3.1. Natural Product Library Screen
An assay for chaperone-mediated protein renaturation was developed in 1994 using rabbit reticulocyte lysate (RRL) to refold thermally denatured firefly luciferase [
34]. RRL had been used for decades for
in vitro protein synthesis and was known to contain abundant quantities of the heat shock proteins. After the discovery that geldanamycin was an Hsp90 inhibitor, it was also demonstrated to inhibit the refolding of luciferase in RRL, confirming that luciferase renaturation was an Hsp90-dependent process [
28]. The assay was demonstrated to be sensitive to compounds that bound and inhibited Hsp70, indicating that the assay could also detect compounds that interacted with other critical components of the Hsp90 chaperone machinery [
27]. Subsequently, it was demonstrated that inhibitors that bind to the C-terminal domain of Hsp90 (e.g., novobiocin) also inhibited luciferase refolding in RRL [
29]. The assay was then miniaturized and developed as a high-throughput screen for inhibitors of the Hsp90 chaperone machine [
29]. Thus, while we generically refer to the compounds discussed below as Hsp90 inhibitors, the molecular target of the potential inhibitor could be any component of the Hsp90 chaperone machinery that is required for the refolding of luciferase.
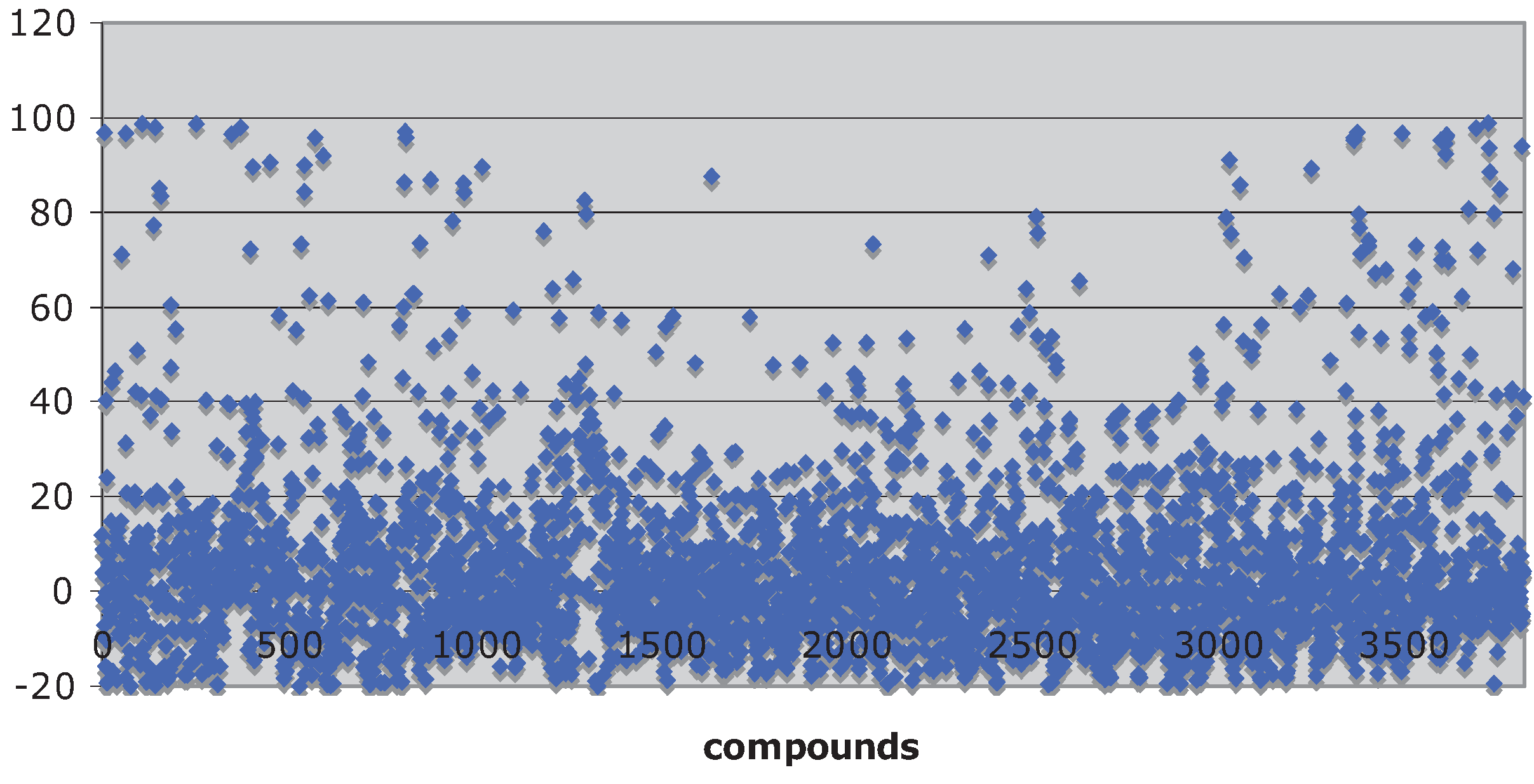
Natural product libraries proved to be a fertile ground for identifying inhibitors of the Hsp90 chaperone machine. A scatter plot of percent inhibition of luciferase refolding
versus 3,859 compounds screened from the TimTec, Biofocus, BioMol and Analyticon libraries is shown in
Figure 1 (median = −0.041 ± 13 rSD). The Microsource compounds were part of a previously published screen [
29] that had a Z-factor of 0.62 ± 0.09. The Z-factors for the TimTec and Analyticon screens were 0.77 ± 0.18 and 0.64 ± 0.14, respectively. The %rCV for the screens of the TimTec, Biofocus, BioMol and Analyticon screens were 8.9 ± 3.9, 20 ± 6.4, 18 ± 5.0 and 13 ± 6.2, respectively, indicating that the assay had a good signal-to-noise ration.
Figure 1.
Scatter plot showing the activities of 3,859 of the compounds screened.
Figure 1.
Scatter plot showing the activities of 3,859 of the compounds screened.
The compounds, whose structures are shown in
Figure 2,
Figure 3,
Figure 4,
Figure 5,
Figure 6,
Figure 7,
Figure 8,
Figure 9,
Figure 10 and
Figure 11 and are listed in
Table 1,
Table 2,
Table 3,
Table 4,
Table 5,
Table 6,
Table 7,
Table 8,
Table 9, and
Table 10, inhibited Hsp90-dependent refolding of luciferase by 60% or greater in the initial screen (a value greater than 4 rSD from the median), without inhibiting luciferase itself, with the exception of the screening of the Microsource compounds which used a cut-off of 70% inhibition [
29]. The compounds have been sorted largely by structural classification, although some do not fit well into any particular structural group. Some compounds were present in more than one library. Curcumin (2 screens), gambogic acid (3 screens) and plumbagin (3 screens) were consistently identified as hits. Of the compounds that were present in more than one library, but were not identified as hits, six were present in the Microsource screen that used a cut-off of 70% inhibition, and four compounds failed to be identified as hits because they inhibited luciferase refolding less than the 60% cut-off used for the other libraries. The lone exception was luteolin, which exhibited no inhibitory activity in one of the screens, indicating that its identification as a hit should be viewed with some skepticism.
Because of Hsp90’s role in modulating proteins associated with signal transduction and protein shuttling, its inhibition has widespread and sometimes varied effects within a cell. Depending on the conditions, the physiological manifestations of this inhibition can also vary. Thus, Hsp90 inhibitors display not only anti-proliferative and anti-tumor properties, but they can also display anti-inflammatory, anti-metastatic [
20,
21,
35], immunosuppressant, anti-viral [
22], anti-bacterial, anti-fungal [
23] and/or anti-malarial [
24,
25] properties depending on the system investigated. Another property of inhibitors that bind to Hsp90’s N-terminal ATP-binding domain is the induction of a robust heat shock response [
10].
The significance of this screen, is that novel compounds not known to have any activity against Hsp90 or its co-chaperones can be implicated to exhibit multiple, seemingly unrelated, medically relevant biological activities. Subsequent to the screen, the literature was mined to identify reports on the physiological effect of potential compounds. As shown below, some of the compounds identified have been specifically shown to inhibit the activities of proteins known to be dependent upon Hsp90 for their function [
9,
10,
20]. These proteins include Akt, STAT-3, Her2 (ErbB2), Insulin-like Growth Factor Receptor (IGFR), Endothelial Growth Factor Receptor (EGFR), telomerase and others. Compounds reported to block the actions of these proteins, or their downstream signaling partners, such as NF-κB, are of special interest. Since Hsp90 is required for the activity of viral polymerases [
22,
36,
37], anti-viral activity is another hallmark manifested by Hsp90 inhibitors [
22]. VEGFR1 and 2 [
38], and HIF1α [
35] are also Hsp90-dependent clients, and as such Hsp90 inhibitors are accordingly anti-angiogenic [
39]. Even though they have not been identified as Hsp90 inhibitors, many of the compounds identified from this screen belong to structural families that contain known Hsp90 inhibitors. Below are examples of the biological activities manifested that make each class of compounds, or specific compounds, potential candidates as Hsp90 inhibitors.
3.1.1. Sesquiterpene Lactones, Tetracyclic Sesterpenoids and Sesterpines
Some of the compound hits in this screen belong to the sesquiterpene lactone family of compounds (
Figure 2,
Table 1). These molecules are characterized by a fifteen-carbon skeleton formed by the union of three isoprene units that contain a lactone group. These compounds are present in many types of plants, and have long been used for various purposes in traditional medicine. Given their effectiveness in the treatment of a wide variety of ailments, and their observed activity on multiple cellular functions and molecular targets, these compounds represent promising candidates as Hsp90 inhibitors.
Figure 2.
Sesquiterpine lactones, tetracyclic sestorpenoids and sesterpines.
Figure 2.
Sesquiterpine lactones, tetracyclic sestorpenoids and sesterpines.
