Are There Circadian Clocks in Non-Photosynthetic Bacteria?

 and
and 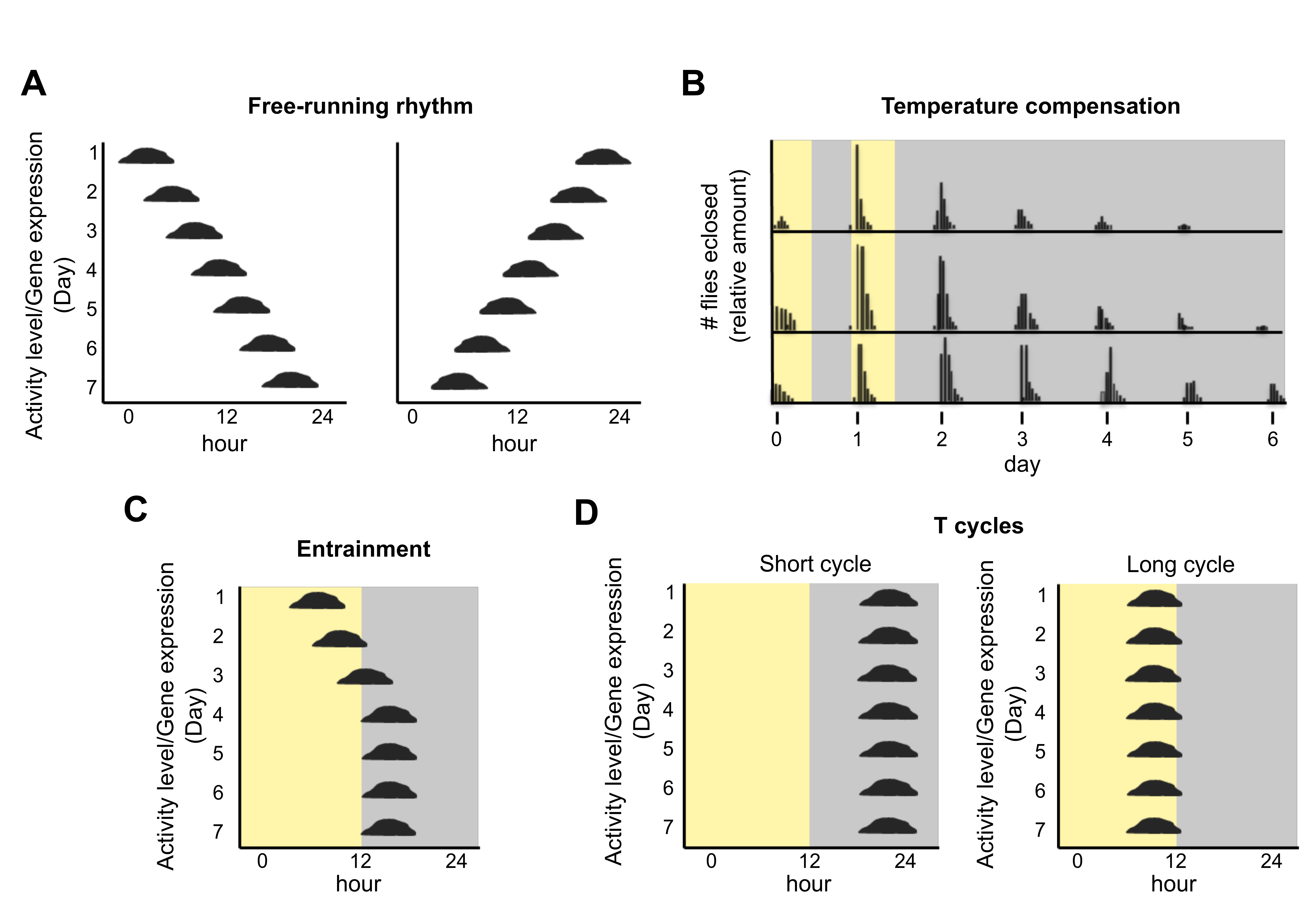{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
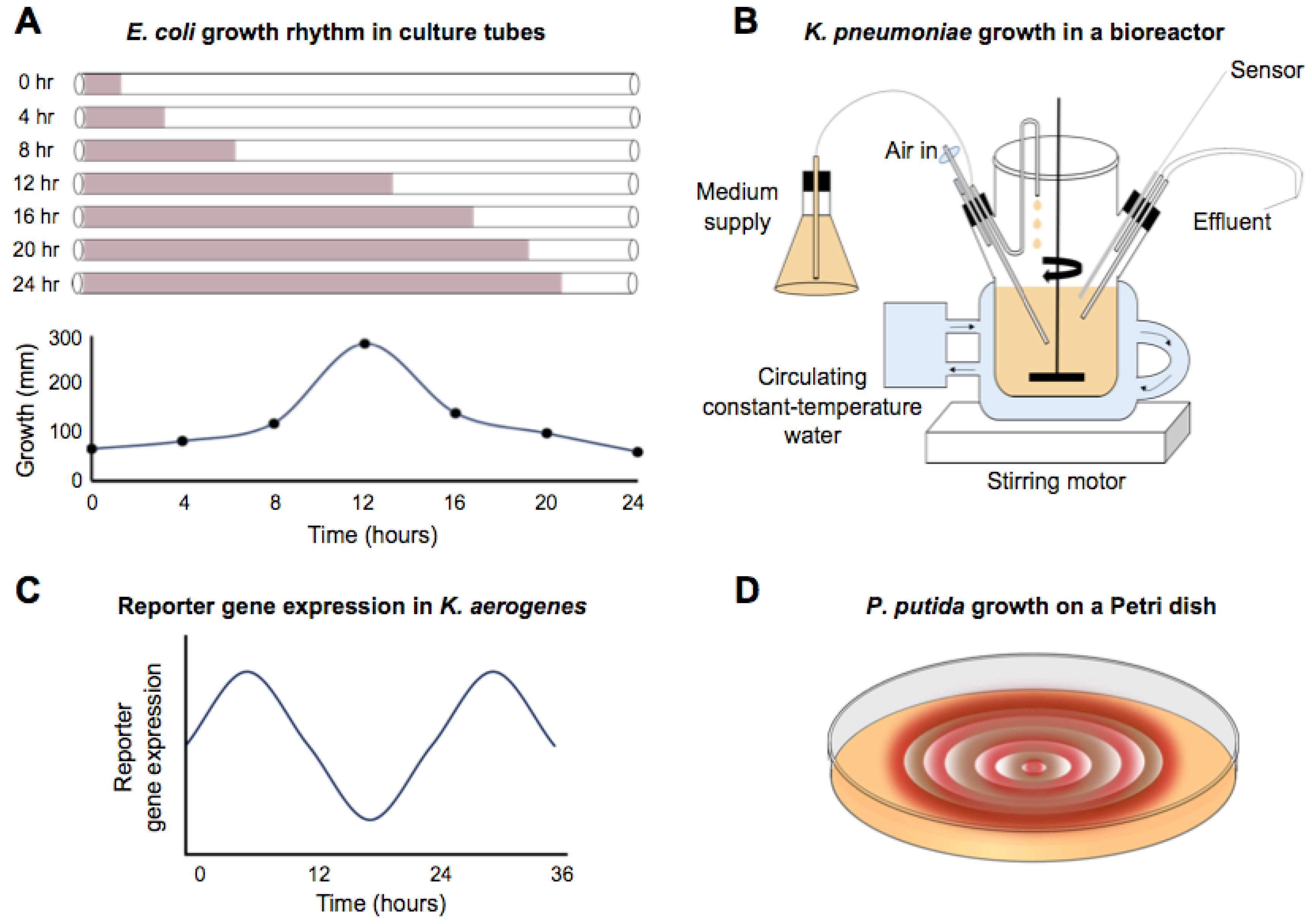
2. Endogenous Circadian Rhythms in Non-Photosynthetic Prokaryotes
3. Possible Timekeeping Components in Non-Photosynthetic Eubacteria
4. Bacillus subtilis—A Potential Model to Investigate Circadian Systems in Non-Photosynthetic Bacteria
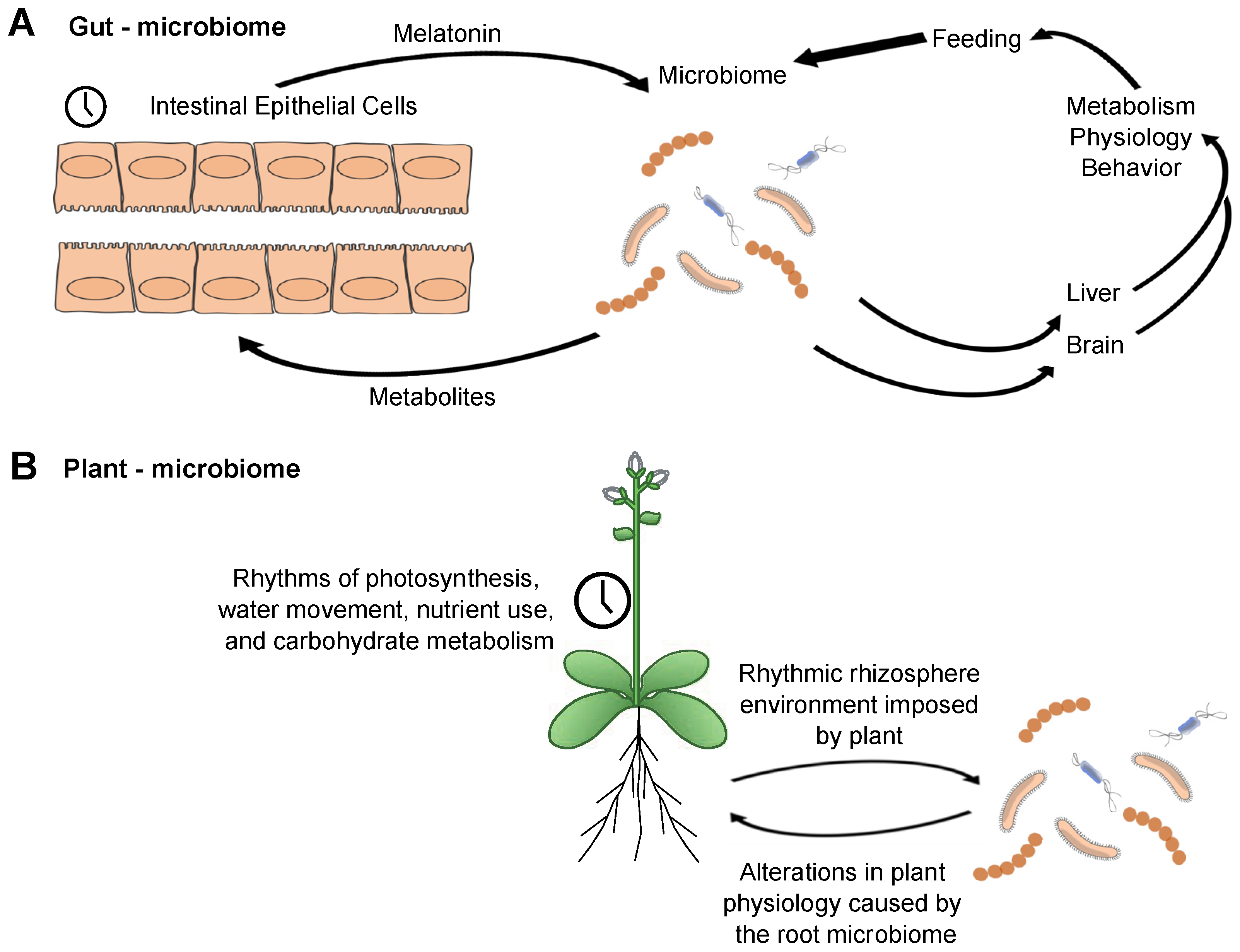
5. Microbiomes and Daily Rhythms
6. The Microbiome as an Organism and Temporal Structuring of Mutualism
7. Why Do We Care?
8. A Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Abraham, D.; Dallmann, R.; Steinlechner, S.; Albrecht, U.; Eichele, G.; Oster, H. Restoration of Circadian Rhythmicity in Circadian Clock-Deficient Mice in Constant Light. J. Biol. Rhythms 2006, 21, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Pittendrigh, C.S.; Caldarola, P.C. General homeostasis of the frequency of circadian oscillations. Proc. Natl. Acad. Sci. USA 1973, 70, 2697–2701. [Google Scholar] [CrossRef]
- Gardner, G.F.; Feldman, J.F. Temperature Compensation of Circadian Period Length in Clock Mutants of Neurospora crassa. Plant Physiol. 1981, 68, 1244–1248. [Google Scholar] [CrossRef]
- Barrett, R.K.; Takahashi, J.S. Temperature compensation and temperature entrainment of the chick pineal cell circadian clock. J. Neurosci. 1995, 15, 5681–5692. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, W.F.; Pittendrigh, C.S.; Pavlidis, T. Temperature compensation of the circadian oscillation in Drosophila pseudoobscura and its entrainment by temperature cycles. J. Insect Physiol. 1968, 14, 669–684. [Google Scholar] [CrossRef]
- Merrow, M.; Spoelstra, K.; Roenneberg, T. The circadian cycle: Daily rhythms from behaviour to genes. EMBO Rep. 2005, 6, 930–935. [Google Scholar] [CrossRef]
- Pittendrigh, C.S. On Temperature Independence in the Clock System Controlling Emergence Time in Drosophila. Proc. Natl. Acad. Sci. USA 1954, 40, 1018–1029. [Google Scholar] [CrossRef]
- Mrosovsky, N. Masking: History, definitions, and measurement. Chronobiol Int. 1999, 16, 415–429. [Google Scholar] [CrossRef]
- Eelderink-Chen, Z.; Mazzotta, G.; Sturre, M.; Bosman, J.; Roenneberg, T.; Merrow, M. A circadian clock in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2010, 107, 2043–2047. [Google Scholar] [CrossRef]
- Aschoff, J.; Pohl, H. Phase relations between a circadian rhythm and its zeitgeber within the range of entrainment. Naturwissenschaften 1978, 65, 80–84. [Google Scholar] [CrossRef]
- Tan, Y.; Dragovic, Z.; Roenneberg, T.; Merrow, M. Entrainment dissociates transcription and translation of a circadian clock gene in Neurospora. Curr. Biol. 2004, 14, 433–438. [Google Scholar] [CrossRef]
- Rémi, J.; Merrow, M.; Roenneberg, T. A circadian surface of entrainment: Varying T, τ, and Photoperiod in Neurospora crassa. J. Biol. Rhythms 2010. [Google Scholar] [CrossRef]
- Kondo, T.; Tsinoremas, N.F.; Golden, S.S.; Johnson, C.H.; Kutsuna, S.; Ishiura, M. Circadian clock mutants of cyanobacteria. Science 1994. [Google Scholar] [CrossRef]
