Lymphatic Vessels and Their Surroundings: How Local Physical Factors Affect Lymph Flow



Abstract
:Simple Summary
Abstract
1. Introduction
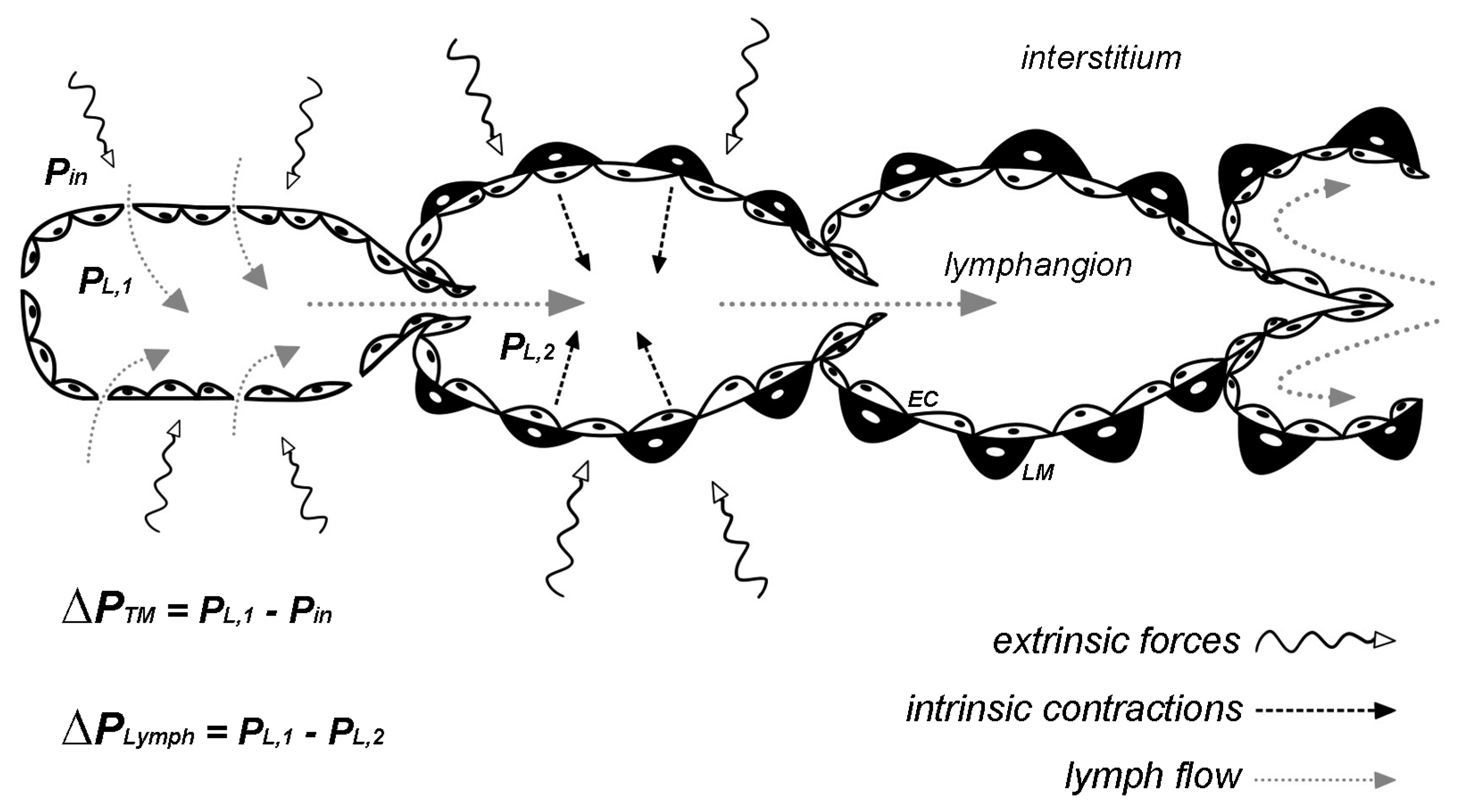
2. Hydraulic Pressure Changes
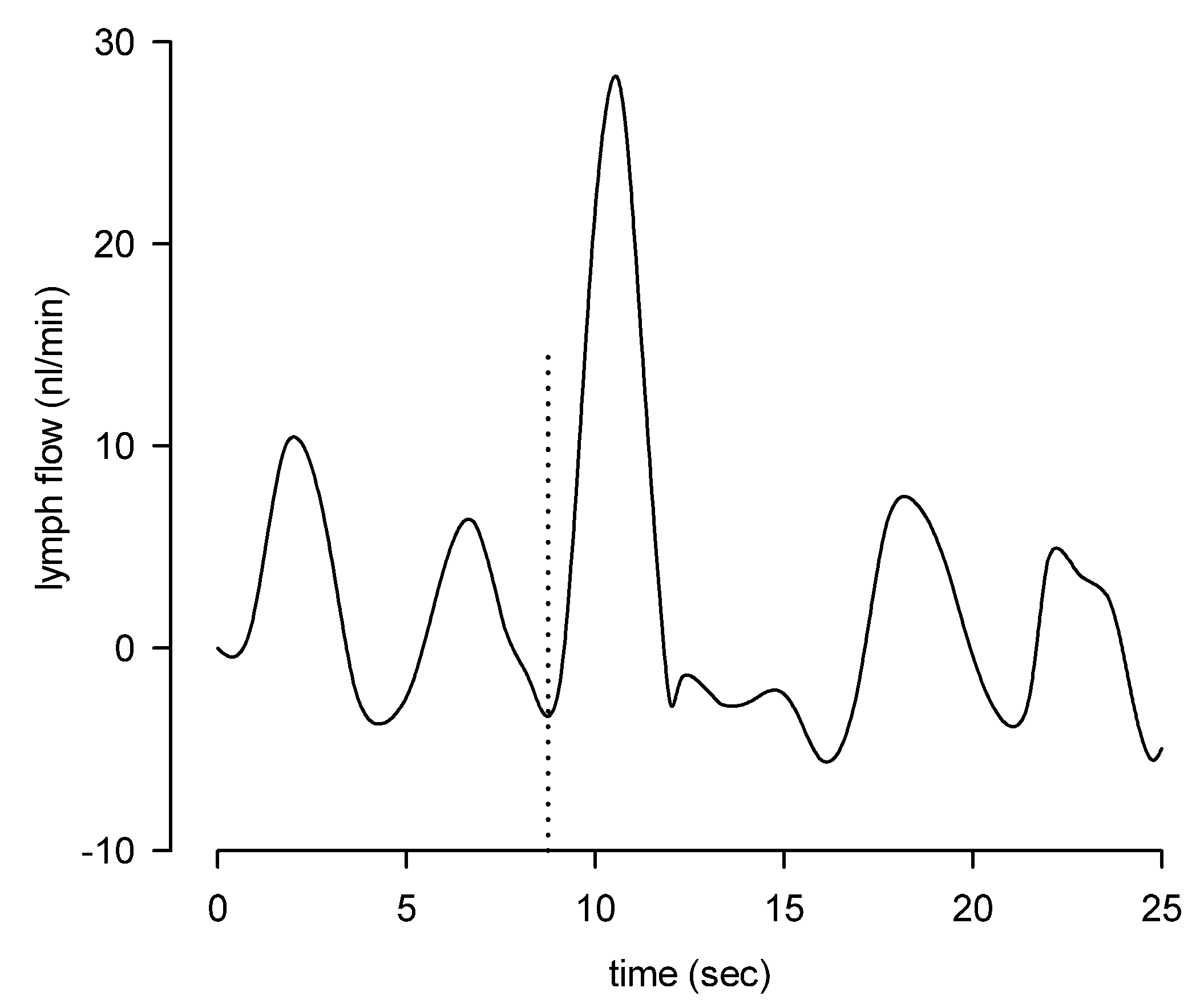
3. Lymph Flow and Shear Stress
4. Fluid Osmolarity in the Microenvironment
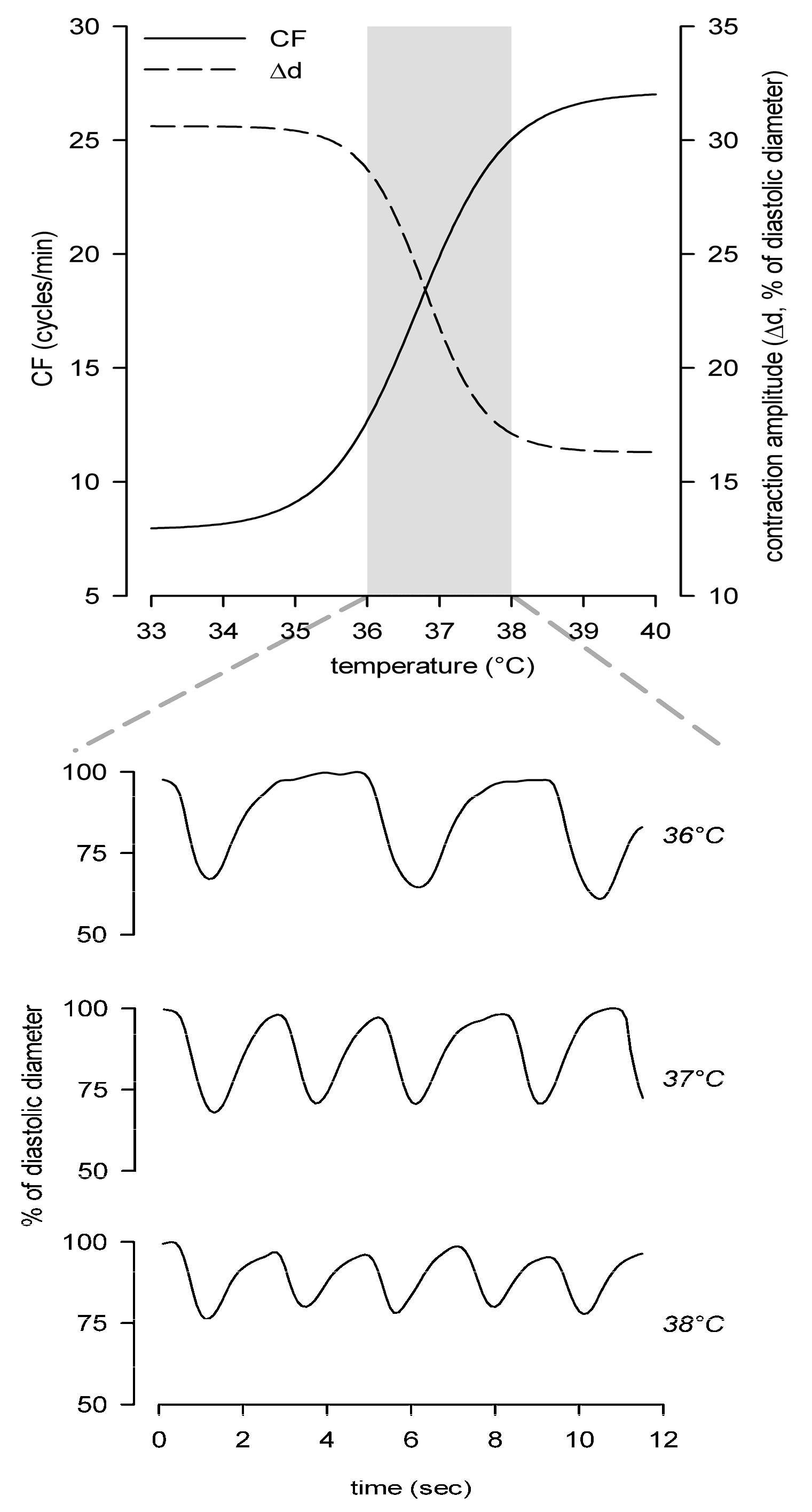
5. Local Tissue Temperature
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Aukland, K.; Reed, R.K. Interstitial-lymphatic mechanisms in the control of extracellular fluid volume. Physiol. Rev. 1993, 73, 1–78. [Google Scholar] [CrossRef] [PubMed]
- Wiig, H.; Swartz, M.A. Interstitial fluid and lymph formation and transport: Physiological regulation and roles in inflammation and cancer. Physiol. Rev. 2012, 92, 1005–1060. [Google Scholar] [CrossRef] [PubMed]
- Leak, L.V. Studies on the permeability of lymphatic capillaries. J. Cell Biol. 1971, 50, 300–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trzewik, J.; Mallipattu, S.K.; Artmann, G.M.; Delano, F.A.; Schmid-Schönbein, G.W. Evidence for a second valve system in lymphatics: Endothelial microvalves. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2001, 15, 1711–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazigou, E.; Wilson, J.T.; Moore, J.E.J. Primary and secondary lymphatic valve development: Molecular, functional and mechanical insights. Microvasc. Res. 2014, 96, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid-Schönbein, G.W. The second valve system in lymphatics. Lymphat. Res. Biol. 2003, 1, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Florey, H. Observations on the contractility of lacteals: Part I. J. Physiol. 1927, 62, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Breslin, J.W. Mechanical forces and lymphatic transport. Microvasc. Res. 2014, 96, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Bridenbaugh, E.A.; Gashev, A.A.; Zawieja, D.C. Lymphatic muscle: A review of contractile function. Lymphat. Res. Biol. 2003, 1, 147–158. [Google Scholar] [CrossRef]
- Muthuchamy, M.; Gashev, A.; Boswell, N.; Dawson, N.; Zawieja, D. Molecular and functional analyses of the contractile apparatus in lymphatic muscle. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 920–922. [Google Scholar] [CrossRef]
- Bridenbaugh, E.A.; Nizamutdinova, I.T.; Jupiter, D.; Nagai, T.; Thangaswamy, S.; Chatterjee, V.; Gashev, A.A. Lymphatic muscle cells in rat mesenteric lymphatic vessels of various ages. Lymphat. Res. Biol. 2013, 11, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargens, A.R.; Zweifach, B.W. Contractile stimuli in collecting lymph vessels. Am. J. Physiol. 1977, 233, H57–H65. [Google Scholar] [CrossRef] [PubMed]
- Van Helden, D.F. Pacemaker potentials in lymphatic smooth muscle of the guinea-pig mesentery. J. Physiol. 1993, 471, 465–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von der Weid, P.-Y.; Rahman, M.; Imtiaz, M.S.; van Helden, D.F. Spontaneous transient depolarizations in lymphatic vessels of the guinea pig mesentery: Pharmacology and implication for spontaneous contractility. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1989–H2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckett, E.A.H.; Hollywood, M.A.; Thornbury, K.D.; McHale, N.G. Spontaneous electrical activity in sheep mesenteric lymphatics. Lymphat. Res. Biol. 2007, 5, 29–43. [Google Scholar] [CrossRef]