Sesquiterpene lactones have been grouped into seven general classes according to their structures. They are germacranolides, eudesmanolides, eremophilanolides, guaianolides, pseudoguaianolides, hypocretenolides, and iso-seco-tanapartholides. Compounds with reported biological activity come from all of the groups, although the germacranolides, guaianolides, and pseudoguaianolides appear most prominent [
40].
Several sesquiterpene lactones with structures similar to those shown in
Figure 1 have demonstrated biological activities. Isodeoxyelephantopin, and its nearly identical analog, deoxyelephantopin, were shown to inhibit the proliferation of mouse fibroblast tumor cells. The two compounds also inhibited DNA replication in both proliferating lymphocytes and tumor ascites [
41]. Another pair of compounds fitting into this family, costunolide and eremanthin that have structures similar to compounds
6 and
7, were extracted from the ornamental plant
Costus speciosus, and displayed anti-fungal activity similar to the standard anti-fungal, fluconazole, against two species of
Trichophyton, and somewhat weaker activity against other fungi [
42]. As nitric oxide is a mediator of inflammation, a compound’s effect on nitric oxide production is also an indicator of its anti-inflammatory potential, which is especially relevant to this study, as nitric oxide synthase is a well-known Hsp90 client. Eudesmanolides isolated from
Taraxacum mongolicum inhibit nitric oxide production in RAW 264.7 mouse macrophages [
43].
Compound
6, parthenolide, is from a different class of sesquiterpene lactones, but presents a similar structure. Additionally, it contains a methane group at the same location as the previously mentioned compound, a functional group that is the sole distinguishing feature from another, less active compound in the same study. Compound
4, an eudesmanolide (MEGxm0_000041), contains a similar moiety at its core, but it also contains an unsaturated eight-carbon fatty acid ester. Two compounds from the plant
Eupatorium lindleyanum, eupalinolide A and eupalinolide B, are of the germacranolide sub-class. They induced expression of several heat shock proteins, including Hsp70 and Hsp90, in mouse squamous cell carcinoma and melanoma cells. The compounds were also shown to activate HSF1 [
44], a potential indicator of Hsp90 inhibition.
Compound
6, parthenolide, has been identified as an anti-tumor and anti-inflammatory agent, and is currently in clinical trials along with several other sesquiterpene lactones for acute myeloid leukemia, acute lymphoblastic leukemia, and other types of blood and lymph node cancers [
40]. Parthenolide’s anti-cancer and anti-inflammatory activities have been attributed to multiple mechanisms. It was shown to inhibit the activation of NF-κB by IKK, even when the kinase was constitutively active [
45]. It was also able to sensitize TRAIL-resistant cancer cells by inhibiting STAT3 activation [
46].
Compound
9, helenalin, suppresses NF-κB activation, promotes ROS generation, and induces apoptosis by bypassing Bcl-2 function [
47]. Helenalin is cytotoxic against a number of cancer cells, and it also manifests immunosuppressant and anti-inflammatory activity [
48]. In addition, helenalin is a potent inhibitor of telomerase [
49], which further supports its potential as an Hsp90 inhibitor. Compound
3, also belongs to this class of compounds, and presents a similarity in the attached moieties.
Grouped with these compounds are two additional hits. Luffariellolide, a sesterterpene from a marine sponge, is cytotoxic to breast cancer cells, and inhibits the activation of the Hsp90-dependent protein HIF-1α [
50]. Compound
11, 12-epi-scalardial, a tetracyclic sesterpenoid, inhibits EGFR-mediated activation of Akt [
51].
Table 1.
Sesquiterpine lactones, tetracyclic sesterpenoids and sesterterpenes.
Table 1.
Sesquiterpine lactones, tetracyclic sesterpenoids and sesterterpenes.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 1 | Guaianolide | ~60 | |
| 2 | Germacranolide | ~40 | |
| 3 | Guaianolide | ~40 | |
| 4 | Eudesmanolide: MEGxm0_000041 | ~60 | Cytotoxic to L5178Y lymphoblastic, PC12 neuroendrocrine, HeLa cervical cancer cells [52] |
| 5 | Tagitinin C | ~10 | Anti-malaria [53] |
| 6 | Parthenolide | >100 | Anti-tumor, anti-inflammatory [40,45,46] |
| 7 | 17-C4 | ~30 | |
| 8 | 16-H2 | ~10 | |
| 9 | Helenalin | ~80 | Inhibition of NF-κB and suppression of Bcl-2-mediated resistance to apoptosis [47]; anti-leukemic [54]; inhibition of telomerase [49]; induction of autophagy and cell cycle arrest [55] |
| 10 | Luffariellolide | ~10–20 | Cytotoxic, inhibits HIF-1α [50] |
| 11 | Scalaradial, 12-epi- | ~40–60 | Inhibits EGFR activation of Akt independent of PLA2 [51] |
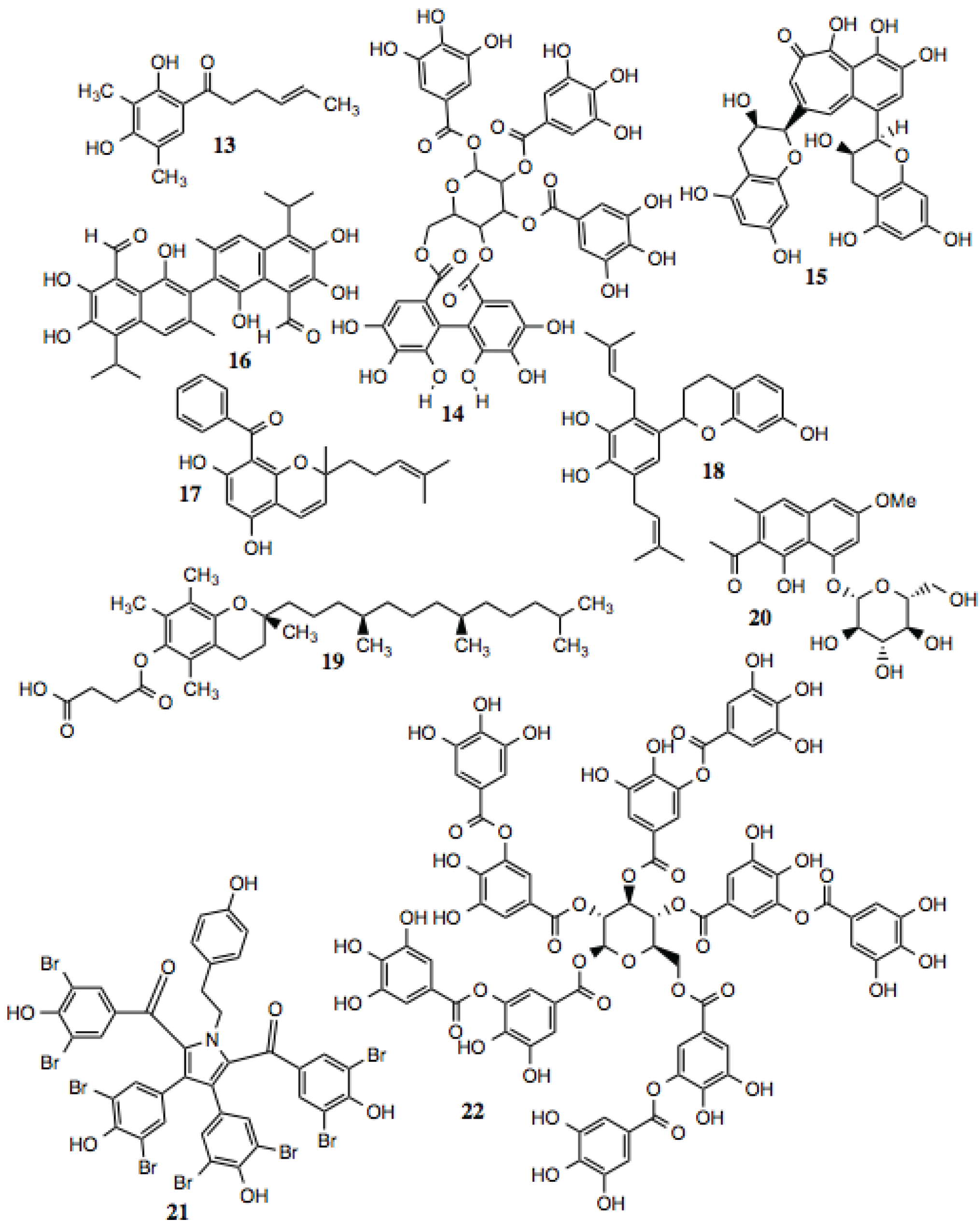
3.1.2. Polyphenols
Polyphenols are defined as compounds that contain multiple phenolic moieties and often are poly-hydroxylated. This family of compounds is large, and contains multiple subtypes. Polyphenols generally exhibit anti-oxidant activity and can protect against ROS
in vitro. The actual mechanisms behind these activities, however, have not been fully evaluated. Several of the compounds shown in
Figure 3 and listed in
Table 2 inhibit Hsp90-dependent proteins, and manifest cytotoxic, anti-proliferative, anti-inflammatory, and anti-viral activities, among others, that might be explained by their ability to inhibit Hsp90. Theaflavin (compound
15) has recently been reported to be an inhibitor of Hsp90 [
56].
Figure 3.
Polyphenols and related compounds.
Figure 3.
Polyphenols and related compounds.
Table 2.
Polyphenols and related compounds.
Table 2.
