Scaling the Feeding Mechanism of Captive Alligator mississippiensis from Hatchling to Juvenile


Abstract
:1. Introduction
2. Experimental Section
2.1. Scaling of the Cranial Elements

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimated Size Class (cm) | Cranium Length (cm) | Lower Jaw Length (cm) | Lower Jaw Width (cm) | Upper Jaw Width (cm) | |
|---|---|---|---|---|---|
| n | Average ± SE | Average ± SE | Average ± SE | Average ± SE | |
| 30.5–60.5 | 3 | 5.5 ± 1.04 | 5.6 ± 1.04 | 2.1 ± 1.05 | 2.1 ± 1.02 |
| 60.5–91.5 | 3 | 7.4 ± 1.01 | 8.1 ± 1.03 | 2.3 ± 1.04 | 3.1 ± 1.02 |
| 91.5–122.0 | 3 | 9.8 ± 1.08 | 11.0 ± 1.07 | 3.4 ± 1.07 | 4.1 ± 1.05 |
| 152.0–182.6 | 2 | 21.4 ± 0.00 | 24.5 ± 1.04 | 7.9 ± 1.11 | 8.1 ± 0.00 |
| 213.0–244.0 | 3 | 33.1 ± 1.04 | 40.7 ± 1.04 | 13.2 ± 1.03 | 13.5 ± 1.03 |
| 244.0–274.3 | 2 | 40.7 ± 1.02 | 50.1 ± 1.02 | 16.2 ± 1.05 | 16.6 ± 1.04 |
2.2. Kinematic Scaling of the Feeding Mechanism


| Variable | Units | Definition |
|---|---|---|
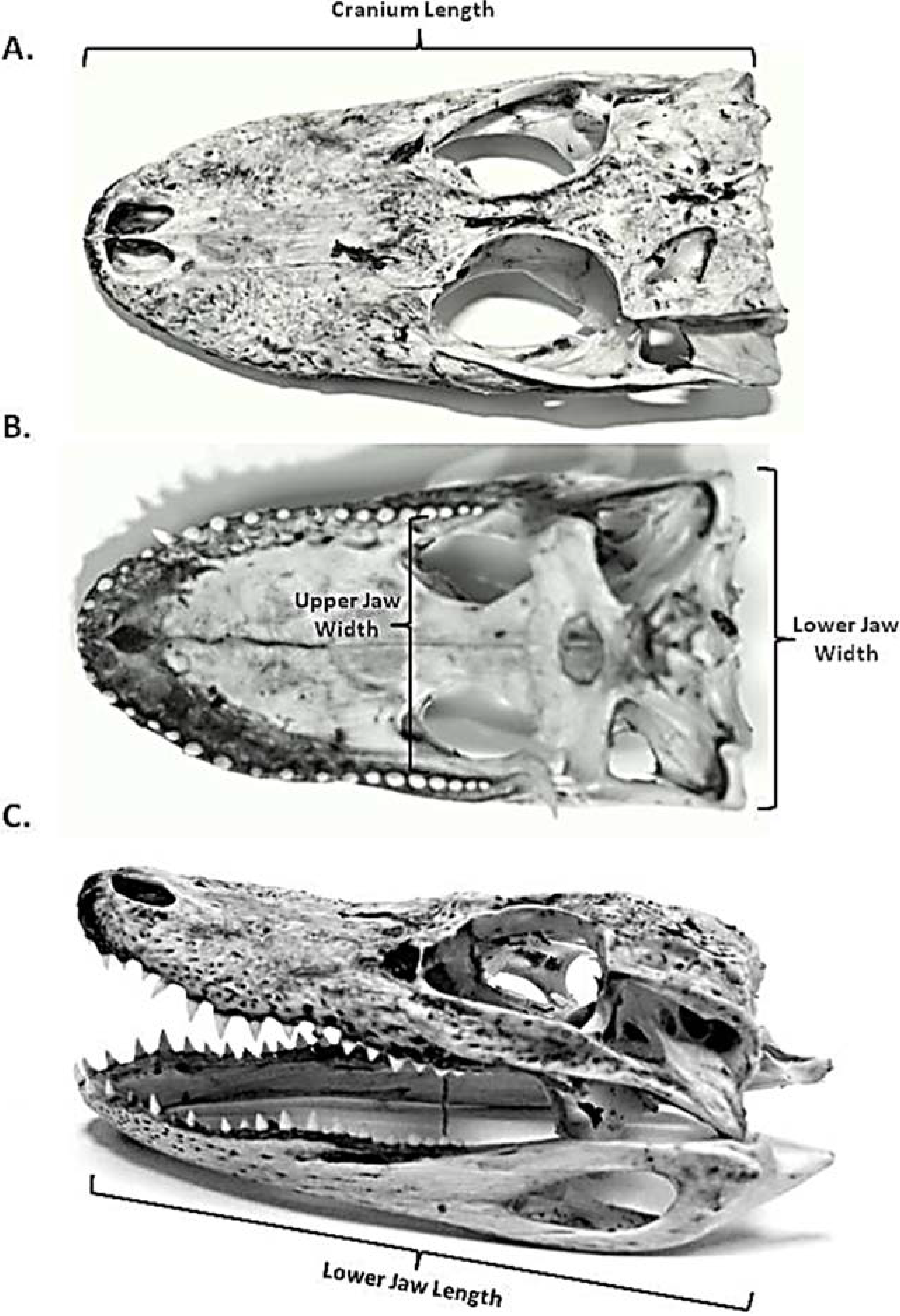
| Cranium Length | mm | Length from the anterior tip of the premaxilla (A) to the posterior portion of the parietal bone (D). See Figure 2. |
| Maximum Gape | mm | Maximum distance from the anterior tip of the premaxilla (A) to the anterior tip of the dentary bone (B). See Figure 2. |
| Lower Jaw Displacement | degree | Maximum postero-ventral rotation of the lower jaw relative to the neurocranium, measured by the angle formed by line segments AC and BC. See Figure 2. |
| Cranial Rotation | degree | Maximum postero-dorsal rotation of the neurocranium relative to the body, measured by the angle formed by line segments AD and DE. See Figure 2. |