- McCloskey, K.D.; Toland, H.M.; Hollywood, M.A.; Thornbury, K.D.; McHale, N.G. Hyperpolarisation-activated inward current in isolated sheep mesenteric lymphatic smooth muscle. J. Physiol. 1999, 521 Pt 1, 201–211. [Google Scholar] [CrossRef]
- Negrini, D.; Marcozzi, C.; Solari, E.; Bossi, E.; Cinquetti, R.; Reguzzoni, M.; Moriondo, A. Hyperpolarization-activated cyclic nucleotide-gated channels in peripheral diaphragmatic lymphatics. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H892–H903. [Google Scholar] [CrossRef] [Green Version]
- Zawieja, D.C.; Davis, K.L.; Schuster, R.; Hinds, W.M.; Granger, H.J. Distribution, propagation, and coordination of contractile activity in lymphatics. Am. J. Physiol. 1993, 264, H1283–H1291. [Google Scholar] [CrossRef]
- Mazzoni, M.C.; Skalak, T.C.; Schmid-Schonbein, G.W. Effects of skeletal muscle fiber deformation on lymphatic volumes. Am. J. Physiol. 1990, 259, H1860–H1868. [Google Scholar] [CrossRef]
- Leak, L.V.; Burke, J.F. Ultrastructural studies on the lymphatic anchoring filaments. J. Cell Biol. 1968, 36, 129–149. [Google Scholar] [CrossRef]
- Schmid-Schönbein, G.W. Microlymphatics and lymph flow. Physiol. Rev. 1990, 70, 987–1028. [Google Scholar] [CrossRef] [PubMed]
- Swartz, M.A. The physiology of the lymphatic system. Adv. Drug Deliv. Rev. 2001, 50, 3–20. [Google Scholar] [CrossRef]
- Negrini, D.; Moriondo, A.; Mukenge, S. Transmural Pressure During Cardiogenic Oscillations in Rodent Diaphragmatic Lymphatic Vessels. Lymphat. Res. Biol. 2004, 2, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Moriondo, A.; Mukenge, S.; Negrini, D. Transmural pressure in rat initial subpleural lymphatics during spontaneous or mechanical ventilation. Am. J. Physiol. Circ. Physiol. 2005, 289, H263–H269. [Google Scholar] [CrossRef] [PubMed]
- Skalak, T.C.; Schmid-Schönbein, G.W.; Zweifach, B.W. New morphological evidence for a mechanism of lymph formation in skeletal muscle. Microvasc. Res. 1984, 28, 95–112. [Google Scholar] [CrossRef]
- McGeown, J.G.; McHale, N.G.; Thornbury, K.D. The role of external compression and movement in lymph propulsion in the sheep hind limb. J. Physiol. 1987, 387, 83–93. [Google Scholar] [CrossRef]
- Moriondo, A.; Solari, E.; Marcozzi, C.; Negrini, D. Lymph flow pattern in pleural diaphragmatic lymphatics during intrinsic and extrinsic isotonic contraction. Am. J. Physiol. Hear. Circ. Physiol. 2016, 310, H60–H70. [Google Scholar] [CrossRef] [Green Version]
- Moriondo, A.; Solari, E.; Marcozzi, C.; Negrini, D. Diaphragmatic lymphatic vessel behavior during local skeletal muscle contraction. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H193–H205. [Google Scholar] [CrossRef] [Green Version]
- Moriondo, A.; Solari, E.; Marcozzi, C.; Negrini, D. Spontaneous activity in peripheral diaphragmatic lymphatic loops. Am. J. Physiol. Heart Circ. Physiol. 2013, 305, H987–H995. [Google Scholar] [CrossRef] [Green Version]