Polyphenols and related compounds.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 13 | 2'3'-dihydrosorbicillin | >100 | Moderate cytotoxic activity against cancer cell lines [57]. |
| 14 | Tellimagrandin II | ~70 | Anti-HIV [58]; suppression of sarcoma tumor cell growth [57]. |
| 15 | Theaflavin | ~25 | Anti-viral, anti-inflammatory [59]; anti-proliferative against leukemia cells via Akt down-regulation via Hsp90 inhibition [56]; inhibits NF-κB and MAPK signaling [60]; |
| 16 | Gossypol | ~50 | Anti-oxidant; broad anti-cancer activity; anti-viral; anti-protozoan; anti-bacterial; contraceptive [61]. |
| 17 | Similar to catechin (CID 10670714) | ~50 | |
| 18 | Flavan-3-ol (AC1MR5D9, CID 3512639) | ~60 | |
| 19 | (+)-α-Tocopherol acid succinate | ~50 | |
| 20 | Torachrysone 8-O-Glucoside (CID 11972479) | ~25 | Anti-oxidant [62]; inhibits LPS-induced NO production and NF-κB activation [63]. |
| 21 | Polycitone A | ~10–20 | |
| 22 | Tannic acid | ~5 | |
3.1.3. Flavonoids
Flavonoids (
Table 3,
Figure 4) represent a large and diverse group of compounds that reside within the class of polyphenols. Flavonoids comprise approximately one half of all identified polyphenolic compounds. Aside from their aromaticity, the molecules have no unifying characteristic, except that they contain two or more six-membered rings, as well as at least one oxygen atom in the form of an ether or ketone. Many of these compounds contain multiple ketones and hydroxyl groups. This family of compounds is abundant in a number of substances used in traditional medicine. These substances have been shown to exhibit activities against allergy, inflammation, infection, tumors, diarrhea, and others. They have also been credited with wound healing and other beneficial properties. As ubiquitous as flavonoids are in plants, they are found in many foods. Examples are quercitin, epigallocatechin gallate (EGCG), resveratrol, and others.
Several hits from the screening belong to the flavonoid family (
Table 3,
Figure 4). Some of the compounds contain the typical bicyclic core along with a benzene ring fused to a pyran or pyrone, as well as a phenyl group attached to the flavan, isoflavan, or neoflavan. They also contain phenyl or aliphatic groups of varying saturation and oxygen incorporation. Additionally, some of the compounds fall into the subgroup of flavonoids known as chalcones (
Table 4,
Figure 5), which are metabolic precursors to flavonoids. These chalcones are characterized by two benzenes bridged by a 2-propen-1-one group.
Flavonoids are found throughout the plant kingdom. They have been used for the treatment of disease for centuries. Many flavonoids also demonstrate anti-microbial activity. Argentinian folk medicine has made use of a plant containing the glycosylated flavonol, quercetagetin-7-arabinosyl-galactoside, for the treatment of infectious diseases [
64]. In another study, epigallocatechin gallate, a type of flavonoid found abundantly in green tea, demonstrated strong anti-bacterial activity, resulting from damage to the lipid bilayer [
65]. However, EGCG has also been shown to bind Hsp90 and to induce degradation of Hsp90-dependent substrates [
66].
Table 3.
Flavonoids.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 23 | Glabranine (CAS 41983-91-9) | >75 | Anti-viral [67]. |
| 24 | 5-Methoxyflavone | ~100 | |
| 25 | MolPort-005-945-561 (CID 4560115) | ~70 | |
| 26 | 2'-hydroxy-b-naphtho-flavone | ~350 | |
| 27 | 7,8-dihydroxy-2-(2-hydroxyphenyl)chromen-4-one | ~10 | |
| 28 | Biochanin A (5,7-dihydroxy-3-(4-methoxyphenyl)-chromen-4-one) | >90 | Anti-proliferative, anti-inflammatory, cytotoxic; inhibits iNOS expression, MAPK phosphorylation and NF-κB activation [68]. |
| 29 | 2',3',6-Trimethoxyflavone (2-(2,3-Dimethoxyphenyl)-6-methoxy-4H-chromen-4-one) | >90 | |
| 30 | 3',4'-Dimethoxy-3-hydroxy-6-methylflavone | ~30–80 | |
| 31 | Luteolin | >100 | Induction of unfolded protein response and apoptosis in neuroblastoma [69]; inhibits LPS-activated, Akt-mediated activation of NF-κB in macrophages [70]. Anti-tumor activity through EGFR pathway suppression in breast cancer cells [71]. Shown to inhibit Hsp90 [72]. |
| 32 | Mangostin [1,7-bis(3-methylbut-2-enyl)-3,6,8-trihydroxy-2-methoxy-xanthen-9-one] | ~60 | Xanthanoid–Induces cell cycle arrest and apoptosis in colon [73] and prostate cancer cells [74]. Blocks activation of MAPK and Akt pathways [75]. |
| 33 | MolPort-001-742-269 (CID 38356110) | ~65 | |
| 34 | 6-Hydroxy-7-methoxyflavone | ~45 | |
| 35 | Gambogic acid | ~2 | Demonstrated to inhibit Hsp90 [33,76]. |
| 36 | Tetrahydrogambogic acid | ~10 | |
| 37 | Dimethyl Gambogate | ~2 | |
| 38 | Derrubone | ~0.2 | Inhibitor of Hsp90 [77]. |
Figure 4.
Flavonoids and related compounds.
Figure 4.
Flavonoids and related compounds.
Over the course of twenty years, apigenin, a simple flavone, was assayed for its anti-bacterial activity, and was found active against more than fifteen types of pathogenic bacteria, including
S. aureus, MRSA,
E. coli,
P. aeruginosa, and
K. pneumonia [
78]. In addition to anti-microbial activity, these compounds also demonstrate anti-cancer activity. For example, apigenin (
Figure 3) exhibited strong
in vitro anti-tumor and anti-angiogenic activity against human lung, prostate, and ovarian cancer cells. In each of these cases, the expression of VEGF and HIF-1alpha were suppressed [
79,
80], both of which are Hsp90-dependent clients.
Among the hits from this screen, compounds
23,
24,
27,
29,
30 and
34, contain the same flavone core as apigenin. Two contain a flavonone core that is nearly identical, except for saturation of the C-2 double bond found in flavones. The physiological effects of several of the flavonoid hits implicating them as possible Hsp90 inhibitors are noted in
Table 3.
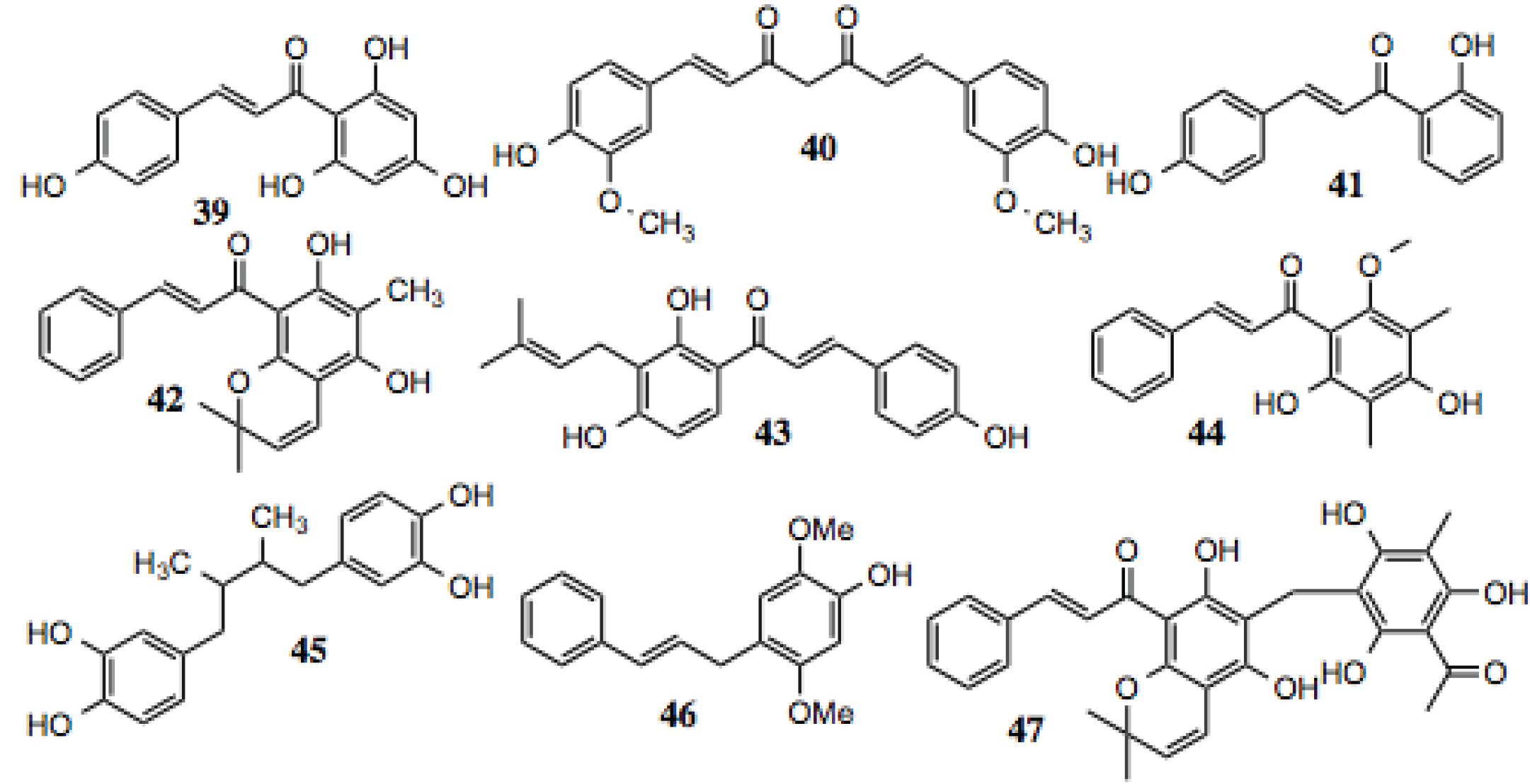
3.1.4.Chalcones
Chalcones are a structurally distinct subclass of flavonoids from which several hits were identified (
Table 4,
Figure 5). Chalcones share many of biochemical characteristics with other flavonoids, as they exhibit anti-fungal [
81], anti-inflammatory [
82], anti-tumorogenic [
83], anti-HIV, and anti-plasmodial activities [
84], amongst others (
Table 4). A number of cellular proteins were identified as targets for the chalcones, many of which are known to be dependent upon Hsp90 (
Table 4), including Akt, NF-κB, mTOR, STAT3, HIF-1α, iNOS, and others [
85].