| Time Zero (t₀) | The frame prior to mouth opening. | |
| Time to Maximum Gape | s | Time measured from t₀ until maximum gape. |
| Duration of Feeding Bout | s | Times measured from t₀ until individual fully closed its mouth. |
| Maximum Gape Velocity | mm s−1 | Velocity measured from t0 until maximum gape. |
| Lower Jaw Displacement Velocity | degree s−1 | Velocity measured from t0 until maximum displacement. |
| Cranial Rotation Velocity | degree s−1 | Velocity measured from t0 until maximum rotation. |
2.3. Geometric Similarity Model
2.4. Statistics
3. Results and Discussion
3.1. Scaling of the Cranial Elements
3.2. Kinematic Scaling of the Feeding Mechanism

 ). The solid regression line represents the relationship between total length and lower jaw length, dashed line between total length and upper jaw width and the dotted line between total length and lower jaw width.
). The solid regression line represents the relationship between total length and lower jaw length, dashed line between total length and upper jaw width and the dotted line between total length and lower jaw width.
). The solid regression line represents the relationship between total length and lower jaw length, dashed line between total length and upper jaw width and the dotted line between total length and lower jaw width.
). The solid regression line represents the relationship between total length and lower jaw length, dashed line between total length and upper jaw width and the dotted line between total length and lower jaw width.

