgon, I.; Singh, S.K.; Alsanie, W.F.; Alamri, A.S.; Alhomrani, M.; Alsharif, A.; Basalamah, M.A.M.; et al. Therapeutic Potential of Green-Engineered ZnO Nanoparticles on Rotenone-Exposed D. melanogaster (Oregon R+): Unveiling Ameliorated Biochemical, Cellular, and Behavioral Parameters. Antioxidants 2023, 12, 1679. [Google Scholar] [CrossRef] [PubMed]
- Himalian, R.; Singh, M.P. A Comparative account on Antioxidant activities, Total phenolic and Flavonoid contents of Punica granatum, Carica papaya, Foeniculum vulgare, Trigonella foenum-graecum, and Urtica dioica: An in vitro Evaluation. Res. J. Pharm. Technol. 2022, 15, 1175–1183. [Google Scholar] [CrossRef]
- Singh, M.P.; Mishra, M.; Sharma, A.; Shukla, A.K.; Mudiam, M.K.; Patel, D.K.; Ram, K.R.; Chowdhuri, D.K. Genotoxicity, and apoptosis in Drosophila melanogaster exposed to benzene, toluene, and xylene: Attenuation by quercetin and curcumin. Toxicol. Appl. Pharmacol. 2011, 253, 14–30. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, L.C.; Dwivedi, S.; Raihan, F.; Yathisha, U.G.; Raghu, S.V.; Mamatha, B.S.; Sharma, A. Hsp70 overexpression in Drosophila hemocytes attenuates benzene-induced immune and developmental toxicity by regulating ROS/JNK signalling pathway. Environ. Toxicol. 2022, 37, 1723–1739. [Google Scholar] [CrossRef] [PubMed]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Siddique, H.R.; Saxena, D.K.; Kar Chowdhuri, D. Hazardous effect of organophosphate compound, dichlorvos in transgenic Drosophila melanogaster (hsp70-lacZ): Induction of hsp70, antioxidant enzymes and inhibition of acetylcholinesterase. Biochim. Biophys. Acta 2005, 1725, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Academic Press: Cambridge, MA, USA, 1974; pp. 673–684. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Shabir, S.; Yousuf, S.; Singh, S.K.; Vamanu, E.; Singh, M.P. Ethnopharmacological effects of Urtica dioica, Matricaria chamomilla, and Murraya koenigii on rotenone-exposed D. melanogaster: An attenuation of cellular, biochemical, and organismal markers. Antioxidants 2022, 11, 1623. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, I.; Siddique, H.R.; Bajpai, V.K.; Saxena, D.K.; Chowdhuri, D.K. Synthetic pyrethroid cypermethrin induced cellular damage in reproductive tissues of Drosophila melanogaster: Hsp70 as a marker of cellular damage. Arch. Environ. Contam. Toxicol. 2006, 51, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, M.V.; Krishnamurthy, N.B. Studies on the toxicity of mercurial fungicide Agallol3 in Drosophila melanogaster. Environ. Res. 1981, 24, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Reddy, M.K.; Mathur, N.; Saxena, D.K.; Chowdhuri, D.K. Induction of hsp70, hsp60, hsp83 and hsp26 and oxidative stress markers in benzene, toluene and xylene exposed Drosophila melanogaster: Role of ROS generation. Toxicol. Appl. Pharmacol. 2009, 235, 226–243. [Google Scholar] [CrossRef]
- Song, Y.; Cao, C.; Qiu, R.; Hu, J.; Liu, M.; Lu, S.; Shi, H.; Raley-Susman, K.M.; He, D. Uptake and adverse effects of polyethylene terephthalate microplastics fibres on terrestrial snails (Achatina fulica) after soil exposure. Environ. Pollut. 2019, 250, 447–455. [Google Scholar] [CrossRef]
- Kim, D.; Kim, H.; An, Y.J. Effects of synthetic and natural microfibers on Daphnia magna—Are they dependent on microfiber type? Aquat. Toxicol. 2021, 240, 105968. [Google Scholar] [PubMed]
