Cuttlefish Buoyancy in Response to Food Availability and Ocean Acidification






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Collection and CO2 Acclimation Conditions
2.3. Sampling Endpoints
2.4. Sampling and Analysis of the Cuttlebone
2.5. Statistical Analysis
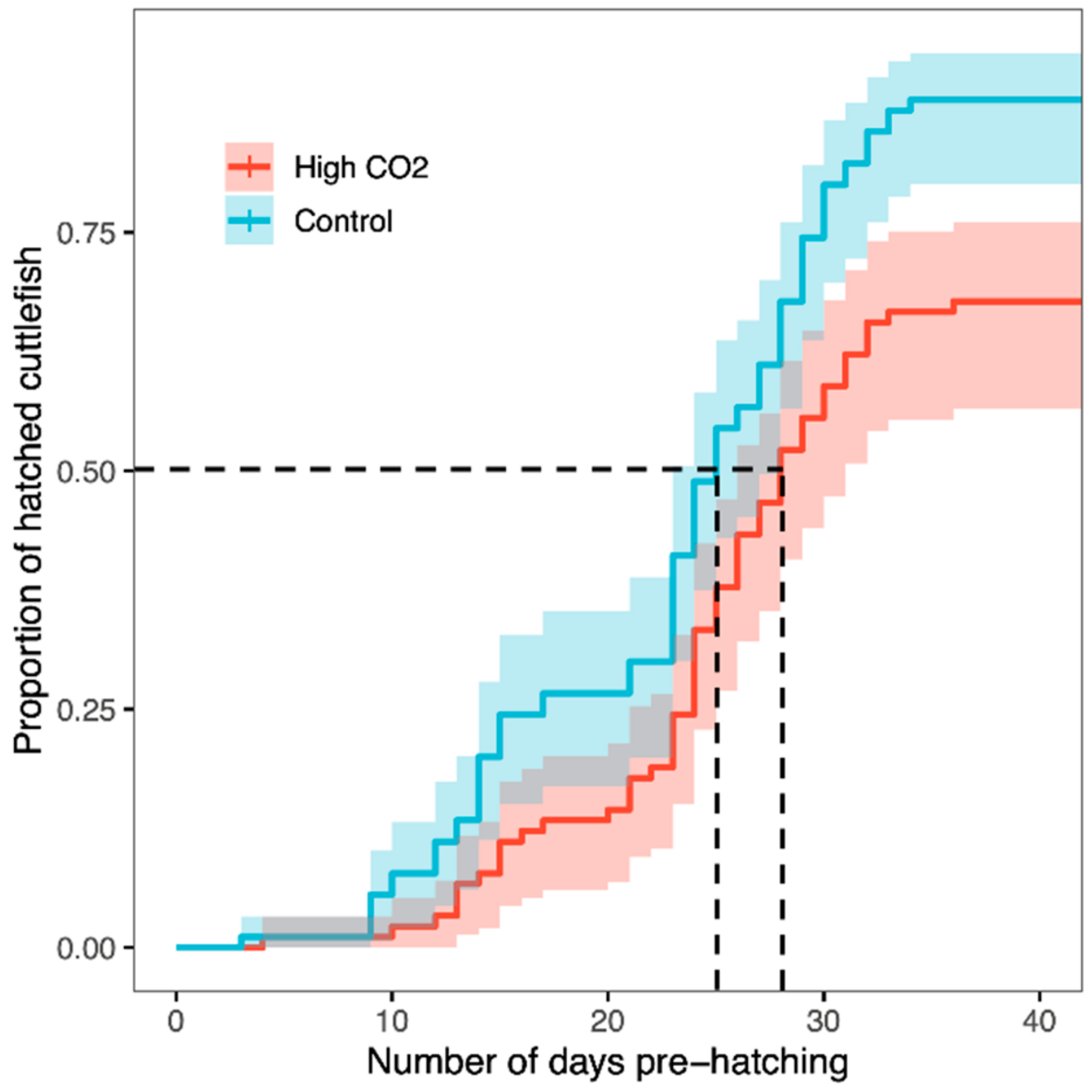
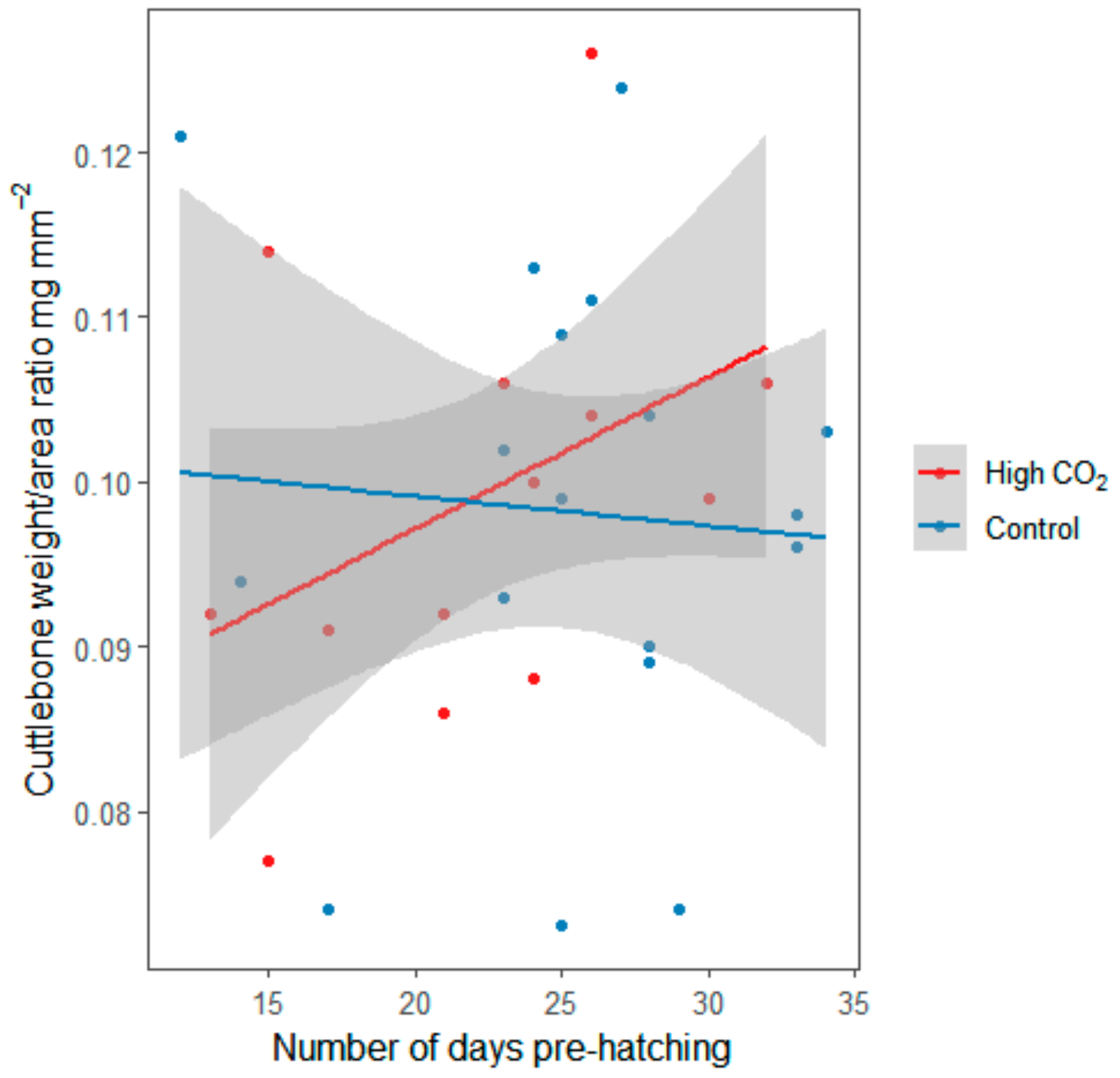
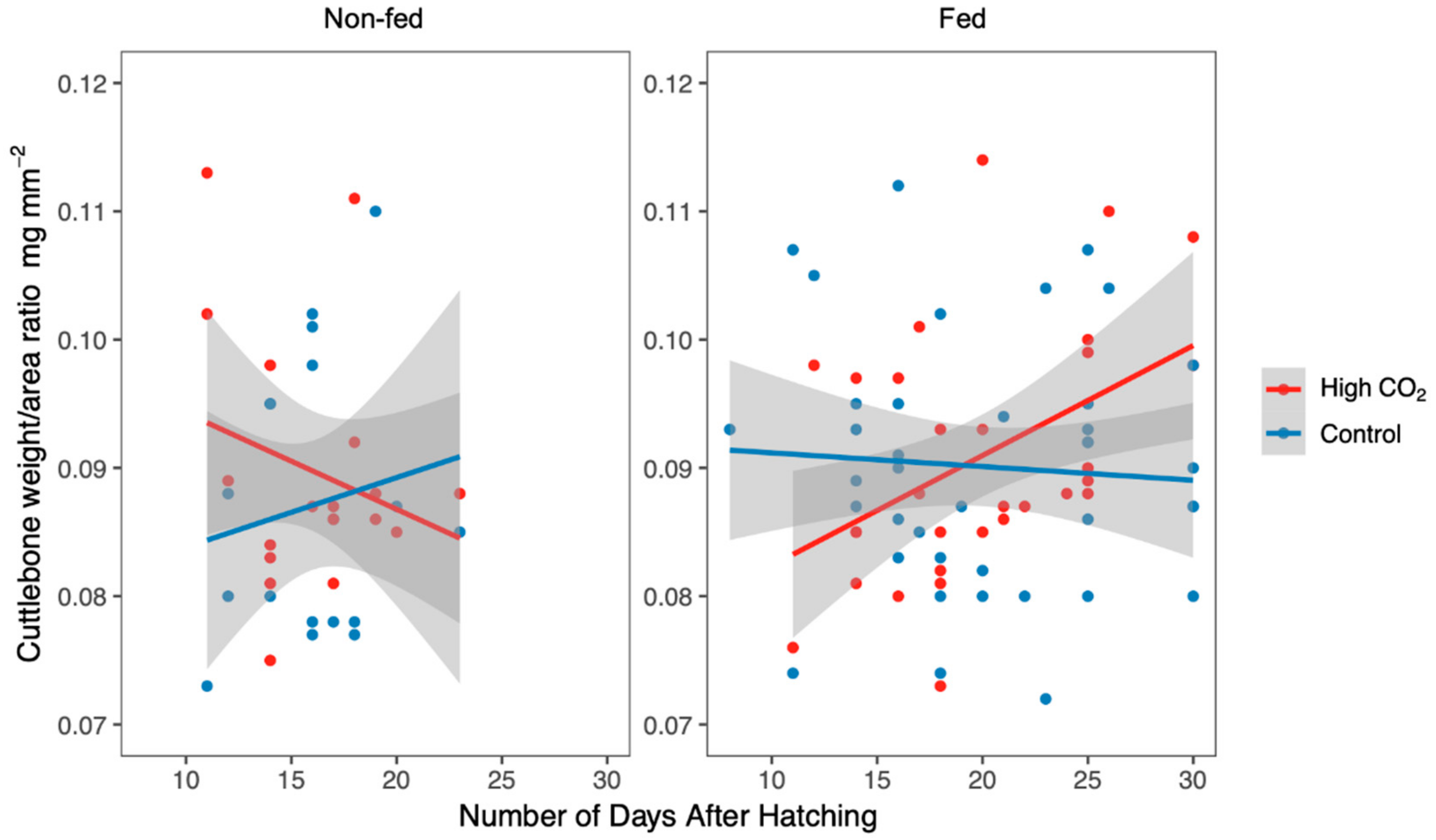
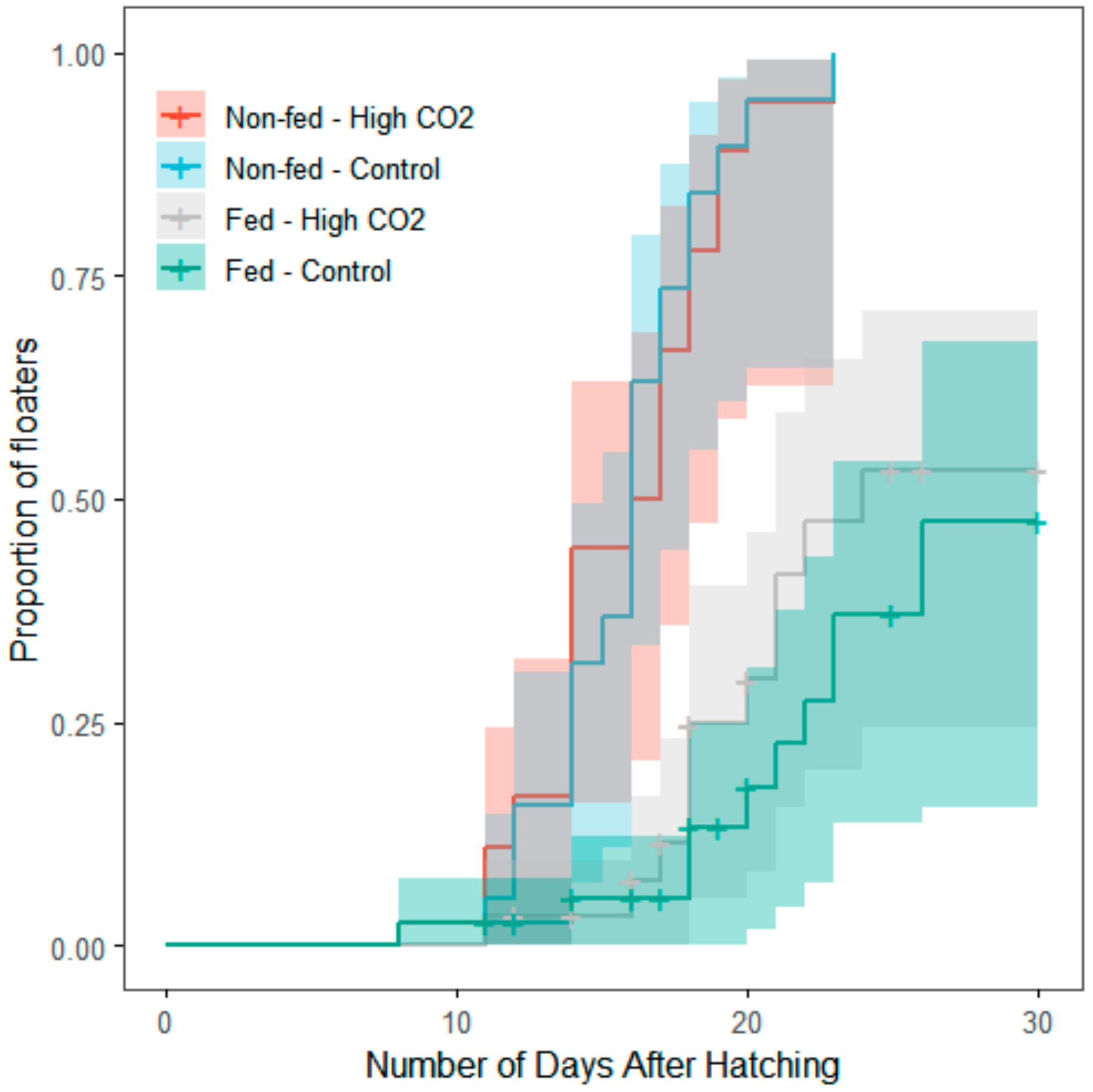
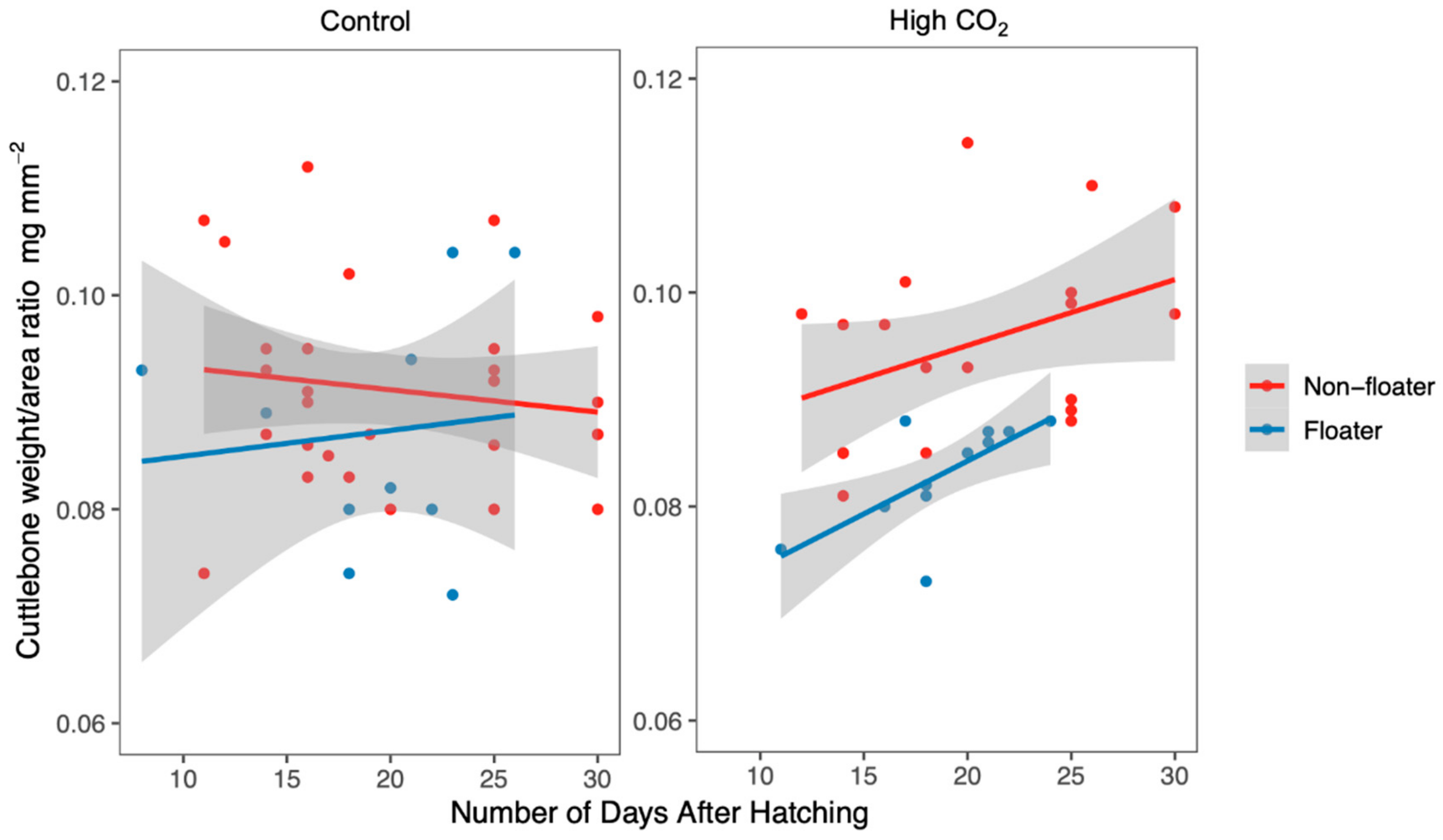
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Data Availability
References
- IPCC. Climate Change 2013 The Physical Science Basis Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Allen, S.K., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; ISBN 9781107661820. [Google Scholar]
- Bruno, J.F.; Harley, C.D.G.; Burrows, M.T. Climate change in marine communities. In Marine Community Ecology and Conservation; Bertness, M.D., Bruno, J.F., Silliman, B.R., Stachowicz, J.J., Eds.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2014; pp. 427–447. [Google Scholar]
- NOAA. Trends in Atmospheric Carbon Dioxide. In Earth Syst. Res. Lab. Glob. Monit. Lab. Available online: https://www.esrl.noaa.gov/gmd/ccgg/trends/ (accessed on 6 March 2020).
- Thor, P.; Dupont, S. Ocean Acidification. In Handbook on Marine Environment Protection; Salomon, M., Markus, T., Eds.; Springer: Cham, Switzerland, 2018; pp. 375–394. ISBN 9783319601564. [Google Scholar]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.C.; Feely, R.A.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F.; et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Fabry, V.J.; Seibel, B.A.; Feely, R.A.; Orr, J.C. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES J. Mar. Sci. 2008, 65, 414–432. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.N.; Singh, G.G. Meta-analysis reveals negative yet variable effects of ocean acidification on marine organisms. Ecol. Lett. 2010, 13, 1419–1434. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, J.; Buddemeier, R.W. Ocean biogeochemistry: Calcification and CO2. Nature 2000, 407, 311–313. [Google Scholar] [CrossRef]
