The Cytoplasmic LIM Domain Protein Espinas Contributes to Photoreceptor Layer Selection in the Visual System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Genetics
2.2. DNA Constructs
2.3. Immunohistochemistry
2.4. Photoreceptor Phenotype Quantification and Statistics
3. Results
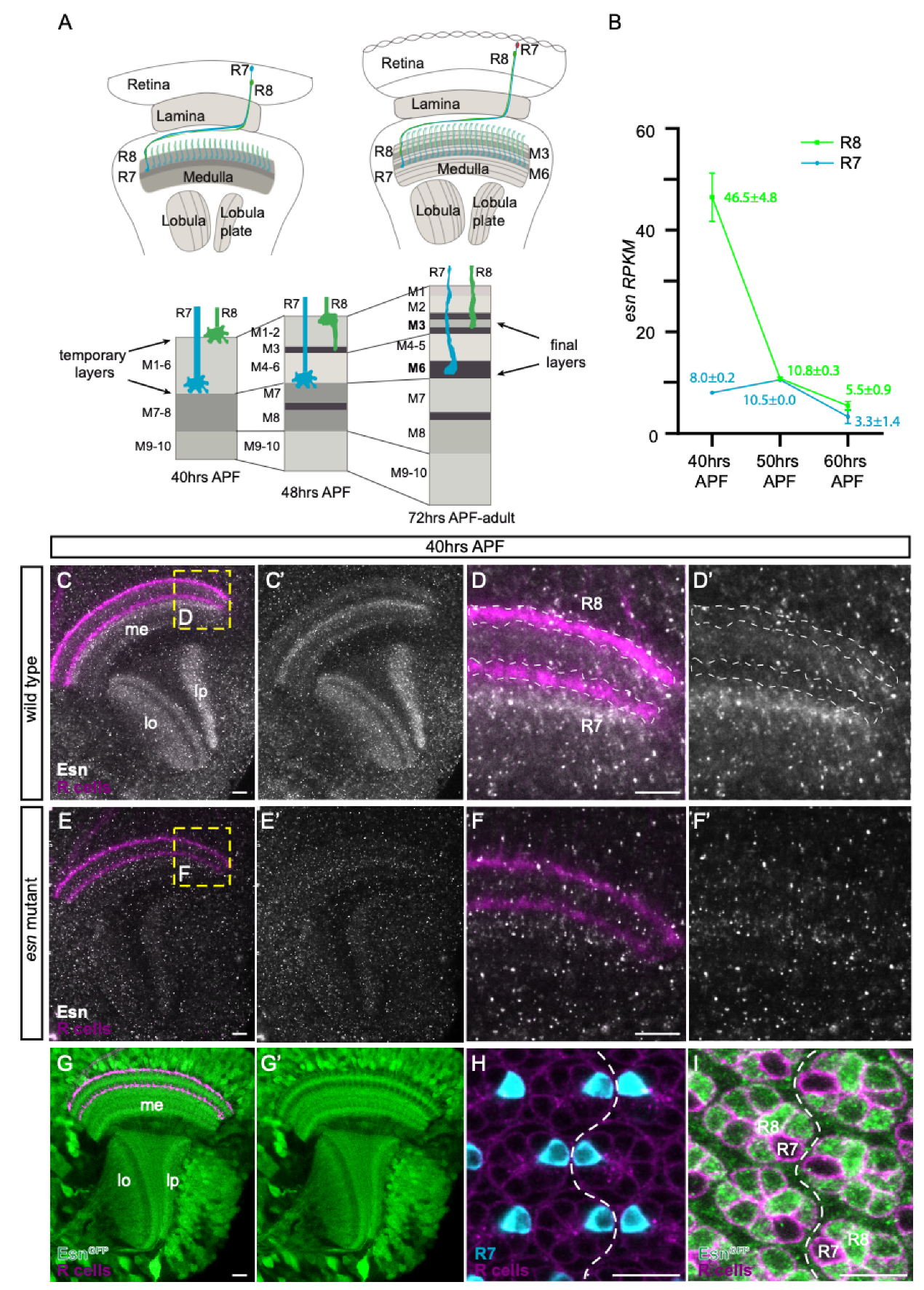
3.1. esn Is Expressed in the Optic Lobe and Enriched in R8 versus R7 Photoreceptors
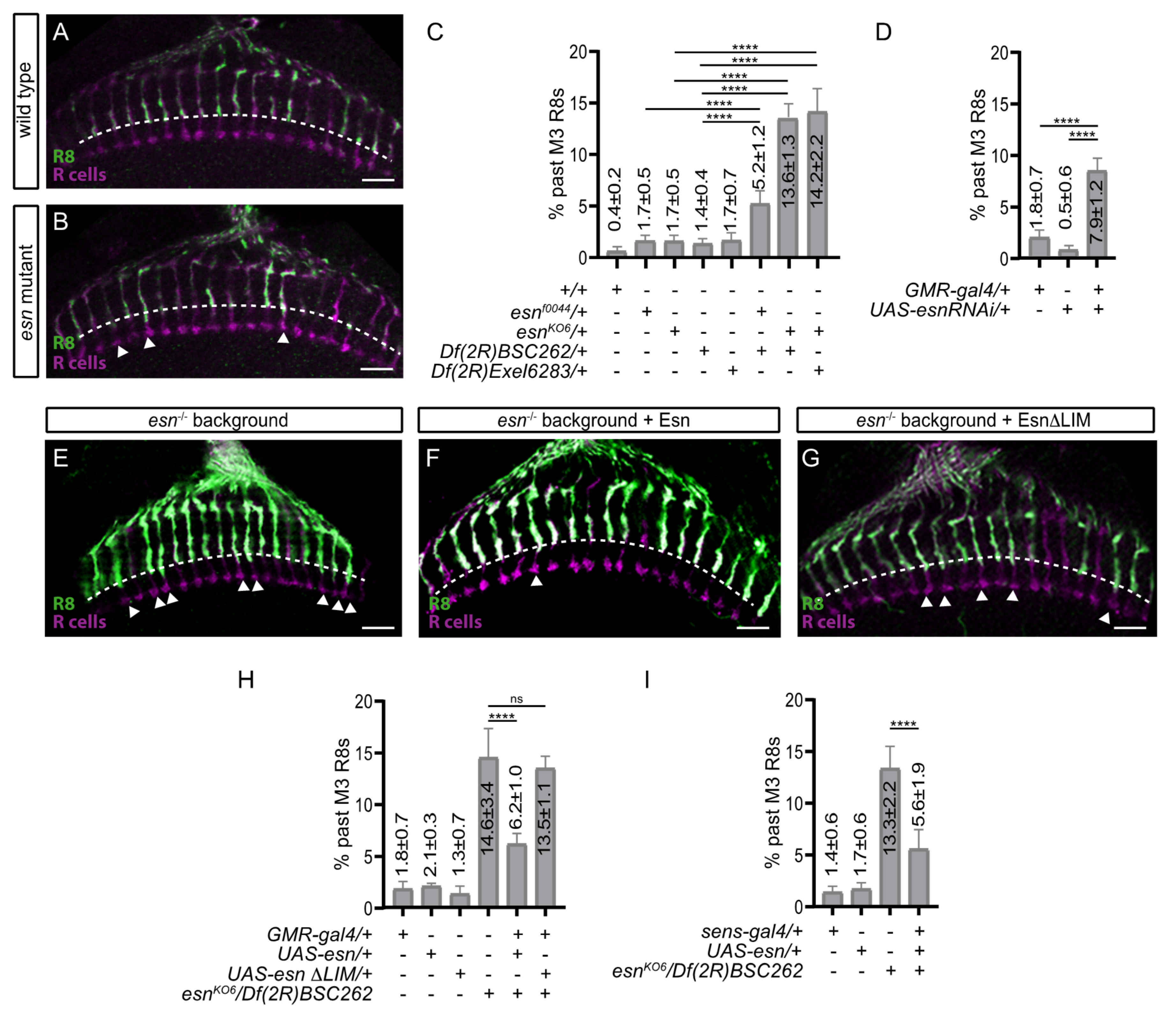
3.2. R8 Photoreceptor Axons in esn Mutant Animals Mistarget Past Their Final Target Layer
3.3. esn Is Required in the R8 Photoreceptor for Final Layer Selection
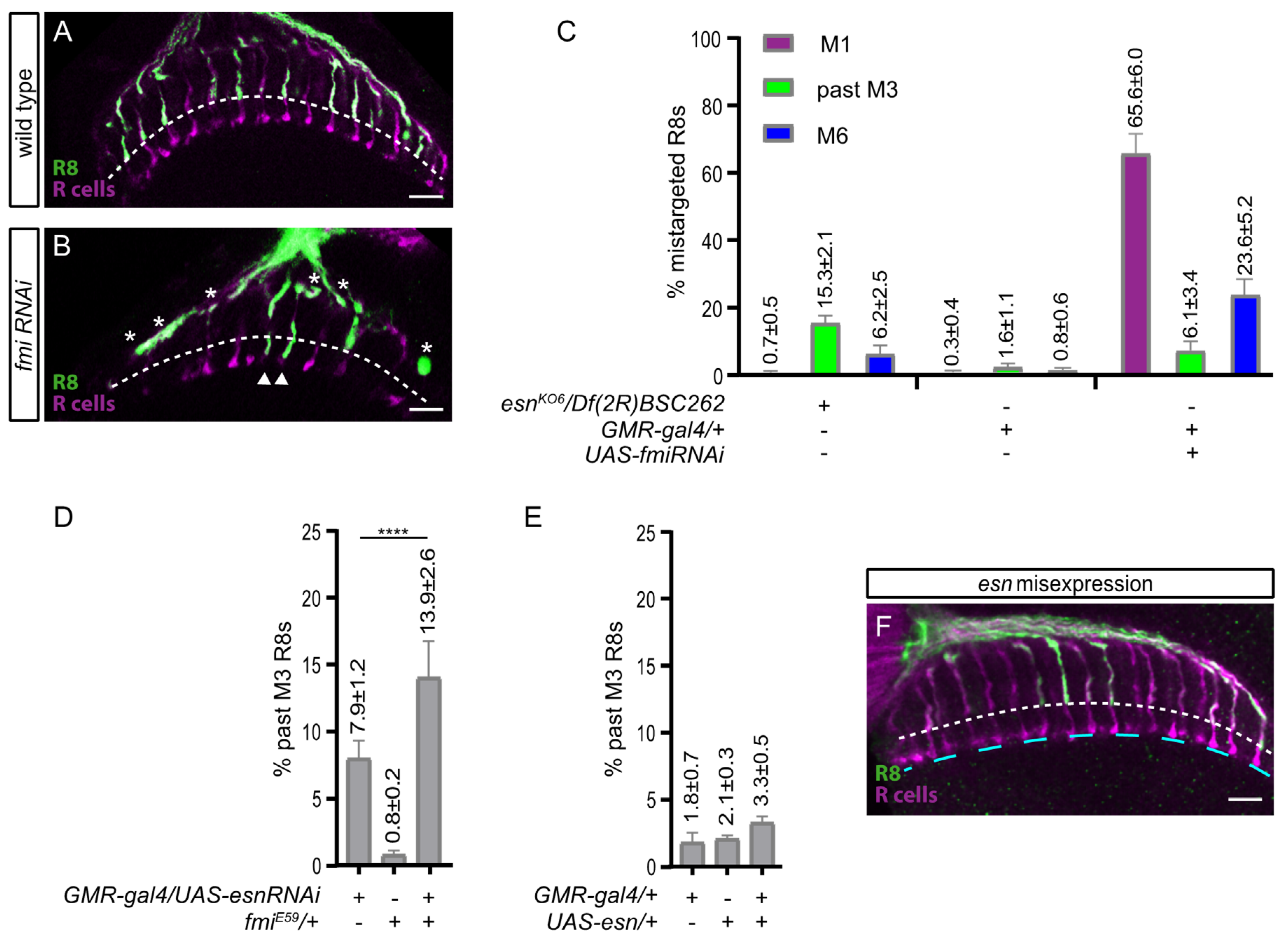
3.4. An esn/fmi Interaction Contributes R8 Final Layer Selection but esn Is Not Instructive
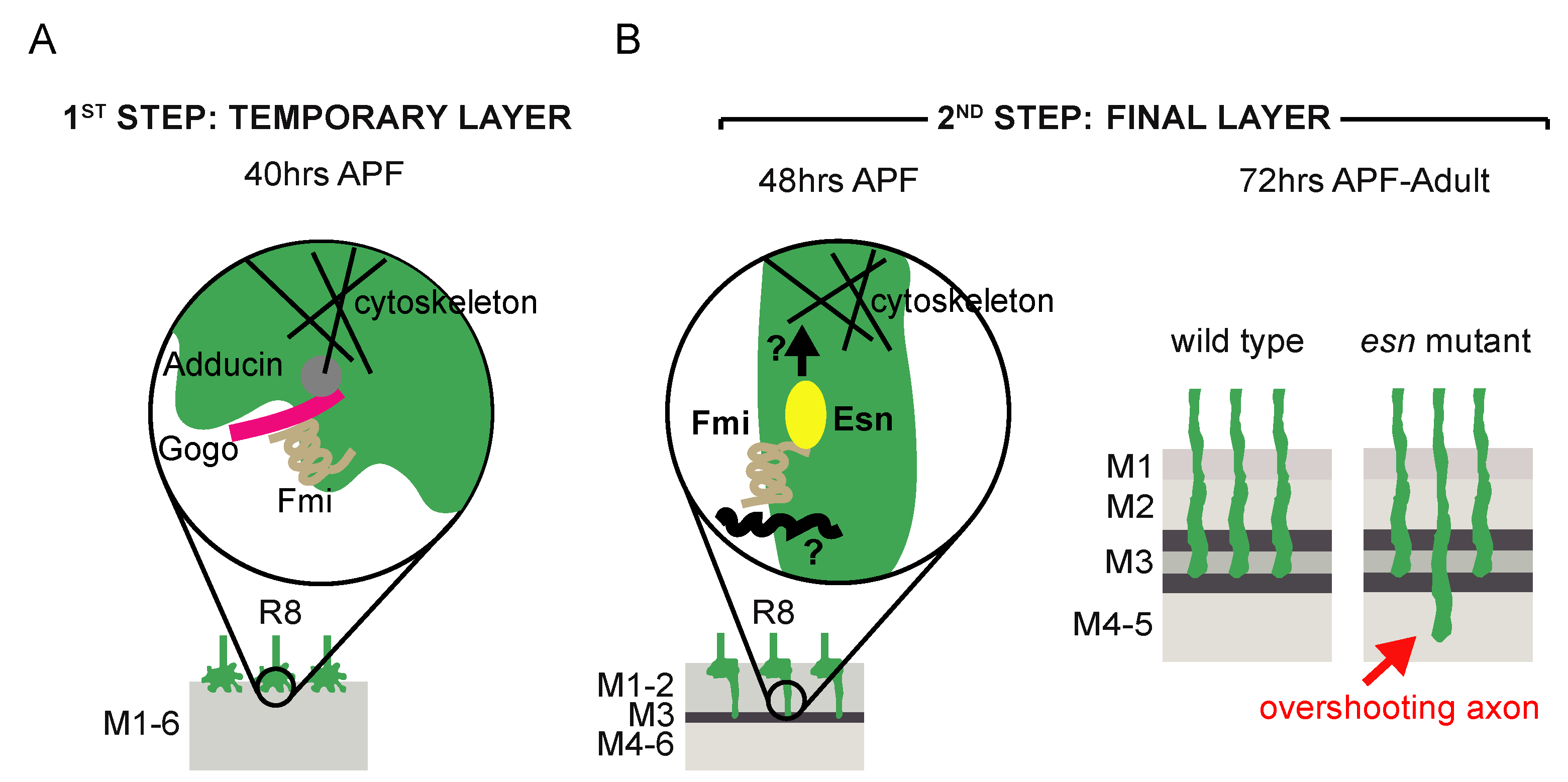
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Kolodkin, A.L.; Hiesinger, P.R. Wiring visual systems: Common and divergent mechanisms and principles. Curr. Opin. Neurobiol. 2017, 42, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Millard, S.S.; Pecot, M.Y. Strategies for assembling columns and layers in the Drosophila visual system. Neural Dev. 2018, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Plazaola-Sasieta, H.; Fernández-Pineda, A.; Zhu, Q.; Morey, M. Untangling the wiring of the Drosophila visual system: Developmental principles and molecular strategies. J. Neurogenet. 2017, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanes, J.R.; Zipursky, S.L. Design Principles of Insect and Vertebrate Visual Systems. Neuron 2010, 66, 15–36. [Google Scholar] [CrossRef] [Green Version]
- Balaskas, N.; Abbott, L.F.; Jessell, T.M.; Ng, D. Positional Strategies for Connection Specificity and Synaptic Organization in Spinal Sensory-Motor Circuits. Neuron 2019, 102, 1143–1156. [Google Scholar] [CrossRef]
