Pre- and Postsynaptic Role of Dopamine D2 Receptor DD2R in Drosophila Olfactory Associative Learning

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Strains
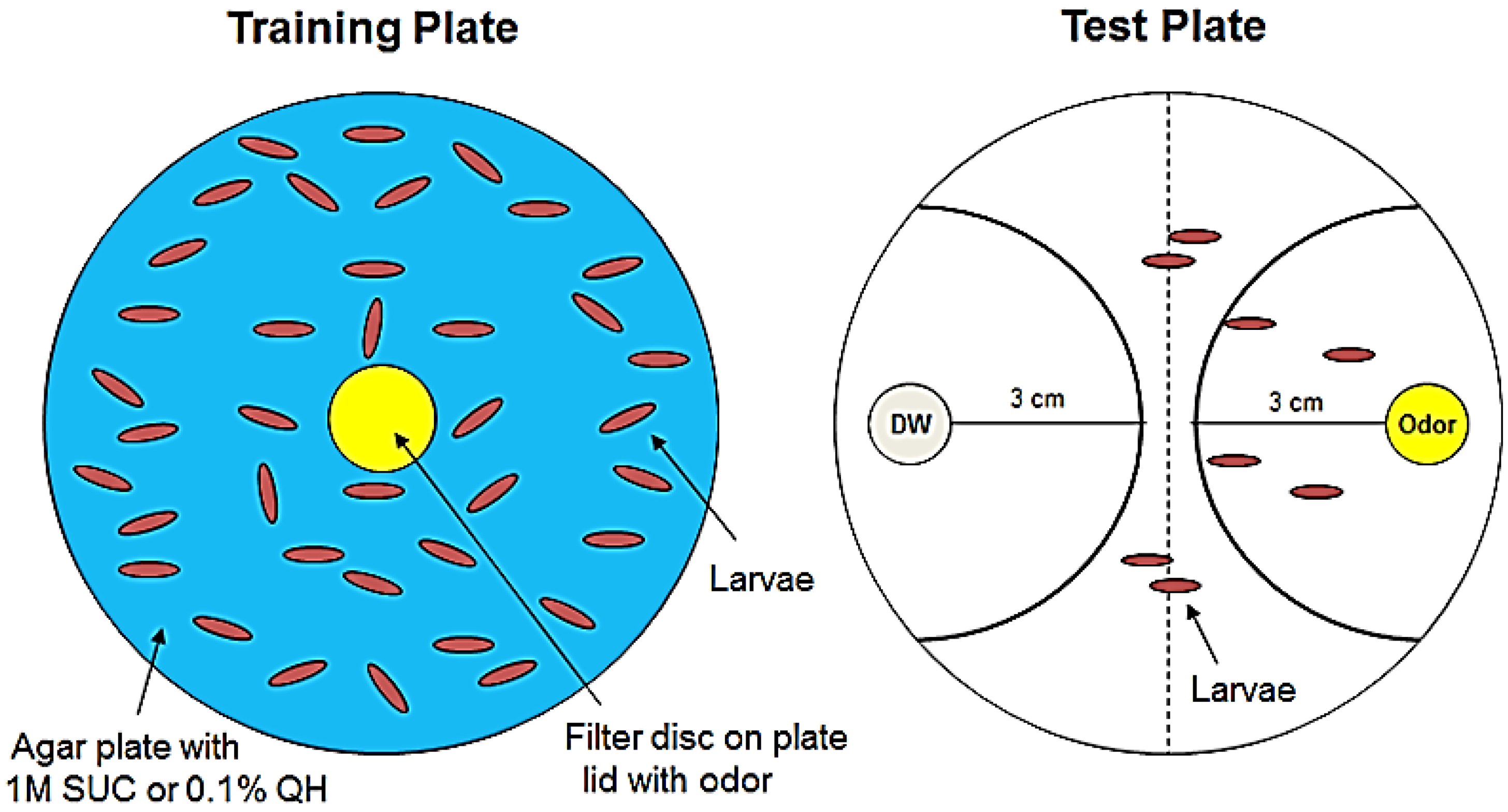
2.2. Larval Olfactory Learning
2.3. Naïve Olfactory Test

2.5. Locomotion Assay
3. Results
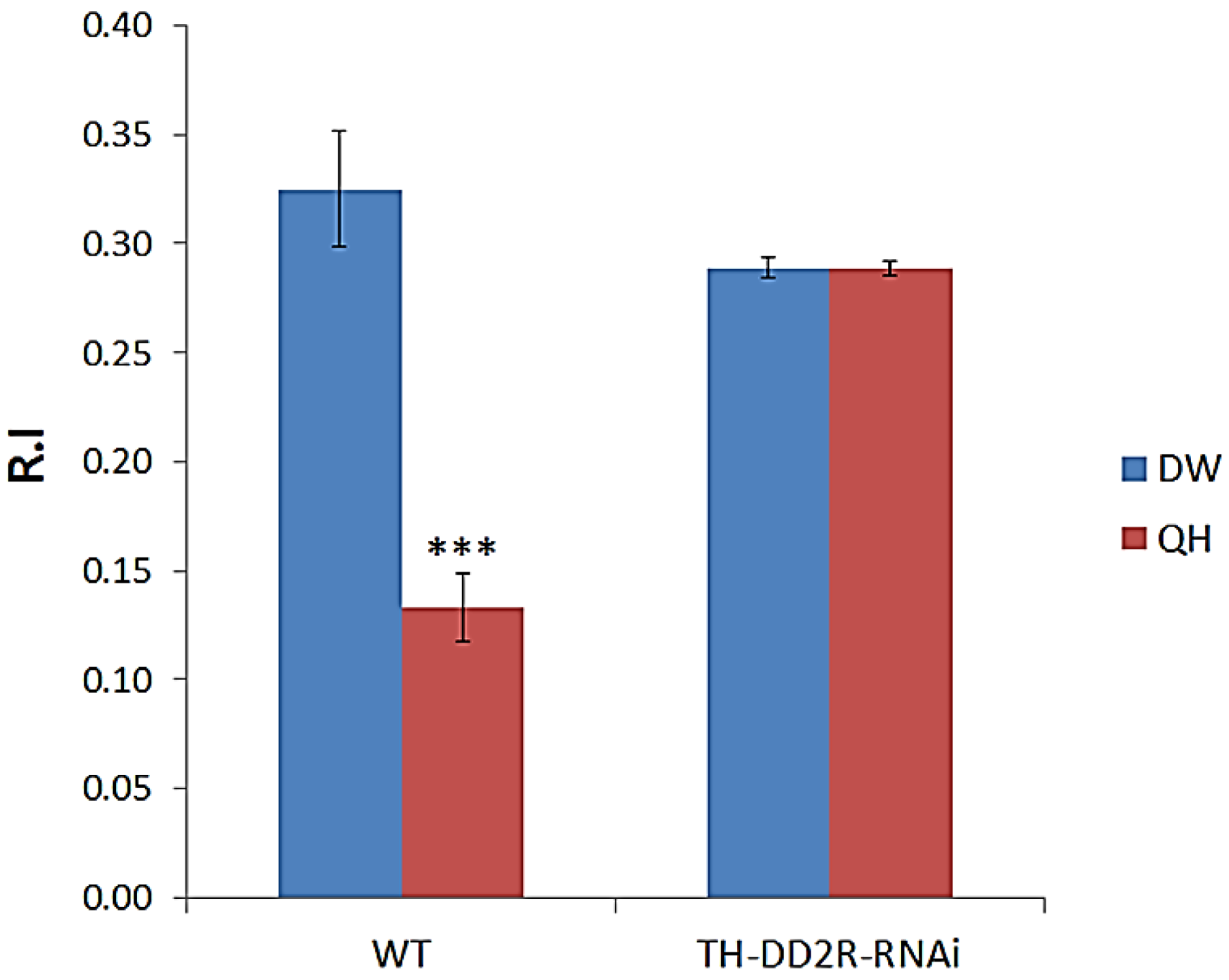
3.1. Aversive Olfactory Learning is Impaired by Down-Regulation of Drosophila DD2R in Dopaminergic Neurons
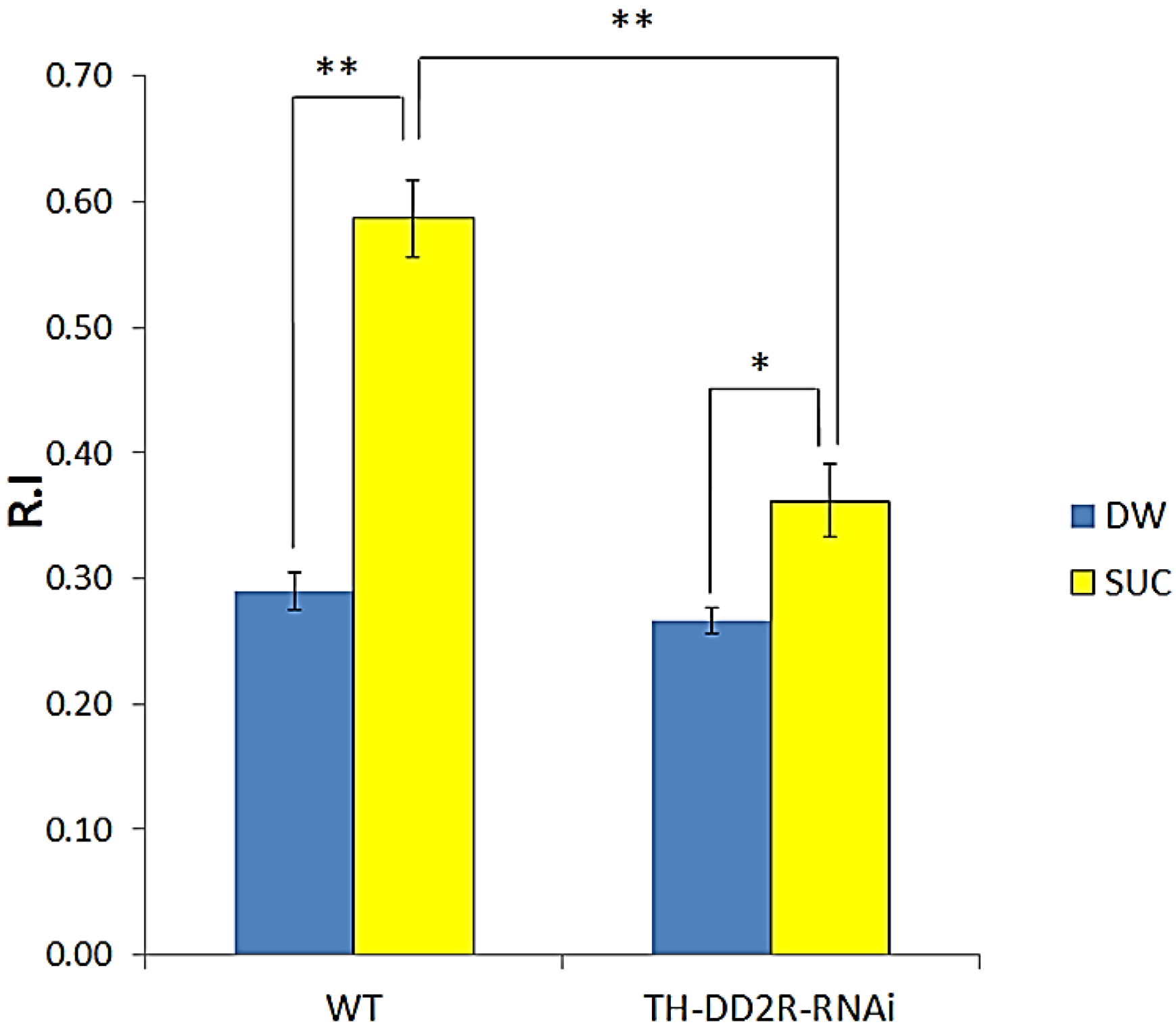
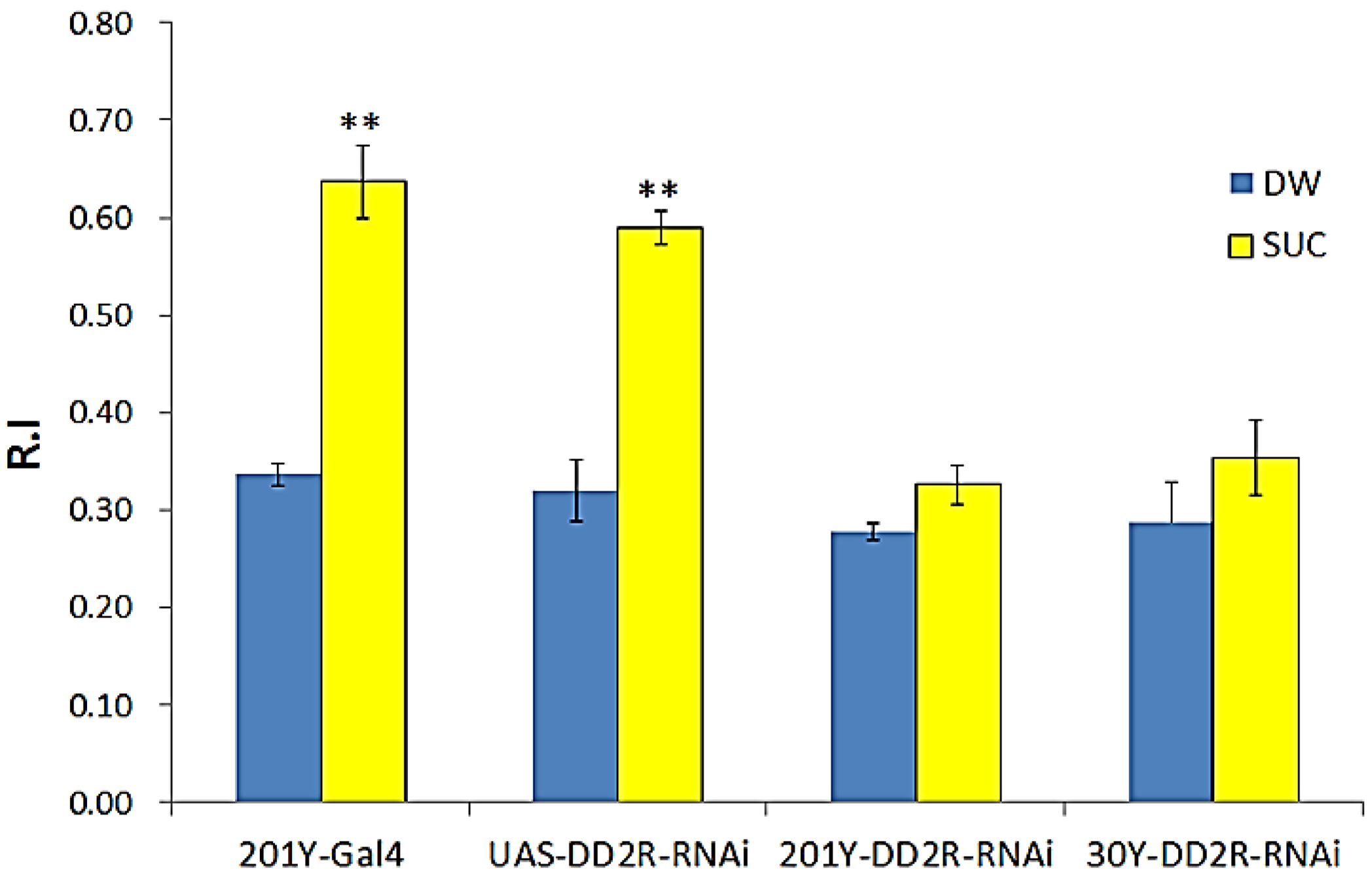
3.2. Appetitive Olfactory Learning is Impaired by Down-Regulation of Drosophila DD2R in Dopaminergic Neurons


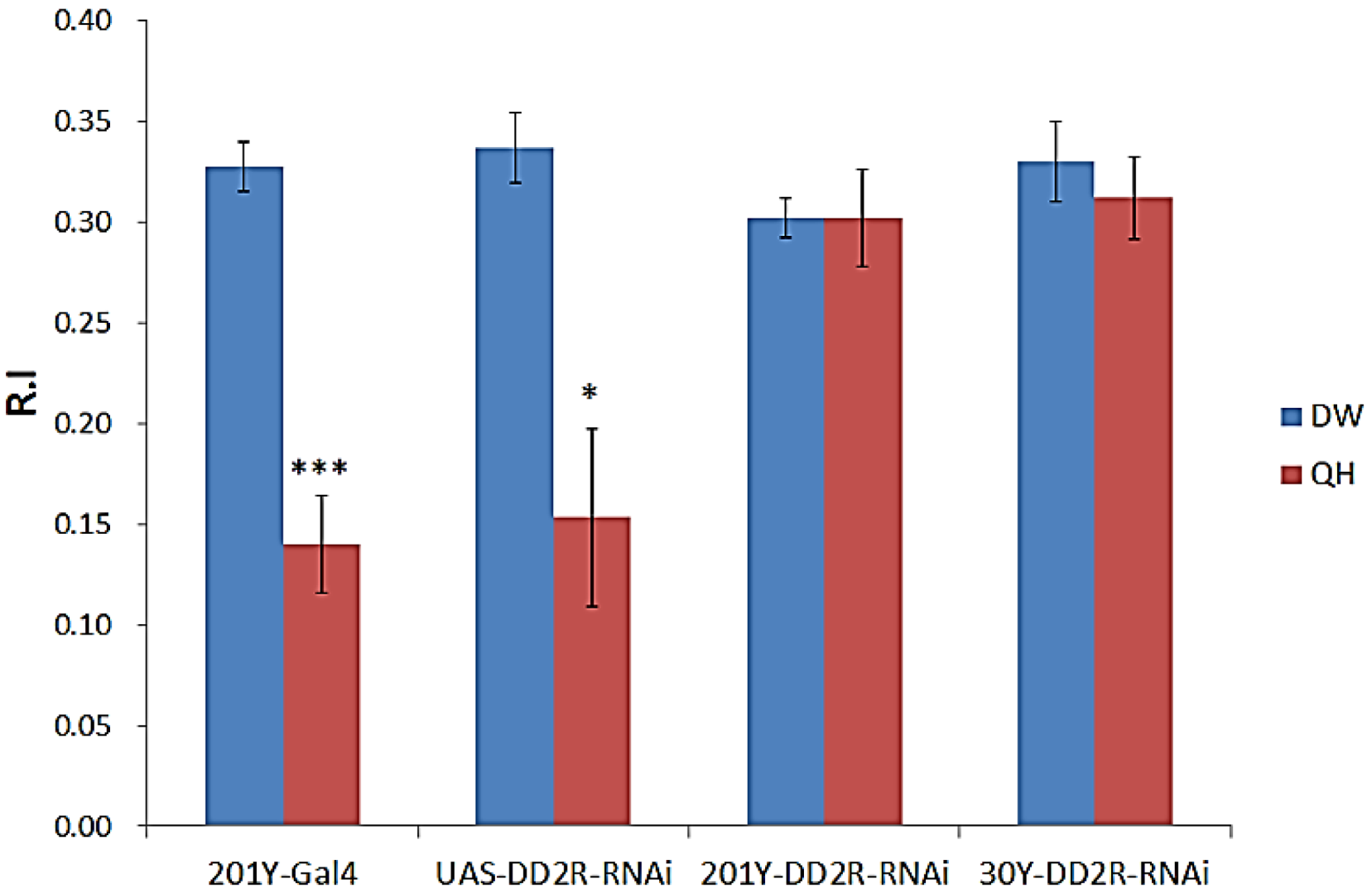
3.3. Aversive Olfactory Learning is Impaired by Down-Regulation of Drosophila DD2R in Mushroom Body (MB) Neurons

3.4. Appetitive Olfactory Learning is Impaired by Down-Regulation of Drosophila DD2R in Mushroom Body Neurons

3.5. Sensory and Motor Functions Are Not Affected by Expression of DD2R-RNAi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | TH-DD2R-RNAi | 201Y-DD2R-RNAi | 30Y-DD2R-RNAi | |
|---|---|---|---|---|
| Naive Olfactory (R.I.) | 0.34 ± 0.02 (n = 9) | 0.31 ± 0.02 (n = 3) | 0.36 ± 0.02 (n = 3) | 0.35 ± 0.054 (n = 4) |
| Naive Gustatory (R.I.) Appetitive | 0.44 ± 0.10 (n = 3) | 0.38 ± 0.05 (n = 3) | 0.65 ± 0.13 (n = 3) | 0.35 ± 0.08 (n = 3) |
