The Biology of Neisseria Adhesins


Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
| Neisseria species | Colonisation sites | Clinical disease | |
|---|---|---|---|
| Obligate pathogen | N. meningitidis | Nasopharynx | See text for details |
| N. gonorrhoeae | Urogenital tract | See text for details | |
| Opportunistic | N. lactamica | Nasopharynx | Meningitis, septicaemia |
| N. sicca | Nasopharynx | Endocarditis [6], bacteremia [7], meningitis | |
| N. subflava, which contains previous spp. N. flava, N. perflava and N. subflava | Nasopharynx Urogenital tract | Bacteremia [7], meningitis | |
| N. cinerea | Nasopharynx | Newborn ocular infection | |
| N. elongate, including 3 subspecies: N. elongate subsp. elongate, subsp. glycolytica and subsp. nitroreducens. | Nasopharynx | Septicaemia, endocarditis, brain abscess [8] | |
| N. flavescens | Nasopharynx | Meningitis, septicaemia, endocarditis | |
| N. mucosa | Nasopharynx | Pneumonia | |
| N. pharyngis | Nasopharynx | Septic arthritis | |
| N. polysaccharea | Nasopharynx | Not known | |
| N. canis | Isolated from the throats of cats | Cat-bite wound infection | |
| N. weaveri | Normal oral flora in dogs | Dog-bite wound infection | |
| N. iguana | Zoonotic | Not known | |
| N. animalis | Isolated from the throats of guinea pigs | Not known | |
| N. denitrificans | Isolated from the throats of guinea pigs | Not known | |
| N. dentiae | Zoonotic; isolated from dental plaque of domestic cows | Not known | |
| N. macacae | Isolated from the oropharynges of rhesus monkeys | Not known | |
| N. zoodegmatis & N. animaloris | Commensal organisms in the oral cavity of dogs and cats | Systemic infections in humans and animals; dog-bite wound infection [9] | |
| N. bacilliformis | Not known | Wound infection, respiratory tract infection [10], endocarditis [11,12] | |
| N. skkuensis | Not known | Isolated from a foot ulcer in a diabetic patient [13] | |
| N. wadsworthii & N. shayeganii | Not known | Wound infection [14] | |
| N. oralis | Healthy subgingival plaque [15] | Not known | |
| False | N. caviae, N. ovis, N. cuniculi [16] | ||
1.1. Meningococcal Disease
1.2. Gonococcal Disease
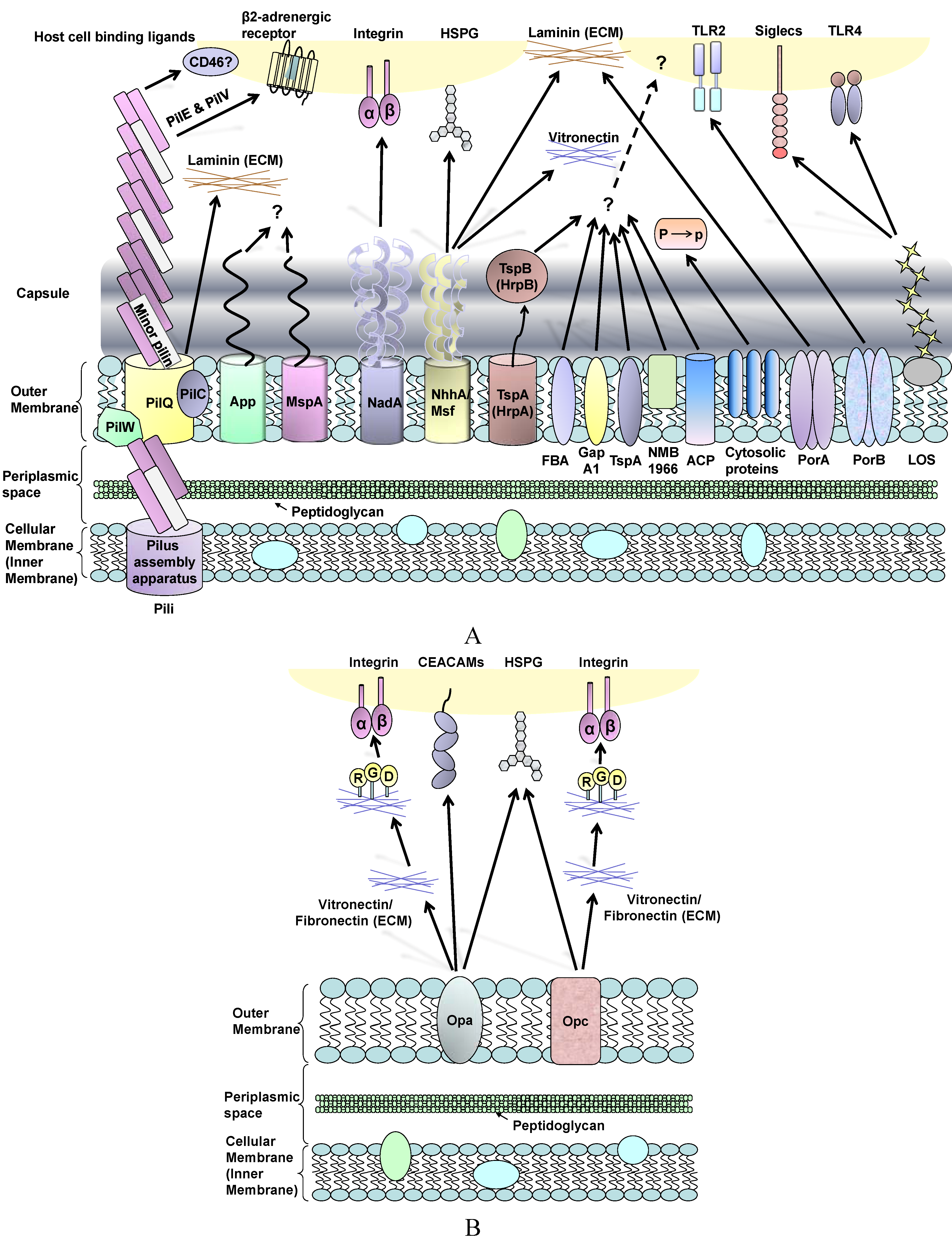
2. Neisseria Surface Structures Involved in Adhesion
2.1. Pilus
2.2. Opacity-Associated Proteins: Opa and Opc
2.3. Classical Monomeric Autotransporters: App and MspA/AusI
2.4. Trimeric Autotransporter Adhesins: NadA and NhhA
2.5. Two-Partner Secretion (TPS) System: HrpA and HrpB
2.6. Other Neisseria Adhesins
2.7. Other Neisseria Surface Structures Influencing Adhesion
2.7.1. Polysaccharide Capsule
2.7.2. Lipo-Oligosaccharide (LOS)
2.7.3. Porin (Por)
3. Mechanisms of Neisseria Adhesion with Host Cells

3.1. Neisseria meningitidis Adherence to Nasopharyngeal Epithelial Cells
3.2. Neisseria Gonorrhoeae Adhesion to the Urogenital Tract
3.2.1. Gonococcal Infection of the Human Lower Reproductive Tract
3.2.2. Ascending Gonococcal Infection
3.3. Adherence of Neisseria spp. to Endothelial Cells

3.4. Interactions of Neisseriae with Immune Effector Cells
| Neisseria ligand | TLR, cell expression and host response | Reference |
|---|---|---|
| Neisseria LPS | TLR4 | See text for details |
| Neisseria PorB | TLR2 | See text for details |
| Meningococcal NhhA | TLR4 activation in macrophages | [348] |
| Product of meningococcal NMB1468 (Ag-473) | TLR4 activation in bone marrow-derived dendritic cells | [349] |
| Meningococcal penicillin-binding proteins | TLR4 activation of dendritic cells | [350] |
| Meningococcal recombinant NadA(Δ351-405) | Binds to monocyte HSP90 and forms a transducing complex of HSP90/HSP70/TLR4 | [351] |
| Meningococcal CPS | Macrophage recognition via TLR2- and TLR4-MD-2 pathways. | [352] |
| Meningococcal membrane-associated proteins (LPS− background) | Activate TLR2-CD14 in monocytes/macrophages | [353,354] |
| Gonococcal Lip (H.8) protein | Stimulates human endocervical epithelial cells in TLR2-dependent manner to secrete cytokines | [355] |
3.5. Interaction of Neisseria with the Meninges
4. Conclusions

Acknowledgements
Conflict of Interest
References
- Weichselbaum, A. Ueber die Aetiologie der akuten meningitis cerebro-spinalis. Fortschr. Med. 1887, 5, 573–583; 620–626. [Google Scholar]
- Marchiafava, E.; Celli, A. Spra i micrococchi della meningite cerebrospinale epidemica. Gazzetta Osp. 1884, 5, 59. [Google Scholar]
- Heubner, J.O.L. Beobachtungen und versuche über den meningokokkus intracellularis (Weichselbaum-Jaeger). Jb Kinderheilk 1896, 43, 1–22. [Google Scholar]
- Kiefer, F. Zur differentialdiagnose des erregers der epidemischen cerebrospinal-meningitis und der gonorrhoe. Berl. Klin. Wochenschr. 1896, 33, 628–630. [Google Scholar]
- Goodwin, M.E.; von Sholly, A.I. The frequent occurrence of meningococci in the nasal cavities of meningitis patients and of those in direct contact with them. Public Health Pap. Rep. 1905, 31, 21–34. [Google Scholar]
- Sommerstein, R.; Ramsay, D.; Dubuis, O.; Waser, S.; Aebersold, F.; Vogt, M. Fatal Neisseria sicca endocarditis. Infection 2013, 41, 747–749. [Google Scholar] [CrossRef]
- Jung, J.J.; Vu, D.M.; Clark, B.; Keller, F.G.; Spearman, P. Neisseria sicca/subflava bacteremia presenting as cutaneous nodules in an immunocompromised host. Pediatr. Infect. Dis. J. 2009, 28, 661–663. [Google Scholar] [CrossRef]
- Hsiao, J.F.; Lee, M.H.; Chia, J.H.; Ho, W.J.; Chu, J.J.; Chu, P.H. Neisseria elongata endocarditis complicated by brain embolism and abscess. J. Med. Microbiol. 2008, 57, 376–381. [Google Scholar] [CrossRef]
- Vandamme, P.; Holmes, B.; Bercovier, H.; Coenye, T. Classification of centers for disease control group eugonic fermenter (EF)-4a and EF-4b as Neisseria animaloris sp. nov. and Neisseria zoodegmatis sp . nov., respectively. Int. J. Syst. Evol. Microbiol. 2006, 56, 1801–1805. [Google Scholar] [CrossRef]
- Han, X.Y.; Hong, T.; Falsen, E. Neisseria bacilliformis sp. nov. isolated from human infections. J. Clin. Microbiol. 2006, 44, 474–479. [Google Scholar] [CrossRef]
- Masliah-Planchon, J.; Breton, G.; Jarlier, V.; Simon, A.; Benveniste, O.; Herson, S.; Drieux, L. Endocarditis due to Neisseria bacilliformis in a patient with a bicuspid aortic valve. J. Clin. Microbiol. 2009, 47, 1973–1975. [Google Scholar] [CrossRef]
- Abandeh, F.I.; Balada-Llasat, J.M.; Pancholi, P.; Risaliti, C.M.; Maher, W.E.; Bazan, J.A. A rare case of Neisseria bacilliformis native valve endocarditis. Diagn. Microbiol. Infect. Dis. 2012, 73, 378–379. [Google Scholar] [CrossRef]
- Lee, M.Y.; Park, E.G.; Choi, J.Y.; Cheong, H.S.; Chung, D.R.; Peck, K.R.; Song, J.H.; Ko, K.S. “Neisseria skkuensis” sp. nov., isolated from the blood of a diabetic patient with a foot ulcer. J. Med. Microbiol. 2010, 59, 856–859. [Google Scholar] [CrossRef]
- Wolfgang, W.J.; Carpenter, A.N.; Cole, J.A.; Gronow, S.; Habura, A.; Jose, S.; Nazarian, E.J.; Kohlerschmidt, D.J.; Limberger, R.; Schoonmaker-Bopp, D.; et al. Neisseria wadsworthii sp. nov. and Neisseria shayeganii sp. nov., isolated from clinical specimens. Int. J. Syst. Evol. Microbiol. 2011, 61, 91–98. [Google Scholar] [CrossRef]
- Wolfgang, W.J.; Passaretti, T.V.; Jose, R.; Cole, J.; Coorevits, A.; Carpenter, A.N.; Jose, S.; van Landschoot, A.; Izard, J.; Kohlerschmidt, D.J.; et al. Neisseria oralis sp. nov. isolated from healthy gingival plaque and clinical samples. Int. J. Syst. Evol. Microbiol. 2013, 63, 1323–1328. [Google Scholar] [CrossRef]
- Veron, M.; Lenvoise-Furet, A.; Coustere, C.; Ged, C.; Grimont, F. Relatedness of three species of “false Neisseriae,” Neisseria caviae, Neisseria cuniculi, and Neisseria ovis, by DNA-DNA hybridizations and fatty acid analysis. Int. J. Syst. Bacteriol. 1993, 43, 210–220. [Google Scholar] [CrossRef]
- Initiative for Vaccine Research (IVR). Bacterial Infections—Meningococcal Disease. Available online: http://www.who.int/vaccine_research/diseases/soa_bacterial/en/index1.html/ (accessed on 1 July 2013).
- Stephens, D.S. Conquering the meningococcus. FEMS Microbiol. Rev. 2007, 31, 3–14. [Google Scholar] [CrossRef]
- Vieusseux, M. Memoire sur la maladie qui a regne a Geneve au printemps de 1805. J. Med. Chir. Pharmacol. 1805, 11, 163–182. [Google Scholar]
- Wolf, R.E.; Birbara, C.A. Meningococcal infections at an army training center. Am. J. Med. 1968, 44, 243–255. [Google Scholar] [CrossRef]
- Apicella, M.A. Neisseria meningitidis. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 7th ed.; Mandell, G.L., Douglas, R.G., Bennett, J.E., Dolin, R., Eds.; Elsevier/Churchill Livingstone: Philadelphia, PA, USA, 2010; Chapter 211; pp. 2737–2752. [Google Scholar]
- Brandtzaeg, P.; van Deuren, M. Classification and Pathogenesis of Meningococcal Infections. In Neisseria meningitidis: Advanced Methods and Protocols; Christodoulides, M., Ed.; Humana Press: New York, NY, USA, 2012; Chapter 4; pp. 21–35. [Google Scholar]
- Stephens, D.S.; Greenwood, B.; Brandtzaeg, P. Epidemic meningitis, meningococcaemia, and Neisseria meningitidis. Lancet 2007, 369, 2196–2210. [Google Scholar] [CrossRef]
- Rosenstein, N.E.; Perkins, B.A.; Stephens, D.S.; Popovic, T.; Hughes, J.M. Meningococcal disease. N. Engl. J. Med. 2001, 344, 1378–1388. [Google Scholar] [CrossRef]
- Brasier, A.R.; Macklis, J.D.; Vaughan, D.; Warner, L.; Kirshenbaum, J.M. Myopericarditis as an initial presentation of meningococcemia. Unusual manifestation of infection with serotype W135. Am. J. Med. 1987, 82, 641–644. [Google Scholar] [CrossRef]
- Varon, J.; Chen, K.; Sternbach, G.L. Rupert waterhouse and carl friderichsen: Adrenal apoplexy. J. Emerg. Med. 1998, 16, 643–647. [Google Scholar] [CrossRef]
- Girard, M.P.; Preziosi, M.P.; Aguado, M.T.; Kieny, M.P. A review of vaccine research and development: Meningococcal disease. Vaccine 2006, 24, 4692–4700. [Google Scholar] [CrossRef]
- Fellick, J.M.; Sills, J.A.; Marzouk, O.; Hart, C.A.; Cooke, R.W.; Thomson, A.P. Neurodevelopmental outcome in meningococcal disease: A case-control study. Arch. Dis. Child. 2001, 85, 6–11. [Google Scholar] [CrossRef]
- Koomen, I.; van Furth, A.M.; Kraak, M.A.; Grobbee, D.E.; Roord, J.J.; Jennekens-Schinkel, A. Neuropsychology of academic and behavioural limitations in school-age survivors of bacterial meningitis. Dev. Med. Child. Neurol. 2004, 46, 724–732. [Google Scholar]
- Initiative for Vaccine Research (IVR). Sexually Transmitted Diseases—Gonorrhoea. Available online: http://www.who.int/vaccine_research/diseases/soa_std/en/index2.html/ (accessed on 1 July 2013).