- Waldbusser, G.G.; Voigt, E.P.; Bergschneider, H.; Green, M.A.; Newell, R.I.E. Biocalcification in the Eastern oyster (Crassostrea virginica) in relation to long-term trends in Chesapeake Bay pH. Estuaries Coasts 2011, 34, 221–231. [Google Scholar] [CrossRef]
- Kleypas, J.A.; Buddemeier, R.W.; Archer, D.; Gattuso, J.P.; Langdon, C.; Opdyke, B.N. Geochemical consequences of increased atmospheric carbon dioxide on coral reefs. Science 1999, 284, 118–120. [Google Scholar] [CrossRef]
- Riebesell, U.; Zondervan, I.; Rost, B.; Tortell, P.D.; Zeebe, R.E.; Morel, F.M.M. Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature 2000, 407, 364–366. [Google Scholar] [CrossRef] [Green Version]
- Zondervan, I.; Zeebe, R.E.; Rost, B.; Riebesell, U. Decreasing marine biogenic calcification: A negative feedback on rising atmospheric pCO2. Glob. Biogeochem. Cycles 2001, 15, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Shirayama, Y.; Thornton, H. Effect of increased atmospheric CO2 on shallow water marine benthos. J. Geophys. Res. 2005, 110, 1–7. [Google Scholar] [CrossRef]
- Michaelidis, B.; Ouzounis, C.; Paleras, A.; Pörtner, H.O. Effects of long-term moderate hypercapnia on acid-base balance and growth rate in marine mussels Mytilus galloprovincialis. Mar. Ecol. Prog. Ser. 2005, 293, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazeau, F.; Quiblier, C.; Jansen, J.M.; Gattuso, J.P.; Middelburg, J.J.; Heip, C.H.R. Impact of elevated CO2 on shellfish calcification. Geophys. Res. Lett. 2007, 34, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Dupont, S.; Ortega-Martínez, O.; Thorndyke, M. Impact of near-future ocean acidification on echinoderms. Ecotoxicology 2010, 19, 449–462. [Google Scholar] [CrossRef]