- Hassan, B.A.; Hiesinger, P.R. Beyond Molecular Codes: Simple Rules to Wire Complex Brains. Cell 2015, 163, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Petrovic, M.; Hummel, T. Temporal identity in axonal target layer recognition. Nature 2008, 456, 800–803. [Google Scholar] [CrossRef]
- Roberts, A.; Conte, D.; Hull, M.; Merrison-Hort, R.; al Azad, A.K.; Buhl, E.; Soffe, S.R. Can simple rules control development of a pioneer vertebrate neuronal network generating behavior? J. Neurosci. 2014, 34, 608–621. [Google Scholar] [CrossRef] [Green Version]
- Sanes, J.R.; Zipursky, S.L. Synaptic Specificity, Recognition Molecules, and Assembly of Neural Circuits. Cell 2020, 181, 536–556. [Google Scholar] [CrossRef]
- Morey, M.; Yee, S.K.; Herman, T.; Nern, A.; Blanco, E.; Zipursky, S.L. Coordinate control of synaptic-layer specificity and rhodopsins in photoreceptor neurons. Nature 2008, 456, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Zhang, K.X.; Pecot, M.Y.; Nagarkar-Jaiswal, S.; Lee, P.T.; Takemura, S.Y.; Zipursky, S.L. Ig Superfamily Ligand and Receptor Pairs Expressed in Synaptic Partners in Drosophila. Cell 2015, 163, 1756–1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ting, C.Y.; Yonekura, S.; Chung, P.; Hsu, S.N.; Robertson, H.M.; Chiba, A.; Lee, C.H. Drosophila N-cadherin functions in the first stage of the two-stage layer-selection process of R7 photoreceptor afferents. Development 2005, 132, 953–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akin, O.; Lawrence Zipursky, S. Frazzled promotes growth cone attachment at the source of a Netrin gradient in the Drosophila visual system. eLife 2016, 5, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Clandinin, T.R.; Lee, C.H.; Chen, P.L.; Meinertzhagen, I.A.; Zipursky, S.L. The protocadherin Flamingo is required for axon target selection in the Drosophila visual system. Nat. Neurosci. 