
Stearoyl-CoA Desaturase-1: Is It the Link between Sulfur Amino Acids and Lipid Metabolism?

Abstract
:1. Introduction


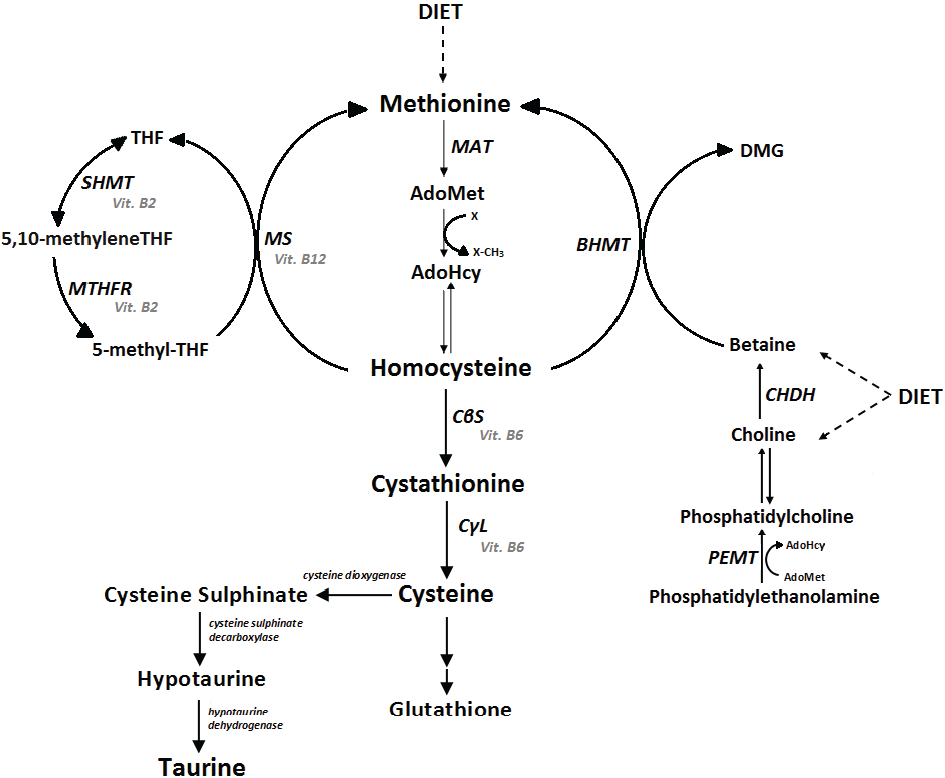{kind=link}
| Etiology | Characteristics |
|---|---|
| Vitamin B12 deficiency | Vitamin B12 is used in homocysteine remethylation. The groups at higher risk for this deficiency are vegetarians and the elderly, with the deficiency rates reaching 20% for the latter. Deficient absorption is likely the main etiological factor in elderly individuals because intrinsic-factor deficiency, which is fundamental for the absorption of B12, is common in this age group [62]. |
| Folate deficiency | Folate is also involved in the remethylation of homocysteine, and its deficiency is more common in individuals with increased needs, such as pregnant women and alcoholics. In addition, a frequent polymorphism in the MTHFR gene (c.677C > T) makes the enzyme thermolabile and decreases enzymatic activity in homozygotes to 70%. If their folate status is low to normal, homocysteine can increase dramatically [63]. |
| Medicines | The chronic use of certain drugs, such as methotrexate, theophylline, phenytoin, carbamazepine and valproic acid, is known to increase homocysteine levels [64,65]. |
| Other causes | Additional causes include chronic renal disease, cancer, hypothyroidism, liver failure, smoking, alcohol and coffee intake, age and hormonal disorders [64,65]. |
2. Sulfur Amino Acids and Lipid Metabolism
| Amino Acid | Effect on Lipid Metabolism | Effect on SCD-1 Regulators |
|---|---|---|
| Methionine |
|
|
| Homocysteine |
| |
| Cysteine |
|
|
| Taurine |
|
3. SCD-1: Localization, Function and Regulation
4. Evidence that Sulfur Amino Acids Influence SCD-1 Expression
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Castro, R.; Rivera, I.; Blom, H.J.; Jakobs, C.; Tavares de Almeida, I. Homocysteine metabolism, hyperhomocysteinaemia and vascular disease: An overview. J. Inherit. Metab. Dis. 2006, 29, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: An overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [PubMed]
- Blom, H.J.; Smulders, Y. Overview of homocysteine and folate metabolism. With special references to cardiovascular disease and neural tube defects. J. Inherit. Metab. Dis. 2011, 34, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Guo, J.X.; Chang, P. The effect of taurine on cholesterol metabolism. Mol. Nutr. Food. Res. 2012, 56, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Oh da, H.; Kim, J.Y.; Lee, B.G.; You, J.S.; Chang, K.J.; Chung, H.J.; Yoo, M.C.; Yang, H.I.; Kang, J.H.; et al. Taurine ameliorates hyperglycemia and dyslipidemia by reducing insulin resistance and leptin level in Otsuka Long-Evans Tokushima fatty (OLETF) rats with long-term diabetes. Exp. Mol. Med. 2012, 44, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Graham, I.M.; Palma Reis, R.; Sales Luis, A.; Smith, A.D.; Refsum, H. The association of fasting plasma sulfur-containing compounds with BMI, serum lipids and apolipoproteins. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- El-Khairy, L.; Ueland, P.M.; Refsum, H.; Graham, I.M.; Vollset, S.E.; European Concerted Action Project. Plasma total cysteine as a risk factor for vascular disease: The European Concerted Action Project. Circulation 2001, 103, 2544–2549. [Google Scholar] [CrossRef] [PubMed]
- Fowler, B. Disorders of homocysteine metabolism. J. Inherit. Metab. Dis. 1997, 20, 270–285. [Google Scholar] [CrossRef] [PubMed]