| Naive Gustatory (R.I.) Aversive | −0.60 ± 0.07 (n = 4) | −0.58 ± 0.05 (n = 3) | −0.43 ± 0.02 (n = 2) | −0.60 ± 0.08 (n = 3) |
| Locomotion Speed (mm/min) | 95.56 ± 3.89 (n = 36) | 90.53 ± 3.66 (n = 18) | 92.78 ± 3.34 (n = 8) | 99.74 ± 2.38 (n = 5) |
4. Discussion
4.1. Dopamine D2 Receptor DD2R Mediates Olfactory Learning in Drosophila Larvae
4.2. Dopamine Signaling in Drosophila Aversive and Appetitive Learning
4.3. No Change in Locomotion by Expression of DD2R-RNAi in DA or MB Neurons
4.4. Role of Presynaptic DD2R Autoreceptors in Olfactory Learning

4.5. Role of Postsynaptic DD2R in Olfactory Learning
5. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Monastirioti, M. Biogenic amine systems in the fruit fly Drosophila melanogaster. Micro. Res. Tech. 1999, 45, 106–121. [Google Scholar] [CrossRef]
- Neve, K.A.; Seamans, J.K.; Trantham-Davidson, H. Dopamine receptor signaling. J. Recept. Signal Transduct. Res. 2004, 24, 165–205. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Seto, E.S. Dopamine dynamics and signaling in Drosophila: An overview of genes, drugs and behavioral paradigms. Exp. Anim. 2014, 63, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Mustard, J.A.; Beggs, K.T.; Mercer, A.R. Molecular biology of the invertebrate dopamine receptors. Arch. Insect Biochem. Physiol. 