- Cattano, C.; Claudet, J.; Domenici, P.; Milazzo, M. Living in a high CO2 world: A global meta-analysis shows multiple trait-mediated fish responses to ocean acidification. Ecol. Monogr. 2018, 88, 320–335. [Google Scholar] [CrossRef]
- Munday, P.L.; Jarrold, M.D.; Nagelkerken, I. Ecological effects of elevated CO2 on marine and freshwater fishes: From individual to community effects. In Fish Physiology; Grosell, M., Munday, P.L., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2019; Volume 37, pp. 323–368. ISBN 9780128176092. [Google Scholar]
- Melzner, F.; Mark, F.C.; Seibel, B.A.; Tomanek, L. Ocean Acidification and Coastal Marine Invertebrates: Tracking CO2 Effects from Seawater to the Cell. Ann. Rev. Mar. Sci. 2020, 12, 499–523. [Google Scholar] [CrossRef]
- Clark, T.D.; Raby, G.D.; Roche, D.G.; Binning, S.A.; Speers-Roesch, B.; Jutfelt, F.; Sundin, J. Ocean acidification does not impair the behaviour of coral reef fishes. Nature 2020, 577, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Spady, B.L.; Munday, P.L.; Watson, S.A. Predatory strategies and behaviours in cephalopods are altered by elevated CO2. Glob. Chang. Biol. 2018, 24, 2585–2596. [Google Scholar] [CrossRef]
- Spady, B.L.; Watson, S.A.; Chase, T.J.; Munday, P.L. Projected near-future CO2 levels increase activity and alter defensive behaviours in the tropical squid Idiosepius pygmaeus. Biol. Open 2014, 3, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Spady, B.L.; Munday, P.L.; Watson, S.-A. Elevated seawater pCO2 affects reproduction and embryonic development in the pygmy squid, Idiosepius pygmaeus. Mar. Environ. Res. 2019, 153, 104812. [Google Scholar] [CrossRef]
- Kaplan, M.B.; Mooney, T.A.; McCorkle, D.C.; Cohen, A.L. Adverse Effects of Ocean Acidification on Early Development of Squid (Doryteuthis pealeii). PLoS ONE 2013, 8, e63714. [Google Scholar] [CrossRef] [Green Version]