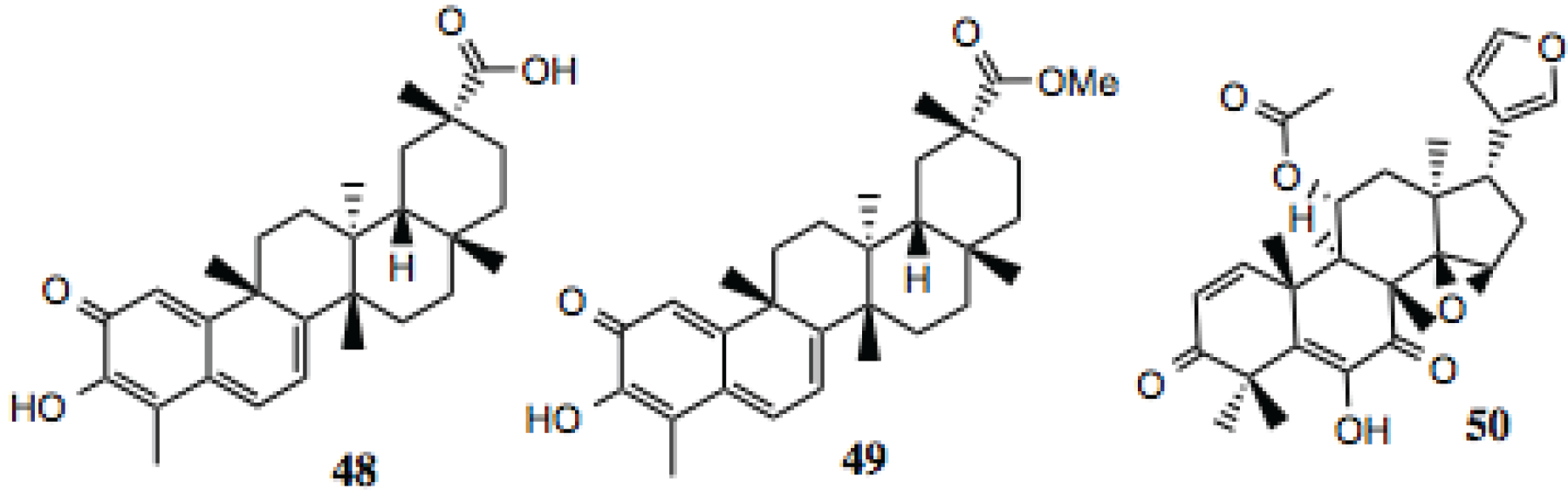
3.1.5. Pentacyclic Triterpenoids
Three pentacyclic triterpenoids were identified in the screening (
Table 5,
Figure 6), celastrol, its methyl ester and anthothecol. Celastrol is a well-established Hsp90 inhibitor [
86]. Anthothecol is a limonoid related to degunin. Degunin has also been identified as an inhibitor of the Hsp90 machine [
87,
88].
Table 4.
Chalone compounds.
Table 4.
Chalone compounds.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 39 | Phloretin | ~35–90 | Induction of apoptosis in breast cancer cells via Bcl-xL degradation [89]. |
| 40 | Curcumin | ~70 | Anti-proliferative, anti-tumor, anti-inflammatory via suppression of NF-κB activation [90]; reported Hsp90 inhibitor [91]. |
| 41 | 2',4-Dihydroxychalone | ~120 | |
| 42 | [6-methyl-5, 7-dihydroxy-2,2-dimethylchromen-8-yl]-3-phenylprop-2-en-1-one Similar to rotterlin and catechin | ~60 | |
| 43 | (CID 193568) Corylifolinin; isobacachalone | ~50 | Inhibition of LPS-induced NO production [92]. |
| 44 | Dimethyl cardamonin (CID 10424762) | ~70 | Antibacterial; anti-fungal [93,94]; anti-proliferative via cell cycle arrest; anti-inflammatory via blocking NF-κB activation [95,96,97,98]. |
| 45 | Nordihydroguaiaretic acid | ~35–80 | Phase II study for effect on prostate cancer. Increased doubling time of PSA. Thought to inhibit IGF1R and HER2 [99]. Repressed breast tumor growth via mTORC1 inhibition [100]. |
| 46 | Violastyrene | ~20 | |
| 47 | Rottlerin | ~60 | Anti-proliferative [101,102]; cytotoxic to pancreatic cancer cells via PI3K/Akt/mTOR inhibition [103]; inhibits NF-κB; STAT and amyloid aggregation [104]. |
Figure 5.
Chalones and related compounds.
Figure 5.
Chalones and related compounds.
Table 5.
Pentacyclic triterpenoids.
Table 5.
Pentacyclic triterpenoids.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 48 | Celastrol | ~2 | Hsp90 inhibitor [86]. |
| 49 | Celastrol methyl ester | ~2 | |
| 50 | Anthothecol | ~6 | Antimalarial [105]. |
Figure 6.
Pentacyclic triterpenoids.
Figure 6.
Pentacyclic triterpenoids.
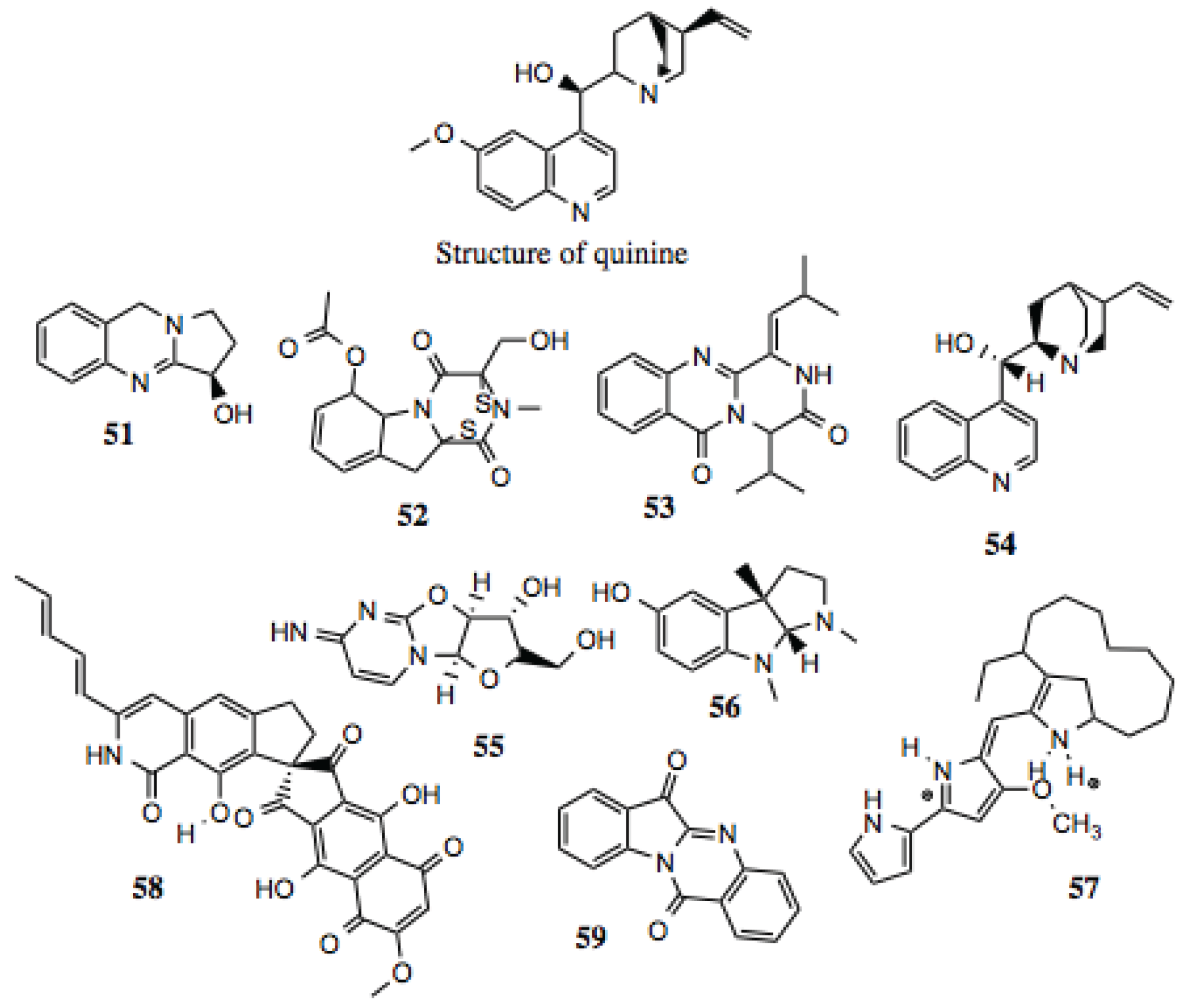
3.1.6. Alkaloids
Alkaloids are broadly defined as containing basic nitrogen atoms within their ring structures. As this definition includes a large number of potential compounds, the family is further broken down into smaller subdivisions. Regardless, the compounds within the family that have demonstrated biological activities are diverse, with no single structure or group of structures requisite for activity. For the purposes of this screen, alkaloids were regarded as molecules that contain a cyclic nitrogen atom. Similar to other groups identified in this screen, alkaloids (
Table 6,
Figure 7) demonstrate a wide range of medically relevant bioactivities, which include anti-tumor, anti-hypertensive, anti-depressant, anti-microbial, anti-inflammatory, and other activities [
106], as well as inhibiting the function of some well known Hsp90-dependent proteins (
Table 6). A well-known example of a medicinal alkaloid is quinine (
Figure 7), isolated from the tropical medicinal plant
Cinchona succirubra, which has been used to treat malaria for hundreds of years [
53].