- Stipanuk, M.H.; Ueki, I. Dealing with methionine/homocysteine sulfur: Cysteine metabolism to taurine and inorganic sulfur. J. Inherit. Metab. Dis. 2011, 34, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, J.D. The metabolism of homocysteine: Pathways and regulation. Eur. J. Pediatr. 1998, 157, 40–44. [Google Scholar] [CrossRef]
- Booth, G.L.; Wang, E.E. Preventive health care, 2000 update: Screening and management of hyperhomocysteinemia for the prevention of coronary artery disease events. The Canadian Task Force on Preventive Health Care. CMAJ 2000, 163, 21–29. [Google Scholar] [PubMed]
- Selhub, J. The many facets of hyperhomocysteinemia: Studies from the Framingham cohorts. J. Nutr. 2006, 136, 1726S–1730S. [Google Scholar] [PubMed]
- Mudd, S.H.; Levy, H.L.; Kraus, J.P. Disorders of transsulfuration. In The Metabolic and Molecular Bases of Inherited Disease; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Childs, B., Kinzler, K.W., Vogelstein, B., Eds.; McGraw Hill Inc.: New York, NY, USA, 2001; pp. 2007–2056. [Google Scholar]
- Wilcken, B. Disorders of Sulfur Amino Acids. In Physician’s Guide to the Treatment and Follow-Up of Metabolic Diseases; Blau, N., Hoffman, G.F., Leonard, J., Clarke, J.T.R., Eds.; Springer-Verlag Berlin Heidelberg: Berlin, Germany, 2006; pp. 105–115. [Google Scholar]
- Brenton, D.P.; Dow, C.J.; James, J.I.; Hay, R.L.; Wynne-Davies, R. Homocystinuria and Marfan’s syndrome—A comparison. J. Bone Jt. Surg. Br. 1972, 54, 277–298. [Google Scholar]
- Rosenblatt, D.S.; Fowler, B. Disorders of cobalamin and folate transport and metabolism. In Inborn Metabolic Diseases. Diagnosis and Treatment; Fernandes, J., Saudubray, J.M., van den Berghe, G., Walter, J.H., Eds.; Hardcover; Springer: Berlin, Germany, 2006; pp. 341–356. [Google Scholar]
- Orphanet. Prevalence of rare diseases: Bibliographic data. Orphanet Rep. Ser. 2014, 1, 20. [Google Scholar]
- Hirche, F.; Schroder, A.; Knoth, B.; Stangl, G.I.; Eder, K. Effect of dietary methionine on plasma and liver cholesterol concentrations in rats and expression of hepatic genes involved in cholesterol metabolism. Br. J. Nutr. 2006, 95, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Moat, S.J.; Bonham, J.R.; Allen, J.C.; Powers, H.J.; McDowell, I.F. Decreased circulating plasma lipids in patients with homocystinuria. J. Inherit. Metab. Dis. 1999, 22, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Tan, H.; Hui, R.; Li, Z.; Jiang, X.; Gaubatz, J.; Yang, F.; Durante, W.; Chan, L.; Schafer, A.I.; et al. Hyperhomocysteinemia decreases circulating high-density lipoprotein by inhibiting apolipoprotein A-I Protein synthesis and enhancing HDL cholesterol clearance. Circ. Res. 2006, 99, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Poloni, S.; Mendes, R.H.; Belló-Klein, A.; Schwartz, I.V. Is lipid metabolism altered in classical homocystinuria? Mol. Genet. Metab. 2012, 106, 382–383. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Osada, J.; Aratani, Y.; Kluckman, K.; Reddick, R.; Malinow, M.R.; Maeda, N. Mice deficient in cystathionine beta-synthase: Animal models for mild and severe homocyst(e)inemia. Proc. Natl. Acad. Sci. USA 1995, 92, 1585–1589. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Herrmann, W. Homocysteine and lipids: S-adenosyl methionine as a key intermediate. FEBS Lett. 2009, 583, 1215–1225. [Google Scholar] [CrossRef] [PubMed]
- Namekata, K.; Enokido, Y.; Ishii, I.; Nagai, Y.; Harada, T.; Kimura, H. Abnormal lipid metabolism in cystathionine beta-synthase-deficient mice, an animal model for hyperhomocysteinemia. J. Biol. Chem. 2004, 279, 52961–52969. [Google Scholar] [CrossRef] [PubMed]