- Boriek, A.M.; Rodarte, J.R.; Reid, M.B. Shape and tension distribution of the passive rat diaphragm. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R33–R41. [Google Scholar] [CrossRef]
- Ohtani, O.; Ohtani, Y. Organization and developmental aspects of lymphatic vessels. Arch. Histol. Cytol. 2008, 71, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solari, E.; Marcozzi, C.; Bartolini, B.; Viola, M.; Negrini, D.; Moriondo, A. Acute Exposure of Collecting Lymphatic Vessels to Low-Density Lipoproteins Increases Both Contraction Frequency and Lymph Flow: An In Vivo Mechanical Insight. Lymphat. Res. Biol. 2020, 18, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Telinius, N.; Baandrup, U.; Rumessen, J.; Pilegaard, H.; Hjortdal, V.; Aalkjaer, C.; Boedtkjer, D.B. The human thoracic duct is functionally innervated by adrenergic nerves. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H206–H213. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, S.B.; Gsponer, D.; Montoya-Zegarra, J.A.; Schneider, M.; Scholkmann, F.; Tacconi, C.; Noerrelykke, S.F.; Proulx, S.T.; Detmar, M. A Distinct Role of the Autonomic Nervous System in Modulating the Function of Lymphatic Vessels under Physiological and Tumor-Draining Conditions. Cell Rep. 2019, 27, 3305–3314.e13. [Google Scholar] [CrossRef] [PubMed]
- Bazigou, E.; Makinen, T. Flow control in our vessels: Vascular valves make sure there is no way back. Cell. Mol. Life Sci. 2013, 70, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Marshall, M.V.; Rasmussen, J.C.; Tan, I.-C.; Aldrich, M.B.; Adams, K.E.; Wang, X.; Fife, C.E.; Maus, E.A.; Smith, L.A.; Sevick-Muraca, E.M. Near-Infrared Fluorescence Imaging in Humans with Indocyanine Green: A Review and Update. Open Surg. Oncol. J. 2010, 2, 12–25. [Google Scholar] [CrossRef]
- Pujari, A.; Smith, A.F.; Hall, J.D.; Mei, P.; Chau, K.; Nguyen, D.T.; Sweet, D.T.; Jiménez, J.M. Lymphatic Valves Bifurcate Lymph Flow Into a Central Jet and a Slow-Moving Peri-Valvular Milieu. J. Biomech. Eng. 2020, 142. [Google Scholar] [CrossRef]
- Dixon, J.B.; Greiner, S.T.; Gashev, A.A.; Cote, G.L.; Moore, J.E.; Zawieja, D.C. Lymph flow, shear stress, and lymphocyte velocity in rat mesenteric prenodal lymphatics. Microcirculation 2006, 13, 597–610. [Google Scholar] [CrossRef]
- Zawieja, D.C. Contractile physiology of lymphatics. Lymphat. Res. Biol. 2009, 7, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Benoit, J.N.; Zawieja, D.C.; Goodman, A.H.; Granger, H.J. Characterization of intact mesenteric lymphatic pump and its responsiveness to acute edemagenic stress. Am. J. Physiol. 1989, 257, H2059–H2069. [Google Scholar] [CrossRef]
- Zweifach, B.W.; Prather, J.W. Micromanipulation of pressure in terminal lymphatics in the mesentery. Am. J. Physiol. 1975, 228, 1326–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gashev, A.A.; Davis, M.J.; Zawieja, D.C. Inhibition of the active lymph pump by flow in rat mesenteric lymphatics and thoracic duct. J. Physiol. 2002, 540, 1023–1037. [Google Scholar] [CrossRef] [PubMed]