- Liu, Y.; Tsinoremas, N.F.; Johnson, C.H.; Lebedeva, N.V.; Golden, S.S.; Ishiura, M.; Kondo, T. Circadian orchestration of gene expression in cyanobacteria. Genes Dev. 1995, 9, 1469–1478. [Google Scholar] [CrossRef]
- Nakajima, M.; Imai, K.; Ito, H.; Nishiwaki, T.; Murayama, Y.; Iwasaki, H.; Oyama, T.; Kondo, T. Reconstitution of circadian oscillation of cyanobacterial KaiC phosphorylation in vitro. Science 2005, 308, 414–415. [Google Scholar] [CrossRef]
- Chen, A.H.; Lubkowicz, D.; Yeong, V.; Chang, R.L.; Silver, P.A. Transplantability of a circadian clock to a noncircadian organism. Sci. Adv. 2015, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Aoki, S.; Kondo, T.; Wada, H.; Ishiura, M. Circadian rhythm of the cyanobacterium Synechocystis sp. strain PCC 6803 in the dark. J. Bacteriol. 1997. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Andersson, C.R.; Kondo, T.; Golden, S.S.; Hirschie, C. Resonating circadian clocks enh; ince fitness in cyanobacteria. Evolution 2009. [Google Scholar] [CrossRef]
- Woelfle, M.A.; Ouyang, Y.; Phanvijhitsiri, K.; Johnson, C.H. The adaptive value of circadian clocks: An experimental assessment in cyanobacteria. Curr. Biol. 2004, 14, 1481–1486. [Google Scholar] [CrossRef]
- Welkie, D.G.; Rubin, B.E.; Chang, Y.-G.; Diamond, S.; Rifkin, S.A.; LiWang, A.; Golden, S.S. Genome-wide fitness assessment during diurnal growth reveals an expanded role of the cyanobacterial circadian clock protein KaiA. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef]
- Van Praag, E.; Agosti, R.D.; Bachofen, R. Rhythmic activity of uptake hydrogenase in the prokaryote Rhodospirillum rubrum. J. Biol. Rhythms 2000. [Google Scholar] [CrossRef]
- Min, H.; Guo, H.; Xiong, J. Rhythmic gene expression in a purple photosynthetic bacterium, Rhodobacter sphaeroides. FEBS Lett. 2005. [Google Scholar] [CrossRef]
- Ehret, C.F.; Wille, J.J. The Photobiology of Circadian Rhythms in Protozoa and other Eukaryotic Microorganisms. In Photobiology of Microorganisms; Halldal, P., Ed.; Wiley-Interscience: New York, NY, USA, 1970; pp. 369–416. [Google Scholar]
- Kondo, T.; Mori, T.; Lebedeva, N.V.; Aoki, S.; Ishiura, M.; Golden, S.S. Circadian rhythms in rapidly dividing cyanobacteria. Science 1997. [Google Scholar] [CrossRef]
- Norman, T.M.; Lord, N.D.; Paulsson, J.; Losick, R. Stochastic Switching of Cell Fate in Microbes. Annu. Rev. Microbiol. 2015. [Google Scholar] [CrossRef]
- Rogers, L.A.; Greenbank, G.R. The intermitent growth of bacterial cultures. J. Bacteriol. 1930, 16, 181–190. [Google Scholar]
- Halberg, F.; Conner, R.L. Circadian organization and microbiology: Variance spectra and a periodogram on behavior of Escherichia coli growing in fluid culture. Proc. Minn. Acad. Sci. USA 1961, 29, 227–239. [Google Scholar]
- Sturtevant, R. Circadian patterns in linear growth of Escherichia coli. Anat Rec. 1973, 175, 453. [Google Scholar]
- Bibb, L.B. Long Tube Method of Cultivating Microorganisms with Observations on Mobile Colonies in Liquid Medium. J. Bacteriol. 1925, 10, 561. [Google Scholar]