- Zakroff, C.; Mooney, T.A.; Berumen, M.L. Dose-dependence and small-scale variability in responses to ocean acidification during squid, Doryteuthis pealeii, development. Mar. Biol. 2019, 166, 62. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.; Trubenbach, K.; Pimentel, M.S.; Boavida-Portugal, J.; Faleiro, F.; Baptista, M.; Dionisio, G.; Calado, R.; Portner, H.O.; Repolho, T. Differential impacts of ocean acidification and warming on winter and summer progeny of a coastal squid (Loligo vulgaris). J. Exp. Biol. 2014, 217, 518–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutowska, M.A.; Melzner, F. Abiotic conditions in cephalopod (Sepia officinalis) eggs: Embryonic development at low pH and high pCO2. Mar. Biol. 2009, 156, 515–519. [Google Scholar] [CrossRef]
- Rosa, R.; Trubenbach, K.; Repolho, T.; Pimentel, M.; Faleiro, F.; Boavida-Portugal, J.; Baptista, M.; Lopes, V.M.; Dionísio, G.; Leal, M.C.; et al. Lower hypoxia thresholds of cuttlefish early life stages living in a warm acidified ocean. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131695. [Google Scholar] [CrossRef] [PubMed]
- Denton, E.J.; Gilpin-Brown, J.B. The buoyancy of the cuttlefish Sepia officinalis (L.). J. Mar. Biol. Assoc. UK 1961, 41, 319–342. [Google Scholar] [CrossRef] [Green Version]
- Denton, E.J.; Gilpin-Brown, J.B. Buoyancy of the cuttlefish. Nature 1959, 184, 1330–1331. [Google Scholar] [CrossRef]
- Denton, E.J.; Gilpin-Brown, J.B. The distribution of gas and liquid within the cuttlebone. J. Mar. Biol. Assoc. UK 1961, 41, 365–381. [Google Scholar] [CrossRef] [Green Version]
- Denton, E.J.; Gilpin-Brown, J.B.; Howarth, J.V. The osmotic mechanism of the cuttlebone. J. Mar. Biol. Assoc. UK 1961, 41, 351–364. [Google Scholar] [CrossRef] [Green Version]
- Gutowska, M.A.; Pörtner, H.O.; Melzner, F. Growth and calcification in the cephalopod Sepia officinalis under elevated seawater pCO2. Mar. Ecol. Prog. Ser. 2008, 373, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Gutowska, M.A.; Melzner, F.; Pörter, H.O.; Meier, S. Cuttlebone calcification increases during exposure to elevated seawater pCO2 in the cephalopod Sepia officinalis. Mar. Biol. 2010, 157, 1653–1663. [Google Scholar] [CrossRef]