- Gashev, A.A.; Davis, M.J.; Delp, M.D.; Zawieja, D.C. Regional variations of contractile activity in isolated rat lymphatics. Microcirculation 2004, 11, 477–492. [Google Scholar] [CrossRef] [PubMed]
- McHale, N.G.; Roddie, I.C. The effect of transmural pressure on pumping activity in isolated bovine lymphatic vessels. J. Physiol. 1976, 261, 255–269. [Google Scholar] [CrossRef] [Green Version]
- Zawieja, D.C.; Kossmann, E.; Pullin, J. Dynamics of the Microlymphatic System. In Progress in Applied Microcirculation; Karger Medical and Scientific Publishers: Basel, Switzerland, 1999; Volume 23, pp. 33–41. [Google Scholar]
- Zhang, R.-Z.; Gashev, A.A.; Zawieja, D.C.; Davis, M.J. Length-tension relationships of small arteries, veins, and lymphatics from the rat mesenteric microcirculation. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1943–H1952. [Google Scholar] [CrossRef]
- Gashev, A.A.; Zhang, R.-Z.; Muthuchamy, M.; Zawieja, D.C.; Davis, M.J. Regional heterogeneity of length-tension relationships in rat lymph vessels. Lymphat. Res. Biol. 2012, 10, 14–19. [Google Scholar] [CrossRef]
- Gashev, A.A.; Wang, W.; Laine, G.A.; Stewart, R.H.; Zawieja, D.C. Characteristics of the active lymph pump in bovine prenodal mesenteric lymphatics. Lymphat. Res. Biol. 2007, 5, 71–79. [Google Scholar] [CrossRef]
- Gasheva, O.Y.; Knippa, K.; Nepiushchikh, Z.V.; Muthuchamy, M.; Gashev, A.A. Age-related alterations of active pumping mechanisms in rat thoracic duct. Microcirculation 2007, 14, 827–839. [Google Scholar] [CrossRef]
- Quick, C.M.; Venugopal, A.M.; Gashev, A.A.; Zawieja, D.C.; Stewart, R.H. Intrinsic pump-conduit behavior of lymphangions. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1510–R1518. [Google Scholar] [CrossRef] [Green Version]
- Moriondo, A.; Bianchin, F.; Marcozzi, C.; Negrini, D. Kinetics of fluid flux in the rat diaphragmatic submesothelial lymphatic lacunae. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1182–H1190. [Google Scholar] [CrossRef]
- Sweet, D.T.; Jiménez, J.M.; Chang, J.; Hess, P.R.; Mericko-Ishizuka, P.; Fu, J.; Xia, L.; Davies, P.F.; Kahn, M.L. Lymph flow regulates collecting lymphatic vessel maturation in vivo. J. Clin. Investig. 2015, 125, 2995–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planas-Paz, L.; Lammert, E. Mechanosensing in developing lymphatic vessels. Adv. Anat. Embryol. Cell Biol. 2014, 214, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Sabine, A.; Saygili Demir, C.; Petrova, T.V. Endothelial Cell Responses to Biomechanical Forces in Lymphatic Vessels. Antioxid. Redox Signal. 2016, 25, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, R.; Koller, A.; Kaley, G. Regulation of the vasomotor activity of lymph microvessels by nitric oxide and prostaglandins. Am. J. Physiol. 1998, 274, R790–R796. [Google Scholar] [CrossRef] [PubMed]