- Sturtevant, R.P. Circadian variability in Klebsiella demonstrated by cosinor analysis. Int. J. Chronobiol. 1973, 1, 141–146. [Google Scholar]
- Paulose, J.K.; Wright, J.M.; Patel, A.G.; Cassone, V.M. Human gut bacteria are sensitive to melatonin and express endogenous circadian rhythmicity. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Süel, G.M.; Garcia-Ojalvo, J.; Liberman, L.M.; Elowitz, M.B. An excitable gene regulatory circuit induces transient cellular differentiation. Nature 2006. [Google Scholar] [CrossRef] [PubMed]
- Norman, T.M.; Lord, N.D.; Paulsson, J.; Losick, R. Memory and modularity in cell-fate decision making. Nature 2013, 503, 481–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischofs, I.B.; Hug, J.A.; Liu, A.W.; Wolf, D.M.; Arkin, A.P. Complexity in bacterial cell-cell communication: Quorum signal integration and subpopulation signaling in the Bacillus subtilis phosphorelay. Proc. Natl. Acad. Sci. USA 2009, 106, 6459–6464. [Google Scholar] [CrossRef]
- Kuchina, A.; Espinar, L.; Çağatay, T.; Balbin, A.O.; Zhang, F.; Alvarado, A.; Garcia-Ojalvo, J.; Süel, G.M.; Çağatay, T.; Balbin, A.O.; et al. Temporal competition between differentiation programs determines cell fate choice. Mol. Syst. Biol. 2011, 7, 557. [Google Scholar] [CrossRef] [PubMed]
- Soriano, M.I.; Roibás, B.; García, A.B.; Espinosa-Urgel, M. Evidence of circadian rhythms in non-photosynthetic bacteria? J. Circadian Rhythms 2010, 8. [Google Scholar] [CrossRef] [PubMed]
- Dvornyk, V.; Vinogradova, O.; Nevo, E. Origin and evolution of circadian clock genes in prokaryotes. Proc. Natl. Acad. Sci. USA 2003. [Google Scholar] [CrossRef]
- Loza-Correa, M.; Gomez-Valero, L.; Buchrieser, C. Circadian clock proteins in prokaryotes: Hidden rhythms? Front. Microbiol. 2010. [Google Scholar] [CrossRef]
- Cleyet-Marel, J.C.; Rome, S.; Salducci, X.; Wery, J. Importance des composés phénoliques dans les interactions entre plantes et microorganismes: Exemple des relations Rhizobium/légumineuses. Acta Bot. Gallica 1996. [Google Scholar] [CrossRef]
- Rome, S.; Fernandez, M.P.; Brunel, B.; Normand, P.; Cleyet-Marel, J.-C. Sinorhizobium medicae sp. nov., Isolated from Annual Medicago spp. Int. J. Syst. Bacteriol. 1996. [Google Scholar] [CrossRef]
- Bernal, P.; Allsopp, L.P.; Filloux, A.; Llamas, M.A. The Pseudomonas putida T6SS is a plant warden against phytopathogens. ISME J. 2017. [Google Scholar] [CrossRef]
- Edgar, R.S.; Green, E.W.; Zhao, Y.; Van Ooijen, G.; Olmedo, M.; Qin, X.; Xu, Y.; Pan, M.; Valekunja, U.K.; Feeney, K.A.; et al. Peroxiredoxins are conserved markers of circadian rhythms. Nature 2012, 485. [Google Scholar] [CrossRef]
- Hall, A.; Sankaran, B.; Poole, L.B.; Karplus, P.A. Structural Changes Common to Catalysis in the Tpx Peroxiredoxin Subfamily. J. Mol. Biol. 2009. [Google Scholar] [CrossRef]
- Davis, S.J.; Vener, A.V.; Vierstra, R.D. Bacteriophytochromes: Phytochrome-like photoreceptors from nonphotosynthetic eubacteria. Science 1999, 286, 2517–2520. [Google Scholar] [CrossRef]