| Linear Regression | t-test | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | Average (mean ± SE) | Expected GSM Slope | Observed Least Square Slope | y-Intercept | r2 | F | p-value | t | p-value |
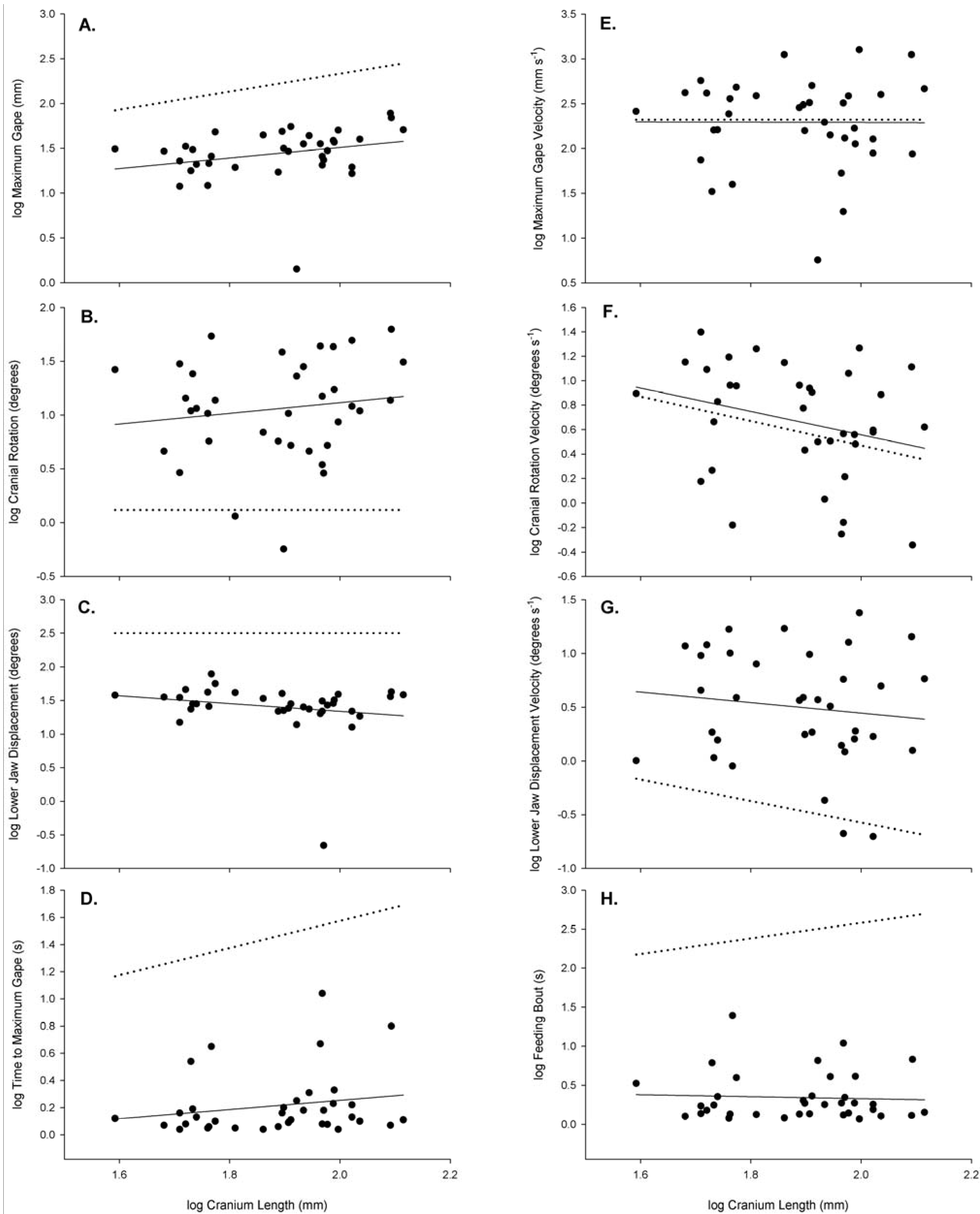
| log Lower Jaw Length (cm) | 1.23 ± 0.159 | 1 | 1.050 | −0.940 | 0.910 | 40.652 | 0.003* | 0.9198 | >0.200 |
| log Upper Jaw Length (cm) | 0.78 ± 0.146 | 1 | 0.970 | −1.220 | 0.917 | 44.383 | 0.003* | −0.4799 | >0.500 |
| log Lower Jaw Width (cm) | 0.74 ± 0.158 | 1 | 1.010 | −1.360 | 0.955 | 23.656 | 0.008* | 0.1935 | >0.500 |
| log Maximum Gape (mm) | 1.44 ± 0.049 | 1 | 0.589 | 0.332 | 0.071 | 2.599 | 0.116 | −16.023 | <0.001* |
| log Cranial Rotation (degree) | 1.06 ± 0.078 | 0 | 0.498 | 0.119 | 0.020 | 0.696 | 0.410 | 11.964 | <0.0025* |
| log Lower Jaw Displacement (degree) | 1.40 ± 0.065 | 0 | −0.581 | 2.500 | 0.040 | 1.405 | 0.244 | −14.225 | <0.001* |
| log Maximum Gape Velocity (mm·s−1) | 2.29 ± 0.082 | 0 | −0.015 | 2.320 | 2.000 × 10−5 | 0.001 | 0.981 | −0.3068 | >0.250 |
| log Cranial Rotation | |||||||||
| Velocity (degrees·s−1) | 0.67 ± 0.077 | −1 | −0.956 | 2.469 | 0.076 | 2.799 | 0.103 | 0.969 | >0.250 |
| log Lower Jaw Displacement | |||||||||
| Velocity (degree·s−1) | 0.50 ± 0.088 | −1 | −0.491 | 1.426 | 0.016 | 0.538 | 0.468 | 10.495 | <0.005* |
| log Time to Maximum Gape (s) | 0.21 ± 0.039 | 1 | 0.340 | -0.427 | 0.037 | 1.319 | 0.259 | 42.357 | <0.001* |
| log Length of Feeding Bout (s) | 0.34 ± 0.052 | 1 | −0.127 | 0.580 | 0.003 | 0.102 | 0.752 | −40.078 | <0.001* |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koehl, M.A.R. When does morphology matter? Annu. Rev. Ecol. Syst. 1996, 27, 501–542. [Google Scholar] [CrossRef]
- Richard, B.A.; Wainwright, P.C. Scaling the feeding mechanism of Largemouth Bass (Micropterus. salmoides): Kinematics of prey capture. J. Exp. Biol. 1995, 198, 419–433. [Google Scholar] [PubMed]
- Dodson, P. Functional and ecological significance of relative growth in alligator. J. Zool. 1975, 175, 315–355. [Google Scholar] [CrossRef]
- Erickson, G.M.; Lappin, A.K.; Vliet, K.A. The ontogeny of bite-force performance in American alligator (Alligator mississippiensis). J. Zool. Lond. 2003, 260, 317–327. [Google Scholar] [CrossRef]