- Dorey, N.; Melzner, F.; Martin, S.; Oberhänsli, F.; Teyssié, J.-L.; Bustamante, P.; Gattuso, J.-P.; Lacoue-Labarthe, T. Ocean acidification and temperature rise: Effects on calcification during early development of the cuttlefish Sepia officinalis. Mar. Biol. 2013, 160, 2007–2022. [Google Scholar] [CrossRef] [Green Version]
- Sigwart, J.D.; Lyons, G.; Fink, A.; Gutowska, M.A.; Murray, D.; Melzner, F.; Houghton, J.D.R.; Hu, M.Y. Elevated pCO2 drives lower growth and yet increased calcification in the early life history of the cuttlefish Sepia officinalis (Mollusca: Cephalopoda). ICES J. Mar. Sci. 2016, 73, 970–980. [Google Scholar] [CrossRef] [Green Version]
- Von Boletzky, S. Effets de la sous-nutrition prolongée sur le développement de la coquille de Sepia officinalis L. (Mollusca, Cephalopoda). Bull. Soc. Zool. Fr. 1974, 99, 667–673. (In French) [Google Scholar]
- Von Boletzky, S. Growth and life-span of Sepia officinalis under artificial conditions (Mollusca, Cephalopoda). Rapp. Proces-Verbaux Reun. Conceil Int. Explor. Sci. Mediterr. 1979, 25, 159–168. [Google Scholar]
- Maes, J.; Van De Putte, A.; Hecq, J.H.; Volckaert, F.A.M. State-dependent energy allocation in the pelagic Antarctic silverfish Pleuragramma antarcticum: Trade-off between winter reserves and buoyancy. Mar. Ecol. Prog. Ser. 2006, 326, 269–282. [Google Scholar] [CrossRef]
- Dubreuil, J.; Petitgas, P. Energy density of anchovy Engraulis encrasicolus in the Bay of Biscay. J. Fish Biol. 2009, 74, 521–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, R.; Gonzalez, L.; Broitman, B.R.; Garrido, S.; Santos, A.M.P.; Nunes, M.L. Bioenergetics of small pelagic fishes in upwelling systems: Relationship between fish condition, coastal ecosystem dynamics and fisheries. Mar. Ecol. Prog. Ser. 2010, 410, 205–218. [Google Scholar] [CrossRef]
