Circadian Photoentrainment in Mice and Humans


Abstract
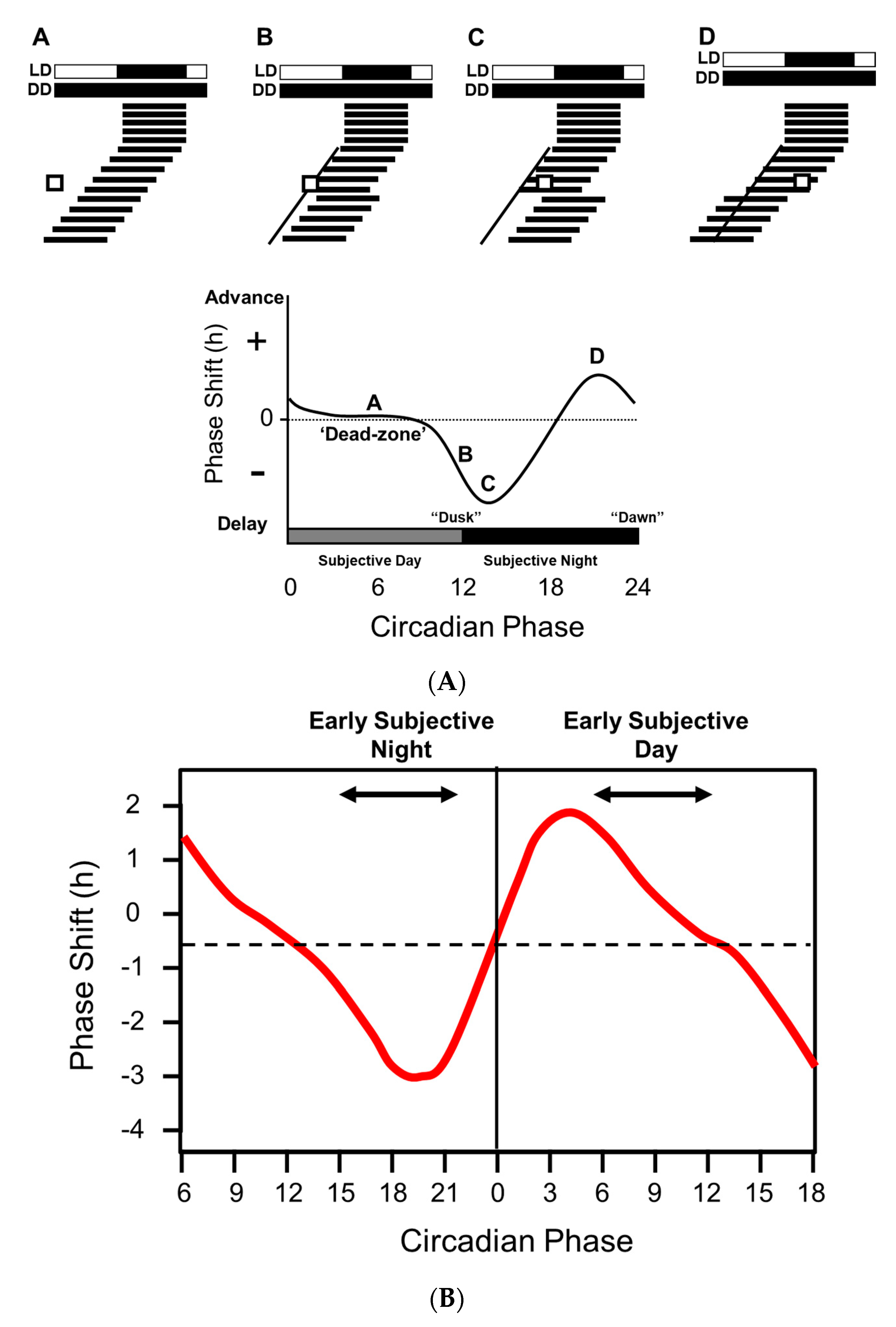
:1. Shedding Light on the Clock—The Phase Response Curve