- Datar, S.A.; Gong, W.; He, Y.; Johengen, M.; Kameny, R.J.; Raff, G.W.; Maltepe, E.; Oishi, P.E.; Fineman, J.R. Disrupted NOS signaling in lymphatic endothelial cells exposed to chronically increased pulmonary lymph flow. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H137–H145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von der Weid, P.Y. ATP-sensitive K+ channels in smooth muscle cells of guinea-pig mesenteric lymphatics: Role in nitric oxide and beta-adrenoceptor agonist-induced hyperpolarizations. Br. J. Pharmacol. 1998, 125, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Von der Weid, P.Y.; Zhao, J.; Van Helden, D.F. Nitric oxide decreases pacemaker activity in lymphatic vessels of guinea pig mesentery. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H2707–H2716. [Google Scholar] [CrossRef]
- Gasheva, O.Y.; Gashev, A.A.; Zawieja, D.C. Cyclic guanosine monophosphate and the dependent protein kinase regulate lymphatic contractility in rat thoracic duct. J. Physiol. 2013, 591, 4549–4565. [Google Scholar] [CrossRef]
- Coste, B.; Xiao, B.; Santos, J.S.; Syeda, R.; Grandl, J.; Spencer, K.S.; Kim, S.E.; Schmidt, M.; Mathur, J.; Dubin, A.E.; et al. Piezo proteins are pore-forming subunits of mechanically activated channels. Nature 2012, 483, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Ranade, S.S.; Qiu, Z.; Woo, S.-H.; Hur, S.S.; Murthy, S.E.; Cahalan, S.M.; Xu, J.; Mathur, J.; Bandell, M.; Coste, B.; et al. Piezo1, a mechanically activated ion channel, is required for vascular development in mice. Proc. Natl. Acad. Sci. USA 2014, 111, 10347–10352. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.; Park, E.; Jung, E.; Cha, B.; Lee, S.; Yu, J.; Kim, P.M.; Lee, S.; Hong, Y.J.; Koh, C.J.; et al. Piezo1 incorporates mechanical force signals into the genetic program that governs lymphatic valve development and maintenance. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Nonomura, K.; Lukacs, V.; Sweet, D.T.; Goddard, L.M.; Kanie, A.; Whitwam, T.; Ranade, S.S.; Fujimori, T.; Kahn, M.L.; Patapoutian, A. Mechanically activated ion channel PIEZO1 is required for lymphatic valve formation. Proc. Natl. Acad. Sci. USA 2018, 115, 12817–12822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukacs, V.; Mathur, J.; Mao, R.; Bayrak-Toydemir, P.; Procter, M.; Cahalan, S.M.; Kim, H.J.; Bandell, M.; Longo, N.; Day, R.W.; et al. Impaired PIEZO1 function in patients with a novel autosomal recessive congenital lymphatic dysplasia. Nat. Commun. 2015, 6, 8329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Hou, B.; Tumova, S.; Muraki, K.; Bruns, A.; Ludlow, M.J.; Sedo, A.; Hyman, A.J.; McKeown, L.; Young, R.S.; et al. Piezo1 integration of vascular architecture with physiological force. Nature 2014, 515, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Ballermann, B.J.; Dardik, A.; Eng, E.; Liu, A. Shear stress and the endothelium. Kidney Int. Suppl. 1998, 67, S100–S108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Hooks, J.; Nepiyushchikh, Z.; Dixon, J.B. Entrainment of Lymphatic Contraction to Oscillatory Flow. Sci. Rep. 2019, 9, 5840. [Google Scholar] [CrossRef] [PubMed]
- Schacht, V.; Berens von Rautenfeld, D.; Abels, C. The lymphatic system in the dorsal skinfold chamber of the Syrian golden hamster in vivo. Arch. Dermatol. Res. 2004, 295, 542–548. [Google Scholar] [CrossRef]