- Sykes, A.V.; Domingues, P.M.; Loyd, M.; Sommerfield, A.; Andrade, J.P. The influence of culture density and enriched environments on the first stage culture of young cuttlefish, Sepia officinalis (Linnaeus, 1758). Aquac. Int. 2003, 11, 531–544. [Google Scholar] [CrossRef]
- Sykes, A.V.; Domingues, P.; Andrade, J.P. European Cuttlefish, Sepia officinalis. In Cephalopod Culture; Iglesias, J., Fuentes, L., Villanueva, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; ISBN 9789401786485. [Google Scholar]
- Fiorito, G.; Affuso, A.; Basil, J.; Cole, A.; de Girolamo, P.; D’angelo, L.; Dickel, L.; Gestal, C.; Grasso, F.; Kuba, M.; et al. Guidelines for the Care and Welfare of Cephalopods in Research—A consensus based on an initiative by CephRes, FELASA and the Boyd Group. Lab. Anim. 2015, 49, 1–90. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Aho, K.; Derryberry, D.; Peterson, T. Model selection for ecologists: The worldviews of AIC and BIC. Ecology 2014, 95, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N. A protocol for conducting and presenting results of regression-type analyses. Methods Ecol. Evol. 2016, 7, 636–645. [Google Scholar] [CrossRef]
- Moura, É.; Pimentel, M.; Santos, C.P.; Sampaio, E.; Pegado, M.R.; Lopes, V.M.; Rosa, R.; Genta, F.A. Cuttlefish Early Development and Behavior Under Future High CO2 Conditions. Front. Physiol. 2019, 10, 975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaire, J. Etude du Développement Embryonnaire de Sepia officinalis L. (Mollusque Céphalopode). Ph.D. Thesis, Université des Sciences et Techniques de Lille, Lille, France, 25 June 1971. (In French). [Google Scholar]
- Hu, M.Y.; Tseng, Y.C.; Stumpp, M.; Gutowska, M.A.; Kiko, R.; Lucassen, M.; Melzner, F. Elevated seawater pCO2 differentially affects branchial acid-base transporters over the course of development in the cephalopod Sepia officinalis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, 1100–1114. [Google Scholar] [CrossRef] [Green Version]