- Nuño-Ayala, M.; Guillén, N.; Navarro, M.A.; Lou-Bonafonte, J.M.; Arnal, C.; Gascón, S.; Barranquero, C.; Godino, J.; Royo-Cañas, M.; Sarría, A.J.; et al. Cysteinemia, rather than homocysteinemia, is associated with plasma apolipoprotein A-I levels in hyperhomocysteinemia: Lipid metabolism in cystathionine beta-synthase deficiency. Atherosclerosis 2010, 212, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Karaplis, A.C.; Ackerman, S.L.; Pogribny, I.P.; Melnyk, S.; Lussier-Cacan, S.; Chen, M.F.; Pai, A.; John, S.W.; Smith, R.S.; et al. Mice deficient in methylenetetrahydrofolate reductase exhibit hyperhomocysteinemia and decreased methylation capacity, with neuropathology and aortic lipid deposition. Hum. Mol. Genet. 2001, 10, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Mikael, L.G.; Genest, J., Jr.; Rozen, R. Elevated homocysteine reduces apolipoprotein A-I expression in hyperhomocysteinemic mice and in males with coronary artery disease. Circ. Res. 2006, 98, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Fielding, B.A. Stearoyl-Co desaturase: Rogue or innocent bystander? Prog. Lipid Res. 2013, 52, 15–42. [Google Scholar] [CrossRef] [PubMed]
- Paton, C.M.; Ntambi, J.M. Biochemical and physiological function of stearoyl-CoA desaturase. Am. J. Physiol. Endocrinol. Metab. 2009, 297, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Kim, Y.C.; Ntambi, J.M. A lipogenic diet in mice with a disruption of the stearoyl-CoA desaturase 1 gene reveals a stringent requirement of endogenous monounsaturated fatty acids for triglyceride synthesis. J. Lipid Res. 2001, 42, 1018–1024. [Google Scholar] [PubMed]
- Ozols, J. Degradation of hepatic stearyl CoA delta 9-desaturase. Mol. Biol. Cell 1997, 8, 2281–2290. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Friedman, J.M. Leptin and the control of metabolism: Role for stearoyl-CoA desaturase-1 (SCD-1). J. Nutr. 2004, 134, 2455S–2463S. [Google Scholar] [PubMed]
- Biddinger, S.B.; Miyazaki, M.; Boucher, J.; Ntambi, J.M.; Kahn, C.R. Leptin suppresses stearoyl-CoA desaturase 1 by mechanisms independent of insulin and sterol regulatory element-binding protein-1c. Diabetes 2006, 55, 2032–2041. [Google Scholar] [CrossRef] [PubMed]
- Mutch, D.M.; Grigorov, M.; Berger, A.; Fay, L.B.; Roberts, M.A.; Watkins, S.M.; Williamson, G.; German, J.B. An integrative metabolism approach identifies stearoyl-CoA desaturase as a target for an arachidonate-enriched diet. FASEB J. 2005, 19, 599–601. [Google Scholar] [CrossRef] [PubMed]
- Vessby, B.; Gustafsson, I.B.; Tengblad, S.; Berglund, L. Indices of fatty acid desaturase activity in healthy human subjects: Effects of different types of dietary fat. Br. J. Nutr. 2013, 110, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Vinknes, K.J.; Elshorbagy, A.K.; Nurk, E.; Drevon, C.A.; Gjesdal, C.G.; Tell, G.S.; Nygard, O.; Vollset, S.E.; Refsum, H. Plasma Stearoyl-CoA Desaturase Indices: Association with Lifestyle, Diet, and Body Composition. Obesity 2013, 21, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Vinknes, K.J.; Elshorbagy, A.K.; Drevon, C.A.; Nurk, E.; Tell, G.S.; Nygård, O.; Vollset, S.E.; Refsum, H. Associations Between Plasma Polyunsaturated Fatty Acids, Plasma Stearoyl-CoA Desaturase Indices and Body Fat. Obesity 2013, 21, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Ntambi, J.M.; Miyazaki, M.; Stoehr, J.P.; Lan, H.; Kendziorski, C.M.; Yandell, B.S.; Song, Y.; Cohen, P.; Friedman, J.M.; Attie, A.D.; et al. Loss of stearoyl-CoA desaturase-1 function protects mice against adiposity. Proc. Natl. Acad. Sci. USA 2002, 99, 11482–11486. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Flowers, M.T.; Sampath, H.; Chu, K.; Otzelberger, C.; Liu, X.; Ntambi, J.M. Hepatic stearoyl-CoA desaturase-1 deficiency protects mice from carbohydrate-induced adiposity and hepatic steatosis. Cell Metab. 2007, 6, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Flowers, M.T.; Ade, L.; Strable, M.S.; Ntambi, J.M. Combined deletion of SCD1 from adipose tissue and liver does not protect mice from obesity. J. Lipid Res. 2012, 53, 1646–1653. [Google Scholar] [CrossRef] [PubMed]