- Jafarnejad, M.; Cromer, W.E.; Kaunas, R.R.; Zhang, S.L.; Zawieja, D.C.; Moore, J.E.J. Measurement of shear stress-mediated intracellular calcium dynamics in human dermal lymphatic endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H697–H706. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, E.; Akl, T.; Coté, G.L.; Moore, J.E.J.; Zawieja, D.C. Lymph transport in rat mesenteric lymphatics experiencing edemagenic stress. Microcirculation 2014, 21, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.T.; Wang, W.; Hellerstedt, A.H.; Zawieja, D.C.; Moore, J.E. Confocal image-based computational modeling of nitric oxide transport in a rat mesenteric lymphatic vessel. J. Biomech. Eng. 2013, 135, 51005. [Google Scholar] [CrossRef] [Green Version]
- Solari, E.; Marcozzi, C.; Negrini, D.; Moriondo, A. Fluid Osmolarity Acutely and Differentially Modulates Lymphatic Vessels Intrinsic Contractions and Lymph Flow. Front. Physiol. 2018, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Zingg, W.; Morgan, C.D.; Anderson, D.E. Blood viscosity, erythrocyte sedimentation rate, packed cell volume, osmolality, and plasma viscosity of the Wistar rat. Lab. Anim. Sci. 1971, 21, 740–742. [Google Scholar] [PubMed]
- Eggermont, J.; Trouet, D.; Carton, I.; Nilius, B. Cellular function and control of volume-regulated anion channels. Cell Biochem. Biophys. 2001, 35, 263–274. [Google Scholar] [CrossRef]
- Voets, T.; Droogmans, G.; Nilius, B. Membrane currents and the resting membrane potential in cultured bovine pulmonary artery endothelial cells. J. Physiol. 1996, 497 Pt 1, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-X.; Hatton, W.J.; Wang, G.L.; Zhong, J.; Yamboliev, I.; Duan, D.; Hume, J.R. Functional effects of novel anti-ClC-3 antibodies on native volume-sensitive osmolyte and anion channels in cardiac and smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H1453–H1463. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.P. The effects of Na(+)-K(+)-Cl- co-transport and Cl(-)-HCO3-exchange blockade on the membrane potential and intracellular chloride levels of rat arterial smooth muscle, in vitro. Exp. Physiol. 1992, 77, 857–862. [Google Scholar] [CrossRef] [Green Version]
- Chipperfield, A.R.; Harper, A.A. Chloride in smooth muscle. Prog. Biophys. Mol. Biol. 2000, 74, 175–221. [Google Scholar] [CrossRef]
- Behringer, E.J.; Scallan, J.P.; Jafarnejad, M.; Castorena-Gonzalez, J.A.; Zawieja, S.D.; Moore, J.E.J.; Davis, M.J.; Segal, S.S. Calcium and electrical dynamics in lymphatic endothelium. J. Physiol. 2017, 595, 7347–7368. [Google Scholar] [CrossRef] [Green Version]
- Solari, E.; Marcozzi, C.; Bistoletti, M.; Baj, A.; Giaroni, C.; Negrini, D.; Moriondo, A. TRPV4 channels’ dominant role in the temperature modulation of intrinsic contractility and lymph flow of rat diaphragmatic lymphatics. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H507–H518. [Google Scholar] [CrossRef]
- Nilius, B.; Watanabe, H.; Vriens, J. The TRPV4 channel: Structure-function relationship and promiscuous gating behaviour. Pflugers Arch. 2003, 446, 298–303. [Google Scholar] [CrossRef]