- Zhang, Y.; Wolosker, M.B.; Zhao, Y.; Ren, H.; Lemos, B. Exposure to microplastics cause gut damage, locomotor dysfunction, epigenetic silencing, and aggravate cadmium (Cd) toxicity in Drosophila. Sci. Total Environ. 2020, 744, 140979. [Google Scholar] [CrossRef]
- Ferrante, M.C.; Monnolo, A.; Del Piano, F.; Mattace Raso, G.; Meli, R. The pressing issue of micro- and nanoplastic contamination: Profiling the reproductive alterations mediated by oxidative stress. Antioxidants 2022, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Liu, Z.; Wu, D.; Chen, M.; Lv, W.; Zhao, Y. Accumulation of polystyrene microplastics in juvenile Eriocheir sinensis and oxidative stress effects in the liver. Aquat. Toxicol. 2018, 200, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yang, X.; Wang, Y.; Liu, R.; Gu, H.; Mao, L. Influence of polystyrene microplastics on rotifer (Brachionus calyciflorus) growth, reproduction, and antioxidant responses. Aquat. Ecol. 2021, 55, 1097–1111. [Google Scholar] [CrossRef]
- Vimal, D.; Saini, S.; Kristipati, R.R.; Chowdhuri, D.K. Atrazine or bisphenol A mediated negative modulation of mismatch repair gene, mlh1 leads to defective oogenesis and reduced female fertility in Drosophila melanogaster. Chemosphere 2019, 225, 247–258. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Pragya, P.; Ram, K.R.; Chowdhuri, D.K. Environmental chemical mediated male reproductive toxicity: Drosophila melanogaster as an alternate animal model. Theriogenology 2011, 76, 197–216. [Google Scholar] [PubMed]
- Zhao, Q.; Zhu, L.; Weng, J.; Jin, Z.; Cao, Y.; Jiang, H.; Zhang, Z. Detection and characterization of microplastics in the human testis and semen. Sci. Total Environ. 2023, 877, 162713. [Google Scholar] [CrossRef]
- Gupta, P.; Mahapatra, A.; Suman, A.; Ray, S.S.; Malafaia, G.; Singh, R.K. Polystyrene microplastics disrupt female reproductive health and fertility via sirt1 modulation in zebrafish (Danio rerio). J. Hazard. Mater. 2023, 460, 132359. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Zhao, Y.; Zhao, J.; Yu, T.; Yao, Y.; Su, J. Reproductive toxicity of microplastics in female mice and their offspring from induction of oxidative stress. Environ. Pollut. 2023, 327, 121482. [Google Scholar] [CrossRef]
- Li, H.; Zeng, L.; Wang, C.; Shi, C.; Li, Y.; Peng, Y.; Huang, Y. Review of the toxicity and potential molecular mechanisms of parental or successive exposure to environmental pollutants in the model organism Caenorhabditis elegans. Environ. Pollut. 2022, 311, 119927. [Google Scholar] [CrossRef]
- Chen, H.; Gu, Y.; Jiang, Y.; Yu, J.; Chen, C.; Shi, C.; Li, H. Photoaged Polystyrene Nanoplastics Result in Transgenerational Reproductive Toxicity Associated with the Methylation of Histone H3K4 and H3K9 in Caenorhabditis elegans. Environ. Sci. Technol. 2023, 57, 19341–19351. [Google Scholar] [CrossRef]
- Sobhani, Z.; Panneerselvan, L.; Fang, C.; Naidu, R.; Megharaj, M. Chronic and transgenerational effects of polyethylene microplastics at environmentally relevant concentrations in earthworms. Environ. Technol. Innov. 2022, 25, 102226. [Google Scholar] [CrossRef]