2005, 59, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Schwaerzel, M.; Monastirioti, M.; Scholz, H.; Friggi-Grelin, F.; Birman, S.; Heisenberg, M. Dopamine and octopamine differentiate between aversive and appetitive olfactory memory in Drosophila. J. Neurosci. 2003, 23, 10495–10502. [Google Scholar] [PubMed]
- Schroll, C.; Riemensperger, T.; Bucher, D.; Ehmer, J.; Völler, T.; Erbguth, K.; Gerber, B.; Hendel, T.; Nagel, G.; Buchner, E.; et al. Light-induced activation of distinct modulatory neurons triggers appetitive or aversive learning in Drosophila larvae. Curr. Biol. 2006, 16, 1741–1747. [Google Scholar] [CrossRef] [PubMed]
- Honjo, K.; Furukubo-Tokunaga, K. Distinctive neuronal networks and biochemical pathways for appetitive and aversive memory in Drosophila larvae. J. Neurosci. 2009, 29, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Selcho, M.; Pauls, D.; Han, K.A.; Stocker, R.F.; Thum, A.S. The role of dopamine in Drosophila larval classical olfactory conditioning. PLoS One 2009, 4, e5897. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.L. Traces of Drosophila memory. Neuron 2011, 70, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Waddell, S. Reinforcement signaling in Drosophila; dopamine does it all after all. Curr. Opin. Neuro. 2013, 23, 324–329. [Google Scholar] [CrossRef]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure and function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [PubMed]