- Bhoo, S.H.; Davis, S.J.; Walker, J.; Karniol, B.; Vierstra, R.D. Bacteriophytochromes are photochromic histidine kinases using a biliverdin chromophore. Nature 2001, 414, 776–779. [Google Scholar] [CrossRef]
- Rottwinkel, G.; Oberpichler, I.; Lamparter, T. Bathy phytochromes in rhizobial soil bacteria. J. Bacteriol. 2010. [Google Scholar] [CrossRef]
- Lee, H.J.; Ha, J.H.; Kim, S.G.; Choi, H.K.; Kim, Z.H.; Han, Y.J.; Kim, J.I.; Oh, Y.; Fragoso, V.; Shin, K.; et al. Stem-piped light activates phytochrome B to trigger light responses in Arabidopsis thaliana roots. Sci. Signal. 2016. [Google Scholar] [CrossRef]
- Nimmo, H.G. Entrainment of Arabidopsis roots to the light:dark cycle by light piping. Plant Cell Environ. 2018. [Google Scholar] [CrossRef]
- Losi, A.; Polverini, E.; Quest, B.; Gärtner, W. First evidence for phototropin-related blue-light receptors in prokaryotes. Biophys. J. 2002. [Google Scholar] [CrossRef]
- Gomelsky, M.; Klug, G. BLUF: A novel FAD-binding domain involved in sensory transduction in microorganisms. Trends Biochem. Sci. 2002. [Google Scholar] [CrossRef]
- Tschowri, N.; Busse, S.; Hengge, R. The BLUF-EAL protein YcgF acts as a direct anti-repressor in a blue-light response of Escherichia coli. Genes Dev. 2009. [Google Scholar] [CrossRef]
- Memmi, S.; Kyndt, J.; Meyer, T.; Devreese, B.; Cusanovich, M.; Van Beeumen, J. Photoactive yellow protein from the halophilic bacterium Salinibacter ruber. Biochemistry 2008. [Google Scholar] [CrossRef]
- Kumauchi, M.; Hara, M.T.; Stalcup, P.; Xie, A.; Hoff, W.D. Identification of six new photoactive yellow proteins-diversity and structure-function relationships in a bacterial blue light photoreceptor. Photochem. Photobiol. 2008. [Google Scholar] [CrossRef]
- Bonomi, H.R.; Posadas, D.M.; Paris, G.; del Carrica, M.C.; Frederickson, M.; Pietrasanta, L.I.; Bogomolni, R.A.; Zorreguieta, A.; Goldbaum, F.A. Light regulates attachment, exopolysaccharide production, and nodulation in Rhizobium leguminosarum through a LOV-histidine kinase photoreceptor. Proc. Natl. Acad. Sci. USA 2012. [Google Scholar] [CrossRef]
- Müller, S.; Strack, S.N.; Hoefler, B.C.; Straight, P.D.; Kearns, D.B.; Kirby, J.R. Bacillaene and Sporulation Protect Bacillus subtilis from Predation by Myxococcus xanthus. Appl. Environ. Microbiol. 2014. [Google Scholar] [CrossRef]
- Shank, E.A.; Klepac-Ceraj, V.; Collado-Torres, L.; Powers, G.E.; Losick, R.; Kolter, R. Interspecies interactions that result in Bacillus subtilis forming biofilms are mediated mainly by members of its own genus. Proc. Natl. Acad. Sci. USA 2011. [Google Scholar] [CrossRef]
- Gallegos-Monterrosa, R.; Mhatre, E.; Kovács, Á.T. Specific Bacillus subtilis 168 variants form biofilms on nutrient-rich medium. Microbiology 2016, 162, 1922–1932. [Google Scholar] [CrossRef]
- Benoit, I.; van den Esker, M.H.; Patyshakuliyeva, A.; Mattern, D.J.; Blei, F.; Zhou, M.; Dijksterhuis, J.; Brakhage, A.A.; Kuipers, O.P.; de Vries, R.P.; Kovács, Á.T. Bacillus subtilis attachment to Aspergillus niger hyphae results in mutually altered metabolism. Environ. Microbiol. 2015. [Google Scholar] [CrossRef]