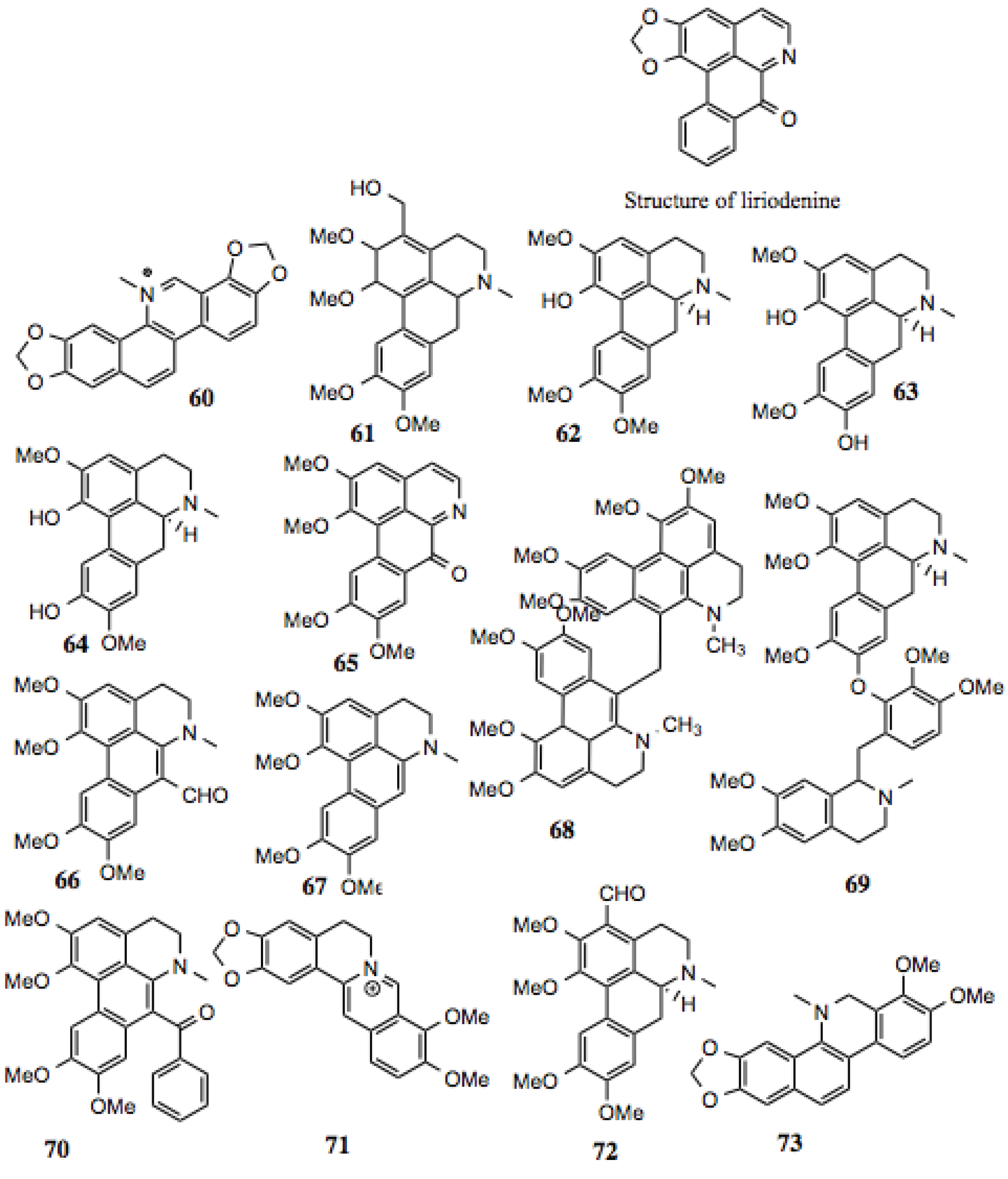
3.1.7. Benzylisoquinoline Alkaloids
Benzylisoquinoline alkaloids (
Table 7,
Figure 8) comprise a subset of alkaloid compounds characterized by an aromatic, tetracyclic skeleton, containing three benzene moieties, and a fourth cycle containing the alkaloid nitrogen. Most of the compounds shown in
Figure 7 are aporphines that are highly similar, and differ only by the presence or location of a hydroxyl, methoxy, or keto group. Some, however, incorporate an additional ring, or exist as a dimer of two aporphine molecules. While there is little known about most of the aporphine compounds identified in this screen, some have no prior designation. One example is glaucine, which manifests a host of activities
in vitro, including relaxation of bronchia via inhibition of its contraction, reduction in superoxide generation in stimulated polymorphonuclear leukocytes and eosinophils, reduction of elastase release, leukotriene production, and intracellular Ca
2+ in PMN’s, platelet aggregation, and eosinophil peroxidase release. These effects make glaucine a likely candidate for the treatment of bronchiodilation and inflammation [
107].
Table 6.
Alkaloid compounds.
Table 6.
Alkaloid compounds.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 51 | Peganine (CID 72610) | ~130 | Modest anti-proliferative and cytotoxic activity [108]; anti-mycobacterial [109]; anti-leishmania [110,111]. |
| 52 | Gliotoxin acetate (CID 21127802) | ~15 | Anti-viral; anti-mycobacterial; inhibition of NF-κB; anti-leukemic; anti-tumor [112,113,114]. |
| 53 | 19-A7: (Z)-4-isopropyl-1-(2-methylpropylidene)-1,2-dihydro-6H-pyrazino[2,1-b]quinazoline-3,6(4H)-dione | ~60 | |
| 54 | (+)-Cinchonine | ~30 | Circumvention of P-glycoprotein mediated multi-drug resistance [115]. |
| 55 | (2S,3R,3aS,9aR)-2-(hydroxymethyl)-6-imino-2,3,3a,9a-tetrahydro-6H-furo[2',3':4,5]oxazolo[3,2-a] pyrimidin-3-ol | ~50 | |
| 56 | (−)-Eseroline | ~50–90 | |
| 57 | Metacycloprodigiosin; Streptorubin | ~10 | Cytoxic activity against cancer cell lines [116,117]. |
| 58 | Fredericamycin | ~3 | Cytotoxic, anti-bacterial, anti-fungal, anti-tumor activities; topoisomerase inhibitor; cell cycle arrest [118,119]. |
| 59 | Tryptanthrin | ~15 | Induced apoptosis in human leukemia cells; suppression of NO production [120]. |
Figure 7.
Alkaloid compounds.
Figure 7.
Alkaloid compounds.
Aporphines also demonstrate
in vitro anti-viral activity. A number of compounds inhibit polio-virus infection of cultured mammalian cells by 50% at low micromolar concentrations [
121,
122]. Additionally, the aporphines dicentrine, glaucine, corydine, and apomorphine, which are analogs of the aporphines presented in this study, demonstrate anti-proliferative activity against mouse leukemia, melanoma, bladder cancer, and colon cancer cells [
123].
As discussed earlier, Hsp90 contains a distinct ATP-binding domain, the specificity of which has been exploited for the development of inhibitors. This domain contains a Bergerat fold, and is shared by only a few protein families, which include DNA gyrase, a type II topoisomerase. Some aporphines are inhibitors of topoisomerase II, increasing the likelihood that highly similar compounds from this screen are also Hsp90 inhibitors. One such example is liriodenine (
Figure 7), which manifests activity against human cancer cells [
124], Gram-positive bacteria, yeast, and fungi [
125,
126,
127].
Table 7.
Benzylisoquinoline Alkaloids.
Table 7.
Benzylisoquinoline Alkaloids.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 60 | Sanguinarine | ~70 | Induces cell cycle arrest and apoptosis in human cervical cancer cells [128]; anti-inflammatory [129]; anti-fungal [130]. |
| 61 | 3-hydroxymethyl-glaucine | ~80 | |
| 62 | Thaliporphine; thalicmidine | ~9 | Inhibits the activity of LPS-induced NOS; inhibits LPS-induced nuclear translocation of NF-κB [131]. |
| 63 | Isoboldine | ~30–80 | Anti-viral [121]. |
| 64 | Bracteoline | ~80 | |
| 65 | 7-oxoglaucine | ~80 | Anti-plasmodial [132]; anti-inflammatory [133]. |
| 66 | 7-Formyl-dehydroglaucine | ~1–3 | |
| 67 | Dehydroglaucine | 0.9 | Anti-microbial, some anti-fungal activity [126]. |
| 68 | Dehydroglaucine dimer | 0.1–0.4 | |
| 69 | Glaucine derivative | >30 | |
| 70 | Dehydroglaucinylphenone | ~7–20 | |
| 71 | Berberine | ~40 | Anti-tumor, anti-metastatic, inhibits HIF1α [129,134,135,136]. |
| 72 | 3-Formyl-glaucine | ~8–20 | |
| 73 | Dihydrosanguinarine derivative | ~40 | Anti-fungal [130]. |
Figure 8.
Benzylisoquinoline Alkaloids.
Figure 8.
Benzylisoquinoline Alkaloids.
3.1.8. Cyclic Peptides
The McAlpine Laboratory has investigated the anti-cancer and Hsp90-inhibiting activities of naturally occurring macrocyclic peptides. These molecules and their derivatives have demonstrated modest activity against a number of cancers [
137]. The rationale behind the pursuit of these particular scaffolds is that similar compounds have been identified as antibiotics and anti-fungals, while maintaining anti-cancer activity. The polypeptide nature of these compounds may confer the ability to mimic hydrophobic portions of client proteins. Four of the compounds identified from this screen (
Table 8,
Figure 9) contain moieties that could mimic hydrophobic amino acids. Of particular interest is antibiotic A83586 C (compound
75), which manifest potent anti-proliferative activity against cancer cells [
138].