2003, 6, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Senti, K.; Usui, T.; Boucke, K.; Greber, U.; Uemura, T.; Dickson, B. Flamingo Regulates R8 Axon-Axon and Axon-Target Interactions in the Drosophila Visual System. Curr. Biol. 2003, 13, 828–832. [Google Scholar] [CrossRef] [Green Version]
- Tomasi, T.; Hakeda-Suzuki, S.; Ohler, S.; Schleiffer, A.; Suzuki, T. The Transmembrane Protein Golden Goal Regulates R8 Photoreceptor Axon-Axon and Axon-Target Interactions. Neuron 2008, 57, 691–704. [Google Scholar] [CrossRef] [Green Version]
- Hakeda-Suzuki, S.; Berger-Müller, S.; Tomasi, T.; Usui, T.; Horiuchi, S.; Uemura, T.; Suzuki, T. Golden Goal collaborates with Flamingo in conferring synaptic-layer specificity in the visual system. Nat. Neurosci. 2011, 14, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Takechi, H.; Hakeda-Suzuki, S.; Nitta, Y.; Ishiwata, Y.; Sato, M.; Sugie, A.; Suzuki, T. Glial insulin regulates cooperative or antagonistic Golden goal/Flamingo interactions during photoreceptor axon navigation. BioRxiv 2020. [Google Scholar] [CrossRef]
- Gubb, D.; Green, C.; Huen, D.; Coulson, D.; Johnson, G.; Tree, D.; Collier, S.; Roote, J. The balance between isoforms of the Prickle LIM domain protein is critical for planar polarity in Drosophila imaginal discs. Genes Dev. 1999, 13, 2315–2327. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, D.; Horiuchi, S.Y.; Shimono, K.; Usui, T.; Uemura, T. The seven-pass transmembrane cadherin Flamingo controls dendritic self-avoidance via its binding to a LIM domain protein, Espinas, in Drosophila sensory neurons. Genes Dev. 2011, 25, 1982–1996. [Google Scholar] [CrossRef] [Green Version]