- Marrazzo, J.M.; Handsfield, H.H.; Sparling, F.P. Neisseria gonorrhoeae. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 7th ed.; Mandell, G.L., Douglas, R.G., Bennett, J.E., Dolin, R., Eds.; Elsevier/Churchill Livingstone: Philadelphia, PA, USA, 2010; Chapter 212; pp. 2753–2769. [Google Scholar]
- Miller, W.C.; Ford, C.A.; Morris, M.; Handcock, M.S.; Schmitz, J.L.; Hobbs, M.M.; Cohen, M.S.; Harris, K.M.; Udry, J.R. Prevalence of chlamydial and gonococcal infections among young adults in the United States. JAMA 2004, 291, 2229–2236. [Google Scholar] [CrossRef]
- Kahn, R.H.; Mosure, D.J.; Blank, S.; Kent, C.K.; Chow, J.M.; Boudov, M.R.; Brock, J.; Tulloch, S. Chlamydia trachomatis and Neisseria gonorrhoeae prevalence and coinfection in adolescents entering selected US juvenile detention centers, 1997–2002. Sex. Transm. Dis. 2005, 32, 255–259. [Google Scholar] [CrossRef]
- Fleming, D.T.; Wasserheit, J.N. From epidemiological synergy to public health policy and practice: The contribution of other sexually transmitted diseases to sexual transmission of HIV infection. Sex. Transm. Infect. 1999, 75, 3–17. [Google Scholar] [CrossRef]
- CDC. Update to CDC’s Sexually transmitted diseases treatment guidelines, 2010: Oral cephalosporins no longer a recommended treatment for gonococcal infections. MMWR Morb. Mortal. Wkly. Rep. 2012, 61, 590–594.
- Campos-Outcalt, D. CDC update on gonorrhea: Expand treatment to limit resistance. J. Fam. Pract. 2011, 60, 736–740. [Google Scholar]
- Strom, M.S.; Lory, S. Structure-function and biogenesis of the type IV pili. Annu. Rev. Microbiol. 1993, 47, 565–596. [Google Scholar] [CrossRef]
- Merz, A.J.; So, M.; Sheetz, M.P. Pilus retraction powers bacterial twitching motility. Nature 2000, 407, 98–102. [Google Scholar] [CrossRef]
- Fussenegger, M.; Rudel, T.; Barten, R.; Ryll, R.; Meyer, T.F. Transformation competence and type-4 pilus biogenesis in Neisseria gonorrhoeae—A review. Gene 1997, 192, 125–134. [Google Scholar] [CrossRef]
- Virji, M.; Heckels, J.E. Antigenic cross-reactivity of Neisseria pili: Investigations with type- and species-specific monoclonal antibodies. J. Gen. Microbiol. 1983, 129, 2761–2768. [Google Scholar]
- Perry, A.C.; Nicolson, I.J.; Saunders, J.R. Neisseria meningitidis C114 contains silent, truncated pilin genes that are homologous to Neisseria gonorrhoeae pil sequences. J. Bacteriol. 1988, 170, 1691–1697. [Google Scholar]
- Hitchcock, P.J. Unified nomenclature for pathogenic Neisseria species. Clin. Microbiol. Rev. 1989, 2, S64–S65. [Google Scholar]
- Virji, M.; Heckels, J.E.; Potts, W.J.; Hart, C.A.; Saunders, J.R. Identification of epitopes recognized by monoclonal antibodies SM1 and SM2 which react with all pili of Neisseria gonorrhoeae but which differentiate between two structural classes of pili expressed by Neisseria meningitidis and the distribution of their encoding sequences in the genomes of Neisseria spp. J. Gen. Microbiol. 1989, 135, 3239–3251. [Google Scholar]
- Aho, E.L.; Keating, A.M.; McGillivray, S.M. A comparative analysis of pilin genes from pathogenic and nonpathogenic Neisseria species. Microb. Pathog. 2000, 28, 81–88. [Google Scholar] [CrossRef]
- Aho, E.L.; Urwin, R.; Batcheller, A.E.; Holmgren, A.M.; Havig, K.; Kulakoski, A.M.; Vomhof, E.E.; Longfors, N.S.; Erickson, C.B.; Anderson, Z.K.; et al. Neisserial pilin genes display extensive interspecies diversity. FEMS Microbiol. Lett. 2005, 249, 327–334. [Google Scholar] [CrossRef]
- Higashi, D.L.; Biais, N.; Weyand, N.J.; Agellon, A.; Sisko, J.L.; Brown, L.M.; So, M. N. elongata produces type IV pili that mediate interspecies gene transfer with N. gonorrhoeae. PLoS One 2011, 6, e21373:1–e21373:7. [Google Scholar]
- Hagblom, P.; Segal, E.; Billyard, E.; So, M. Intragenic recombination leads to pilus antigenic variation in Neisseria gonorrhoeae. Nature 1985, 315, 156–158. [Google Scholar] [CrossRef]
- Haas, R.; Meyer, T.F. The repertoire of silent pilus genes in Neisseria gonorrhoeae: Evidence for gene conversion. Cell 1986, 44, 107–115. [Google Scholar] [CrossRef]
- Segal, E.; Hagblom, P.; Seifert, H.S.; So, M. Antigenic variation of gonococcal pilus involves assembly of separated silent gene segments. Proc. Natl. Acad. Sci. USA 1986, 83, 2177–2181. [Google Scholar] [CrossRef]
- Perry, A.C.; Hart, C.A.; Nicolson, I.J.; Heckels, J.E.; Saunders, J.R. Inter-strain homology of pilin gene sequences in Neisseria meningitidis isolates that express markedly different antigenic pilus types. J. Gen. Microbiol. 1987, 133, 1409–1418. [Google Scholar]
- Carbonnelle, E.; Hill, D.J.; Morand, P.; Griffiths, N.J.; Bourdoulous, S.; Murillo, I.; Nassif, X.; Virji, M. Meningococcal interactions with the host. Vaccine 2009, 27, B78–B89. [Google Scholar] [CrossRef]
- Tinsley, C.R.; Heckels, J.E. Variation in the expression of pili and outer membrane protein by Neisseria meningitidis during the course of meningococcal infection. J. Gen. Microbiol. 1986, 132, 2483–2490. [Google Scholar]
- Criss, A.K.; Kline, K.A.; Seifert, H.S. The frequency and rate of pilin antigenic variation in Neisseria gonorrhoeae. Mol. Microbiol. 2005, 58, 510–519. [Google Scholar] [CrossRef]
- Helm, R.A.; Seifert, H.S. Frequency and rate of pilin antigenic variation of Neisseria meningitidis. J. Bacteriol. 2010, 192, 3822–3823. [Google Scholar] [CrossRef]
- Nassif, X.; Beretti, J.L.; Lowy, J.; Stenberg, P.; Ogaora, P.; Pfeifer, J.; Normark, S.; So, M. Roles of pilin and PilC in adhesion of Neisseria-meningitidis to human epithelial and endothelial cells. Proc. Natl. Acad. Sci. USA 1994, 91, 3769–3773. [Google Scholar]
- Parge, H.E.; McRee, D.E.; Capozza, M.A.; Bernstein, S.L.; Getzoff, E.D.; Tainer, J.A. Three dimensional structure of bacterial pili. Antonie Van Leeuwenhoek 1987, 53, 447–453. [Google Scholar] [CrossRef]
- Parge, H.E.; Bernstein, S.L.; Deal, C.D.; McRee, D.E.; Christensen, D.; Capozza, M.A.; Kays, B.W.; Fieser, T.M.; Draper, D.; So, M. Biochemical purification and crystallographic characterization of the fiber-forming protein pilin from Neisseria gonorrhoeae. J. Biol. Chem. 1990, 265, 2278–2285. [Google Scholar]
- Parge, H.E.; Forest, K.T.; Hickey, M.J.; Christensen, D.A.; Getzoff, E.D.; Tainer, J.A. Structure of the fibre-forming protein pilin at 2.6 A resolution. Nature 1995, 378, 32–38. [Google Scholar] [CrossRef]
- Forest, K.T.; Dunham, S.A.; Koomey, M.; Tainer, J.A. Crystallographic structure reveals phosphorylated pilin from Neisseria: Phosphoserine sites modify type IV pilus surface chemistry and fibre morphology. Mol. Microbiol. 1999, 31, 743–752. [Google Scholar] [CrossRef]
- Craig, L.; Volkmann, N.; Arvai, A.S.; Pique, M.E.; Yeager, M.; Egelman, E.H.; Tainer, J.A. Type IV pilus structure by cryo-electron microscopy and crystallography: Implications for pilus assembly and functions. Mol. Cell. 2006, 23, 651–662. [Google Scholar] [CrossRef]
- Carbonnelle, E.; Helaine, S.; Nassif, X.; Pelicic, V. A systematic genetic analysis in Neisseria meningitidis defines the Pil proteins required for assembly, functionality, stabilization and export of type IV pili. Mol. Microbiol. 2006, 61, 1510–1522. [Google Scholar] [CrossRef]
- Brown, D.R.; Helaine, S.; Carbonnelle, E.; Pelicic, V. Systematic functional analysis reveals that a set of seven genes is involved in fine-tuning of the multiple functions mediated by type IV pili in Neisseria meningitidis. Infect. Immun. 2010, 78, 3053–3063. [Google Scholar] [CrossRef]
- Georgiadou, M.; Castagnini, M.; Karimova, G.; Ladant, D.; Pelicic, V. Large-scale study of the interactions between proteins involved in type IV pilus biology in Neisseria meningitidis: Characterization of a subcomplex involved in pilus assembly. Mol. Microbiol. 2012, 84, 857–873. [Google Scholar] [CrossRef]
- Morand, P.C.; Bille, E.; Morelle, S.; Eugene, E.; Beretti, J.L.; Wolfgang, M.; Meyer, T.F.; Koomey, M.; Nassif, X. Type IV pilus retraction in pathogenic Neisseria is regulated by the PilC proteins. EMBO J. 2004, 23, 2009–2017. [Google Scholar] [CrossRef]
- Tonjum, T.; Freitag, N.E.; Namork, E.; Koomey, M. Identification and characterization of pilG, a highly conserved pilus-assembly gene in pathogenic Neisseria. Mol. Microbiol. 1995, 16, 451–464. [Google Scholar] [CrossRef]
- Drake, S.L.; Koomey, M. The product of the pilQ gene is essential for the biogenesis of type IV pili in Neisseria gonorrhoeae. Mol. Microbiol. 1995, 18, 975–986. [Google Scholar]
- Carbonnelle, E.; Helaine, S.; Prouvensier, L.; Nassif, X.; Pelicic, V. Type IV pilus biogenesis in Neisseria meningitidis: PilW is involved in a step occurring after pilus assembly, essential for fibre stability and function. Mol. Microbiol. 2005, 55, 54–64. [Google Scholar]
- Helaine, S.; Carbonnelle, E.; Prouvensier, L.; Beretti, J.L.; Nassif, X.; Pelicic, V. PilX, a pilus-associated protein essential for bacterial aggregation, is a key to pilus-facilitated attachment of Neisseria meningitidis to human cells. Mol. Microbiol. 2005, 55, 65–77. [Google Scholar]
- Cehovin, A.; Simpson, P.J.; McDowell, M.A.; Brown, D.R.; Noschese, R.; Pallett, M.; Brady, J.; Baldwin, G.S.; Lea, S.M.; Matthews, S.J.; et al. Specific DNA recognition mediated by a type IV pilin. Proc. Natl. Acad. Sci. USA 2013, 110, 3065–3070. [Google Scholar] [CrossRef]
- Collins, R.F.; Ford, R.C.; Kitmitto, A.; Olsen, R.O.; Tonjum, T.; Derrick, J.P. Three-dimensional structure of the Neisseria meningitidis secretin PilQ determined from negative-stain transmission electron microscopy. J. Bacteriol. 2003, 185, 2611–2617. [Google Scholar] [CrossRef]
- Collins, R.F.; Davidsen, L.; Derrick, J.P.; Ford, R.C.; Tonjum, T. Analysis of the PilQ secretin from Neisseria meningitidis by transmission electron microscopy reveals a dodecameric quaternary structure. J. Bacteriol. 2001, 183, 3825–3832. [Google Scholar]
- Collins, R.F.; Frye, S.A.; Kitmitto, A.; Ford, R.C.; Tonjum, T.; Derrick, J.P. Structure of the Neisseria meningitidis outer membrane PilQ secretin complex at 12 A resolution. J. Biol. Chem. 2004, 279, 39750–39756. [Google Scholar]
- Jain, S.; Moscicka, K.B.; Bos, M.P.; Pachulec, E.; Stuart, M.C.; Keegstra, W.; Boekema, E.J.; van der Does, C. Structural characterization of outer membrane components of the type IV pili system in pathogenic Neisseria. PLoS One 2011, 6, e16624:1–e16624:11. [Google Scholar]
- Berry, J.L.; Phelan, M.M.; Collins, R.F.; Adomavicius, T.; Tonjum, T.; Frye, S.A.; Bird, L.; Owens, R.; Ford, R.C.; Lian, L.Y.; et al. Structure and assembly of a trans-periplasmic channel for type IV pili in Neisseria meningitidis. PLoS Pathog. 2012, 8, e1002923:1–e1002923:15. [Google Scholar]
- Balasingham, S.V.; Collins, R.F.; Assalkhou, R.; Homberset, H.; Frye, S.A.; Derrick, J.P.; Tonjum, T. Interactions between the lipoprotein PilP and the secretin PilQ in Neisseria meningitidis. J. Bacteriol. 2007, 189, 5716–5727. [Google Scholar] [CrossRef]
- Golovanov, A.P.; Balasingham, S.; Tzitzilonis, C.; Goult, B.T.; Lian, L.Y.; Homberset, H.; Tonjum, T.; Derrick, J.P. The solution structure of a domain from the Neisseria meningitidis lipoprotein PilP reveals a new beta-sandwich fold. J. Mol. Biol. 2006, 364, 186–195. [Google Scholar] [CrossRef]
- Trindade, M.B.; Job, V.; Contreras-Martel, C.; Pelicic, V.; Dessen, A. Structure of a widely conserved type IV pilus biogenesis factor that affects the stability of secretin multimers. J. Mol. Biol. 2008, 378, 1031–1039. [Google Scholar] [CrossRef]
- Collins, R.F.; Saleem, M.; Derrick, J.P. Purification and three-dimensional electron microscopy structure of the Neisseria meningitidis type IV pilus biogenesis protein PilG. J. Bacteriol. 2007, 189, 6389–6396. [Google Scholar] [CrossRef]
- Helaine, S.; Dyer, D.H.; Nassif, X.; Pelicic, V.; Forest, K.T. 3D structure/function analysis of PilX reveals how minor pilins can modulate the virulence properties of type IV pili. Proc. Natl. Acad. Sci. USA 2007, 104, 15888–15893. [Google Scholar] [CrossRef]
- Forest, K.T.; Satyshur, K.A.; Worzalla, G.A.; Hansen, J.K.; Herdendorf, T.J. The pilus-retraction protein PilT: Ultrastructure of the biological assembly. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 978–982. [Google Scholar] [CrossRef]
- Swanson, J. Studies on gonococcus infection. XIV. Cell wall protein differences among color/opacity colony variants of Neisseria gonorrhoeae. Infect. Immun. 1978, 21, 292–302. [Google Scholar]
- Parkhill, J.; Achtman, M.; James, K.D.; Bentley, S.D.; Churcher, C.; Klee, S.R.; Morelli, G.; Basham, D.; Brown, D.; Chillingworth, T.; et al. Complete DNA sequence of a serogroup A strain of Neisseria meningitidis Z2491. Nature 2000, 404, 502–506. [Google Scholar] [CrossRef]