Table 8.
Cyclic peptides.
Table 8.
Cyclic peptides.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 74 | Tyrothricin | ~5 | Antibiotic [139]. |
| 75 | Antibiotic A83586C | ~10 | Anti-proliferative activity against cancer cells [138]. |
| 76 | P12 | ~10 | |
| 77 | Cyclopeptide L-156373 | ~10 | |
Figure 9.
Cyclic peptides.
Figure 9.
Cyclic peptides.
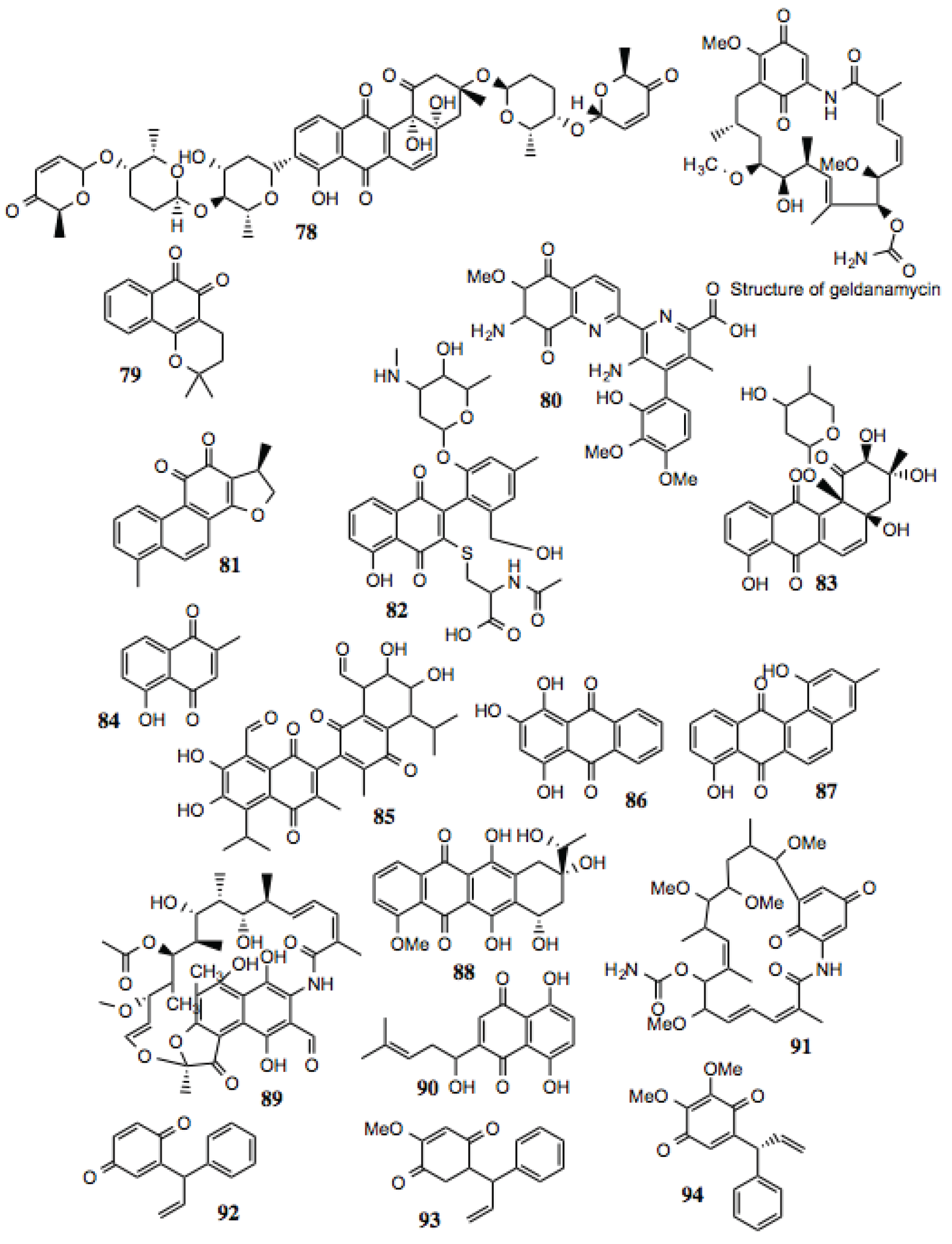
3.1.9. Quinones
Quinones represent an extremely large and diverse family of compounds. Essentially, the only thing that differentiates a quinone from other classes of compounds is the presence of two keto groups on an unsaturated six-membered ring. As such, an enormous array of functional and structural groups that decorate this motif is possible. Generally, quinones are redox-active, making them promising compounds with which to treat cancer, but also potential liabilities that may arise as a consequence of this activity. The production of reactive oxygen species resulting from exposure to quinone-containing compounds is a process that is potentially destructive to any cell. It has not been established whether the redox potential of these compounds contributes to the anti-cancer activity by increasing their ability to inhibit Hsp90, or by increasing cellular stress alongside Hsp90 inhibition. It should be noted, that reduction of 17-DMAG and 17-AAG to the corresponding hydroquinones resulted in increased Hsp90 inhibitory activity. [
140]
Some well-established Hsp90 inhibitors, such as geldanamycin (
Figure 10) and its derivatives, contain quinone moieties. Consequently, we have observed reticulocyte lysate treated with these inhibitors to possess a distinctive dark red color, attributable to met-hemoglobin formation resulting from the oxidative activity of such compounds. Similarly, geldanamycin has been shown to generate reactive oxygen species
in vitro and in cell culture [
141]. Structural studies demonstrate direct binding of these compounds to the ATP-binding site at the N-terminus of Hsp90. Not surprisingly, the antibiotic, rifamycin, contains a reduced quinone moiety within a similar anthroquinone ansamycin structure, and has similar activity as geldanamycin. Seventeen quinones were identified in this screen (
Table 9,
Figure 10). As indicated in
Table 9, a number of these compounds display anti-cancer, anti-trypanosomal, anti-viral, and anti-inflammatory activities, as well as having the capacity to inhibit the activities of several well-known Hsp90-dependent proteins.
3.1.10. Other Compounds
Compounds identified in this section (
Figure 11,
Table 10) do not fit well into any of the previously described families. Some of these are known biochemicals, such as Vitamin D2, 9-cis-retionioc acid, prostaglandin J2 and L-adrenaline, and have not been implicated as Hsp90 inhibitors, although Vitamin D2 [
142,
143] and retinoic acid [
144,
145] have demonstrated anti-cancer activity. Others are more exotic and little, if anything is known about their mechanism of action. Recently, the anti-cancer activity of hypericin has been tied to its ability to inhibit Hsp90 stabilization of HIF-1α [
146].
Table 9.
Quinones.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 78 | Vineomycin A1 | ~100 | |
| 79 | β-Lapachone (CID 3885) | ~20 | Anti-cancer; anti-trypanosomal activity; anti-viral [147,148,149,150,151,152,153,154,155]. |
| 80 | Streptonigrin (CID 5351165) | ~15 | Disrupts NF-κB activation; antibacterial, antifungal, antiviral, anti-glioma [156,157,158,159,160,161]. |
| 81 | 15.16-Dihydrotanshinone | ~80 | Anti-cancer; inhibition of HIF-1α; depletion of Bcl-2 [162,163,164]. |
| 82 | 2-A5: 1,4-Napthoquinone derivative | ~10 | |
| 83 | 30-D10: 9,10-Anthraquinone derivative | ~2 | |
| 84 | Plumbagin | <0.05 | Widely studied anti-cancer activity; reported to target EGFR, STAT-3, Akt, and NF-κB pathways [165,166]. |
| 85 | 7-[8-formyl-6,7-dihydroxy-3-methyl-5-(methylethyl)-1,4-dioxo(2-naphthyl)]-2,3- dihydroxy-6-methyl-4-(methylethyl)-5,8-dioxonaphthalenecarbaldehyde | ~6 | |
| 86 | 1,2,4-trihydroxyanthracene-9,10-dione | ~40–100 | |
| 87 | Tetrangulol G2 | ~20 | |
| 88 | Dihydodaunomycinone; Leukaemomycin-D | >60 | |
| 89 | 3-Formyl Rifamycin SV | ~15–35 | Inhibition of Vaccinia virus assembly [167]. |
| 90 | Shikonin | ~75 | Anti-inflammatory, anti-tumor, and wide-ranging activities reported [166,168]. |
| 91 | Herbimycin | ~40 | Hsp90 inhibitor [1,169,170,171]. |
| 92 | Dalbergione | ~2 | |
| 93 | 4'-Methoxydalbergione | ~5 | Anti-trypanosomal [172]. |
| 94 | 3'4'-Dimethoxydalbergione | ~5 | Anti-trypanosomal [172]. |
Figure 11.
Other miscellaneous compounds.
Figure 11.
Other miscellaneous compounds.
Table 10.
Other compounds.
Table 10.