- Nagarkar-Jaiswal, S.; Deluca, S.Z.; Lee, P.T.; Lin, W.W.; Pan, H.; Zuo, Z.; Bellen, H.J. A genetic toolkit for tagging intronic MiMIC containing genes. eLife 2015, 4, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Nagarkar-Jaiswal, S.; Lee, P.T.; Campbell, M.E.; Chen, K.; Anguiano-Zarate, S.; Cantu Gutierrez, M.; Bellen, H.J. A library of MiMICs allows tagging of genes and reversible, spatial and temporal knockdown of proteins in Drosophila. eLife 2015, 4, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Thibault, S.T.; Singer, M.A.; Miyazaki, W.Y.; Milash, B.; Dompe, N.A.; Singh, C.M.; Buchholz, R.; Demsky, M.; Margolis, J. A complementary transposon tool kit for Drosophila melanogaster using P and piggyBac. Nat. Genet. 2004, 36, 283–287. [Google Scholar] [CrossRef]
- Bazigou, E.; Apitz, H.; Johansson, J.; Lorén, C.E.; Hirst, E.M.A.; Chen, P.L.; Salecker, I. Anterograde Jelly belly and Alk Receptor Tyrosine Kinase Signaling Mediates Retinal Axon Targeting in Drosophila. Cell 2007, 128, 961–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usui, T.; Shima, Y.; Shimada, Y.; Hirano, S.; Burgess, R.W.; Schwarz, T.L.; Takeichi, M.; Uemura, T. Flamingo, a seven-pass transmembrane cadherin, regulates planar cell polarity under the control of Frizzled. Cell 1999, 98, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Bischof, J.; Maeda, R.K.; Hediger, M.; Karch, F.; Basler, K. An optimized transgenesis system for Drosophila using germ-line-specific phiC31 integrases. Proc. Natl. Acad. Sci. USA 2007, 104, 3312–3317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Cardona, A. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Venken, K.J.; Schulze, K.L.; Haelterman, N.A.; Pan, H.; He, Y.; Evans-Holm, M.; Carlson, J.W.; Levis, R.W.; Spradling, A.C.; Hoskins, R.A.; et al. MiMIC: A highly versatile transposon insertion resource for engineering Drosophila melanogaster genes. Nat. Methods 2011, 8, 737–743. [Google Scholar] [CrossRef] [Green Version]