- Sampath, H.; Flowers, M.T.; Liu, X.; Paton, C.M.; Sullivan, R.; Chu, K.; Zhao, M.; Ntambi, J.M. Skin-specific deletion of stearoyl-CoA desaturase-1 alters skin lipid composition and protects mice from high fat diet-induced obesity. J. Biol. Chem. 2009, 284, 19961–19973. [Google Scholar] [CrossRef] [PubMed]
- Warensjö, E.; Ohrvall, M.; Vessby, B. Fatty acid composition and estimated desaturase activities are associated with obesity and lifestyle variables in men and women. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Paillard, F.; Catheline, D.; Duff, F.L.; Bouriel, M.; Deugnier, Y.; Pouchard, M.; Daubert, J.C.; Legrand, P. Plasma palmitoleic acid, a product of stearoyl-CoA desaturase activity, is an independent marker of triglyceridemia and abdominal adiposity. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Warensjö, E.; Risérus, U.; Vessby, B. Fatty acid composition of serum lipids predicts the development of the metabolic syndrome in men. Diabetologia 2005, 48, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, Y.; Ågren, J.; Uusitupa, M.; Cederberg, H.; Vangipurapu, J.; Stančáková, A.; Schwab, U.; Kuusisto, J.; Laakso, M. Association of erythrocyte membrane fatty acids with changes in glycemia and risk of type 2 diabetes. Am. J. Clin. Nutr. 2014, 99, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K. Body composition in gene knockouts of sulfur amino acid-metabolizing enzymes. Mamm. Genome 2014, 25, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kruger, W.D. Cystathionine beta-synthase deficiency causes fat loss in mice. PLoS ONE 2011, 6, e27598. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Refsum, H.; Smith, A.D.; Mattocks, D.A.; Perrone, C.E. Sulfur amino acids in methionine-restricted rats: Hyperhomocysteinemia. Nutrition 2010, 26, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Mattocks, D.A.; Plummer, J.D.; Smith, A.D.; Drevon, C.A.; Refsum, H.; Perrone, C.E. Cysteine supplementation reverses methionine restriction effects on rat adiposity: Significance of stearoyl-coenzyme A desaturase. J. Lipid Res. 2011, 52, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Mattocks, D.A.; Plummer, J.D.; Orentreich, D.S.; Orentreich, N.; Refsum, H.; Perrone, C.E. Effect of taurine and N-acetylcysteine on methionine restriction-mediated adiposity resistance. Metabolism 2013, 62, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Dieter, M.Z.; Chen, Y.; Shertzer, H.G.; Nebert, D.W.; Dalton, T.P. Initial characterization of the glutamatecysteine ligase modifier subunit Gclm(−/−) knockout mouse. Novel modelsystem for a severely compromised oxidative stress response. J. Biol. Chem. 2002, 277, 49446–49452. [Google Scholar] [CrossRef] [PubMed]