- Tettelin, H.; Saunders, N.J.; Heidelberg, J.; Jeffries, A.C.; Nelson, K.E.; Eisen, J.A.; Ketchum, K.A.; Hood, D.W.; Peden, J.F.; Dodson, R.J.; et al. Complete genome sequence of Neisseria meningitidis serogroup B strain MC58. Science 2000, 287, 1809–1815. [Google Scholar] [CrossRef]
- Bhat, K.S.; Gibbs, C.P.; Barrera, O.; Morrison, S.G.; Jahnig, F.; Stern, A.; Kupsch, E.M.; Meyer, T.F.; Swanson, J. The opacity proteins of Neisseria gonorrhoeae strain MS11 are encoded by a family of 11 complete genes. Mol. Microbiol. 1991, 5, 1889–1901. [Google Scholar] [CrossRef]
- Stern, A.; Meyer, T.F. Common mechanism controlling phase and antigenic variation in pathogenic Neisseriae. Mol. Microbiol. 1987, 1, 5–12. [Google Scholar] [CrossRef]
- Wolff, K.; Stern, A. Identification and characterization of specific sequences encoding pathogenicity associated proteins in the genome of commensal Neisseria species. FEMS Microbiol. Lett. 1995, 125, 255–263. [Google Scholar] [CrossRef]
- Toleman, M.; Aho, E.; Virji, M. Expression of pathogen-like Opa adhesins in commensal Neisseria: Genetic and functional analysis. Cell Microbiol. 2001, 3, 33–44. [Google Scholar] [CrossRef]
- Malorny, B.; Morelli, G.; Kusecek, B.; Kolberg, J.; Achtman, M. Sequence diversity, predicted two-dimensional protein structure, and epitope mapping of neisserial Opa proteins. J. Bacteriol. 1998, 180, 1323–1330. [Google Scholar]
- De Jonge, M.I.; Bos, M.P.; Hamstra, H.J.; Jiskoot, W.; van Ulsen, P.; Tommassen, J.; van, A.L.; van der Ley, P. Conformational analysis of opacity proteins from Neisseria meningitidis. Eur. J. Biochem. 2002, 269, 5215–5223. [Google Scholar] [CrossRef]
- Vandeputte-Rutten, L.; Bos, M.P.; Tommassen, J.; Gros, P. Crystal structure of Neisserial surface protein A (NspA), a conserved outer membrane protein with vaccine potential. J. Biol. Chem. 2003, 278, 24825–24830. [Google Scholar] [CrossRef]
- Hobbs, M.M.; Malorny, B.; Prasad, P.; Morelli, G.; Kusecek, B.; Heckels, J.E.; Cannon, J.G.; Achtman, M. Recombinational reassortment among opa genes from ET-37 complex Neisseria meningitidis isolates of diverse geographical origins. Microbiology 1998, 144, 157–166. [Google Scholar] [CrossRef]
- Bilek, N.; Ison, C.A.; Spratt, B.G. Relative contributions of recombination and mutation to the diversification of the opa gene repertoire of Neisseria gonorrhoeae. J. Bacteriol. 2009, 191, 1878–1890. [Google Scholar] [CrossRef]
- Stern, A.; Brown, M.; Nickel, P.; Meyer, T.F. Opacity genes in Neisseria gonorrhoeae: Control of phase and antigenic variation. Cell 1986, 47, 61–71. [Google Scholar] [CrossRef]
- Belland, R.J.; Morrison, S.G.; van der Ley, P.; Swanson, J. Expression and phase variation of gonococcal P.II genes in Escherichia coli involves ribosomal frameshifting and slipped-strand mispairing. Mol. Microbiol. 1989, 3, 777–786. [Google Scholar] [CrossRef]
- Mayer, L.W. Rates in vitro changes of gonococcal colony opacity phenotypes. Infect. Immun. 1982, 37, 481–485. [Google Scholar]
- Belland, R.J.; Morrison, S.G.; Carlson, J.H.; Hogan, D.M. Promoter strength influences phase variation of neisserial opa genes. Mol. Microbiol. 1997, 23, 123–135. [Google Scholar]
- Callaghan, M.J.; Jolley, K.A.; Maiden, M.C. Opacity-associated adhesin repertoire in hyperinvasive Neisseria meningitidis. Infect. Immun. 2006, 74, 5085–5094. [Google Scholar]
- Callaghan, M.J.; Buckee, C.; McCarthy, N.D.; Ibarz Pavon, A.B.; Jolley, K.A.; Faust, S.; Gray, S.J.; Kaczmarski, E.B.; Levin, M.; Kroll, J.S.; et al. Opa protein repertoires of disease-causing and carried meningococci. J. Clin. Microbiol. 2008, 46, 3033–3041. [Google Scholar] [CrossRef]
- Zhu, P.; Morelli, G.; Achtman, M. The opcA and (psi)opcB regions in Neisseria: Genes, pseudogenes, deletions, insertion elements and DNA islands. Mol. Microbiol. 1999, 33, 635–650. [Google Scholar] [CrossRef]
- Zhu, P.; Klutch, M.J.; Derrick, J.P.; Prince, S.M.; Tsang, R.S.; Tsai, C.M. Identification of opcA gene in Neisseria polysaccharea: Interspecies diversity of Opc protein family. Gene 2003, 307, 31–40. [Google Scholar] [CrossRef]
- Sarkari, J.; Pandit, N.; Moxon, E.R.; Achtman, M. Variable expression of the Opc outer membrane protein in Neisseria meningitidis is caused by size variation of a promoter containing poly-cytidine. Mol. Microbiol. 1994, 13, 207–217. [Google Scholar] [CrossRef]
- Prince, S.M.; Achtman, M.; Derrick, J.P. Crystal structure of the OpcA integral membrane adhesin from Neisseria meningitidis. Proc. Natl. Acad. Sci. USA 2002, 99, 3417–3421. [Google Scholar] [CrossRef]
- Luan, B.; Caffrey, M.; Aksimentiev, A. Structure refinement of the OpcA adhesin using molecular dynamics. Biophys. J. 2007, 93, 3058–3069. [Google Scholar] [CrossRef]
- Cherezov, V.; Liu, W.; Derrick, J.P.; Luan, B.; Aksimentiev, A.; Katritch, V.; Caffrey, M. In meso crystal structure and docking simulations suggest an alternative proteoglycan binding site in the OpcA outer membrane adhesin. Proteins 2008, 71, 24–34. [Google Scholar] [CrossRef]
- Hadi, H.A.; Wooldridge, K.G.; Robinson, K.; Ala’Aldeen, D.A. Identification and characterization of App: An immunogenic autotransporter protein of Neisseria meningitidis. Mol. Microbiol. 2001, 41, 611–623. [Google Scholar] [CrossRef]
- Serruto, D.; Adu-Bobie, J.; Scarselli, M.; Veggi, D.; Pizza, M.; Rappuoli, R.; Arico, B. Neisseria meningitidis App, a new adhesin with autocatalytic serine protease activity. Mol. Microbiol. 2003, 48, 323–334. [Google Scholar] [CrossRef]
- Turner, D.P.; Marietou, A.G.; Johnston, L.; Ho, K.K.; Rogers, A.J.; Wooldridge, K.G.; Ala'Aldeen, D.A. Characterization of MspA, an immunogenic autotransporter protein that mediates adhesion to epithelial and endothelial cells in Neisseria meningitidis. Infect. Immun. 2006, 74, 2957–2964. [Google Scholar] [CrossRef]
- Pohlner, J.; Halter, R.; Beyreuther, K.; Meyer, T.F. Gene structure and extracellular secretion of Neisseria gonorrhoeae IgA protease. Nature 1987, 325, 458–462. [Google Scholar] [CrossRef]
- Van Ulsen, P. Protein Folding in Bacterial Adhesion: Secretion and Folding of Classical Monomeric Autotransporters. In Bacterial Adhesion: Chemistry, Biology and Physics; Linke, D., Goldman, A., Eds.; Springer: Dordrecht, Germany, 2011; Chapter 8; pp. 125–142. [Google Scholar]
- Meng, G.; Spahich, N.; Kenjale, R.; Waksman, G.; St. Geme, J.W. Crystal structure of the Haemophilus influenzae Hap adhesin reveals an intercellular oligomerization mechanism for bacterial aggregation. EMBO J. 2011, 30, 3864–3874. [Google Scholar] [CrossRef]
- Cotter, S.E.; Surana, N.K.; St. Geme, J.W. Trimeric autotransporters: A distinct subfamily of autotransporter proteins. Trends Microbiol. 2005, 13, 199–205. [Google Scholar] [CrossRef]
- Cotter, S.E.; Surana, N.K.; Grass, S.; St. Geme, J.W. Trimeric autotransporters require trimerization of the passenger domain for stability and adhesive activity. J. Bacteriol. 2006, 188, 5400–5407. [Google Scholar] [CrossRef]
- Lyskowski, A.; Leo, J.C.; Goldman, A. Structure and biology of trimeric autotransporter adhesins. In Bacterial Adhesion: Chemistry, Biology and Physics; Linke, D., Goldman, A., Eds.; Springer: Dordrecht, Germany, 2011; Chapter 9; pp. 143–158. [Google Scholar]
- El Tahir, Y.; Skurnik, M. YadA, the multifaceted Yersinia adhesin. Int. J. Med. Microbiol. 2001, 291, 209–218. [Google Scholar] [CrossRef]
- Koretke, K.K.; Szczesny, P.; Gruber, M.; Lupas, A.N. Model structure of the prototypical non-fimbrial adhesin YadA of Yersinia enterocolitica. J. Struct. Biol. 2006, 155, 154–161. [Google Scholar] [CrossRef]
- St. Geme, J.W.; Cutter, D. The Haemophilus influenzae Hia adhesin is an autotransporter protein that remains uncleaved at the C terminus and fully cell associated. J. Bacteriol. 2000, 182, 6005–6013. [Google Scholar] [CrossRef]
- Cotter, S.E.; Yeo, H.J.; Juehne, T.; St. Geme, J.W. Architecture and adhesive activity of the Haemophilus influenzae Hsf adhesin. J. Bacteriol. 2005, 187, 4656–4664. [Google Scholar] [CrossRef]
- Comanducci, M.; Bambini, S.; Brunelli, B.; du-Bobie, J.; Arico, B.; Capecchi, B.; Giuliani, M.M.; Masignani, V.; Santini, L.; Savino, S.; et al. NadA, a novel vaccine candidate of Neisseria meningitidis. J. Exp. Med. 2002, 195, 1445–1454. [Google Scholar] [CrossRef]
- Tavano, R.; Capecchi, B.; Montanari, P.; Franzoso, S.; Marin, O.; Sztukowska, M.; Cecchini, P.; Segat, D.; Scarselli, M.; Arico, B.; et al. Mapping of the Neisseria meningitidis NadA cell-binding site: Relevance of predicted {alpha}-helices in the NH2-terminal and dimeric coiled-coil regions. J. Bacteriol. 2011, 193, 107–115. [Google Scholar] [CrossRef]
- Comanducci, M.; Bambini, S.; Caugant, D.A.; Mora, M.; Brunelli, B.; Capecchi, B.; Ciucchi, L.; Rappuoli, R.; Pizza, M. NadA diversity and carriage in Neisseria meningitidis. Infect. Immun. 2004, 72, 4217–4223. [Google Scholar] [CrossRef]
- Peak, I.R.; Srikhanta, Y.; Dieckelmann, M.; Moxon, E.R.; Jennings, M.P. Identification and characterisation of a novel conserved outer membrane protein from Neisseria meningitidis. FEMS Immunol. Med. Microbiol. 2000, 28, 329–334. [Google Scholar] [CrossRef]
- Scarselli, M.; Serruto, D.; Montanari, P.; Capecchi, B.; Adu-Bobie, J.; Veggi, D.; Rappuoli, R.; Pizza, M.; Arico, B. Neisseria meningitidis NhhA is a multifunctional trimeric autotransporter adhesin. Mol. Microbiol. 2006, 61, 631–644. [Google Scholar] [CrossRef]
- Griffiths, N.J.; Hill, D.J.; Borodina, E.; Sessions, R.B.; Devos, N.I.; Feron, C.M.; Poolman, J.T.; Virji, M. Meningococcal surface fibril (Msf) binds to activated vitronectin and inhibits the terminal complement pathway to increase serum resistance. Mol. Microbiol. 2011, 82, 1129–1149. [Google Scholar] [CrossRef]
- Hodak, H.; Jacob-Dubuisson, F. Current challenges in autotransport and two-partner protein secretion pathways. Res. Microbiol. 2007, 158, 631–637. [Google Scholar] [CrossRef]
- Hodak, H.; Clantin, B.; Willery, E.; Villeret, V.; Locht, C.; Jacob-Dubuisson, F. Secretion signal of the filamentous haemagglutinin, a model two-partner secretion substrate. Mol. Microbiol. 2006, 61, 368–382. [Google Scholar] [CrossRef]
- Jacob-Dubuisson, F.; Fernandez, R.; Coutte, L. Protein secretion through autotransporter and two-partner pathways. Biochim. Biophys. Acta 2004, 1694, 235–257. [Google Scholar] [CrossRef]
- Jacob-Dubuisson, F.; Locht, C.; Antoine, R. Two-partner secretion in Gram-negative bacteria: A thrifty, specific pathway for large virulence proteins. Mol. Microbiol. 2001, 40, 306–313. [Google Scholar] [CrossRef]
- Clantin, B.; Hodak, H.; Willery, E.; Locht, C.; Jacob-Dubuisson, F.; Villeret, V. The crystal structure of filamentous hemagglutinin secretion domain and its implications for the two-partner secretion pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 6194–6199. [Google Scholar]
- Schmitt, C.; Turner, D.; Boesl, M.; Abele, M.; Frosch, M.; Kurzai, O. A functional two-partner secretion system contributes to adhesion of Neisseria meningitidis to epithelial cells. J. Bacteriol. 2007, 189, 7968–7976. [Google Scholar] [CrossRef]
- Snyder, L.A.; Saunders, N.J. The majority of genes in the pathogenic Neisseria species are present in non-pathogenic Neisseria lactamica, including those designated as “virulence genes”. BMC Genomics 2006, 7, 128:1–128:11. [Google Scholar]
- Van Ulsen, P.; Rutten, L.; Feller, M.; Tommassen, J.; van der Ende, A. Two-partner secretion systems of Neisseria meningitidis associated with invasive clonal complexes. Infect. Immun. 2008, 76, 4649–4658. [Google Scholar] [CrossRef]
- Ur Rahman, S.; van Ulsen, P. System specificity of the TpsB transporters of coexpressed two-partner secretion systems of Neisseria meningitidis. J. Bacteriol. 2013, 195, 788–797. [Google Scholar] [CrossRef]
- Kizil, G.; Todd, I.; Atta, M.; Borriello, S.P.; Ait-Tahar, K.; Ala’aldeen, D.A. Identification and characterization of TspA, a major CD4(+) T-cell- and B-cell-stimulating Neisseria-specific antigen. Infect. Immun. 1999, 67, 3533–3541. [Google Scholar]
- Oldfield, N.J.; Bland, S.J.; Taraktsoglou, M.; Dos Ramos, F.J.; Robinson, K.; Wooldridge, K.G.; Ala'aldeen, D.A. T-cell stimulating protein A (TspA) of Neisseria meningitidis is required for optimal adhesion to human cells. Cell Microbiol. 2007, 9, 463–478. [Google Scholar] [CrossRef]
- Semmler, A.B.; Whitchurch, C.B.; Leech, A.J.; Mattick, J.S. Identification of a novel gene, fimV, involved in twitching motility in Pseudomonas aeruginosa. Microbiology 2000, 146, 1321–1332. [Google Scholar]