- Blenau, W.; Baumann, A. Molecular and pharmacological properties of insect biogenic amine receptors: Lessons from Drosophila melanogaster and Apis melifera. Arch. Insect Biochem. Physiol. 2001, 48, 13–38. [Google Scholar] [CrossRef] [PubMed]
- Aghajanian, G.K.; Bunney, B.S. Dopamine “autoreceptors”: Pharmacological characterization by microiontophoretic single cell recording studies. Naunyn Schmiedebergs Arch. Pharmacol. 1977, 297, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chiodo, L.A.; Freeman, A.S.; Bunney, B.S. Dopamine autoreceptor signal transduction and regulation. Psychopharmacology—4th Generation of Progress. Available online: http://www.acnp.org/g4/GN401000020/Default.htm (accessed on 10 October 2014).
- Feuerstein, T.J. Presynaptic receptors for dopamine, histamine, and serotonin. Handb. Exp. Pharmacol. 2008, 184, 289–338. [Google Scholar] [PubMed]
- Gotzes, F.; Balfanz, S.; Baumann, A. Primary structure and functional characterization of a Drosophila dopamine receptor with high homology to human D1/5 receptors. Recept. Chan. 1994, 2, 131–141. [Google Scholar]
- Feng, G.; Hannan, F.; Reale, V.; Hon, Y.Y.; Kousky, C.T.; Evans, P.D.; Hall, L.M. Cloning and functional characterization of a novel dopamine receptor from Drosophila melanogaster. J. Neurosci. 1996, 16, 3925–3933. [Google Scholar] [PubMed]