- Liedtke, W.; Choe, Y.; Martí-Renom, M.A.; Bell, A.M.; Denis, C.S.; Sali, A.; Hudspeth, A.J.; Friedman, J.M.; Heller, S. Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell 2000, 103, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Vriens, J.; Watanabe, H.; Janssens, A.; Droogmans, G.; Voets, T.; Nilius, B. Cell swelling, heat, and chemical agonists use distinct pathways for the activation of the cation channel TRPV4. Proc. Natl. Acad. Sci. USA 2004, 101, 396–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonkusare, S.K.; Bonev, A.D.; Ledoux, J.; Liedtke, W.; Kotlikoff, M.I.; Heppner, T.J.; Hill-Eubanks, D.C.; Nelson, M.T. Elementary Ca2+ signals through endothelial TRPV4 channels regulate vascular function. Science 2012, 336, 597–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilius, B.; Prenen, J.; Wissenbach, U.; Bödding, M.; Droogmans, G. Differential activation of the volume-sensitive cation channel TRP12 (OTRPC4) and volume-regulated anion currents in HEK-293 cells. Pflugers Arch. 2001, 443, 227–233. [Google Scholar] [CrossRef]
- Zakaria, E.R.; Hunt, C.M.; Li, N.; Harris, P.D.; Garrison, R.N. Disparity in osmolarity-induced vascular reactivity. J. Am. Soc. Nephrol. 2005, 16, 2931–2940. [Google Scholar] [CrossRef] [Green Version]
- Levine, S.E.; Granger, D.N.; Brace, R.A.; Taylor, A.E. Effect of hyperosmolality on vascular resistance and lymph flow in the cat ileum. Am. J. Physiol. 1978, 234, H14–H20. [Google Scholar] [CrossRef]
- Zakaria, E.R.; Althani, A.; Fawzi, A.A.; Fituri, O.M. Hyperosmolality-mediated peritoneal microvascular vasodilation is linked to aquaporin function. Adv. Perit. Dial. 2014, 30, 63–74. [Google Scholar]
- Zakaria, E.R.; Althani, A.; Fawzi, A.A.; Fituri, O.M. Molecular mechanisms of peritoneal dialysis-induced microvascular vasodilation. Adv. Perit. Dial. 2014, 30, 98–109. [Google Scholar]
- Toda, N.; Ayajiki, K.; Toda, H.; Hatano, Y.; Okamura, T. Mechanism underlying mannitol-induced relaxation in isolated monkey cerebral arteries. Am. J. Physiol. 1992, 262, H897–H902. [Google Scholar] [CrossRef]
- De Clerck, I.; Boussery, K.; Pannier, J.-L.; Van De Voorde, J. Hyperosmolarity increases K+-induced vasodilations in rat skeletal muscle arterioles. Med. Sci. Sports Exerc. 2005, 37, 220–226. [Google Scholar] [CrossRef]
- Moriondo, A.; Pelosi, P.; Passi, A.; Viola, M.; Marcozzi, C.; Severgnini, P.; Ottani, V.; Quaranta, M.; Negrini, D. Proteoglycan fragmentation and respiratory mechanics in mechanically ventilated healthy rats. J. Appl. Physiol. 2007, 103, 747–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelosi, P.; Rocco, P.R.M.; Negrini, D.; Passi, A. The extracellular matrix of the lung and its role in edema formation. An. Acad. Bras. Cienc. 2007, 79, 285–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solari, E.; Marcozzi, C.; Negrini, D.; Moriondo, A. Temperature-dependent modulation of regional lymphatic contraction frequency and flow. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H879–H889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuda, A.; Ohshima, N. In situ observations of spontaneous contractions of the peripheral lymphatic vessels in the rat mesentery: Effects of temperature. Experientia 1984, 40, 342–343. [Google Scholar] [CrossRef] [PubMed]