- Tunio, S.A.; Oldfield, N.J.; Berry, A.; Ala’Aldeen, D.A.; Wooldridge, K.G.; Turner, D.P. The moonlighting protein fructose-1, 6-bisphosphate aldolase of Neisseria meningitidis: Surface localization and role in host cell adhesion. Mol. Microbiol. 2010, 76, 605–615. [Google Scholar] [CrossRef]
- Tunio, S.A.; Oldfield, N.J.; Ala’Aldeen, D.A.; Wooldridge, K.G.; Turner, D.P. The role of glyceraldehyde 3-phosphate dehydrogenase (GapA-1) in Neisseria meningitidis adherence to human cells. BMC Microbiol. 2010, 10, 280:1–280:10. [Google Scholar]
- Li, M.S.; Chow, N.Y.; Sinha, S.; Halliwell, D.; Finney, M.; Gorringe, A.R.; Watson, M.W.; Kroll, J.S.; Langford, P.R.; Webb, S.A. A Neisseria meningitidis NMB1966 mutant is impaired for invasion of respiratory epithelial cells, survival in human blood and for virulence in vivo. Med. Microbiol. Immunol. 2009, 198, 57–67. [Google Scholar] [CrossRef]
- Serino, L.; Nesta, B.; Leuzzi, R.; Fontana, M.R.; Monaci, E.; Mocca, B.T.; Cartocci, E.; Masignani, V.; Jerse, A.E.; Rappuoli, R.; et al. Identification of a new OmpA-like protein in Neisseria gonorrhoeae involved in the binding to human epithelial cells and in vivo colonization. Mol. Microbiol. 2007, 64, 1391–1403. [Google Scholar] [CrossRef]
- Grizot, S.; Buchanan, S.K. Structure of the OmpA-like domain of RmpM from Neisseria meningitidis. Mol. Microbiol. 2004, 51, 1027–1037. [Google Scholar] [CrossRef]
- Danoff, E.J.; Fleming, K.G. The soluble, periplasmic domain of OmpA folds as an independent unit and displays chaperone activity by reducing the self-association propensity of the unfolded OmpA transmembrane beta-barrel. Biophys. Chem. 2011, 159, 194–204. [Google Scholar] [CrossRef]
- Hung, M.C.; Heckels, J.E.; Christodoulides, M. The Adhesin Complex Protein (ACP) of Neisseria meningitidis is a new adhesin with vaccine potential. mBio 2013, 4, e00041:1–e00041:13. [Google Scholar]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinf. 2010, 11, 595:1–595:11. [Google Scholar]
- Schoen, C.; Blom, J.; Claus, H.; Schramm-Gluck, A.; Brandt, P.; Muller, T.; Goesmann, A.; Joseph, B.; Konietzny, S.; Kurzai, O.; et al. Whole-genome comparison of disease and carriage strains provides insights into virulence evolution in Neisseria meningitidis. Proc. Natl. Acad. Sci. USA 2008, 105, 3473–3478. [Google Scholar] [CrossRef]
- Swain, C.L.; Martin, D.R. Survival of meningococci outside of the host: Implications for acquisition. Epidemiol. Infect. 2007, 135, 315–320. [Google Scholar] [CrossRef]
- Claus, H.; Maiden, M.C.; Maag, R.; Frosch, M.; Vogel, U. Many carried meningococci lack the genes required for capsule synthesis and transport. Microbiology 2002, 148, 1813–1819. [Google Scholar]
- Frosch, M.; Vogel, U. Structure and Genetics of the Meningococcal Capsule. In Handbook of Meningococcal Disease: Infection Biology, Vaccination, Clinical Management; Frosch, M., Maiden, M.C.J., Eds.; Wiley-VCH: Weinheim, Germany, 2006; Chapter 8; pp. 145–162. [Google Scholar]
- Harrison, O.B.; Claus, H.; Jiang, Y.; Bennett, J.S.; Bratcher, H.B.; Jolley, K.A.; Corton, C.; Care, R.; Poolman, J.T.; Zollinger, W.D.; et al. Description and nomenclature of Neisseria meningitidis capsule locus. Emerg. Infect. Dis. 2013, 19, 566–573. [Google Scholar] [CrossRef]
- Swartley, J.S.; Marfin, A.A.; Edupuganti, S.; Liu, L.J.; Cieslak, P.; Perkins, B.; Wenger, J.D.; Stephens, D.S. Capsule switching of Neisseria meningitidis. Proc. Natl. Acad. Sci. USA 1997, 94, 271–276. [Google Scholar] [CrossRef]
- Finne, J.; Leinonen, M.; Makela, P.H. Antigenic similarities between brain components and bacteria causing meningitis. Implications for vaccine development and pathogenesis. Lancet 1983, 2, 355–357. [Google Scholar] [CrossRef]
- Finne, J.; Finne, U.; Deagostini-Bazin, H.; Goridis, C. Occurrence of alpha 2–8 linked polysialosyl units in a neural cell adhesion molecule. Biochem. Biophys. Res. Commun. 1983, 112, 482–487. [Google Scholar]
- Yamasaki, R.; Bacon, B. Three-dimensional structural analysis of the group B polysaccharide of Neisseria meningitidis 6275 by two-dimensional NMR: The polysaccharide is suggested to exist in helical conformations in solution. Biochemistry 1991, 30, 851–857. [Google Scholar] [CrossRef]
- Lemercinier, X.; Jones, C. Full 1H NMR assignment and detailed O-acetylation patterns of capsular polysaccharides from Neisseria meningitidis used in vaccine production. Carbohydr. Res. 1996, 296, 83–96. [Google Scholar] [CrossRef]
- Gudlavalleti, S.K.; Szymanski, C.M.; Jarrell, H.C.; Stephens, D.S. In vivo determination of Neisseria meningitidis serogroup A capsular polysaccharide by whole cell high-resolution magic angle spinning NMR spectroscopy. Carbohydr. Res. 2006, 341, 557–562. [Google Scholar] [CrossRef]
- Xie, O.; Bolgiano, B.; Gao, F.; Lockyer, K.; Swann, C.; Jones, C.; Delrieu, I.; Njanpop-Lafourcade, B.M.; Tamekloe, T.A.; Pollard, A.J.; et al. Characterization of size, structure and purity of serogroup X Neisseria meningitidis polysaccharide, and development of an assay for quantification of human antibodies. Vaccine 2012, 30, 5812–5823. [Google Scholar] [CrossRef]
- Garrido, R.; Puyada, A.; Fernandez, A.; Gonzalez, M.; Ramirez, U.; Cardoso, F.; Valdes, Y.; Gonzalez, D.; Fernandez, V.; Verez, V.; et al. Quantitative proton nuclear magnetic resonance evaluation and total assignment of the capsular polysaccharide Neisseria meningitidis serogroup X. J. Pharm. Biomed. Anal. 2012, 70, 295–300. [Google Scholar] [CrossRef]
- Kahler, C.M.; Stephens, D.S. Genetic basis for biosynthesis, structure, and function of meningococcal lipooligosaccharide (endotoxin). Crit. Rev. Microbiol. 1998, 24, 281–334. [Google Scholar]
- Arking, D.; Tong, Y.; Stein, D.C. Analysis of lipooligosaccharide biosynthesis in the Neisseriaceae. J. Bacteriol. 2001, 183, 934–941. [Google Scholar] [CrossRef]
- Zhu, P.; Klutch, M.J.; Bash, M.C.; Tsang, R.S.; Ng, L.K.; Tsai, C.M. Genetic diversity of three lgt loci for biosynthesis of lipooligosaccharide (LOS) in Neisseria species. Microbiology 2002, 148, 1833–1844. [Google Scholar]
- Jennings, M.P.; Srikhanta, Y.N.; Moxon, E.R.; Kramer, M.; Poolman, J.T.; Kuipers, B.; van der Ley, P. The genetic basis of the phase variation repertoire of lipopolysaccharide immunotypes in Neisseria meningitidis. Microbiology 1999, 145, 3013–3021. [Google Scholar]
- Mandrell, R.E.; Griffiss, J.M.; Macher, B.A. Lipooligosaccharides (LOS) of Neisseria gonorrhoeae and Neisseria meningitidis have components that are immunochemically similar to precursors of human blood group antigens. Carbohydrate sequence specificity of the mouse monoclonal antibodies that recognize crossreacting antigens on LOS and human erythrocytes. J. Exp. Med. 1988, 168, 107–126. [Google Scholar] [CrossRef]
- Vogel, U.; Hammerschmidt, S.; Rosch, M. Sialic acids of both the capsule and the sialylated lipooligosaccharide of Neisseria meningitis serogroup B are prerequisites for virulence of meningococci in the infant rat. Med. Microbiol. Immunol. 1996, 185, 81–87. [Google Scholar] [CrossRef]
- Hill, D.J.; Griffiths, N.J.; Borodina, E.; Virji, M. Cellular and molecular biology of Neisseria meningitidis colonization and invasive disease. Clin. Sci. (Lond.) 2010, 118, 547–564. [Google Scholar]
- Takayama, K.; Qureshi, N.; Hyver, K.; Honovich, J.; Cotter, R.J.; Mascagni, P.; Schneider, H. Characterization of a structural series of lipid A obtained from the lipopolysaccharides of Neisseria gonorrhoeae. Combined laser desorption and fast atom bombardment mass spectral analysis of high performance liquid chromatography-purified dimethyl derivatives. J. Biol. Chem. 1986, 261, 10624–10631. [Google Scholar]
- Michon, F.; Beurret, M.; Gamian, A.; Brisson, J.R.; Jennings, H.J. Structure of the L5 lipopolysaccharide core oligosaccharides of Neisseria meningitidis. J. Biol. Chem. 1990, 265, 7243–7247. [Google Scholar]
- Yamasaki, R.; Bacon, B.E.; Nasholds, W.; Schneider, H.; Griffiss, J.M. Structural determination of oligosaccharides derived from lipooligosaccharide of Neisseria gonorrhoeae F62 by chemical, enzymatic, and two-dimensional NMR methods. Biochemistry 1991, 30, 10566–10575. [Google Scholar] [CrossRef]
- Gamian, A.; Beurret, M.; Michon, F.; Brisson, J.R.; Jennings, H.J. Structure of the L2 lipopolysaccharide core oligosaccharides of Neisseria meningitidis. J. Biol. Chem. 1992, 267, 922–925. [Google Scholar]
- Kulshin, V.A.; Zahringer, U.; Lindner, B.; Frasch, C.E.; Tsai, C.M.; Dmitriev, B.A.; Rietschel, E.T. Structural characterization of the lipid A component of pathogenic Neisseria meningitidis. J. Bacteriol. 1992, 174, 1793–1800. [Google Scholar]
- Wakarchuk, W.W.; Gilbert, M.; Martin, A.; Wu, Y.; Brisson, J.R.; Thibault, P.; Richards, J.C. Structure of an alpha-2,6-sialylated lipooligosaccharide from Neisseria meningitidis immunotype L1. Eur. J. Biochem. 1998, 254, 626–633. [Google Scholar]
- Cox, A.D.; Li, J.; Brisson, J.R.; Moxon, E.R.; Richards, J.C. Structural analysis of the lipopolysaccharide from Neisseria meningitidis strain BZ157 galE: Localisation of two phosphoethanolamine residues in the inner core oligosaccharide. Carbohydr. Res. 2002, 337, 1435–1444. [Google Scholar] [CrossRef]
- Leavell, M.D.; Leary, J.A.; Yamasaki, R. Mass spectrometric strategy for the characterization of lipooligosaccharides from Neisseria gonorrhoeae 302 using FTICR. J. Am. Soc. Mass Spectrom. 2002, 13, 571–576. [Google Scholar] [CrossRef]
- Cox, A.D.; Wright, J.C.; Gidney, M.A.; Lacelle, S.; Plested, J.S.; Martin, A.; Moxon, E.R.; Richards, J.C. Identification of a novel inner-core oligosaccharide structure in Neisseria meningitidis lipopolysaccharide. Eur. J. Biochem. 2003, 270, 1759–1766. [Google Scholar] [CrossRef]
- Swanson, K.V.; Griffiss, J.M. Separation and identification of neisserial lipooligosaccharide oligosaccharides using high-performance anion-exchange chromatography with pulsed amperometric detection. Carbohydr. Res. 2006, 341, 388–396. [Google Scholar] [CrossRef]
- Choudhury, B.; Kahler, C.M.; Datta, A.; Stephens, D.S.; Carlson, R.W. The structure of the L9 immunotype lipooligosaccharide from Neisseria meningitidis NMA Z2491. Carbohydr. Res. 2008, 343, 2971–2979. [Google Scholar] [CrossRef]
- Kahler, C.M.; Datta, A.; Tzeng, Y.L.; Carlson, R.W.; Stephens, D.S. Inner core assembly and structure of the lipooligosaccharide of Neisseria meningitidis: Capacity of strain NMB to express all known immunotype epitopes. Glycobiology 2005, 15, 409–419. [Google Scholar]
- Jennings, H.J.; Beurret, M.; Gamian, A.; Michon, F. Structure and immunochemistry of meningococcal lipopolysaccharides. Antonie Van Leeuwenhoek 1987, 53, 519–522. [Google Scholar] [CrossRef]
- Pavliak, V.; Brisson, J.R.; Michon, F.; Uhrin, D.; Jennings, H.J. Structure of the sialylated L3 lipopolysaccharide of Neisseria meningitidis. J. Biol. Chem. 1993, 268, 14146–14152. [Google Scholar]
- Phillips, N.J.; John, C.M.; Reinders, L.G.; Gibson, B.W.; Apicella, M.A.; Griffiss, J.M. Structural models for the cell surface lipooligosaccharides of Neisseria gonorrhoeae and Haemophilus influenzae. Biomed. Environ. Mass Spectrom. 1990, 19, 731–745. [Google Scholar] [CrossRef]
- Kerwood, D.E.; Schneider, H.; Yamasaki, R. Structural analysis of lipooligosaccharide produced by Neisseria gonorrhoeae, strain MS11mk (variant A): A precursor for a gonococcal lipooligosaccharide associated with virulence. Biochemistry 1992, 31, 12760–12768. [Google Scholar] [CrossRef]
- Johnson, K.G.; Perry, M.B.; McDonald, I.J.; Russel, R.R. Cellular and free lipopolysaccharides of some species of Neisseria. Can. J. Microbiol. 1975, 21, 1969–1980. [Google Scholar] [CrossRef]
- Johnson, K.G.; Perry, M.B.; McDonald, I.J. Studies of the cellular and free lipopolysaccharides form Neisseria canis and N. subflava. Can. J. Microbiol. 1976, 22, 189–196. [Google Scholar] [CrossRef]
- Tong, Y.; Reinhold, V.; Reinhold, B.; Brandt, B.; Stein, D.C. Structural and immunochemical characterization of the lipooligosaccharides expressed by Neisseria subflava 44. J. Bacteriol. 2001, 183, 942–950. [Google Scholar] [CrossRef]
- Tommassen, J.; Vermeij, P.; Struyve, M.; Benz, R.; Poolman, J.T. Isolation of Neisseria meningitidis mutants deficient in class 1 (porA) and class 3 (porB) outer membrane proteins. Infect. Immun. 1990, 58, 1355–1359. [Google Scholar]
- Kattner, C.; Zaucha, J.; Jaenecke, F.; Zachariae, U.; Tanabe, M. Identification of a cation transport pathway in Neisseria meningitidis porb. Proteins 2013, 81, 830–840. [Google Scholar] [CrossRef]