- Klobutcher, L.A.; Ragkousi, K.; Setlow, P. The Bacillus subtilis spore coat provides “eat resistance” during phagocytic predation by the protozoan Tetrahymena thermophila. Proc. Natl. Acad. Sci. USA 2005. [Google Scholar] [CrossRef]
- Laaberki, M.H.; Dworkin, J. Role of spore coat proteins in the resistance of Bacillus subtilis spores to Caenorhabditis elegans predation. J. Bacteriol. 2008. [Google Scholar] [CrossRef]
- Donato, V.; Ayala, F.R.; Cogliati, S.; Bauman, C.; Costa, J.G.; Leñini, C.; Grau, R. Bacillus subtilis biofilm extends Caenorhabditis elegans longevity through downregulation of the insulin-like signalling pathway. Nat. Commun. 2017. [Google Scholar] [CrossRef]
- Lopez, D.; Vlamakis, H.; Kolter, R. Generation of multiple cell types in Bacillus subtilis. FEMS Microbiol. Rev. 2009, 33, 152–163. [Google Scholar] [CrossRef]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 2010. [Google Scholar] [CrossRef]
- Hölscher, T.; Kovács, Á.T. Sliding on the surface: Bacterial spreading without an active motor. Environ. Microbiol. 2017. [Google Scholar] [CrossRef]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.; Dworkin, J. Recent progress in Bacillus subtilis sporulation. FEMS Microbiol. Rev. 2012. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.H.; Fontes, M.E.; Dworkin, J.; Elowitz, M.B. Pulsed feedback defers cellular differentiation. PLoS Biol. 2012. [Google Scholar] [CrossRef]
- Narula, J.; Kuchina, A.; Lee, D.Y.D.; Fujita, M.; Süel, G.M.; Igoshin, O.A. Chromosomal Arrangement of Phosphorelay Genes Couples Sporulation and DNA Replication. Cell 2015. [Google Scholar] [CrossRef]
- Pittendrigh, C.S. Circadian rhythms and the circadian organization of living systems. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 159–184. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Mori, T.; Zhao, C.; Thiel, T.; Johnson, C.H. Evolution of KaiC-Dependent Timekeepers: A Proto-circadian Timing Mechanism Confers Adaptive Fitness in the Purple Bacterium Rhodopseudomonas palustris. PLoS Genet. 2016. [Google Scholar] [CrossRef]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and feeding pattern affect the diurnal dynamics of the gut microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef]
- Liang, X.; Bushman, F.D.; FitzGerald, G.A. Rhythmicity of the intestinal microbiota is regulated by gender and the host circadian clock. Proc. Natl. Acad. Sci. USA 2015, 112, 10479–10484. [Google Scholar] [CrossRef] [Green Version]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom control of microbiota diurnal oscillations promotes metabolic homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef]
- Mukherji, A.; Kobiita, A.; Ye, T.; Chambon, P. Homeostasis in intestinal epithelium is orchestrated by the circadian clock and microbiota cues transduced by TLRs. Cell 2013. [Google Scholar] [CrossRef] [PubMed]