- Jaikumar, I.M.; Periyakali, S.B.; Rajendran, U.; Joen-Rong, S.; Thanasekaran, J.; Tsorng-Harn, F. Effects of microplastics, polystyrene, and polyethylene on antioxidants, metabolic enzymes, HSP-70, and myostatin expressions in the giant river prawn Macrobrachium rosenbergii: Impact on survival and growth. Arch. Environ. Contam. Toxicol. 2021, 80, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Gong, C.; Zheng, X.; Hu, F.; Liu, J.; Wang, T.; Li, R. Early clues and molecular mechanism involved in neurodegenerative diseases induced in immature mice by combined exposure to polypropylene microplastics and DEHP. Environ. Pollut. 2023, 336, 122406. [Google Scholar] [CrossRef] [PubMed]
- Ciocca, D.R.; Calderwood, S.K. Heat shock proteins in cancer: Diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperon 2005, 10, 86–103. [Google Scholar] [CrossRef]
- Tausif, Y.M.; Thekkekkara, D.; Sai, T.E.; Jahagirdar, V.; Arjun, H.R.; Meheronnisha, S.K.; Banerjee, A. Heat shock protein paradigms in cancer progression: Future therapeutic perspectives. 3 Biotech 2024, 14, 96. [Google Scholar] [CrossRef]
- Somu, P.; Mohanty, S.; Basavegowda, N.; Yadav, A.K.; Paul, S.; Baek, K.H. The Interplay between Heat Shock Proteins and Cancer Pathogenesis: A Novel Strategy for Cancer Therapeutics. Cancers 2024, 16, 638. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Sumi, S.; Seervi, M. Heat shock proteins-driven stress granule dynamics: Yet another avenue for cell survival. Apoptosis 2021, 26, 371–384. [Google Scholar] [CrossRef]
- Kuppuswami, J.; Senthilkumar, G.P. Nutri-stress, mitochondrial dysfunction, and insulin resistance—Role of heat shock proteins. Cell Stress. Chaperon 2023, 28, 35–48. [Google Scholar] [CrossRef]
- Mohamed, S.; Sabita, U.; Rajendra, S.; Raman, D. Genotoxicity: Mechanisms, testing guidelines and methods. Glob. J. Pharm. Pharm. Sci. 2017, 1, 133–138. [Google Scholar] [CrossRef]
- Tagorti, G.; Kaya, B. Genotoxic effect of microplastics and COVID-19: The hidden threat. Chemosphere 2022, 286 Pt 3, 131898. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| hsp83 Forward | 5′CCCGTGGCTTCGAGGTGGTCT3′ |
| hsp83 Reverse | 5′TCTGGGCATCGTCGGTAGTCATAGG3′ |
| hsp70 Forward | 5′GAACGGGCCAAGCGCACACTCTC3′ |
| hsp70 Reverse | 5′TCCTGGATCTTGCCGCTCTGGTCTC3′ |
| hsp60 Forward | 5′CCTCCGGCGGCATTGTCTTC3′ |
| hsp60 Reverse | 5′AGCGCATCGTAGCCGTAGTCACC3′ |
| hsp26 Forward | 5′CAAGCAGCTGAACAAGCTAACAATCTG3′ |
| hsp26 Reverse | 5′GCATGATGTGACCATGGTCGTCCTGG3′ |
| β actin Forward | 5′CCTCCGGCGGCATTGTCTTC3′ |
| β actin Reverse | 5′GGGCGGTGATCTCCTTCTGC3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kauts, S.; Mishra, Y.; Singh, M.P. Impact of Polyethylene Terephthalate Microplastics on Drosophila melanogaster Biological Profiles and Heat Shock Protein Levels. Biology 2024, 13, 293. https://doi.org/10.3390/biology13050293
Kauts S, Mishra Y, Singh MP. Impact of Polyethylene Terephthalate Microplastics on Drosophila melanogaster Biological Profiles and Heat Shock Protein Levels. Biology. 2024; 13(5):293. https://doi.org/10.3390/biology13050293
Chicago/Turabian StyleKauts, Simran, Yachana Mishra, and Mahendra P. Singh. 2024. "Impact of Polyethylene Terephthalate Microplastics on Drosophila melanogaster Biological Profiles and Heat Shock Protein Levels" Biology 13, no. 5: 293. https://doi.org/10.3390/biology13050293
APA StyleKauts, S., Mishra, Y., & Singh, M. P. (2024). Impact of Polyethylene Terephthalate Microplastics on Drosophila melanogaster Biological Profiles and Heat Shock Protein Levels. Biology, 13(5), 293. https://doi.org/10.3390/biology13050293