- Cannon, J.G.; Buchanan, T.M.; Sparling, P.F. Confirmation of association of protein I serotype of Neisseria gonorrhoeae with ability to cause disseminated infection. Infect. Immun. 1983, 40, 816–819. [Google Scholar]
- Feavers, I.M.; Maiden, M.C. A gonococcal porA pseudogene: Implications for understanding the evolution and pathogenicity of Neisseria gonorrhoeae. Mol. Microbiol. 1998, 30, 647–656. [Google Scholar] [CrossRef]
- Derrick, J.P.; Urwin, R.; Suker, J.; Feavers, I.M.; Maiden, M.C. Structural and evolutionary inference from molecular variation in Neisseria porins. Infect. Immun. 1999, 67, 2406–2413. [Google Scholar]
- Bennett, J.S.; Callaghan, M.J.; Derrick, J.P.; Maiden, M.C. Variation in the Neisseria lactamica porin, and its relationship to meningococcal PorB. Microbiology 2008, 154, 1525–1534. [Google Scholar] [CrossRef]
- Minetti, C.A.; Tai, J.Y.; Blake, M.S.; Pullen, J.K.; Liang, S.M.; Remeta, D.P. Structural and functional characterization of a recombinant PorB class 2 protein from Neisseria meningitidis. Conformational stability and porin activity. J. Biol. Chem. 1997, 272, 10710–10720. [Google Scholar]
- Tanabe, M.; Iverson, T.M. Expression, purification and preliminary X-ray analysis of the Neisseria meningitidis outer membrane protein PorB. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2009, 65, 996–1000. [Google Scholar] [CrossRef]
- Tanabe, M.; Nimigean, C.M.; Iverson, T.M. Structural basis for solute transport, nucleotide regulation, and immunological recognition of Neisseria meningitidis PorB. Proc. Natl. Acad. Sci. USA 2010, 107, 6811–6816. [Google Scholar]
- Zeth, K.; Kozjak-Pavlovic, V.; Faulstich, M.; Fraunholz, M.; Hurwitz, R.; Kepp, O.; Rudel, T. Structure and function of the PorB porin from disseminating Neisseria gonorrhoeae. Biochem. J. 2013, 449, 631–642. [Google Scholar] [CrossRef]
- Derrick, J.P.; Maiden, M.C.; Feavers, I.M. Crystal structure of an Fab fragment in complex with a meningococcal serosubtype antigen and a protein G domain. J. Mol. Biol. 1999, 293, 81–91. [Google Scholar] [CrossRef]
- Tzitzilonis, C.; Prince, S.M.; Collins, R.F.; Achtman, M.; Feavers, I.M.; Maiden, M.C.; Derrick, J.P. Structural variation and immune recognition of the P1.2 subtype meningococcal antigen. Proteins 2006, 62, 947–955. [Google Scholar]
- Bartley, S.N.; Tzeng, Y.L.; Heel, K.; Lee, C.W.; Mowlaboccus, S.; Seemann, T.; Lu, W.; Lin, Y.H.; Ryan, C.S.; Peacock, C.; et al. Attachment and invasion of Neisseria meningitidis to host cells is related to surface hydrophobicity, bacterial cell size and capsule. PLoS One 2013, 8, e55798:1–e55798:15. [Google Scholar]
- Deghmane, A.E.; Giorgini, D.; Larribe, M.; Alonso, J.M.; Taha, M.K. Down-regulation of pili and capsule of Neisseria meningitidis upon contact with epithelial cells is mediated by CrgA regulatory protein. Mol. Microbiol. 2002, 43, 1555–1564. [Google Scholar] [CrossRef]
- Virji, M.; Saunders, J.R.; Sims, G.; Makepeace, K.; Maskell, D.; Ferguson, D.J. Pilus-facilitated adherence of Neisseria meningitidis to human epithelial and endothelial cells: Modulation of adherence phenotype occurs concurrently with changes in primary amino acid sequence and the glycosylation status of pilin. Mol. Microbiol. 1993, 10, 1013–1028. [Google Scholar] [CrossRef]
- Nassif, X.; Pujol, C.; Morand, P.; Eugene, E. Interactions of pathogenic Neisseria with host cells. Is it possible to assemble the puzzle? Mol. Microbiol. 1999, 32, 1124–1132. [Google Scholar] [CrossRef]
- Rudel, T.; Scheurerpflug, I.; Meyer, T.F. Neisseria PilC protein identified as type-4 pilus tip-located adhesin. Nature 1995, 373, 357–359. [Google Scholar] [CrossRef]
- Wolfgang, M.; Lauer, P.; Park, H.S.; Brossay, L.; Hebert, J.; Koomey, M. PilT mutations lead to simultaneous defects in competence for natural transformation and twitching motility in piliated Neisseria gonorrhoeae. Mol. Microbiol. 1998, 29, 321–330. [Google Scholar] [CrossRef]
- Wolfgang, M.; Park, H.S.; Hayes, S.F.; van Putten, J.P.; Koomey, M. Suppression of an absolute defect in type IV pilus biogenesis by loss-of-function mutations in pilT, a twitching motility gene in Neisseria gonorrhoeae. Proc. Natl. Acad. Sci. USA 1998, 95, 14973–14978. [Google Scholar]
- Pujol, C.; Eugene, E.; Marceau, M.; Nassif, X. The meningococcal PilT protein is required for induction of intimate attachment to epithelial cells following pilus-mediated adhesion. Proc. Natl. Acad. Sci. USA 1999, 96, 4017–4022. [Google Scholar] [CrossRef]
- Kallstrom, H.; Liszewski, M.K.; Atkinson, J.P.; Jonsson, A.B. Membrane cofactor protein (MCP or CD46) is a cellular pilus receptor for pathogenic Neisseria. Mol. Microbiol. 1997, 25, 639–647. [Google Scholar] [CrossRef]
- Johansson, L.; Rytkonen, A.; Bergman, P.; Albiger, B.; Kallstrom, H.; Hokfelt, T.; Agerberth, B.; Cattaneo, R.; Jonsson, A.B. CD46 in meningococcal disease. Science 2003, 301, 373–375. [Google Scholar] [CrossRef]
- Tobiason, D.M.; Seifert, H.S. Inverse relationship between pilus-mediated gonococcal adherence and surface expression of the pilus receptor, CD46. Microbiology 2001, 147, 2333–2340. [Google Scholar]
- Kirchner, M.; Heuer, D.; Meyer, T.F. CD46-independent binding of neisserial type IV pili and the major pilus adhesin, PilC, to human epithelial cells. Infect. Immun. 2005, 73, 3072–3082. [Google Scholar] [CrossRef]
- Capecchi, B.; du-Bobie, J.; di Marcello, F.; Ciucchi, L.; Masignani, V.; Taddei, A.; Rappuoli, R.; Pizza, M.; Arico, B. Neisseria meningitidis NadA is a new invasin which promotes bacterial adhesion to and penetration into human epithelial cells. Mol. Microbiol. 2005, 55, 687–698. [Google Scholar]
- Nagele, V.; Heesemann, J.; Schielke, S.; Jimenez-Soto, L.F.; Kurzai, O.; Ackermann, N. Neisseria meningitidis adhesin NadA targets beta1 integrins: Functional similarity to Yersinia invasin. J. Biol. Chem. 2011, 286, 20536–20546. [Google Scholar]
- Knaust, A.; Weber, M.V.; Hammerschmidt, S.; Bergmann, S.; Frosch, M.; Kurzai, O. Cytosolic proteins contribute to surface plasminogen recruitment of Neisseria meningitidis. J. Bacteriol. 2007, 189, 3246–3255. [Google Scholar] [CrossRef]
- Morelle, S.; Carbonnelle, E.; Nassif, X. The REP2 repeats of the genome of Neisseria meningitidis are associated with genes coordinately regulated during bacterial cell interaction. J. Bacteriol. 2003, 185, 2618–2627. [Google Scholar] [CrossRef]
- Ieva, R.; Alaimo, C.; Delany, I.; Spohn, G.; Rappuoli, R.; Scarlato, V. CrgA is an inducible LysR-type regulator of Neisseria meningitidis, acting both as a repressor and as an activator of gene transcription. J. Bacteriol. 2005, 187, 3421–3430. [Google Scholar] [CrossRef]
- Virji, M.; Makepeace, K.; Ferguson, D.J.P.; Achtman, M.; Moxon, E.R. Meningococcal Opa and Opc proteins: Their role in colonization and invasion of human epithelial and endothelial cells. Mol. Microbiol. 1993, 10, 499–510. [Google Scholar] [CrossRef]
- Chen, T.; Belland, R.J.; Wilson, J.; Swanson, J. Adherence of pilus- Opa+ gonococci to epithelial cells in vitro involves heparan sulfate. J. Exp. Med. 1995, 182, 511–517. [Google Scholar] [CrossRef]
- Van Putten, J.P.; Paul, S.M. Binding of syndecan-like cell surface proteoglycan receptors is required for Neisseria gonorrhoeae entry into human mucosal cells. EMBO J. 1995, 14, 2144–2154. [Google Scholar]
- Gomez-Duarte, O.G.; Dehio, M.; Guzman, C.A.; Chhatwal, G.S.; Dehio, C.; Meyer, T.F. Binding of vitronectin to opa-expressing Neisseria gonorrhoeae mediates invasion of HeLa cells. Infect. Immun. 1997, 65, 3857–3866. [Google Scholar]
- Duensing, T.D.; Putten, J.P. Vitronectin binds to the gonococcal adhesin OpaA through a glycosaminoglycan molecular bridge. Biochem. J. 1998, 334, 133–139. [Google Scholar]
- Moore, J.; Bailey, S.E.; Benmechernene, Z.; Tzitzilonis, C.; Griffiths, N.J.; Virji, M.; Derrick, J.P. Recognition of saccharides by the OpcA, OpaD, and OpaB outer membrane proteins from Neisseria meningitidis. J. Biol. Chem. 2005, 280, 31489–31497. [Google Scholar] [CrossRef]
- Gray-Owen, S.D.; Blumberg, R.S. CEACAM1: Contact-dependent control of immunity. Nat. Rev. Immunol. 2006, 6, 433–446. [Google Scholar] [CrossRef]
- Gold, P.; Freedman, S.O. Demonstration of tumor-specific antigens in human colonic carcinomata by immunological tolerance and absorption techniques. J. Exp. Med. 1965, 121, 439–462. [Google Scholar] [CrossRef]
- Chen, T.; Gotschlich, E.C. CGM1a antigen of neutrophils, a receptor of gonococcal opacity proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 14851–14856. [Google Scholar] [CrossRef]
- Virji, M.; Makepeace, K.; Ferguson, D.J.; Watt, S.M. Carcinoembryonic antigens (CD66) on epithelial cells and neutrophils are receptors for Opa proteins of pathogenic Neisseriae. Mol. Microbiol. 1996, 22, 941–950. [Google Scholar]
- Virji, M.; Watt, S.M.; Barker, S.; Makepeace, K.; Doyonnas, R. The N-domain of the human CD66a adhesion molecule is a target for Opa proteins of Neisseria meningitidis and Neisseria gonorrhoeae. Mol. Microbiol. 1996, 22, 929–939. [Google Scholar]
- Bos, M.P.; Grunert, F.; Belland, R.J. Differential recognition of members of the carcinoembryonic antigen family by Opa variants of Neisseria gonorrhoeae. Infect. Immun. 1997, 65, 2353–2361. [Google Scholar]
- Chen, T.; Grunert, F.; Medina-Marino, A.; Gotschlich, E.C. Several carcinoembryonic antigens (CD66) serve as receptors for gonococcal opacity proteins. J. Exp. Med. 1997, 185, 1557–1564. [Google Scholar] [CrossRef]
- Gray-Owen, S.D.; Dehio, C.; Haude, A.; Grunert, F.; Meyer, T.F. CD66 carcinoembryonic antigens mediate interactions between Opa-expressing Neisseria gonorrhoeae and human polymorphonuclear phagocytes. EMBO J. 1997, 16, 3435–3445. [Google Scholar] [CrossRef]
- Gray-Owen, S.D.; Lorenzen, D.R.; Haude, A.; Meyer, T.F.; Dehio, C. Differential Opa specificities for CD66 receptors influence tissue interactions and cellular response to Neisseria gonorrhoeae. Mol. Microbiol. 1997, 26, 971–980. [Google Scholar] [CrossRef]
- Popp, A.; Dehio, C.; Grunert, F.; Meyer, T.F.; Gray-Owen, S.D. Molecular analysis of neisserial Opa protein interactions with the CEA family of receptors: Identification of determinants contributing to the differential specificities of binding. Cell Microbiol. 1999, 1, 169–181. [Google Scholar] [CrossRef]
- Muenzner, P.; Dehio, C.; Fujiwara, T.; Achtman, M.; Meyer, T.F.; Gray-Owen, S.D. Carcinoembryonic antigen family receptor specificity of Neisseria meningitidis Opa variants influences adherence to and invasion of proinflammatory cytokine-activated endothelial cells. Infect. Immun. 2000, 68, 3601–3607. [Google Scholar] [CrossRef]
- Rowe, H.A.; Griffiths, N.J.; Hill, D.J.; Virji, M. Co-ordinate action of bacterial adhesins and human carcinoembryonic antigen receptors in enhanced cellular invasion by capsulate serum resistant Neisseria meningitidis. Cell Microbiol. 2007, 9, 154–168. [Google Scholar] [CrossRef]
- Griffiths, N.J.; Bradley, C.J.; Heyderman, R.S.; Virji, M. IFN-gamma amplifies NFkappaB-dependent Neisseria meningitidis invasion of epithelial cells via specific upregulation of CEA-related cell adhesion molecule 1. Cell Microbiol. 2007, 9, 2968–2983. [Google Scholar] [CrossRef]
- Slevogt, H.; Zabel, S.; Opitz, B.; Hocke, A.; Eitel, J.; N’guessan, P.D.; Lucka, L.; Riesbeck, K.; Zimmermann, W.; Zweigner, J.; et al. CEACAM1 inhibits Toll-like receptor 2-triggered antibacterial responses of human pulmonary epithelial cells. Nat. Immunol. 2008, 9, 1270–1278. [Google Scholar] [CrossRef]
- Bos, M.P.; Kuroki, M.; Krop-Watorek, A.; Hogan, D.; Belland, R.J. CD66 receptor specificity exhibited by neisserial Opa variants is controlled by protein determinants in CD66 N-domains. Proc. Natl. Acad. Sci. USA 1998, 95, 9584–9589. [Google Scholar] [CrossRef]
- Virji, M.; Evans, D.; Hadfield, A.; Grunert, F.; Teixeira, A.M.; Watt, S.M. Critical determinants of host receptor targeting by Neisseria meningitidis and Neisseria gonorrhoeae: Identification of Opa adhesiotopes on the N-domain of CD66 molecules. Mol. Microbiol. 1999, 34, 538–551. [Google Scholar] [CrossRef]
- Bos, M.P.; Kao, D.; Hogan, D.M.; Grant, C.C.; Belland, R.J. Carcinoembryonic antigen family receptor recognition by gonococcal Opa proteins requires distinct combinations of hypervariable Opa protein domains. Infect. Immun. 2002, 70, 1715–1723. [Google Scholar] [CrossRef]
- De Jonge, M.I.; Hamstra, H.J.; van Alphen, L.; Dankert, J.; van der Ley, P. Mapping the binding domains on meningococcal Opa proteins for CEACAM1 and CEA receptors. Mol. Microbiol. 2003, 50, 1005–1015. [Google Scholar] [CrossRef]