- Han, K.A.; Millar, N.S.; Grotewiel, M.S.; Davis, R.L. DAMB, a novel dopamine receptor expressed specifically in Drosophila mushroom bodies. Neuron 1996, 16, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Hearn, M.G.; Ren, Y.; McBride, E.W.; Reveillaud, I.; Beinborn, M.; Kopin, A.S. A Drosophila dopamine 2-like receptor: Molecular characterization and identification of multiple alternatively spliced variants. Proc. Natl. Acad. Sci. USA 2002, 99, 14554–14559. [Google Scholar] [CrossRef] [PubMed]
- Vickrey, T.L.; Venton, B.J. Drosophila dopamine2-like receptors function as autoreceptors. ACS Chem. Neurosci. 2011, 2, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Wiemerslage, L.; Schultz, B.J.; Ganguly, A.; Lee, D. Selective degeneration of dopaminergic neurons by MPP (+) and its rescue by D2 autoreceptors in Drosophila primary culture. J. Neurochem. 2013, 126, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Lee, H.G.; Han, K.A. D1 dopamine receptor dDA1 is required in the mushroom body neurons for aversive and appetitive learning in Drosophila. J. Neurosci. 2007, 27, 7640–7647. [Google Scholar] [CrossRef] [PubMed]
- Seugnet, L.; Suzuki, Y.; Vine, L.; Gottschalk, L.; Shaw, P.J. D1 receptor activation in the mushroom bodies rescues sleep-loss-induced learning impairments in Drosophila. Curr. Biol. 2008, 18, 1110–1117. [Google Scholar] [CrossRef]
- Honjo, K.; Furukubo-Tokunaga, K. Induction of cAMP response element-binding protein-dependent medium-term memory by appetitive gustatory reinforcement in Drosophila larvae. J. Neurosci. 2005, 25, 7905–7913. [Google Scholar] [CrossRef] [PubMed]
- Varga, S.J.; Qi, C.; Podolsky, E.; Lee, D. A new Drosophila model to study the interaction between genetic and environmental factors in Parkinson’s disease. Brain Res. 2014, 1583, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [PubMed]
- Friggi-Grelin, F.; Coulom, H.; Meller, M.; Gomez, D.; Hirsh, J.; Birman, S. Targeted gene expression in Drosophila dopaminergic cells using regulatory sequences from tyrosine hydroxylase. J. Neurobiol. 2003, 54, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Draper, I.; Kurshan, P.T.; McBride, E.; Jackson, F.R.; Kopin, A.S. Locomotor activity is regulated by D2-like receptors in Drosophila: An anatomic and functional analysis. Dev. Neurobiol. 2007, 67, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Grübel, K.; Busch, S.; Friedrich, A.B.; Siwanowicz, I.; Tanimoto, H. The mushroom body of adult Drosophila characterized by GAL4 drivers. J. Neurogen. 2009, 23, 156–172. [Google Scholar] [CrossRef]