- Montagner, A.; Korecka, A.; Polizzi, A.; Lippi, Y.; Blum, Y.; Canlet, C.; Tremblay-Franco, M.; Gautier-Stein, A.; Burcelin, R.; Yen, Y.C.; et al. Hepatic circadian clock oscillators and nuclear receptors integrate microbiome-derived signals. Sci. Rep. 2016. [Google Scholar] [CrossRef]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of diurnal variation of gut microbes and high-fat feeding on host circadian clock function and metabolism. Cell Host Microbe 2015. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Tognini, P.; Liu, Y.; Eckel-Mahan, K.L.; Baldi, P.; Sassone-Corsi, P. Gut microbiota directs PPARγ-driven reprogramming of the liver circadian clock by nutritional challenge. EMBO Rep. 2016. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Nobs, S.P.; Elinav, E. NFIL-trating the Host Circadian Rhythm—Microbes Fine-Tune the Epithelial Clock. Cell Metab. 2017, 26, 699–700. [Google Scholar] [CrossRef] [PubMed]
- Weger, B.D.; Gobet, C.; Yeung, J.; Martin, E.; Jimenez, S.; Betrisey, B.; Foata, F.; Berger, B.; Balvay, A.; Foussier, A.; et al. The Mouse Microbiome Is Required for Sex-Specific Diurnal Rhythms of Gene Expression and Metabolism. Cell Metab. 2018. [Google Scholar] [CrossRef] [PubMed]
- Konturek, S.J.; Konturek, P.C.; Brzozowska, I.; Pawlik, M.; Sliwowski, Z.; Cześnikiewicz-Guzik, M.; Kwiecień, S.; Brzozowski, T.; Bubenik, G.A.; Pawlik, W.W. Localization and biological activities of melatonin in intact and diseased Gastrointestinal Tract (GIT). J. Physiol. Pharmacol. 2007. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016, 167, 1495–1510.e12. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.Y.; Harmer, S.L. Wheels within wheels: The plant circadian system. Trends Plant Sci. 2014. [Google Scholar] [CrossRef]
- Hennessey, T.L.; Field, C.B. Circadian Rhythms in Photosynthesis: Oscillations in Carbon Assimilation and Stomatal Conductance under Constant Conditions. Plant Physiol. 1991. [Google Scholar] [CrossRef]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kévei, E.; Tóth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A.R. Plant Circadian Clocks Increase Photosynthesis, Growth, Survival. Science 2007, 630, 630–633. [Google Scholar] [CrossRef]
- Graf, A.; Schlereth, A.; Stitt, M.; Smith, A.M. Circadian control of carbohydrate availability for growth in Arabidopsis plants at night. Proc. Natl. Acad. Sci. USA 2010. [Google Scholar] [CrossRef] [PubMed]
- Green, R.M.; Tingay, S.; Wang, Z.-Y.; Tobin, E.M. Circadian rhythms confer a higher level of fitness to Arabidopsis plants. Plant Physiol. 2002, 129, 576–584. [Google Scholar] [CrossRef]
- Hubbard, C.J.; Brock, M.T.; Van Diepen, L.T.; Maignien, L.; Ewers, B.E.; Weinig, C. The plant circadian clock influences rhizosphere community structure and function. ISME J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xie, Q.; Anderson, R.G.; Ng, G.; Seitz, N.C.; Peterson, T.; McClung, C.R.; McDowell, J.M.; Kong, D.; Kwak, J.M.; et al. Crosstalk between the Circadian Clock and Innate Immunity in Arabidopsis. PLoS Pathog. 2013. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Merrow, M. The circadian clock and human health. Curr. Biol. 2016, 26. [Google Scholar] [CrossRef]