- Virji, M.; Makepeace, K.; Ferguson, D.J.; Achtman, M.; Sarkari, J.; Moxon, E.R. Expression of the Opc protein correlates with invasion of epithelial and endothelial cells by Neisseria meningitidis. Mol. Microbiol. 1992, 6, 2785–2795. [Google Scholar] [CrossRef]
- De Vries, F.P.; Cole, R.; Dankert, J.; Frosch, M.; van Putten, J.P. Neisseria meningitidis producing the Opc adhesin binds epithelial cell proteoglycan receptors. Mol. Microbiol. 1998, 27, 1203–1212. [Google Scholar] [CrossRef]
- Virji, M.; Makepeace, K.; Moxon, R. Distinct mechanisms of interactions of Opc-expressing meningococci at apical and basolateral surfaces of human endothelial cells; the role of integrins in apical interactions. Mol. Microbiol. 1994, 14, 173–184. [Google Scholar] [CrossRef]
- Unkmeir, A.; Latsch, K.; Dietrich, G.; Wintermeyer, E.; Schinke, B.; Schwender, S.; Kim, K.S.; Eigenthaler, M.; Frosch, M. Fibronectin mediates Opc-dependent internalization of Neisseria meningitidis in human brain microvascular endothelial cells. Mol. Microbiol. 2002, 46, 933–946. [Google Scholar] [CrossRef]
- Sa Cunha, C.E.; Griffiths, N.J.; Murillo, I.; Virji, M. Neisseria meningitidis Opc invasin binds to the cytoskeletal protein alpha-actinin. Cell Microbiol. 2009, 11, 389–405. [Google Scholar] [CrossRef]
- De Vries, F.P.; van der Ende, A.; van Putten, J.P.; Dankert, J. Invasion of primary nasopharyngeal epithelial cells by Neisseria meningitidis is controlled by phase variation of multiple surface antigens. Infect. Immun. 1996, 64, 2998–3006. [Google Scholar]
- Merz, A.J.; Enns, C.A.; So, M. Type IV pili of pathogenic Neisseriae elicit cortical plaque formation in epithelial cells. Mol. Microbiol. 1999, 32, 1316–1332. [Google Scholar] [CrossRef]
- Chamot-Rooke, J.; Mikaty, G.; Malosse, C.; Soyer, M.; Dumont, A.; Gault, J.; Imhaus, A.F.; Martin, P.; Trellet, M.; Clary, G.; et al. Posttranslational modification of pili upon cell contact triggers N. meningitidis dissemination. Science 2011, 331, 778–782. [Google Scholar] [CrossRef]
- Ayala, B.P.; Vasquez, B.; Clary, S.; Tainer, J.A.; Rodland, K.; So, M. The pilus-induced Ca2+ flux triggers lysosome exocytosis and increases the amount of Lamp1 accessible to Neisseria IgA1 protease. Cell Microbiol. 2001, 3, 265–275. [Google Scholar] [CrossRef]
- Ayala, P.; Vasquez, B.; Wetzler, L.; So, M. Neisseria gonorrhoeae porin P1.B induces endosome exocytosis and a redistribution of Lamp1 to the plasma membrane. Infect. Immun. 2002, 70, 5965–5971. [Google Scholar] [CrossRef]
- Greenfield, S.; Sheehe, P.R.; Feldman, H.A. Meningococcal carriage in a population of “normal” families. J. Infect. Dis. 1971, 123, 67–73. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Apicella, M.A.; Shao, J.; Neil, R.B. Methods for Studying Neisseria meningitidis Biofilms. In Neisseria meningitidis: Advanced Methods and Protocols; Christodoulides, M., Ed.; Humana Press: New York, NY, USA, 2012; Chapter 11; pp. 169–184. [Google Scholar]
- Neil, R.B.; Apicella, M.A. Clinical and laboratory evidence for Neisseria meningitidis biofilms. Future. Microbiol. 2009, 4, 555–563. [Google Scholar] [CrossRef]
- Yi, K.; Rasmussen, A.W.; Gudlavalleti, S.K.; Stephens, D.S.; Stojiljkovic, I. Biofilm formation by Neisseria meningitidis. Infect. Immun. 2004, 72, 6132–6138. [Google Scholar] [CrossRef]
- Lappann, M.; Haagensen, J.A.; Claus, H.; Vogel, U.; Molin, S. Meningococcal biofilm formation: Structure, development and phenotypes in a standardized continuous flow system. Mol. Microbiol. 2006, 62, 1292–1309. [Google Scholar] [CrossRef]
- Neil, R.B.; Shao, J.Q.; Apicella, M.A. Biofilm formation on human airway epithelia by encapsulated Neisseria meningitidis serogroup B. Microbes. Infect. 2009, 11, 281–287. [Google Scholar] [CrossRef]
- Lappann, M.; Vogel, U. Biofilm formation by the human pathogen Neisseria meningitidis. Med. Microbiol. Immunol. 2010, 199, 173–183. [Google Scholar] [CrossRef]
- Neil, R.B.; Apicella, M.A. Role of HrpA in biofilm formation of Neisseria meningitidis and regulation of the hrpBAS transcripts. Infect. Immun. 2009, 77, 2285–2293. [Google Scholar] [CrossRef]
- Lappann, M.; Claus, H.; van Alen, T.; Harmsen, M.; Elias, J.; Molin, S.; Vogel, U. A dual role of extracellular DNA during biofilm formation of Neisseria meningitidis. Mol. Microbiol. 2010, 75, 1355–1371. [Google Scholar] [CrossRef]
- Edwards, J.L.; Apicella, M.A. I-domain-containing integrins serve as pilus receptors for Neisseria gonorrhoeae adherence to human epithelial cells. Cell Microbiol. 2005, 7, 1197–1211. [Google Scholar] [CrossRef]
- Harvey, H.A.; Swords, W.E.; Apicella, M.A. The mimicry of human glycolipids and glycosphingolipids by the lipooligosaccharides of pathogenic Neisseria and Haemophilus. J. Autoimmun. 2001, 16, 257–262. [Google Scholar] [CrossRef]
- James-Holmquest, A.N.; Swanson, J.; Buchanan, T.M.; Wende, R.D.; Williams, R.P. Differential attachment by piliated and nonpiliated Neisseria gonorrhoeae to human sperm. Infect. Immun. 1974, 9, 897–902. [Google Scholar]
- Harvey, H.A.; Porat, N.; Campbell, C.A.; Jennings, M.; Gibson, B.W.; Phillips, N.J.; Apicella, M.A.; Blake, M.S. Gonococcal lipooligosaccharide is a ligand for the asialoglycoprotein receptor on human sperm. Mol. Microbiol. 2000, 36, 1059–1070. [Google Scholar] [CrossRef]
- Liu, J.H.; Li, H.Y.; Cao, Z.G.; Duan, Y.F.; Li, Y.; Ye, Z.Q. Influence of several uropathogenic microorganisms on human sperm motility parameters in vitro. Asian J. Androl. 2002, 4, 179–182. [Google Scholar]
- Edwards, J.L.; Butler, E.K. The pathobiology of Neisseria gonorrhoeae lower female genital tract infection. Front. Microbiol. 2011, 2, 102:1–102:12. [Google Scholar]
- Jarvis, G.A. Analysis of C3 deposition and degradation on Neisseria meningitidis and Neisseria gonorrhoeae. Infect. Immun. 1994, 62, 1755–1760. [Google Scholar]
- Edwards, J.L.; Brown, E.J.; Ault, K.A.; Apicella, M.A. The role of complement receptor 3 (CR3) in Neisseria gonorrhoeae infection of human cervical epithelia. Cell Microbiol. 2001, 3, 611–622. [Google Scholar] [CrossRef]
- Edwards, J.L.; Brown, E.J.; Uk-Nham, S.; Cannon, J.G.; Blake, M.S.; Apicella, M.A. A co-operative interaction between Neisseria gonorrhoeae and complement receptor 3 mediates infection of primary cervical epithelial cells. Cell Microbiol. 2002, 4, 571–584. [Google Scholar] [CrossRef]
- Jennings, M.P.; Jen, F.E.; Roddam, L.F.; Apicella, M.A.; Edwards, J.L. Neisseria gonorrhoeae pilin glycan contributes to CR3 activation during challenge of primary cervical epithelial cells. Cell Microbiol. 2011, 13, 885–896. [Google Scholar] [CrossRef]
- Van Putten, J.P.; Duensing, T.D.; Carlson, J. Gonococcal invasion of epithelial cells driven by P.IA, a bacterial ion channel with GTP binding properties. J. Exp. Med. 1998, 188, 941–952. [Google Scholar] [CrossRef]
- Agarwal, S.; Ram, S.; Ngampasutadol, J.; Gulati, S.; Zipfel, P.F.; Rice, P.A. Factor H facilitates adherence of Neisseria gonorrhoeae to complement receptor 3 on eukaryotic cells. J. Immunol. 2010, 185, 4344–4353. [Google Scholar] [CrossRef]
- Harvey, H.A.; Jennings, M.P.; Campbell, C.A.; Williams, R.; Apicella, M.A. Receptor-mediated endocytosis of Neisseria gonorrhoeae into primary human urethral epithelial cells: The role of the asialoglycoprotein receptor. Mol. Microbiol. 2001, 42, 659–672. [Google Scholar]
- Swanson, K.V.; Jarvis, G.A.; Brooks, G.F.; Barham, B.J.; Cooper, M.D.; Griffiss, J.M. CEACAM is not necessary for Neisseria gonorrhoeae to adhere to and invade female genital epithelial cells. Cell Microbiol. 2001, 3, 681–691. [Google Scholar] [CrossRef]
- Minor, S.Y.; Banerjee, A.; Gotschlich, E.C. Effect of alpha-oligosaccharide phenotype of Neisseria gonorrhoeae strain MS11 on invasion of Chang conjunctival, HEC-1-B endometrial, and ME-180 cervical cells. Infect. Immun. 2000, 68, 6526–6534. [Google Scholar] [CrossRef]
- Bessen, D.; Gotschlich, E.C. Interactions of gonococci with HeLa cells: Attachment, detachment, replication, penetration, and the role of protein II. Infect. Immun. 1986, 54, 154–160. [Google Scholar]
- Grassme, H.; Gulbins, E.; Brenner, B.; Ferlinz, K.; Sandhoff, K.; Harzer, K.; Lang, F.; Meyer, T.F. Acidic sphingomyelinase mediates entry of N. gonorrhoeae into nonphagocytic cells. Cell 1997, 91, 605–615. [Google Scholar] [CrossRef]
- Van Putten, J.P.; Duensing, T.D.; Cole, R.L. Entry of OpaA+ gonococci into HEp-2 cells requires concerted action of glycosaminoglycans, fibronectin and integrin receptors. Mol. Microbiol. 1998, 29, 369–379. [Google Scholar] [CrossRef]
- Greiner, L.L.; Edwards, J.L.; Shao, J.; Rabinak, C.; Entz, D.; Apicella, M.A. Biofilm Formation by Neisseria gonorrhoeae. Infect. Immun. 2005, 73, 1964–1970. [Google Scholar] [CrossRef]
- Pruthi, V.; Al-Janabi, A.; Pereira, B.M. Characterization of biofilm formed on intrauterine devices. Indian J. Med. Microbiol. 2003, 21, 161–165. [Google Scholar]
- Steichen, C.T.; Shao, J.Q.; Ketterer, M.R.; Apicella, M.A. Gonococcal cervicitis: A role for biofilm in pathogenesis. J. Infect. Dis. 2008, 198, 1856–1861. [Google Scholar] [CrossRef]
- Falsetta, M.L.; Steichen, C.T.; McEwan, A.G.; Cho, C.; Ketterer, M.; Shao, J.; Hunt, J.; Jennings, M.P.; Apicella, M.A. The composition and metabolic phenotype of Neisseria gonorrhoeae biofilms. Front. Microbiol. 2011, 2, 75:1–75:11. [Google Scholar]
- Shaw, J.H.; Falkow, S. Model for invasion of human tissue culture cells by Neisseria gonorrhoeae. Infect. Immun. 1988, 56, 1625–1632. [Google Scholar]
- Christodoulides, M.; Everson, J.S.; Liu, B.L.; Lambden, P.R.; Watt, P.J.; Thomas, E.J.; Heckels, J.E. Interaction of primary human endometrial cells with Neisseria gonorrhoeae expressing green fluorescent protein. Mol. Microbiol. 2000, 35, 32–43. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Arvidson, C.G. Inhibition of Neisseria gonorrhoeae epithelial cell interactions by vaginal Lactobacillus species. Infect. Immun. 2008, 76, 3124–3130. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Arvidson, C.G. Lactobacillus jensenii surface-associated proteins inhibit Neisseria gonorrhoeae adherence to epithelial cells. Infect. Immun. 2010, 78, 3103–3111. [Google Scholar] [CrossRef]
- Griffiss, J.M.; Lammel, C.J.; Wang, J.; Dekker, N.P.; Brooks, G.F. Neisseria gonorrhoeae coordinately uses Pili and Opa to activate HEC-1-B cell microvilli, which causes engulfment of the gonococci. Infect. Immun. 1999, 67, 3469–3480. [Google Scholar]
- Timmerman, M.M.; Shao, J.Q.; Apicella, M.A. Ultrastructural analysis of the pathogenesis of Neisseria gonorrhoeae endometrial infection. Cell Microbiol. 2005, 7, 627–636. [Google Scholar] [CrossRef]
- McCormack, W.M. Pelvic inflammatory disease. N. Engl. J. Med. 1994, 330, 115–119. [Google Scholar] [CrossRef]
- Ward, M.E.; Watt, P.J.; Robertson, J.N. The human fallopian tube: A laboratory model for gonococcal infection. J. Infect. Dis. 1974, 129, 650–659. [Google Scholar] [CrossRef]
- McGee, Z.A.; Stephens, D.S.; Hoffman, L.H.; Schlech, W.F., III; Horn, R.G. Mechanisms of mucosal invasion by pathogenic Neisseria. Rev. Infect. Dis. 1983, 5, S708–S714. [Google Scholar] [CrossRef]
- Gorby, G.L.; Schaefer, G.B. Effect of attachment factors (pili plus Opa) on Neisseria gonorrhoeae invasion of human fallopian tube tissue in vitro: Quantitation by computerized image analysis. Microb. Pathog. 1992, 13, 93–108. [Google Scholar] [CrossRef]
- Velasquez, L.; Garcia, K.; Morales, F.; Heckels, J.E.; Orihuela, P.; Rodas, P.I.; Christodoulides, M.; Cardenas, H. Neisseria gonorrhoeae pilus attenuates cytokine response of human fallopian tube explants. J. Biomed. Biotechnol. 2012, 2012, 491298: 1–491298:7. [Google Scholar]
- Dekker, N.P.; Lammel, C.J.; Mandrell, R.E.; Brooks, G.F. Opa (protein II) influences gonococcal organization in colonies, surface appearance, size and attachment to human fallopian tube tissues. Microb. Pathog. 1990, 9, 19–31. [Google Scholar] [CrossRef]
- Quan, D.N.; Cooper, M.D.; Potter, J.L.; Roberts, M.H.; Cheng, H.; Jarvis, G.A. TREM-2 binds to lipooligosaccharides of Neisseria gonorrhoeae and is expressed on reproductive tract epithelial cells. Mucosal. Immunol. 2008, 1, 229–238. [Google Scholar] [CrossRef]
- Muenzner, P.; Bachmann, V.; Zimmermann, W.; Hentschel, J.; Hauck, C.R. Human-restricted bacterial pathogens block shedding of epithelial cells by stimulating integrin activation. Science 2010, 329, 1197–1201. [Google Scholar] [CrossRef]
- Morales, P.; Reyes, P.; Vargas, M.; Rios, M.; Imarai, M.; Cardenas, H.; Croxatto, H.; Orihuela, P.; Vargas, R.; Fuhrer, J.; et al. Infection of human fallopian tube epithelial cells with Neisseria gonorrhoeae protects cells from tumor necrosis factor alpha-induced apoptosis. Infect. Immun. 2006, 74, 3643–3650. [Google Scholar] [CrossRef]