- Pauls, D.; Selcho, M.; Gendre, N.; Stocker, R.F.; Thum, A.S. Drosophila larvae establish appetitive olfactory memories via mushroom body neurons of embryonic origin. J. Neurosci. 2010, 30, 10655–10666. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Lee, D.; Ohio University, Athens, OH, USA. Unpublished data. 2014.
- Alekseyenko, O.V.; Chan, Y.B.; Li, R.; Kravitz, E.A. Single dopaminergic neurons that modulate aggression in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 6151–6156. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Lee, H.G.; Seong, C.S.; Han, K.A. Expression of a D1 dopamine receptor dDA1/DmDOP1 in the central nervous system of Drosophila melanogaster. Gene Expr. Patterns 2003, 3, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yin, Y.; Lu, H.; Guo, A. Increased dopaminergic signaling impairs aversive olfactory memory retention in Drosophila. Biochem. Biophys. Res. Comm. 2008, 370, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, R.A.; Roche, K.W. Long-term potentiation: Peeling the onion. Neuropharmacology 2013, 74, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Kandel, K.R.; Dudai, Y.; Mayford, M.R. The molecular and systems biology of memory. Cell 2014, 157, 163–186. [Google Scholar] [CrossRef] [PubMed]
- Barco, A.; Bailey, C.H.; Kandel, E.R. Common molecular mechanisms in explicit and implicit memory. J. Neurochem. 2006, 97, 1520–1533. [Google Scholar] [CrossRef]
- Lee, D.; O’Dowd, D.K. cAMP-dependent plasticity at excitatory cholinergic synapses in Drosophila neurons: Alterations in the memory mutant dunce. J. Neurosci. 2000, 20, 2104–2111. [Google Scholar] [PubMed]
- Gervasi, N.; Tchénio, P.; Preat, T. PKA dynamics in a Drosophila learning center: Coincidence detection by rutabaga adenylyl cyclase and spatial regulation by dunce phosphodiesterase. Neuron 2010, 65, 516–529. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, C.; Lee, D. Pre- and Postsynaptic Role of Dopamine D2 Receptor DD2R in Drosophila Olfactory Associative Learning. Biology 2014, 3, 831-845. https://doi.org/10.3390/biology3040831
Qi C, Lee D. Pre- and Postsynaptic Role of Dopamine D2 Receptor DD2R in Drosophila Olfactory Associative Learning. Biology. 2014; 3(4):831-845. https://doi.org/10.3390/biology3040831
Chicago/Turabian StyleQi, Cheng, and Daewoo Lee. 2014. "Pre- and Postsynaptic Role of Dopamine D2 Receptor DD2R in Drosophila Olfactory Associative Learning" Biology 3, no. 4: 831-845. https://doi.org/10.3390/biology3040831
APA StyleQi, C., & Lee, D. (2014). Pre- and Postsynaptic Role of Dopamine D2 Receptor DD2R in Drosophila Olfactory Associative Learning. Biology, 3(4), 831-845. https://doi.org/10.3390/biology3040831