- Jenny, A.; Darken, R.S.; Wilson, P.A.; Mlodzik, M. Prickle and Strabismus form a functional complex to generate a correct axis during planar cell polarity signaling. EMBO J. 2003, 22, 4409–4420. [Google Scholar] [CrossRef] [Green Version]
- Rawls, A.S.; Wolff, T. Strabismus requires Flamingo and Prickle function to regulate tissue polarity in the Drosophila eye. Development 2003, 130, 1877–1887. [Google Scholar] [CrossRef] [Green Version]
- Mencarelli, C.; Pichaud, F. Orthodenticle Is Required for the Expression of Principal Recognition Molecules That Control Axon Targeting in the Drosophila Retina. PLoS Genet. 2015, 11, e1005303. [Google Scholar] [CrossRef] [PubMed]
- Ohler, S.; Hakeda-Suzuki, S.; Suzuki, T. Hts, the Drosophila homologue of Adducin, physically interacts with the transmembrane receptor Golden goal to guide photoreceptor axons. Dev. Dyn. 2011, 240, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Strutt, H.; Strutt, D. Differential stability of Flamingo protein complexes underlies the establishment of planar polarity. Curr. Biol. 2008, 18, 1555–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, J. Wnt/PCP proteins regulate stereotyped axon branch extension in Drosophila. Development 2012, 139, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, K.; Sato, M.; Tabata, T. The Wnt5/planar cell polarity pathway regulates axonal development of the Drosophila mushroom body neuron. J. Neurosci. 2011, 31, 4944–4954. [Google Scholar] [CrossRef] [Green Version]