- Kendig, E.L.; Chen, Y.; Krishan, M.; Johansson, E.; Schneider, S.N.; Genter, M.B.; Nebert, D.W.; Shertzer, H.G. Lipid metabolism and body composition in Gclm(−/−) mice. Toxicol. Appl. Pharmacol. 2011, 257, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H. Metabolic crosstalk between choline/1-carbon metabolism and energy homeostasis. Clin. Chem. Lab. Med. 2013, 51, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.W.; Mehedint, M.G.; Garrow, T.A.; Zeisel, S.H. Deletion of betaine-homocysteine S-methyltransferase in mice perturbs choline and 1-carbon metabolism, resulting in fatty liver and hepatocellular carcinomas. J. Biol. Chem. 2011, 286, 36258–36267. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.W.; Ellis, J.M.; Coleman, R.A.; Zeisel, S.H. Mouse betaine-homocysteine S-methyltransferase deficiency reduces body fat via increasing energy expenditure and impairing lipid synthesis and enhancing glucose oxidation in white adipose tissue. J. Biol. Chem. 2012, 287, 16187–16198. [Google Scholar] [CrossRef] [PubMed]
- Rizki, G.; Arnaboldi, L.; Gabrielli, B.; Yan, J.; Lee, G.S.; Ng, R.K.; Turner, S.M.; Badger, T.M.; Pitas, R.E.; Maher, J.J.; et al. Mice fed a lipogenic methionine-choline-deficient diet develop hypermetabolism coincident with hepatic suppression of SCD-1. J. Lipid Res. 2006, 47, 2280–2290. [Google Scholar] [CrossRef] [PubMed]
- Vinknes, K.J.; Dekker, J.M.; Drevon, C.A.; Refsum, H.; Nurk, E.; Nijpels, G.; Stehouwer, C.D.; Teerlink, T.; Tell, G.S.; Nygård, O.; et al. Plasma sulfur amino acids and stearoyl-CoA desaturase activity in two Caucasian populations. Prostaglandins Leukot. Essent. Fat. Acids 2013, 89, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Nurk, E.; Gjesdal, C.G.; Tell, G.S.; Ueland, P.M.; Nygård, O.; Tverdal, A.; Vollset, S.E.; Refsum, H. Homocysteine, cysteine, and body composition in the Hordaland Homocysteine Study: Does cysteine link amino acid and lipid metabolism? Am. J. Clin. Nutr. 2008, 88, 738–746. [Google Scholar] [PubMed]
- Elshorbagy, A.K.; Refsum, H.; Smith, A.D.; Graham, I.M. The association of plasma cysteine and gamma-glutamyltransferase with BMI and obesity. Obesity 2009, 17, 435–440. [Google Scholar]
- Elshorbagy, A.K.; Kozich, V.; Smith, A.D.; Refsum, H. Cysteine and obesity: Consistency of the evidence across epidemiologic, animal and cellular studies. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Poloni, S.; Leistner-Segal, S.; Bandeira, I.C.; D’Almeida, V.; de Souza, C.F.; Spritzer, P.M.; Castro, K.; Tonon, T.; Nalin, T.; Imbard, A.; et al. Body composition in patients with classical homocystinuria: Body mass relates to homocysteine and choline metabolism. Gene 2014, 546, 443–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan-Harshman, M.; Aldoori, W. Vitamin B12 and health. Can. Fam. Physician 2008, 54, 536–541. [Google Scholar] [PubMed]
- Stover, P.J. Physiology of folate and vitamin B12 in health and disease. Nutr. Rev. 2004, 62, S3–S12. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.V.; Oudit, G.Y.; Evans, M. Homocysteine, vitamins, and coronary artery disease. Comprehensive review of the literature. Can. Fam. Physician 2000, 46, 2236–2245. [Google Scholar] [PubMed]
- Brustolin, S.; Giugliani, R.; Félix, T.M. Genetics of homocysteine metabolism and associated disorders. Braz. J. Med. Biol. Res. 2010, 43, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Oda, H. Functions of sulfur-containing amino acids in lipid metabolism. J. Nutr. 2006, 136, 1666S–1669S. [Google Scholar] [PubMed]
- Sung, H.J.; Kim, J.; Kim, Y.; Jang, S.W.; Ko, J. N-acetyl cysteine suppresses the foam cell formation that is induced by oxidized low density lipoprotein via regulation of gene expression. Mol. Biol. Rep. 2012, 39, 3001–3007. [Google Scholar] [CrossRef] [PubMed]