- Britigan, B.E.; Cohen, M.S.; Sparling, P.F. Gonococcal infection: A model of molecular pathogenesis. N. Engl. J. Med. 1985, 312, 1683–1694. [Google Scholar] [CrossRef]
- Morello, J.A.; Bohnhoff, M. Serovars and serum resistance of Neisseria gonorrhoeae from disseminated and uncomplicated infections. J. Infect. Dis. 1989, 160, 1012–1017. [Google Scholar] [CrossRef]
- Rechner, C.; Kuhlewein, C.; Muller, A.; Schild, H.; Rudel, T. Host glycoprotein Gp96 and scavenger receptor SREC interact with PorB of disseminating Neisseria gonorrhoeae in an epithelial invasion pathway. Cell Host. Microbe 2007, 2, 393–403. [Google Scholar] [CrossRef]
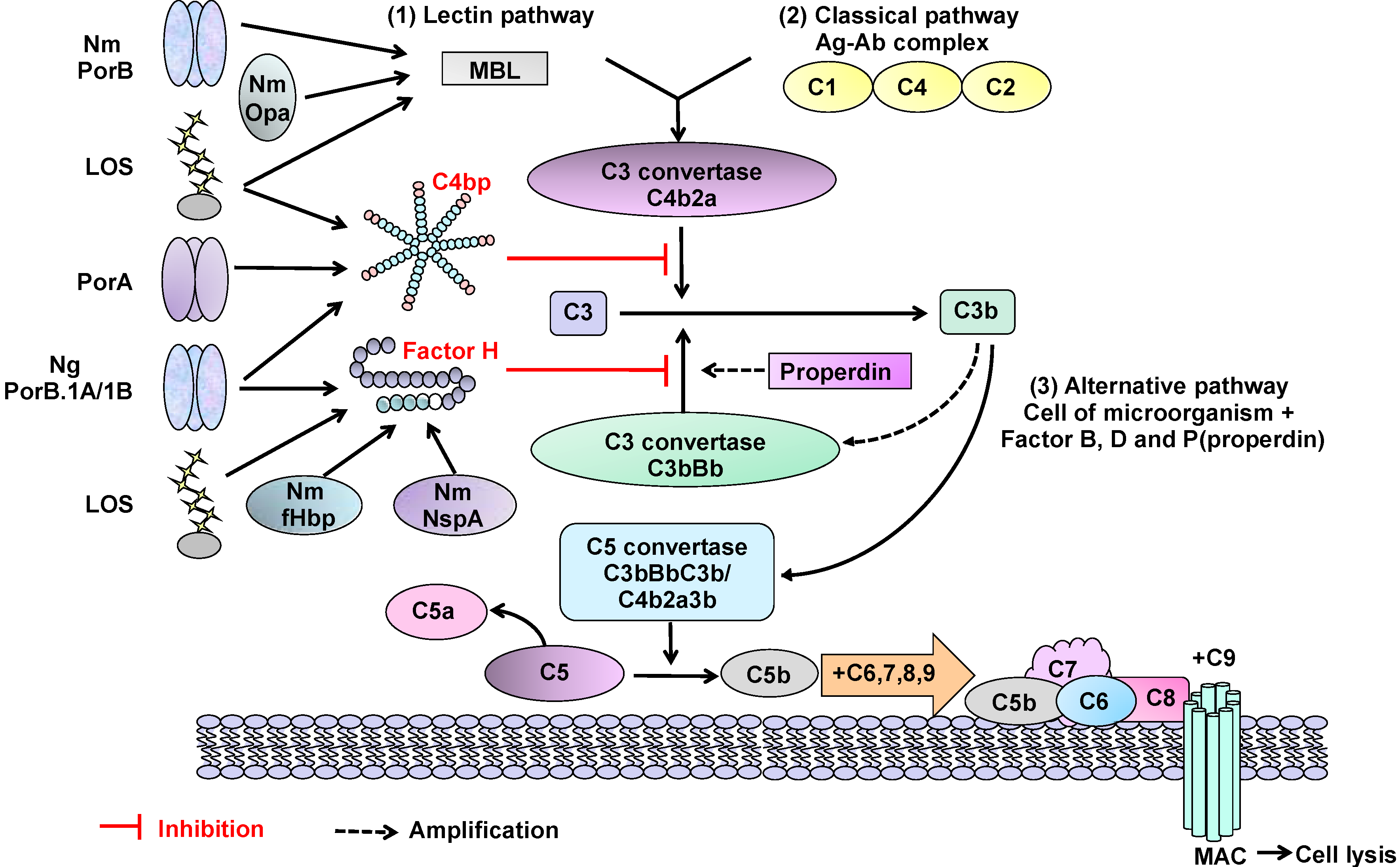
- Van Deuren, M.; Brandtzaeg, P.; van der Meer, J. Update on meningococcal disease with emphasis on pathogenesis and clinical management. Clin. Microbiol. Rev. 2000, 13, 144–166. [Google Scholar] [CrossRef]
- Schneider, M.C.; Exley, R.M.; Ram, S.; Sim, R.B.; Tang, C.M. Interactions between Neisseria meningitidis and the complement system. Trends Microbiol. 2007, 15, 233–240. [Google Scholar] [CrossRef]
- Kugelberg, E.; Gollan, B.; Tang, C.M. Mechanisms in Neisseria meningitidis for resistance against complement-mediated killing. Vaccine 2008, 26, I34–I39. [Google Scholar]
- Stephens, D.S.; Edwards, K.M.; Morris, F.M.G. Pili and outer membrane appendages on Neisseria meningitidis in the cerebrospinal fluid of an infant. J. Infect. Dis. 1982, 146, 568. [Google Scholar] [CrossRef]
- Devoe, I.W. The meningococcus and mechanisms of pathogenicity. Microbiol. Rev. 1982, 46, 162–190. [Google Scholar]
- Brandtzaeg, P.; van Deuren, M. Current concepts in the role of the host response in Neisseria meningitidis septic shock. Curr. Opin. Infect. Dis. 2002, 15, 247–252. [Google Scholar] [CrossRef]
- Jarva, H.; Ram, S.; Vogel, U.; Blom, A.M.; Meri, S. Binding of the complement inhibitor C4bp to serogroup B Neisseria meningitidis. J. Immunol. 2005, 174, 6299–6307. [Google Scholar]
- Madico, G.; Welsch, J.A.; Lewis, L.A.; McNaughton, A.; Perlman, D.H.; Costello, C.E.; Ngampasutadol, J.; Vogel, U.; Granoff, D.M.; Ram, S. The meningococcal vaccine candidate GNA1870 binds the complement regulatory protein factor H and enhances serum resistance. J. Immunol. 2006, 177, 501–510. [Google Scholar]
- Lewis, L.A.; Ngampasutadol, J.; Wallace, R.; Reid, J.E.; Vogel, U.; Ram, S. The meningococcal vaccine candidate neisserial surface protein A (NspA) binds to factor H and enhances meningococcal resistance to complement. PLoS Pathog 2010, 6, e1001027:1–e1001027:20. [Google Scholar]
- Ram, S.; McQuillen, D.P.; Gulati, S.; Elkins, C.; Pangburn, M.K.; Rice, P.A. Binding of complement factor H to loop 5 of porin protein 1A: A molecular mechanism of serum resistance of nonsialylated Neisseria gonorrhoeae. J. Exp. Med. 1998, 188, 671–680. [Google Scholar]
- Ram, S.; Sharma, A.K.; Simpson, S.D.; Gulati, S.; McQuillen, D.P.; Pangburn, M.K.; Rice, P.A. A novel sialic acid binding site on factor H mediates serum resistance of sialylated Neisseria gonorrhoeae. J. Exp. Med. 1998, 187, 743–752. [Google Scholar] [CrossRef]
- Ram, S.; Cullinane, M.; Blom, A.M.; Gulati, S.; McQuillen, D.P.; Monks, B.G.; O’Connell, C.; Boden, R.; Elkins, C.; Pangburn, M.K.; et al. Binding of C4b-binding protein to porin: A molecular mechanism of serum resistance of Neisseria gonorrhoeae. J. Exp. Med. 2001, 193, 281–295. [Google Scholar] [CrossRef]
- Sjolinder, H.; Eriksson, J.; Maudsdotter, L.; Aro, H.; Jonsson, A.B. Meningococcal outer membrane protein NhhA is essential for colonization and disease by preventing phagocytosis and complement attack. Infect. Immun. 2008, 76, 5412–5420. [Google Scholar] [CrossRef]
- Estabrook, M.M.; Jack, D.L.; Klein, N.J.; Jarvis, G.A. Mannose-binding lectin binds to two major outer membrane proteins, opacity protein and porin, of Neisseria meningitidis. J. Immunol. 2004, 172, 3784–3792. [Google Scholar]
- Devyatyarova-Johnson, M.; Rees, I.H.; Robertson, B.D.; Turner, M.W.; Klein, N.J.; Jack, D.L. The lipopolysaccharide structures of Salmonella enterica serovar Typhimurium and Neisseria gonorrhoeae determine the attachment of human mannose-binding lectin to intact organisms. Infect. Immun. 2000, 68, 3894–3899. [Google Scholar]
- Melican, K.; Michea, V.P.; Martin, T.; Bruneval, P.; Dumenil, G. Adhesion of Neisseria meningitidis to dermal vessels leads to local vascular damage and purpura in a humanized mouse model. PLoS Pathog. 2013, 9, e1003139:1–e1003139:11. [Google Scholar]
- Sa E Cunha, C.; Griffiths, N.J.; Virji, M. Neisseria meningitidis Opc invasin binds to the sulphated tyrosines of activated vitronectin to attach to and invade human brain endothelial cells. PLoS Pathog. 2010, 6, e1000911:1–e1000911:18. [Google Scholar]
- Muenzner, P.; Naumann, M.; Meyer, T.F.; Gray-Owen, S.D. Pathogenic Neisseria trigger expression of their carcinoembryonic antigen-related cellular adhesion molecule 1 (CEACAM1; previously CD66a) receptor on primary endothelial cells by activating the immediate early response transcription factor, nuclear factor-kappaB. J. Biol. Chem. 2001, 276, 24331–24340. [Google Scholar]
- Soyer, M.; Dumenil, G. A laminar-Flow Chamber Assay for Measuring Bacterial Adhesion under Shear Stress. In Neisseria meningitidis: Advanced Methods and Protocols; Christodoulides, M., Ed.; Humana Press: New York, NY, USA, 2012; Chapter 12; pp. 185–195. [Google Scholar]
- Mikaty, G.; Soyer, M.; Mairey, E.; Henry, N.; Dyer, D.; Forest, K.T.; Morand, P.; Guadagnini, S.; Prevost, M.C.; Nassif, X.; et al. Extracellular bacterial pathogen induces host cell surface reorganization to resist shear stress. PLoS Pathog. 2009, 5, e1000314:1–e1000314:14. [Google Scholar]
- Coureuil, M.; Lecuyer, H.; Scott, M.G.; Boularan, C.; Enslen, H.; Soyer, M.; Mikaty, G.; Bourdoulous, S.; Nassif, X.; Marullo, S. Meningococcus Hijacks a beta2-adrenoceptor/beta-Arrestin pathway to cross brain microvasculature endothelium. Cell 2010, 143, 1149–1160. [Google Scholar]
- Lecuyer, H.; Nassif, X.; Coureuil, M. Two strikingly different signaling pathways are induced by meningococcal type IV pili on endothelial and epithelial cells. Infect. Immun. 2012, 80, 175–186. [Google Scholar] [CrossRef]
- Brissac, T.; Mikaty, G.; Dumenil, G.; Coureuil, M.; Nassif, X. The meningococcal minor pilin PilX is responsible for type IV pilus conformational changes associated with signaling to endothelial cells. Infect. Immun. 2012, 80, 3297–3306. [Google Scholar] [CrossRef]
- Virji, M.; Kayhty, H.; Ferguson, D.J.; Alexandrescu, C.; Heckels, J.E.; Moxon, E.R. The role of pili in the interactions of pathogenic Neisseria with cultured human endothelial cells. Mol. Microbiol. 1991, 5, 1831–1841. [Google Scholar] [CrossRef]
- Scheuerpflug, I.; Rudel, T.; Ryll, R.; Pandit, J.; Meyer, T.F. Roles of PilC and PilE proteins in pilus-mediated adherence of Neisseria gonorrhoeae and Neisseria meningitidis to human erythrocytes and endothelial and epithelial cells. Infect. Immun. 1999, 67, 834–843. [Google Scholar]
- Kupsch, E.M.; Knepper, B.; Kuroki, T.; Heuer, I.; Meyer, T.F. Variable opacity (Opa) outer membrane proteins account for the cell tropisms displayed by Neisseria gonorrhoeae for human leukocytes and epithelial cells. EMBO J. 1993, 12, 641–650. [Google Scholar]
- McNeil, G.; Virji, M. Phenotypic variants of meningococci and their potential in phagocytic interactions: The influence of opacity proteins, pili, PilC and surface sialic acids. Microb. Pathog. 1997, 22, 295–304. [Google Scholar] [CrossRef]
- Swanson, J.; Sparks, E.; Young, D.; King, G. Young, D.; King, G. Studies on Gonococcus infection. X. Pili and leukocyte association factor as mediators of interactions between gonococci and eukaryotic cells in vitro. Infect. Immun. 1975, 11, 1352–1361. [Google Scholar]
- Soderholm, N.; Vielfort, K.; Hultenby, K.; Aro, H. Pathogenic Neisseria hitchhike on the uropod of human neutrophils. PLoS One 2011, 6, e24353:1–e24353:12. [Google Scholar]
- Johnson, M.B.; Criss, A.K. Resistance of Neisseria gonorrhoeae to neutrophils. Front. Microbiol. 2011, 2, 77:1–77:12. [Google Scholar]
- Sadarangani, M.; Pollard, A.J.; Gray-Owen, S.D. Opa proteins and CEACAMs: Pathways of immune engagement for pathogenic Neisseria. FEMS Microbiol. Rev. 2011, 35, 498–514. [Google Scholar] [CrossRef]
- Criss, A.K.; Seifert, H.S. A bacterial siren song: Intimate interactions between Neisseria and neutrophils. Nat. Rev. Microbiol. 2012, 10, 178–190. [Google Scholar] [CrossRef]
- Wiertz, E.J.; Delvig, A.; Donders, E.M.; Brugghe, H.F.; van Unen, L.M.; Timmermans, H.A.; Achtman, M.; Hoogerhout, P.; Poolman, J.T. T-cell responses to outer membrane proteins of Neisseria meningitidis: Comparative study of the Opa, Opc, and PorA proteins. Infect. Immun. 1996, 64, 298–304. [Google Scholar]
- Lee, H.S.; Boulton, I.C.; Reddin, K.; Wong, H.; Halliwell, D.; Mandelboim, O.; Gorringe, A.R.; Gray-Owen, S.D. Neisserial outer membrane vesicles bind the coinhibitory receptor carcinoembryonic antigen-related cellular adhesion molecule 1 and suppress CD4+ T lymphocyte function. Infect. Immun. 2007, 75, 4449–4455. [Google Scholar]
- Boulton, I.C.; Gray-Owen, S.D. Neisserial binding to CEACAM1 arrests the activation and proliferation of CD4+ T lymphocytes. Nat. Immunol. 2002, 3, 229–236. [Google Scholar] [CrossRef]
- Mandrell, R.E.; Zollinger, W.D. Human immune response to meningococcal outer membrane protein epitopes after natural infection or vaccination. Infect. Immun. 1989, 57, 1590–1598. [Google Scholar]
- Pantelic, M.; Kim, Y.J.; Bolland, S.; Chen, I.; Shively, J.; Chen, T. Neisseria gonorrhoeae kills carcinoembryonic antigen-related cellular adhesion molecule 1 (CD66a)-expressing human B cells and inhibits antibody production. Infect. Immun. 2005, 73, 4171–4179. [Google Scholar]
- Al-Bader, T.; Jolley, K.A.; Humphries, H.E.; Holloway, J.; Heckels, J.E.; Semper, A.E.; Friedmann, P.S.; Christodoulides, M. Activation of human dendritic cells by the PorA protein of Neisseria meningitidis. Cell Microbiol. 2004, 6, 651–662. [Google Scholar] [CrossRef]
- Franzoso, S.; Mazzon, C.; Sztukowska, M.; Cecchini, P.; Kasic, T.; Capecchi, B.; Tavano, R.; Papini, E. Human monocytes/macrophages are a target of Neisseria meningitidis Adhesin A (NadA). J. Leukoc. Biol. 2008, 83, 1100–1110. [Google Scholar] [CrossRef]
- Sjolinder, M.; Altenbacher, G.; Hagner, M.; Sun, W.; Schedin-Weiss, S.; Sjolinder, H. Meningococcal outer membrane protein NhhA triggers apoptosis in macrophages. PLoS One 2012, 7, e29586:1–e29586:8. [Google Scholar]
- Quattroni, P.; Li, Y.; Lucchesi, D.; Lucas, S.; Hood, D.W.; Herrmann, M.; Gabius, H.J.; Tang, C.M.; Exley, R.M. Galectin-3 binds Neisseria meningitidis and increases interaction with phagocytic cells. Cell Microbiol. 2012, 14, 1657–1675. [Google Scholar]
- Jones, C.; Virji, M.; Crocker, P.R. Recognition of sialylated meningococcal lipopolysaccharide by siglecs expressed on myeloid cells leads to enhanced bacterial uptake. Mol. Microbiol. 2003, 49, 1213–1225. [Google Scholar] [CrossRef]
- Peiser, L.; Makepeace, K.; Pluddemann, A.; Savino, S.; Wright, J.C.; Pizza, M.; Rappuoli, R.; Moxon, E.R.; Gordon, S. Identification of Neisseria meningitidis nonlipopolysaccharide ligands for class A macrophage scavenger receptor by using a novel assay. Infect. Immun. 2006, 74, 5191–5199. [Google Scholar] [CrossRef]
- Casey, R.; Newcombe, J.; McFadden, J.; Bodman-Smith, K.B. The acute-phase reactant C-reactive protein binds to phosphorylcholine-expressing Neisseria meningitidis and increases uptake by human phagocytes. Infect. Immun. 2008, 76, 1298–1304. [Google Scholar]
- Zughaier, S.M.; Tzeng, Y.L.; Zimmer, S.M.; Datta, A.; Carlson, R.W.; Stephens, D.S. Neisseria meningitidis lipooligosaccharide structure-dependent activation of the macrophage CD14/Toll-like receptor 4 pathway. Infect. Immun. 2004, 72, 371–380. [Google Scholar] [CrossRef]
- Zimmer, S.M.; Zughaier, S.M.; Tzeng, Y.L.; Stephens, D.S. Human MD-2 discrimination of meningococcal lipid A structures and activation of TLR4. Glycobiology 2007, 17, 847–856. [Google Scholar] [CrossRef]
- DeMarco, M.L.; Woods, R.J. From agonist to antagonist: Structure and dynamics of innate immune glycoprotein MD-2 upon recognition of variably acylated bacterial endotoxins. Mol. Immunol. 2011, 49, 124–133. [Google Scholar] [CrossRef]