- Delany, M.F.; Abercrombie, C.L. American alligator food habits in northcentral Florida. J. Wildl. Manag. 1986, 50, 348–353. [Google Scholar] [CrossRef]
- Subalusky, A.L.; Fitzgerald, L.A.; Smith, L.L. Ontogenetic niche shifts in the American alligator establishes functional connectivity between aquatic systems. Biol. Conserv. 2009, 142, 1507–1514. [Google Scholar] [CrossRef]
- Allen, V.; Elsey, R.M.; Jones, N.; Wright, J.; Hutchinson, J.R. Functional specialization and ontogenetic scaling of limb anatomy in Alligator mississippiensis. J. Anat. 2010, 216, 423–445. [Google Scholar] [CrossRef] [PubMed]
- Livingston, V.J.; Bonnan, M.F.; Elsey, R.M.; Sandrik, J.L.; Wilhite, D.R. Differential limb scaling in American alligator (Alligator mississippiensis) and its implications for archosaur locomotor evolution. Anatom. Record 2009, 292, 787–797. [Google Scholar] [CrossRef]
- Gignac, P.M. Biomechanics and the ontogeny of feeding in the American Alligator (Alligator mississippiensis): Reconciling factors contributing to the intraspecific niche differentiation in a large-bodied vertebrate. Ph.D. Thesis, Dissertation, Florida State University, Tallahassee, FL, USA, 2010. [Google Scholar]
- Piras, P.; Buscalioni, A.D.; Teresi, L.; Raia, P.; Sansalone, G.; Kotsakis, T.; Cubo, J. Morphological integration and functional modularity in the crocodilian skull. Integr. Zool. 2014, 9, 481–516. [Google Scholar] [CrossRef] [PubMed]
- Brantley, C.G. Food habits of juvenile and sub-adult American Alligators (Alligator mississippiens) in southeastern Louisiana. M.Sc. Thesis, Southeastern Louisiana University, Hammond, LA, USA, 1989. [Google Scholar]
- Pooley, A.C. Food and feeding habits. In Crocodiles and Alligators; Garner, R., Ed.; Facts on File: New York, NY, USA, 1989; pp. 76–91. [Google Scholar]
- Elsey, R.M.; McNease, L.; Joanen, T.; Kinler, N. Food habits of native wild and farm-released juvenile alligators. In Proceedings of the Annual Conference of the Southeastern Association of Fish and Wildlife Agencies, Corpus Christi, TX, USA, 25–28 October 1992; Volume 46, pp. 57–66.