- Mrkusich, E.M.; Flanagan, D.J.; Whitington, P.M. The core planar cell polarity gene prickle interacts with flamingo to promote sensory axon advance in the Drosophila embryo. Dev. Biol. 2011, 358, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Ehaideb, S.N.; Iyengar, A.; Ueda, A.; Iacobucci, G.J.; Cranston, C.; Bassuk, A.G.; Gubb, D.; Axelrod, J.D.; Gunawardena, S.; Wu, C.F.; et al. Prickle modulates microtubule polarity and axonal transport to ameliorate seizures in flies. Proc. Natl. Acad. Sci. USA. 2014, 111, 11187–11192. [Google Scholar] [CrossRef] [Green Version]
- Pecot, M.Y.; Chen, Y.; Akin, O.; Chen, Z.; Tsui, C.Y.; Zipursky, S.L. Sequential axon-derived signals couple target survival and layer specificity in the Drosophila visual system. Neuron 2014, 82, 320–333. [Google Scholar] [CrossRef] [Green Version]
- Timofeev, K.; Joly, W.; Hadjieconomou, D.; Salecker, I. Localized Netrins act as positional cues to control layer-specific targeting of photoreceptor axons in Drosophila. Neuron 2012, 75, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, A.; Ertekin, D.; Lee, C.H.; Hummel, T. Birth order dependent growth cone segregation determines synaptic layer identity in the Drosophila visual system. eLife 2016, 5, e13715. [Google Scholar] [CrossRef] [Green Version]
- Shinza-Kameda, M.; Takasu, E.; Sakurai, K.; Hayashi, S.; Nose, A. Regulation of layer-specific targeting by reciprocal expression of a cell adhesion molecule, Capricious. Neuron 2006, 49, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger-Müller, S.; Sugie, A.; Takahashi, F.; Tavosanis, G.; Hakeda-Suzuki, S.; Suzuki, T. Assessing the role of cell-surface molecules in central synaptogenesis in the Drosophila visual system. PLoS ONE 2013, 8, e83732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, M.; Katoh, M. Identification and