- Lu, R.; Pan, H.; Shively, J.E. CEACAM1 negatively regulates IL-1beta production in LPS activated neutrophils by recruiting SHP-1 to a SYK-TLR4-CEACAM1 complex. PLoS Pathog. 2012, 8, e1002597:1–e1002597:15. [Google Scholar]
- Massari, P.; Visintin, A.; Gunawardana, J.; Halmen, K.A.; King, C.A.; Golenbock, D.T.; Wetzler, L.M. Meningococcal porin PorB binds to TLR2 and requires TLR1 for signaling. J. Immunol. 2006, 176, 2373–2380. [Google Scholar]
- Wetzler, L.M. Innate immune function of the neisserial porins and the relationship to vaccine adjuvant activity. Fut. Microbiol. 2010, 5, 749–758. [Google Scholar] [CrossRef]
- Macleod, H.; Bhasin, N.; Wetzler, L.M. Role of protein tyrosine kinase and Erk1/2 activities in the Toll-like receptor 2-induced cellular activation of murine B cells by neisserial porin. Clin. Vaccine Immunol. 2008, 15, 630–637. [Google Scholar] [CrossRef]
- Singleton, T.E.; Massari, P.; Wetzler, L.M. Neisserial porin-induced dendritic cell activation is MyD88 and TLR2 dependent. J. Immunol. 2005, 174, 3545–3550. [Google Scholar]
- Sjolinder, M.; Altenbacher, G.; Wang, X.; Gao, Y.; Hansson, C.; Sjolinder, H. The meningococcal adhesin NhhA provokes proinflammatory responses in macrophages via toll-like receptor 4-dependent and -independent pathways. Infect. Immun. 2012, 80, 4027–4033. [Google Scholar]
- Chu, C.L.; Yu, Y.L.; Kung, Y.C.; Liao, P.Y.; Liu, K.J.; Tseng, Y.T.; Lin, Y.C.; Hsieh, S.S.; Chong, P.C.; Yang, C.Y. The immunomodulatory activity of meningococcal lipoprotein Ag473 depends on the conformation made up of the lipid and protein moieties. PLoS One 2012, 7, e40873:1–e40873:11. [Google Scholar]
- Hill, M.; Deghmane, A.E.; Segovia, M.; Zarantonelli, M.L.; Tilly, G.; Blancou, P.; Beriou, G.; Josien, R.; Anegon, I.; Hong, E.; et al. Penicillin binding proteins as danger signals: Meningococcal penicillin binding protein 2 activates dendritic cells through Toll-like receptor 4. PLoS One 2011, 6, e23995:1–e23995:11. [Google Scholar]
- Cecchini, P.; Tavano, R.; Polverino de, L.P.; Franzoso, S.; Mazzon, C.; Montanari, P.; Papini, E. The soluble recombinant Neisseria meningitidis adhesin NadA(Delta351–405) stimulates human monocytes by binding to extracellular Hsp90. PLoS One 2011, 6, e25089:1–e25089:13. [Google Scholar]
- Zughaier, S.M. Neisseria meningitidis capsular polysaccharides induce inflammatory responses via TLR2 and TLR4-MD-2. J. Leukoc. Biol. 2011, 89, 469–480. [Google Scholar] [CrossRef]
- Ingalls, R.R.; Lien, E.; Golenbock, D.T. Membrane-associated proteins of a lipopolysaccharide-deficient mutant of Neisseria meningitidis activate the inflammatory response through toll-like receptor 2. Infect. Immun. 2001, 69, 2230–2236. [Google Scholar] [CrossRef]
- Pridmore, A.C.; Wyllie, D.H.; Abdillahi, F.; Steeghs, L.; van der Ley, P.; Dower, S.K.; Read, R.C. A lipopolysaccharide-deficient mutant of Neisseria meningitidis elicits attenuated cytokine release by human macrophages and signals via toll-like receptor (TLR) 2 but not via TLR4/MD2. J. Infect. Dis. 2001, 183, 89–96. [Google Scholar] [CrossRef]
- Fisette, P.L.; Ram, S.; Andersen, J.M.; Guo, W.; Ingalls, R.R. The Lip lipoprotein from Neisseria gonorrhoeae stimulates cytokine release and NF-kappaB activation in epithelial cells in a Toll-like receptor 2-dependent manner. J. Biol. Chem. 2003, 278, 46252–46260. [Google Scholar]
- Liu, X.; Wetzler, L.M.; Nascimento, L.O.; Massari, P. Human airway epithelial cell responses to Neisseria lactamica and purified porin via Toll-like receptor 2-dependent signaling. Infect. Immun. 2010, 78, 5314–5323. [Google Scholar] [CrossRef]
- Toussi, D.N.; Carraway, M.; Wetzler, L.M.; Lewis, L.A.; Liu, X.; Massari, P. The amino acid sequence of Neisseria lactamica PorB surface-exposed loops influences Toll-like receptor 2-dependent cell activation. Infect. Immun. 2012, 80, 3417–3428. [Google Scholar] [CrossRef]
- Pron, B.; Taha, M.K.; Rambaud, C.; Fournet, J.C.; Pattey, N.; Monnet, J.P.; Musilek, M.; Beretti, J.L.; Nassif, X. Interaction of Neisseria maningitidis with the components of the blood-brain barrier correlates with an increased expression of PilC. J. Infect. Dis. 1997, 176, 1285–1292. [Google Scholar]
- Sjolinder, H.; Jonsson, A.B. Olfactory nerve—A novel invasion route of Neisseria meningitidis to reach the meninges. PLoS One 2010, 5, e14034:1–e14034:10. [Google Scholar]
- Christodoulides, M.; Heckels, J.E.; Weller, R.O. The Role of the Leptomeninges in Meningococcal Meningitis. In Emerging Strategies in the Fight against Meningitis; Ferreiros, C., Criado, M.T., Vazquez, J., Eds.; Horizon Press: Norfolk, UK, 2002; pp. 1–37. [Google Scholar]
- Mairey, E.; Genovesio, A.; Donnadieu, E.; Bernard, C.; Jaubert, F.; Pinard, E.; Seylaz, J.; Olivo-Marin, J.C.; Nassif, X.; Dumenil, G. Cerebral microcirculation shear stress levels determine Neisseria meningitidis attachment sites along the blood-brain barrier. J. Exp. Med. 2006, 203, 1939–1950. [Google Scholar] [CrossRef]
- Orihuela, C.J.; Mahdavi, J.; Thornton, J.; Mann, B.; Wooldridge, K.G.; Abouseada, N.; Oldfield, N.J.; Self, T.; Ala’Aldeen, D.A.; Tuomanen, E.I. Laminin receptor initiates bacterial contact with the blood brain barrier in experimental meningitis models. J. Clin. Invest. 2009, 119, 1638–1646. [Google Scholar] [CrossRef]
- Feurer, D.J.; Weller, R.O. Barrier functions of the leptomeninges: A study of normal meninges and meningiomas in tissue culture. Neuropathol. Appl. Neurobiol. 1991, 17, 391–405. [Google Scholar] [CrossRef]
- Alcolado, R.; Weller, R.O.; Parrish, E.P.; Garrod, D. The cranial arachnoid and pia mater in man: Anatomical and ultrastructural observations. Neuropathol. Appl. Neurobiol. 1988, 14, 1–17. [Google Scholar] [CrossRef]
- Kleihues, P.; Cavenee, W.K. Pathology and Genetics of Tumours of the Nervous System; IARC press: Lyon, France, 1997. [Google Scholar]
- Hardy, S.J.; Christodoulides, M.; Weller, R.O.; Heckels, J.E. Interactions of Neisseria meningitidis with cells of the human meninges. Mol. Microbiol. 2000, 36, 817–829. [Google Scholar] [CrossRef]
- Fowler, M.I.; Yin, K.Y.; Humphries, H.E.; Heckels, J.E.; Christodoulides, M. Comparison of the inflammatory responses of human meningeal cells following challenge withNeisseria lactamica and with Neisseria meningitidis. Infect. Immun. 2006, 74, 6467–6478. [Google Scholar] [CrossRef]
- Lauer, B.A.; Fisher, C.E. Neisseria lactamica meningitis. Am. J. Dis. Child. 1976, 130, 198–199. [Google Scholar]
- Del Rio, C.; Stephens, D.S.; Knapp, J.S.; Rice, R.J.; Schalla, W.O. Comparison of isolates of Neisseria gonorrhoeae causing meningitis and report of gonococcal meningitis in a patient with C8 deficiency. J. Clin. Microbiol. 1989, 27, 1045–1049. [Google Scholar]
- Virji, M.; Alexandrescu, C.; Ferguson, D.J.; Saunders, J.R.; Moxon, E.R. Variations in the expression of pili: The effect on adherence of Neisseria meningitidis to human epithelial and endothelial cells. Mol. Microbiol. 1992, 6, 1271–1279. [Google Scholar] [CrossRef]
- Nassif, X.; Lowy, J.; Stenberg, P.; O’Gaora, P.; Ganji, A.; So, M. Antigenic variation of pilin regulates adhesion of Neisseria meningitidis to human epithelial cells. Mol. Microbiol. 1993, 8, 719–725. [Google Scholar] [CrossRef]
- Virji, M.; Stimson, E.; Makepeace, K.; Dell, A.; Morris, H.R.; Payne, G.; Saunders, J.R.; Moxon, E.R. Posttranslational modifications of meningococcal pili. Identification of a common trisaccharide substitution on variant pilins of strain C311. Ann. N. Y. Acad. Sci. 1996, 797, 53–64. [Google Scholar]
- Weiser, J.N.; Goldberg, J.B.; Pan, N.; Wilson, L.; Virji, M. The phosphorylcholine epitope undergoes phase variation on a 43-kilodalton protein in Pseudomonas aeruginosa and on pili of Neisseria meningitidis and Neisseria gonorrhoeae. Infect. Immun. 1998, 66, 4263–4267. [Google Scholar]
- Marceau, M.; Beretti, J.L.; Nassif, X. High adhesiveness of encapsulated Neisseria meningitidis to epithelial cells is associated with the formation of bundles of pili. Mol. Microbiol. 1995, 17, 855–863. [Google Scholar]
- Humphries, H.E.; Triantafilou, M.; Makepeace, B.L.; Heckels, J.E.; Triantafilou, K.; Christodoulides, M. Activation of human meningeal cells is modulated by lipopolysaccharide (LPS) and non-LPS components of Neisseria meningitidis and is independent of Toll-like receptor (TLR)4 and TLR2 signalling. Cell Microbiol. 2005, 7, 415–430. [Google Scholar] [CrossRef]
- Dietrich, G.; Kurz, S.; Hubner, C.; Aepinus, C.; Theiss, S.; Guckenberger, M.; Panzner, U.; Weber, J.; Frosch, M. Transcriptome analysis of Neisseria meningitidis during infection. J. Bacteriol. 2003, 185, 155–164. [Google Scholar] [CrossRef]
- Schubert-Unkmeir, A.; Sokolova, O.; Panzner, U.; Eigenthaler, M.; Frosch, M. Gene expression pattern in human brain endothelial cells in response to Neisseria meningitidis. Infect. Immun. 2007, 75, 899–914. [Google Scholar] [CrossRef]
- Schubert-Unkmeir, A.; Slanina, H.; Frosch, M. Mammalian cell transcriptome in response to meningitis-causing pathogens. Expert Rev. Mol. Diagn. 2009, 9, 833–842. [Google Scholar] [CrossRef]
- Echenique-Rivera, H.; Muzzi, A.; Del Tordello, E.; Seib, K.L.; Francois, P.; Rappuoli, R.; Pizza, M.; Serruto, D. Transcriptome analysis of Neisseria meningitidis in human whole blood and mutagenesis studies identify virulence factors involved in blood survival. PLoS Pathog. 2011, 7, e10002027:1–e10002027:18. [Google Scholar]
- Del, T.E.; Serruto, D. Functional genomics studies of the human pathogen Neisseria meningitidis. Brief. Funct. Genomics 2013. [Google Scholar] [CrossRef]
- Merz, A.J.; So, M. Interactions of pathogenic Neisseriae with epithelial cell membranes. Annu. Rev. Cell. Dev. Biol. 2000, 16, 423–457. [Google Scholar]
- Pujol, C.; Eugene, E.; Morand, P.; Nassif, X. Do pathogenic Neisseriae need several ways to modify the host cell cytoskeleton? Microbes Infect. 2000, 2, 821–827. [Google Scholar] [CrossRef]
- Virji, M. Pathogenic Neisseriae: Surface modulation, pathogenesis and infection control. Nat. Rev. Microbiol. 2009, 7, 274–286. [Google Scholar] [CrossRef]
- Kline, K.A.; Falker, S.; Dahlberg, S.; Normark, S.; Henriques-Normark, B. Bacterial adhesins in host-microbe interactions. Cell Host Microbe 2009, 5, 580–592. [Google Scholar]
- Hill, D.J.; Virji, M. Meningococcal ligands and molecular targets of the host. In Neisseria meningitidis: Advanced Methods and Protocols; Christodoulides, M., Ed.; Humana Press: New York, NY, USA, 2012; Chapter 9; pp. 143–152. [Google Scholar]
- Miller, F.; Lecuyer, H.; Join-Lambert, O.; Bourdoulous, S.; Marullo, S.; Nassif, X.; Coureuil, M. Neisseria meningitidis colonization of the brain endothelium and cerebrospinal fluid invasion. Cell Microbiol. 2013, 15, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Serruto, D.; Bottomley, M.J.; Ram, S.; Giuliani, M.M.; Rappuoli, R. The new multicomponent vaccine against meningococcal serogroup B, 4CMenB: Immunological, functional and structural characterization of the antigens. Vaccine 2012, 30, B87–B97. [Google Scholar] [CrossRef]
- Mcneil, L.K.; Zagursky, R.J.; Lin, S.L.; Murphy, E.; Zlotnick, G.W.; Hoiseth, S.K.; Jansen, K.U.; Anderson, A.S. Role of Factor H binding protein in Neisseria meningitidis virulence and its potential as a vaccine candidate to broadly protect against meningococcal disease. Microbiol. Mol. Biol. Rev. 2013, 77, 234–252. [Google Scholar]
- Marri, P.R.; Paniscus, M.; Weyand, N.J.; Rendon, M.A.; Calton, C.M.; Hernandez, D.R.; Higashi, D.L.; Sodergren, E.; Weinstock, G.M.; Rounsley, S.D.; et al. Genome sequencing reveals widespread virulence gene exchange among human Neisseria species. PLoS One 2010, 5, e11835:1–e11835:9. [Google Scholar]
- Deasy, A.; Dale, A.; Evans, C.; Guccione, E.; Andrews, N.; Gorringe, A.; Read, R. Induced Nasopharyngeal Colonisation with Neisseria lactamica Protects against Carriage of Neisseria meningitidis in Healthy Adult Volunteers. In Proceedings of the XVIIIth International Pathogenic Neisseria Conference (IPNC), Wurzberg, Germany, 9–14th September 2012; p. 100.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hung, M.-C.; Christodoulides, M. The Biology of Neisseria Adhesins. Biology 2013, 2, 1054-1109. https://doi.org/10.3390/biology2031054
Hung M-C, Christodoulides M. The Biology of Neisseria Adhesins. Biology. 2013; 2(3):1054-1109. https://doi.org/10.3390/biology2031054
Chicago/Turabian StyleHung, Miao-Chiu, and Myron Christodoulides. 2013. "The Biology of Neisseria Adhesins" Biology 2, no. 3: 1054-1109. https://doi.org/10.3390/biology2031054
APA StyleHung, M. -C., & Christodoulides, M. (2013). The Biology of Neisseria Adhesins. Biology, 2(3), 1054-1109. https://doi.org/10.3390/biology2031054