- Margulis, L.; Fester, R. Symbiosis as a Source of Evolutionary Innovation, 1st ed.; MIT Press: Cambridge, UK, 1991. [Google Scholar]
- Simon, J.C.; Marchesi, J.R.; Mougel, C.; Selosse, M.A. Host-microbiota interactions: From holobiont theory to analysis. Microbiome 2019, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Hörnlein, C.; Confurius-Guns, V.; Stal, L.J.; Bolhuis, H. Daily rhythmicity in coastal microbial mats. NPJ Biofilms Microbiomes 2018. [Google Scholar] [CrossRef]
- D’Souza, G.; Shitut, S.; Preussger, D.; Yousif, G.; Waschina, S.; Kost, C. Ecology and evolution of metabolic cross-feeding interactions in bacteria. Nat. Prod. Rep. 2018, 35, 455–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spengler, M.L.; Artemicheva, N.M.; Kuropatwinski, K.K.; Gasparian, A.V.; Fedtsova, N.; Gitlin, I.I.; Antoch, M.P.; Gudkov, A.V.; Deluca, K.A.; Comas, M.; et al. Core circadian protein CLOCK is a positive regulator of NF-κB-mediated transcription. Proc. Natl. Acad. Sci. USA 2012. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Magri, A.; Hill, M.; Lai, A.G.; Kumar, A.; Rambhatla, S.B.; Donald, C.L.; Lopez-Clavijo, A.F.; Rudge, S.; Pinnick, K.; et al. The circadian clock components BMAL1 and REV-ERBα regulate flavivirus replication. Nat. Commun. 2019. [Google Scholar] [CrossRef]
- Edgar, R.S.; Stangherlin, A.; Nagy, A.D.; Nicoll, M.P.; Efstathiou, S.; O’Neill, J.S.; Reddy, A.B. Cell autonomous regulation of herpes and influenza virus infection by the circadian clock. Proc. Natl. Acad. Sci. USA 2016. [Google Scholar] [CrossRef]
- Dijk, D.J.; Duffy, J.F.; Silva, E.J.; Shanahan, T.L.; Boivin, D.B.; Czeisler, C.A. Amplitude reduction and phase shifts of Melatonin, cortisol and other circadian rhythms after a gradual advance of sleep and light exposure in humans. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Merrow, M.W. Process for the Production of a Compound or a Composition Employing a Culture of Microorganisms under Circadian Cultivation Conditions. Patent Application EU2210952A1, 27 January 2009. [Google Scholar]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 573. [Google Scholar] [CrossRef]
- Rudrappa, T.; Czymmek, K.J.; Pare, P.W.; Bais, H.P. Root-Secreted Malic Acid Recruits Beneficial Soil Bacteria. Plant Physiol. 2008. [Google Scholar] [CrossRef]
- Allard-Massicotte, R.; Tessier, L.; Lécuyer, F.; Lakshmanan, V.; Lucier, J.F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B. Bacillus subtilis early colonization of Arabidopsis thaliana roots involves multiple chemotaxis receptors. MBio 2016. [Google Scholar] [CrossRef]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, E1621–E1630. [Google Scholar] [CrossRef]
- Qiao, J.; Yu, X.; Liang, X.; Liu, Y.; Borriss, R.; Liu, Y. Addition of plant-growth-promoting Bacillus subtilis PTS-394 on tomato rhizosphere has no durable impact on composition of root microbiome. BMC Microbiol. 2017. [Google Scholar] [CrossRef]