characterization of human PRICKLE1 and PRICKLE2 genes as well as mouse Prickle1 and Prickle2 genes homologous to Drosophila tissue polarity gene prickle. Int. J. Mol. Med. 2003, 11, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Tissir, F.; De-Backer, O.; Goffinet, A.M.; Lambert de Rouvroit, C. Developmental expression profiles of Celsr (Flamingo) genes in the mouse. Mech. Dev. 2002, 112, 157–160. [Google Scholar] [CrossRef]
- Scheffer, L.K.; Meinertzhagen, I.A. The Fly Brain Atlas. Annu. Rev. Cell Dev. Biol. 2019, 35, 637–653. [Google Scholar] [CrossRef]
- Deng, B.; Li, Q.; Liu, X.; Cao, Y.; Li, B.; Qian, Y.; Xu, R.; Mao, R.; Zhou, E.; Zhang, W.; et al. Chemoconnectomics: Mapping Chemical Transmission in Drosophila. Neuron 2019, 101, 876–893. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Pineda, A.; Monge-Asensio, M.; Rios, M.; Morey, M. The Cytoplasmic LIM Domain Protein Espinas Contributes to Photoreceptor Layer Selection in the Visual System. Biology 2020, 9, 466. https://doi.org/10.3390/biology9120466
Fernández-Pineda A, Monge-Asensio M, Rios M, Morey M. The Cytoplasmic LIM Domain Protein Espinas Contributes to Photoreceptor Layer Selection in the Visual System. Biology. 2020; 9(12):466. https://doi.org/10.3390/biology9120466
Chicago/Turabian StyleFernández-Pineda, Alejandra, Martí Monge-Asensio, Martín Rios, and Marta Morey. 2020. "The Cytoplasmic LIM Domain Protein Espinas Contributes to Photoreceptor Layer Selection in the Visual System" Biology 9, no. 12: 466. https://doi.org/10.3390/biology9120466
APA StyleFernández-Pineda, A., Monge-Asensio, M., Rios, M., & Morey, M. (2020). The Cytoplasmic LIM Domain Protein Espinas Contributes to Photoreceptor Layer Selection in the Visual System. Biology, 9(12), 466. https://doi.org/10.3390/biology9120466