- Ito, E.; Ikemoto, Y.; Yoshioka, T. Thermodynamic implications of high Q 10 of thermo-TRP channels in living cells. Biophys. 2015, 11, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baylie, R.L.; Brayden, J.E. TRPV channels and vascular function. Acta Physiol. 2011, 203, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Benham, C.D.; Gunthorpe, M.J.; Davis, J.B. TRPV channels as temperature sensors. Cell Calcium 2003, 33, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E.; Miller, C. A thermodynamic framework for understanding temperature sensing by transient receptor potential (TRP) channels. Proc. Natl. Acad. Sci. USA 2011, 108, 19492–19497. [Google Scholar] [CrossRef] [Green Version]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2001; ISBN 9780878933211. [Google Scholar]
- DiFrancesco, D.; Ojeda, C. Properties of the current if in the sino-atrial node of the rabbit compared with those of the current iK, in Purkinje fibres. J. Physiol. 1980, 308, 353–367. [Google Scholar] [CrossRef]
- Yanagida, H.; Inoue, R.; Tanaka, M.; Ito, Y. Temperature-sensitive gating of cation current in guinea pig ileal muscle activated by hyperpolarization. Am. J. Physiol. Cell Physiol. 2000, 278, C40–C48. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Vessels | Imposed Flow Gradient (Δ cm H2O) | CF (Cycles min−1) | EF | FPF (min−1) |
|---|---|---|---|---|
| Mesenteric lymphatics | 0 | 9.0 ± 1.6 | 0.59 ± 0.05 | 5.1 ± 0.7 |
| 1 | 6.1 ± 18 | 0.49 ± 0.10 | 3.1 ± 0.7 | |
| 3 | 4.6 ± 2.1 | 0.27 ± 0.11 | 1.9 ± 0.9 | |
| 5 | 3.4 ± 1.5 | 0.23 ± 0.10 | 1.4 ± 0.8 | |
| Femoral lymphatics | 0 | 15.2 ± 2.6 | 0.17 ± 0.03 | 2.5 ± 0.6 |
| 1 | 9.7 ± 3.0 | 0.11 ± 0.03 | 1.5 ± 0.7 | |
| 3 | 7.7 ± 3.0 | 0.08 ± 0.03 | 1.1 ± 0.6 | |
| 5 | 5.0 ± 2.4 | 0.09 ± 0.04 | 0.4 ± 0.2 | |
| Cervical lymphatics | 0 | 18.9 ± 2.7 | 0.11 ± 0.02 | 2.1 ± 0.5 |
| 1 | 11.4 ± 3.8 | 0.08 ± 0.03 | 1.4 ± 0.5 | |
| 3 | 7.4 ± 3.6 | 0.02 ± 0.02 | 0.2 ± 0.2 | |
| 5 | 1.5 ± 1.5 | 0.02 ± 0.02 | 0.2 ± 0.2 | |
| Thoracic duct | 0 | 4.6 ± 0.6 | 0.31 ± 0.03 | 1.4 ± 0.2 |
| 1 | 2.8 ± 0.7 | 0.16 ± 0.04 | 0.5 ± 0.1 | |
| 3 | 0.4 ± 0.3 | 0.03 ± 0.02 | 0.4 ± 0.0 | |
| 5 | 0.1 ± 0.1 | 0.02 ± 0.02 | 0.01 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solari, E.; Marcozzi, C.; Negrini, D.; Moriondo, A. Lymphatic Vessels and Their Surroundings: How Local Physical Factors Affect Lymph Flow. Biology 2020, 9, 463. https://doi.org/10.3390/biology9120463
Solari E, Marcozzi C, Negrini D, Moriondo A. Lymphatic Vessels and Their Surroundings: How Local Physical Factors Affect Lymph Flow. Biology. 2020; 9(12):463. https://doi.org/10.3390/biology9120463
Chicago/Turabian StyleSolari, Eleonora, Cristiana Marcozzi, Daniela Negrini, and Andrea Moriondo. 2020. "Lymphatic Vessels and Their Surroundings: How Local Physical Factors Affect Lymph Flow" Biology 9, no. 12: 463. https://doi.org/10.3390/biology9120463
APA StyleSolari, E., Marcozzi, C., Negrini, D., & Moriondo, A. (2020). Lymphatic Vessels and Their Surroundings: How Local Physical Factors Affect Lymph Flow. Biology, 9(12), 463. https://doi.org/10.3390/biology9120463