- Ramajo, L.; Pérez-León, E.; Hendriks, I.E.; Marbà, N.; Krause-Jensen, D.; Sejr, M.K.; Blicher, M.E.; Lagos, N.A.; Olsen, Y.S.; Duarte, C.M. Food supply confers calcifiers resistance to ocean acidification. Sci. Rep. 2016, 6, 19374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, N.E.M.; Bernhardt, J.R.; Anderson, K.M.; Harley, C.D.G. Increased food supply mitigates ocean acidification effects on calcification but exacerbates effects on growth. Sci. Rep. 2018, 8, 1–9. [Google Scholar]
- Wood, H.L.; Spicer, J.I.; Widdicombe, S. Ocean acidification may increase calcification rates, but at a cost. Proc. R. Soc. B Biol. Sci. 2008, 275, 1767–1773. [Google Scholar] [CrossRef] [Green Version]
- Melzner, F.; Stange, P.; Trübenbach, K.; Thomsen, J.; Casties, I.; Panknin, U.; Gorb, S.N.; Gutowska, M.A. Food supply and seawater pCO2 impact calcification and internal shell dissolution in the blue mussel mytilus edulis. PLoS ONE 2011, 6, e24223. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, J.; Casties, I.; Pansch, C.; Körtzinger, A.; Melzner, F. Food availability outweighs ocean acidification effects in juvenile Mytilus edulis: Laboratory and field experiments. Glob. Chang. Biol. 2013, 19, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Cominassi, L.; Moyano, M.; Claireaux, G.; Howald, S.; Mark, F.C.; Zambonino-infante, J.; Peck, M.A. Food availability modulates the combined effects of ocean acidification and warming on fish growth. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Kraft, S.; Korn, D.; Klug, C. Patterns of ontogenetic septal spacing in Carboniferous ammonoids. Neues Jahrb. Geol. Palaontologie Abh. 2008, 250, 31–44. [Google Scholar] [CrossRef]