- Swain, M.R.; Ray, R.C. Biocontrol and other beneficial activities of Bacillus subtilis isolated from cowdung microflora. Microbiol. Res. 2009. [Google Scholar] [CrossRef]
- Sharaf-Eldin, M.; Elkholy, S.; Fernández, J.A.; Junge, H.; Cheetham, R.; Guardiola, J.; Weathers, P. Bacillus subtilis FZB24® affects flower quantity and quality of saffron (Crocus sativus). Planta Med. 2008. [Google Scholar] [CrossRef]
- Hong, C.; Holtz, B.A.; Morgan, D.P.; Michailides, T.J. Significance of thinned fruit as a source of the secondary inoculum of Monilinia fructicola in California nectarine orchards. Plant Dis. 1997, 81, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Ongena, M.; Jacques, P.; Touré, Y.; Destain, J.; Jabrane, A.; Thonart, P. Involvement of fengycin-type lipopeptides in the multifaceted biocontrol potential of Bacillus subtilis. Appl. Microbiol. Biotechnol. 2005. [Google Scholar] [CrossRef]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef]
- Hanano, S.; Domagalska, M.A.; Nagy, F.; Davis, S.J. Multiple phytohormones influence distinct parameters of the plant circadian clock. Genes Cells 2006, 11, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Haydon, M.J.; Mielczarek, O.; Robertson, F.C.; Hubbard, K.E.; Webb, A.A.R. Photosynthetic entrainment of the Arabidopsis thaliana circadian clock. Nature 2013, 502, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Frank, A.; Matiolli, C.C.; Viana, A.J.C.; Hearn, T.J.; Kusakina, J.; Belbin, F.E.; Wells Newman, D.; Yochikawa, A.; Cano-Ramirez, D.L.; Chembath, A.; et al. Circadian Entrainment in Arabidopsis by the Sugar-Responsive Transcription Factor bZIP63. Curr. Biol. 2018, 28, 2597–2606.e6. [Google Scholar] [CrossRef] [PubMed]
- Thain, S.; Murtas, G.; Lynn, J. The circadian clock that controls gene expression in Arabidopsis is tissue specific. Plant Physiol. 2002, 130, 102–110. [Google Scholar] [CrossRef]
- James, A.B.; Monreal, J.A.; Nimmo, G.A.; Kelly, C.L.; Herzyk, P.; Jenkins, G.I.; Nimmo, H.G. The circadian clock in Arabidopsis roots is a simplified slave version of the clock in shoots. Science 2008, 322, 1832–1835. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sartor, F.; Eelderink-Chen, Z.; Aronson, B.; Bosman, J.; Hibbert, L.E.; Dodd, A.N.; Kovács, Á.T.; Merrow, M. Are There Circadian Clocks in Non-Photosynthetic Bacteria? Biology 2019, 8, 41. https://doi.org/10.3390/biology8020041
Sartor F, Eelderink-Chen Z, Aronson B, Bosman J, Hibbert LE, Dodd AN, Kovács ÁT, Merrow M. Are There Circadian Clocks in Non-Photosynthetic Bacteria? Biology. 2019; 8(2):41. https://doi.org/10.3390/biology8020041
Chicago/Turabian StyleSartor, Francesca, Zheng Eelderink-Chen, Ben Aronson, Jasper Bosman, Lauren E. Hibbert, Antony N. Dodd, Ákos T. Kovács, and Martha Merrow. 2019. "Are There Circadian Clocks in Non-Photosynthetic Bacteria?" Biology 8, no. 2: 41. https://doi.org/10.3390/biology8020041
APA StyleSartor, F., Eelderink-Chen, Z., Aronson, B., Bosman, J., Hibbert, L. E., Dodd, A. N., Kovács, Á. T., & Merrow, M. (2019). Are There Circadian Clocks in Non-Photosynthetic Bacteria? Biology, 8(2), 41. https://doi.org/10.3390/biology8020041