Other compounds.
| # | Location and/or name | IC50 (µM) | Properties |
|---|
| 95 | 2-((4R,6aS,7R,11bR)-4,7,11b-trimethyl-1,2,3,4,4a,5,6,6a,7,11,11a,11b-dodecahydro-phenanthro[3,2-b]furan-4-yl) acetic acid | >75 | |
| 96 | (+)-Dehydroabietylamine; Leelamine | ~30–75 | |
| 97 | Vitamin D2 | ~60 | |
| 98 | Vitamin D2 metabolite | ~60 | |
| 99 | L-Adrenaline | ~55 | |
| 100 | Rolitetracycline | ~20 | |
| 101 | MolPort-005-945-572 | ~50 | |
| 102 | CID 45359640 | ~50 | |
| 103 | (E)-1-(1,3,6,8-tetramethoxy- naphthalen-2-yl)but-2-en-1-one | ~5 | |
| 104 | CID 53984538: 8,11- anacardic acid | ~10 | Anti-cancer; induction of the UPR [173,174]. |
| 105 | (4R,7R,11bS)-4,11b-dimethyl-7-(((Z)-2-methylbut-2-enoyl)oxy)-8-methylenetetradecahydro-6a,9-methanocyclohepta[a]naphthalene-4-carboxylic acid | ~40 | |
| 106 | (4bR,9S,10S)-3,4-dihydroxy-2-isopropyl-10-methoxy-8,8-dimethyl-6,7,8,8a,9,10-hexahydro-5H-9,4b-(epoxymethano) phenanthren-12-one | ~30 | |
| 107 | CID 13818684 | ~15 | |
| 108 | Manumycin A | ~30–50 | Inhibition of STAT-3 and telomerase; down-regulation of Akt and MEK; anti-cancer [175]; induces autophagy [176]. |
| 109 | Striatal A | ~2 | |
| 110 | Striatal B | ~2 | Inhibition of growth of multiple cancer cell lines- NCI cell line growth inhibition assay [PubChem CID 329431]. |
| 111 | Striatal C | ~2 | |
| 112 | CID 45360154 | ~50 | |
| 113 | 9-Cis-retinoic acid | ~30–50 | |
| 114 | Prostaglandin J2 | ~75–100 | |
| 115 | Hypericin | ~50–75 | Inhibits HIF-1α [146]. |
| 116 | Hypocrellin A | ~5–10 | |
3.2. Further Characterization of Select Putative Inhibitors
The HTS of commercially available natural product libraries identified over a 100 compounds as potential Hsp90 Inhibitors. This raises the question inherent to hits identified in HTS: which of the compounds should one select for further characterization using secondary screens? Of these compounds, derrubone [
77], compounds containing the 1,4-naphthoquinone scaffold [
177], and gambogic acid [
33] have been characterized, in addition to silybin [
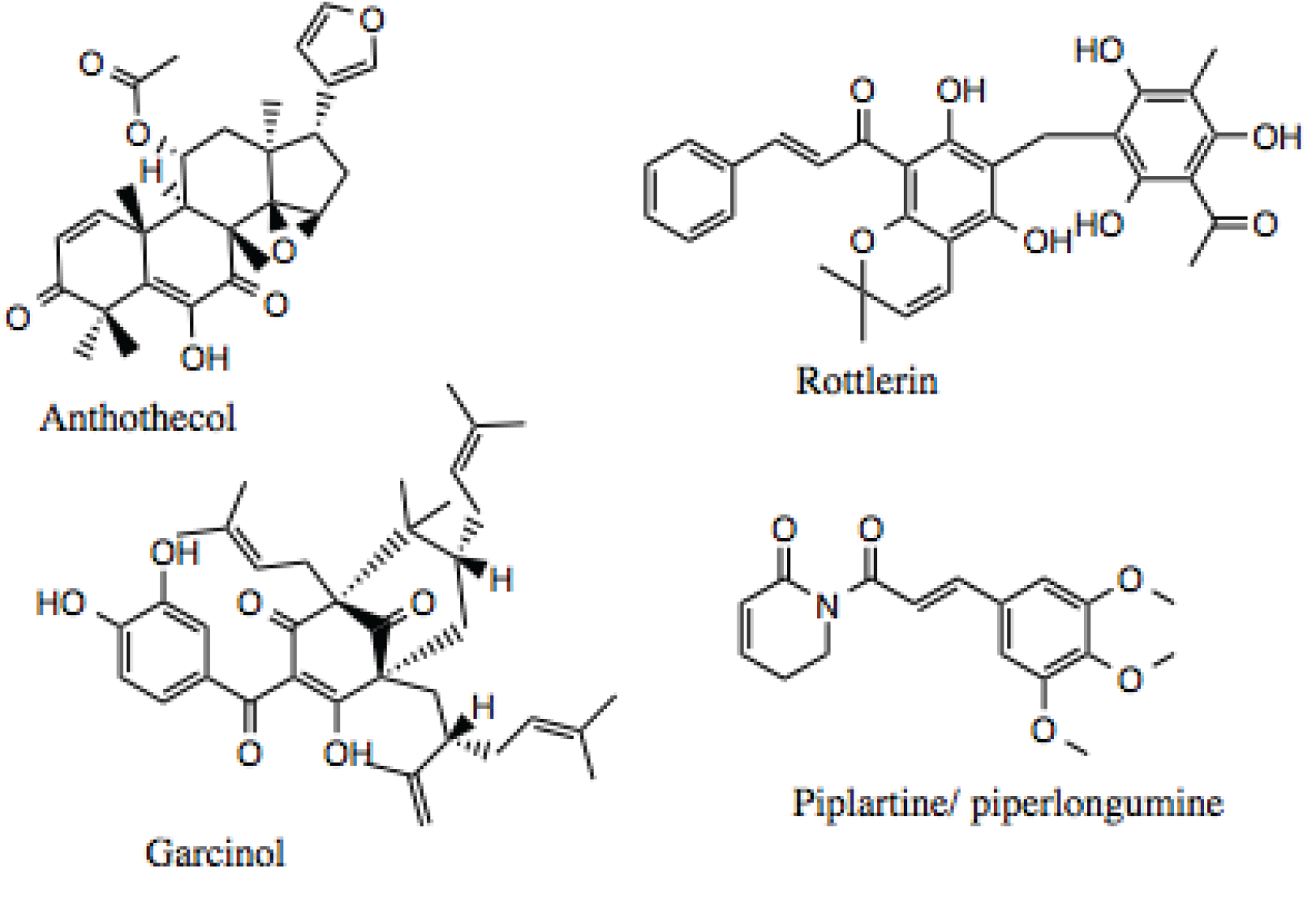
178], which was identified as an Hsp90 inhibitor by mining the literature. Based on the results from our high-throughput screen and data mining, anthothecol and rottlerin were selected for further characterization. In addition, during the search through the literature for natural products that displayed broad biological activities, garcinol, a gambogic acid-like compound, and piplartine/piperlongumine, a polyphenol chalcone-like compound were identified as potential Hsp90 inhibitors. An additional consideration in the selection of rottlerin, garcinol, and piplartine/piperlongumine was that the literature indicated that these compounds were being discussed as possible candidates for clinical trials.
Of the four compounds chosen for additional investigation, rottlerin (
Figure 5 and
Figure 12) is the best studied, and, as noted above has been discussed as a possible candidate for clinical trials [
101]. It has been used in traditional medicine and demonstrates many physiologically significant biological activities. Rottlerin is isolated from the tropical tree
Mallotus philippinensis and displays cytotoxicity against a number of cancer types, including lung, breast, lymphocytic leukemia, and multiple myelomas. While the activity of rottlerin was initially attributed to inhibition of PKCδ [
101], this mechanism of action has largely been called into question [
101,
103]. Rottlerin appears to inhibit a combination of signal transduction pathways at multiple levels [
101], making it a good Hsp90 inhibitor candidate. For example, an array of human malignant tumor cells was treated with rottlerin, and all lines were found to undergo apoptosis mediated by Death Receptor 5 (DR5) [
179]. Rottlerin has also been reported to inhibit the kinases PRAK, MAPKAP-K2, Akt, and CaMK [
180].
Figure 12.
Structure of anthothecol, rottlerin garcinol and piplartine/piperlongumine.
Figure 12.
Structure of anthothecol, rottlerin garcinol and piplartine/piperlongumine.
Anthothecol (
Figure 6 and
Figure 12) is a limonoid natural product isolated from the
Khaya anthotheca tree, and possesses low micromolar activity against the growth of
Plasmodium falciparum in erythrocytes [
105], which is another property manifested by Hsp90 inhibitors. The compound’s structure is similar to that of the Hsp90 inhibitor, gedunin [
88], but it has approximately 10-fold higher activity, as will be demonstrated below.
Garcinol (
Figure 12) was chosen for further study, because it demonstrated the ability to induce apoptosis in a number of cancers, including breast, colon, kidney, prostate, leukemia [
181], pancreatic [
182], and others. The anti-cancer activity of garcinol has been attributed, in part, to its inhibition of STAT and NF-κB signaling [
183,
184]. Additionally, garcinol can inhibit angiogenesis through down-regulation of Prostaglandin E2, VEGF, and IL-8 [
182], which are bonafide Hsp90-dependent clients. Garcinol also exhibits anti-oxidant and anti-inflammatory properties, as it inhibits the production of ROS and nitric oxide [
185].
Piplartine (a.k.a., piperlongumine,
Figure 12) has been shown to suppress platelet-derived growth factor (PDGF) signaling [
186], and inhibit the proliferation of prostate cancer cells through depletion of the androgen receptor, a well-known Hsp90 client protein [
187]. A recent review noted that the pharmacological activities for piplartine reported in the literature include cytotoxic, anti-tumor, anti-angiogenic, anti-metastatic, anti-bacterial, anti-fungal, anti-leishmanial, anti-trypanosomal, and anti-schistosomal activities among others [
188].
In our study, we provide evidence that further implicates these compounds as inhibitors of the Hsp90 complex. We show that, in addition to inhibiting the proliferation of cancer cells, these compounds also inhibit the Hsp90-dependent folding of thermally denatured luciferase in a dose-dependent manner, block the Hsp90-dependent maturation of the heme-regulated eIF2α kinase (HRI), and deplete cells of Hsp90-dependent clients.
3.2.1. Inhibition of Hsp90-Mediated Refolding of Denatured Luciferase
Anthothecol and rottlerin were identified as potential Hsp90 inhibitors by their ability to inhibit refolding of thermally denatured luciferase in screens of natural compound libraries. To more accurately define their inhibitory activity, the compounds were titrated into the assay in a three-fold dilution series. The compounds inhibited refolding of luciferase in a concentration-dependent manner with low µM IC50s (
Table 11). Similarly, garcinol, and piplartine display the ability to inhibit the refolding of luciferase at micromolar concentrations in reticulocyte lysate (
Table 11), implicating them as Hsp90 inhibitors.
Table 11.