- Tajika, A.; Landman, N.H.; Hoffmann, R.; Lemanis, R.; Morimoto, N.; Ifrim, C.; Klug, C. Chamber volume development, metabolic rates, and selective extinction in cephalopods. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherrard, K.M. Cuttlebone morphology limits habitat depth in eleven species of Sepia (Cephalopoda: Sepiidae). Biol. Bull. 2000, 198, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Von Boletzky, S.; Ward, P.D. Shell implosion depth and implosion morphologies in three species of sepia (Cephalopoda) from the Mediterranean sea. J. Mar. Biol. Assoc. UK 1984, 64, 955–966. [Google Scholar]
- Pansch, C.; Schaub, I.; Havenhand, J.; Wahl, M. Habitat traits and food availability determine the response of marine invertebrates to ocean acidification. Glob. Chang. Biol. 2014, 20, 765–777. [Google Scholar] [CrossRef] [Green Version]
- Rossoll, D.; Bermúdez, R.; Hauss, H.; Schulz, K.G.; Riebesell, U.; Sommer, U.; Winder, M. Ocean acidification-induced food quality deterioration constrains trophic transfer. PLoS ONE 2012, 7, e34737. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otjacques, E.; Repolho, T.; Paula, J.R.; Simão, S.; Baptista, M.; Rosa, R. Cuttlefish Buoyancy in Response to Food Availability and Ocean Acidification. Biology 2020, 9, 147. https://doi.org/10.3390/biology9070147
Otjacques E, Repolho T, Paula JR, Simão S, Baptista M, Rosa R. Cuttlefish Buoyancy in Response to Food Availability and Ocean Acidification. Biology. 2020; 9(7):147. https://doi.org/10.3390/biology9070147
Chicago/Turabian StyleOtjacques, Eve, Tiago Repolho, José Ricardo Paula, Silvia Simão, Miguel Baptista, and Rui Rosa. 2020. "Cuttlefish Buoyancy in Response to Food Availability and Ocean Acidification" Biology 9, no. 7: 147. https://doi.org/10.3390/biology9070147
APA StyleOtjacques, E., Repolho, T., Paula, J. R., Simão, S., Baptista, M., & Rosa, R. (2020). Cuttlefish Buoyancy in Response to Food Availability and Ocean Acidification. Biology, 9(7), 147. https://doi.org/10.3390/biology9070147