- Cleuren, J.; de Vree, F. Kinematics of the jaw and hyolingual apparatus during feeding in Caiman crocodiles. J. Morphol. 1992, 212, 141–154. [Google Scholar] [CrossRef]
- Fish, F.E.; Bostic, S.A.; Nicastro, A.J.; Beneski, J.T. Death roll of the Alligator: Mechanics of twist feeding in water. J. Exp. Biol. 2007, 210, 2811–2818. [Google Scholar] [CrossRef] [PubMed]
- Erickson, G.M.; Lappin, A.K.; Parker, T.; Vliet, K.A. Comparison of bite-force performance between long-term captive and wild American alligators (Alligator mississippiensis). J. Zool. Lond. 2004, 262, 21–28. [Google Scholar] [CrossRef]
- Neill, W.T. The Last of the Ruling Reptiles: Alligators, Crocodiles, and Their Kin; Columbia University Press: New York, NY, USA, 1971; p. 498. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Chapter 3. Hypothesis testing. In Experimental Design and Data Analysis for Biologists; Quinn, G.P., Keough, M.J, Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 49–50. [Google Scholar]
- Milnes, M.R.; Woodward, A.R.; Rooney, A.A.; Guillette, L.J., Jr. Plasma steroid concentrations in relation to size and age in juvenile alligators from two Florida lakes. Comp. Biochem. Physiol. Part A 2002, 131, 923–930. [Google Scholar] [CrossRef]
- O’Reilly, J.C.; Lindstedt, S.L.; Nishikawa, K.C. The scaling of feeding kinematics in toads (Anura: Bufonidae). Am. Zool. 1993, 33, 147A. [Google Scholar]
- Hill, A.V. The dimensions of animals and their muscular dynamics. Sci. Prog. Lond. 1950, 38, 209–230. [Google Scholar]
- Busbey, A.B. Form and function of the feeding apparatus of Alligator mississippiensis. J. Morphol. 1989, 202, 99–127. [Google Scholar] [CrossRef] [PubMed]
- Reilly, S.M. The ontogeny of aquatic feeding behavior in Salamandra salamandra: Stereotypy and isometry in feeding kinematics. J. Exp. Biol. 1995, 198, 701–708. [Google Scholar] [PubMed]
- Robinson, M.P.; Motta, P.J. Patterns of growth and the effects of scale on the feeding kinematics of the nurse shark (Ginglymostoma cirratum). J. Zool. Lond. 2002, 256, 449–462. [Google Scholar] [CrossRef]
- Thorbjarnarson, J.B. Fishing behavior of spectacled caiman in the Venezuelan Llanos. Copeia 1993, 4, 1166–1171. [Google Scholar] [CrossRef]
- Nifong, J.C.; Nifong, R.L.; Silliman, B.R.; Lowers, R.H.; Guillette, L.J., Jr.; Ferguson, J.M.; Welsh, M.; Abernathy, K.; Marshall, G. Animal-borne imaging reveals novel insights into the foraging behaviors and diel activity of a large-bodied apex predator, the American alligator (Alligator mississippiensis). PLoS One 2014, 9, 1–11. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerfoot, J.R., Jr.; Fern, M.P.; Elsey, R.M. Scaling the Feeding Mechanism of Captive Alligator mississippiensis from Hatchling to Juvenile. Biology 2014, 3, 724-738. https://doi.org/10.3390/biology3040724
Kerfoot JR Jr., Fern MP, Elsey RM. Scaling the Feeding Mechanism of Captive Alligator mississippiensis from Hatchling to Juvenile. Biology. 2014; 3(4):724-738. https://doi.org/10.3390/biology3040724
Chicago/Turabian StyleKerfoot, James R., Jr., Micah P. Fern, and Ruth M. Elsey. 2014. "Scaling the Feeding Mechanism of Captive Alligator mississippiensis from Hatchling to Juvenile" Biology 3, no. 4: 724-738. https://doi.org/10.3390/biology3040724
APA StyleKerfoot, J. R., Jr., Fern, M. P., & Elsey, R. M. (2014). Scaling the Feeding Mechanism of Captive Alligator mississippiensis from Hatchling to Juvenile. Biology, 3(4), 724-738. https://doi.org/10.3390/biology3040724