IC50 values for inhibition of luciferase refolding and proliferation of MCF7 cells.
Table 11.
IC50 values for inhibition of luciferase refolding and proliferation of MCF7 cells.
| Compound | IC50 luciferase refolding (µM) 1 | IC50 MCF7 proliferation (µM) 2 |
|---|
| Anthothecol | 12 ± 2 | 0.5 ± 0.06 |
| Rottlerin | 63 ± 5 | 7 ± 3 |
| Garcinol | 12 ± 5 | 4 ± 0.6 |
| Piplartine | 80 ± 7 | 10 ± 3 |
3.2.2. Compounds Inhibit Proliferation of Human Cancer Cells
While piplartine, rottlerin and garcinol have well characterized anti-proliferative properties, there were no reports in the literature with regards to the anti-proliferative activity anthothecol. Therefore, the anti-proliferative activity of these four compounds on the growth of MCF7 cells was determined using the MTS assay (
Table 11). In MCF-7 cells, all the compounds cause a 50% reduction in growth in the 0.5–10 µM range (
Table 11). These results confirm the anti-proliferative properties of piplartine, rottlerin and garcinol, and establish anthothecol as anti-proliferative drug.
3.2.3. Inhibition of HRI Maturation
The heme-regulated eIF2α kinase (HRI) is an Hsp90 client kinase which, upon folding by Hsp90, will activate, or mature, by autophosphorylation when heme is deficient [
189]. This activation is dependent on functional Hsp90, and can be detected as an electophoretic mobility shift [
189] when separated by polyacrylamide gel electrophoresis (PAGE) (
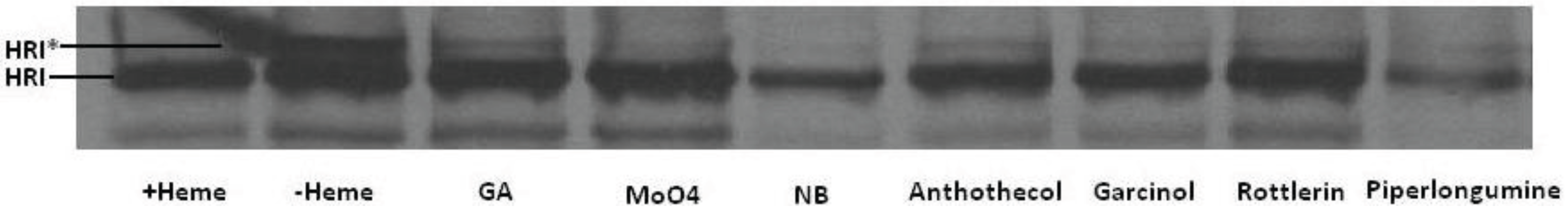
Figure 13). Similar to the known Hsp90 inhibitors, geldanamycin (GA), molybdate and novobiocin, anthothecol, garcinol, rottlerin, and piplartine inhibited the maturation of HRI, as observed by the absence of the slower mobility form of HRI upon PAGE. This result further supports the hypothesis that physiological effects of these four compounds on cells are mediated, at least in part, through their ability to inhibit Hsp90.
Figure 13.
Effect of compounds on HRI’s Hsp90-dependent maturation. [35S]Labeled-HRI was synthesized by TnT in RRL and transferred to heme-deficient lysate for maturation. Translated protein was separated by SDS PAGE, transferred to PVDF membrane, and visualized by X-ray film exposure. The phosphorylated active form of the kinase is indicated with an asterisk. Lanes were treated as follows: heme, no heme, 20 µM geldanamycin (GA), 20 mM sodium molybdate, 20 mM novobiocin, and 100 µM each of anthothecol, garcinol, rottlerin, and piplartine/piperlongumine
Figure 13.
Effect of compounds on HRI’s Hsp90-dependent maturation. [35S]Labeled-HRI was synthesized by TnT in RRL and transferred to heme-deficient lysate for maturation. Translated protein was separated by SDS PAGE, transferred to PVDF membrane, and visualized by X-ray film exposure. The phosphorylated active form of the kinase is indicated with an asterisk. Lanes were treated as follows: heme, no heme, 20 µM geldanamycin (GA), 20 mM sodium molybdate, 20 mM novobiocin, and 100 µM each of anthothecol, garcinol, rottlerin, and piplartine/piperlongumine
3.2.4. Compounds Induce Depletion of Hsp90-Dependent Clients
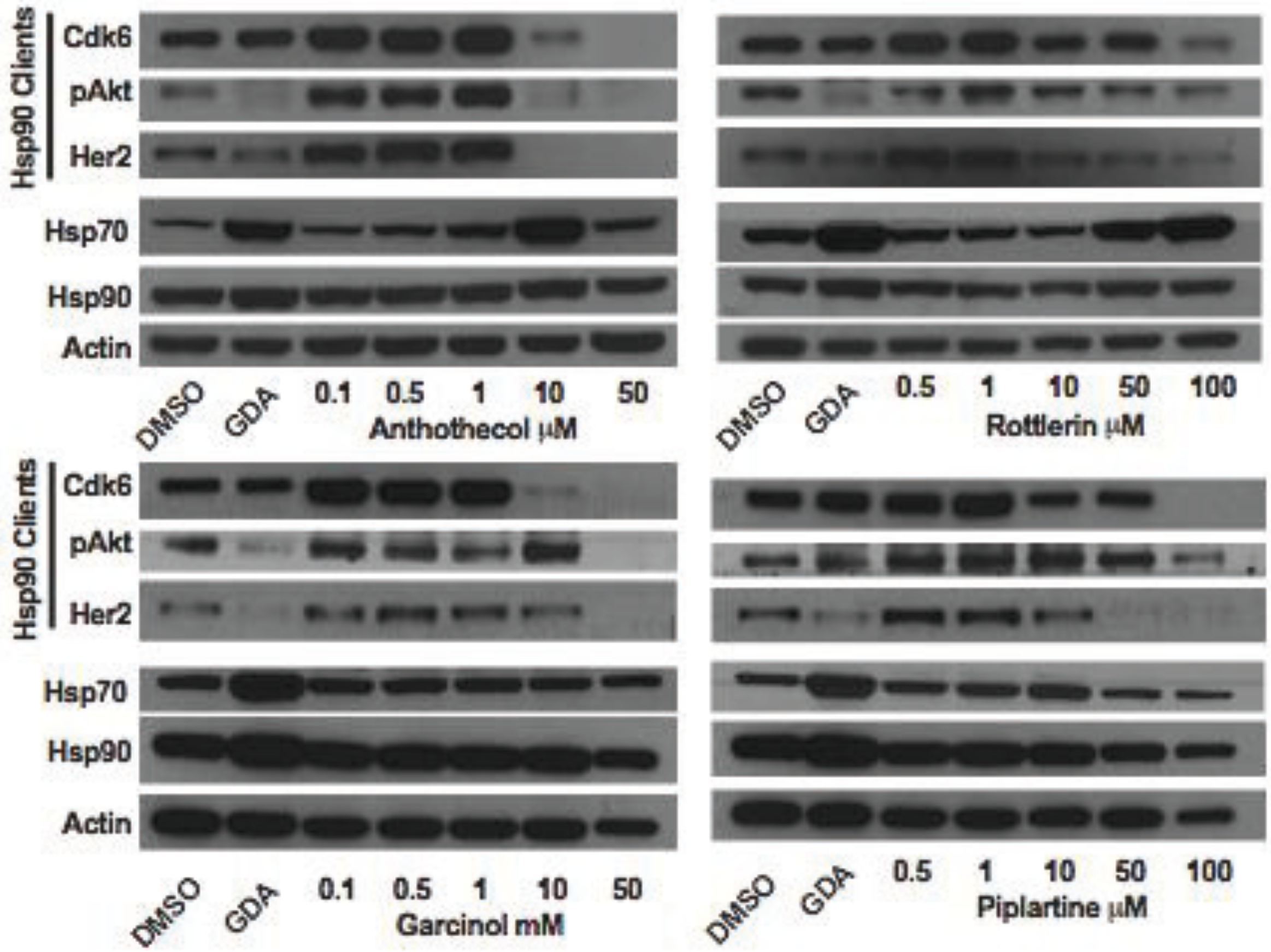
Inhibition of Hsp90 is well known to cause the depletion of Hsp90-depedent client proteins from inhibitor-treated cells. To further test the hypothesis that anthothecol, garcinol, rottlerin and piplartine are inhibitors of Hsp90, the effect of varying concentrations of the compounds on the expression of Hsp90-dependent client proteins Cdk6, pAkt, and Her2 in MCF7 cells was examined (
Figure 14). Consistent with our hypothesis, all four compounds were observed to cause a concentration-dependent depletion of Hsp90 clients from MCF7 cells after 24 h of treatment. None of the compounds increased the expression of Hsp90. However, similar to geldanamycin, anthothecol and rottlerin caused a concentration-dependent increase in the expression of Hsp70, indicating that the compounds likely interact with the ATP binding site in Hsp90’s N-terminal domain. On the other hand, garcinol and piplartine had no effect on the expression of Hsp70, which is a property of compounds that inhibit Hsp90 by binding to its C-terminal domain [
190].
Figure 14.
Western blot for Hsp90-dependent client proteins (Cdk2, pAkt and Her2), and Hsp90 and Hsp70 present in extracts prepared from MCF7 cells treated for 24 h with DMSO (vehicle control), 0.5 µM geldanamycin, or the indicated concentrations of anthothecol, rottlerin, garcinol, or piplartine. Actin was used as the loading control.
Figure 14.
Western blot for Hsp90-dependent client proteins (Cdk2, pAkt and Her2), and Hsp90 and Hsp70 present in extracts prepared from MCF7 cells treated for 24 h with DMSO (vehicle control), 0.5 µM geldanamycin, or the indicated concentrations of anthothecol, rottlerin, garcinol, or piplartine. Actin was used as the loading control.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}