- Narin, F.; Atabek, M.E.; Karakukcu, M.; Narin, N.; Kurtoglu, S.; Gumus, H.; Coksevim, B.; Erez, R. The association of plasma homocysteine levels with serum leptin and apolipoprotein B levels in childhood obesity. Ann. Saudi Med. 2005, 25, 209–214. [Google Scholar] [PubMed]
- Basati, G.; Razavi, A.E.; Abdi, S.; Sarrafzedegan, N. Association of plasma leptin, homocysteine and nitric oxide levels with the presence and unstability of coronary artery disease. Biomark. Med. 2014, 8, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Werstuck, G.H.; Lentz, S.R.; Dayal, S.; Hossain, G.S.; Sood, S.K.; Shi, Y.Y.; Zhou, J.; Maeda, N.; Krisans, S.K.; Malinow, M.R.; et al. Homocysteine-induced endoplasmic reticulum stress causes dysregulation of the cholesterol and triglyceride biosynthetic pathways. J. Clin. Investig. 2001, 107, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, L.; Tan, Y.; Wei, J.; Chang, Y.; Jin, T.; Zhu, H. Betaine supplement alleviates hepatic triglyceride accumulation of apolipoprotein E deficient mice via reducing methylation of peroxisomal proliferator-activated receptor alpha promoter. Lipids Health Dis. 2013, 13, 12–34. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. l-Cysteine and hydrogen sulfide increase PIP3 and AMPK/PPARγ expression and decrease ROS and vascular inflammation markers in high glucose treated human U937 monocytes. J. Cell. Biochem. 2013, 114, 2334–2345. [Google Scholar] [CrossRef] [PubMed]
- Bettzieche, A.; Brandsch, C.; Hirche, F.; Eder, K.; Stangl, G.I. L-cysteine down-regulates SREBP-1c-regulated lipogenic enzymes expression via glutathione in HepG2 cells. Ann. Nutr. Metab. 2008, 2, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Church, C.; Valdivia-Garcia, M.; Smith, A.D.; Refsum, H.; Cox, R. Dietary cystine level affects metabolic rate and glycaemic control in adult mice. J. Nutr. Biochem. 2012, 23, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Peng, K.; Su, J.; Huang, Y.; Xu, Y.; Wang, S. Different effects of homocysteine and oxidized low density lipoprotein on methylation status in the promoter region of the estrogen receptor alpha gene. Acta Biochim. Biophys. Sin. 2007, 39, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Hoang, M.H.; Jia, Y.; Jun, H.J.; Lee, J.H.; Hwang, K.Y.; Choi, D.W.; Um, S.J.; Lee, B.Y.; You, S.G.; Lee, S.J.; et al. Taurine is a liver X receptor-α ligand and activates transcription of key genes in the reverse cholesterol transport without inducing hepatic lipogenesis. Mol. Nutr. Food Res. 2012, 56, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Batista, T.M.; da Silva, P.M.; Amaral, A.G.; Ribeiro, R.A.; Boschero, A.C.; Carneiro, E.M. Taurine supplementation restores insulin secretion and reduces ER stress markers in protein-malnourished mice. Adv. Exp. Med. Biol. 2013, 776, 129–139. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poloni, S.; Blom, H.J.; Schwartz, I.V.D. Stearoyl-CoA Desaturase-1: Is It the Link between Sulfur Amino Acids and Lipid Metabolism? Biology 2015, 4, 383-396. https://doi.org/10.3390/biology4020383
Poloni S, Blom HJ, Schwartz IVD. Stearoyl-CoA Desaturase-1: Is It the Link between Sulfur Amino Acids and Lipid Metabolism? Biology. 2015; 4(2):383-396. https://doi.org/10.3390/biology4020383
Chicago/Turabian StylePoloni, Soraia, Henk J. Blom, and Ida V. D. Schwartz. 2015. "Stearoyl-CoA Desaturase-1: Is It the Link between Sulfur Amino Acids and Lipid Metabolism?" Biology 4, no. 2: 383-396. https://doi.org/10.3390/biology4020383
APA StylePoloni, S., Blom, H. J., & Schwartz, I. V. D. (2015). Stearoyl-CoA Desaturase-1: Is It the Link between Sulfur Amino Acids and Lipid Metabolism? Biology, 4(2), 383-396. https://doi.org/10.3390/biology4020383