2. The Discovery and Characterization of the 3rd Retinal Photoreceptor in Mice
2.1. Identification of a 3rd Ocular Photoreceptor
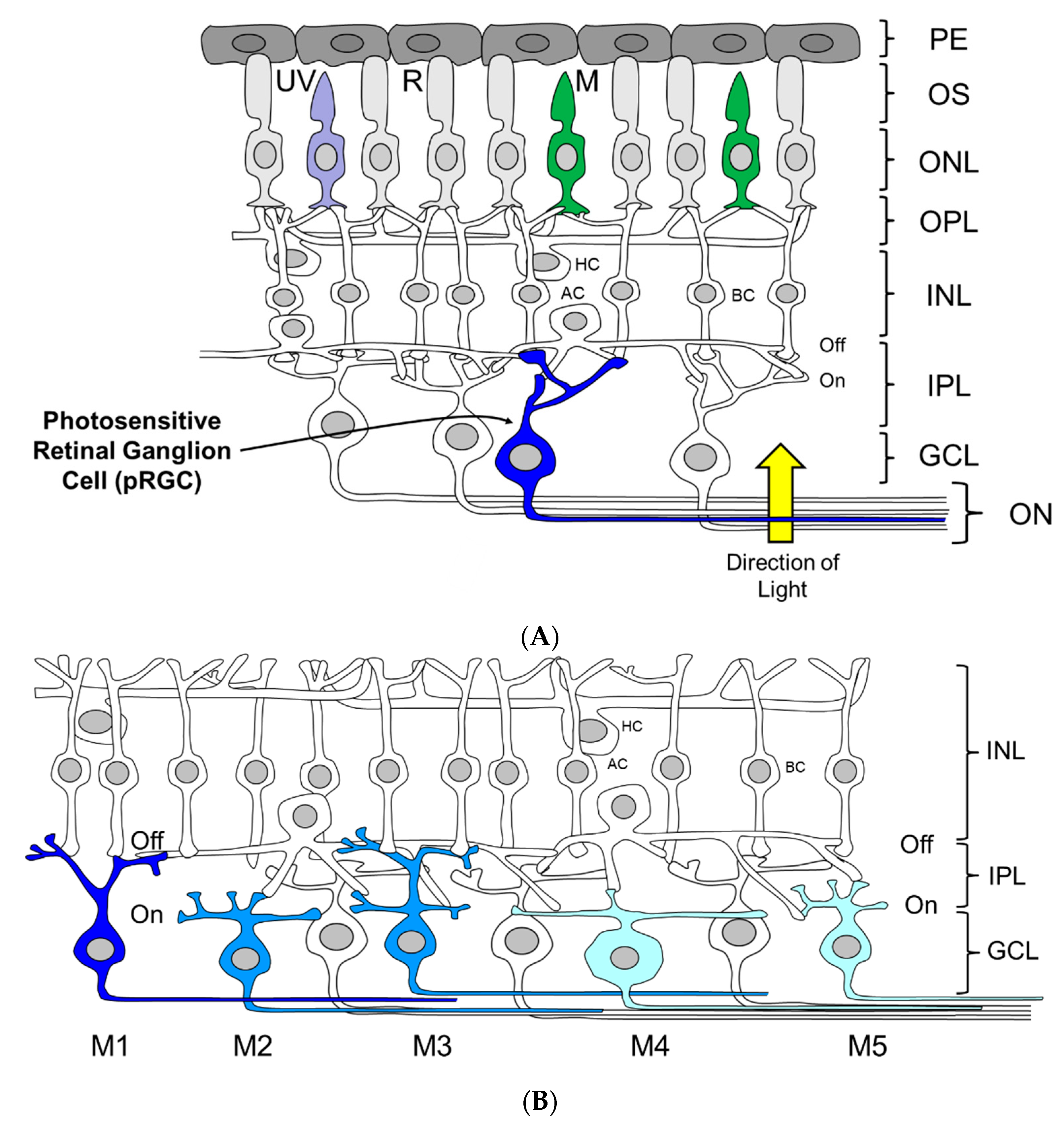
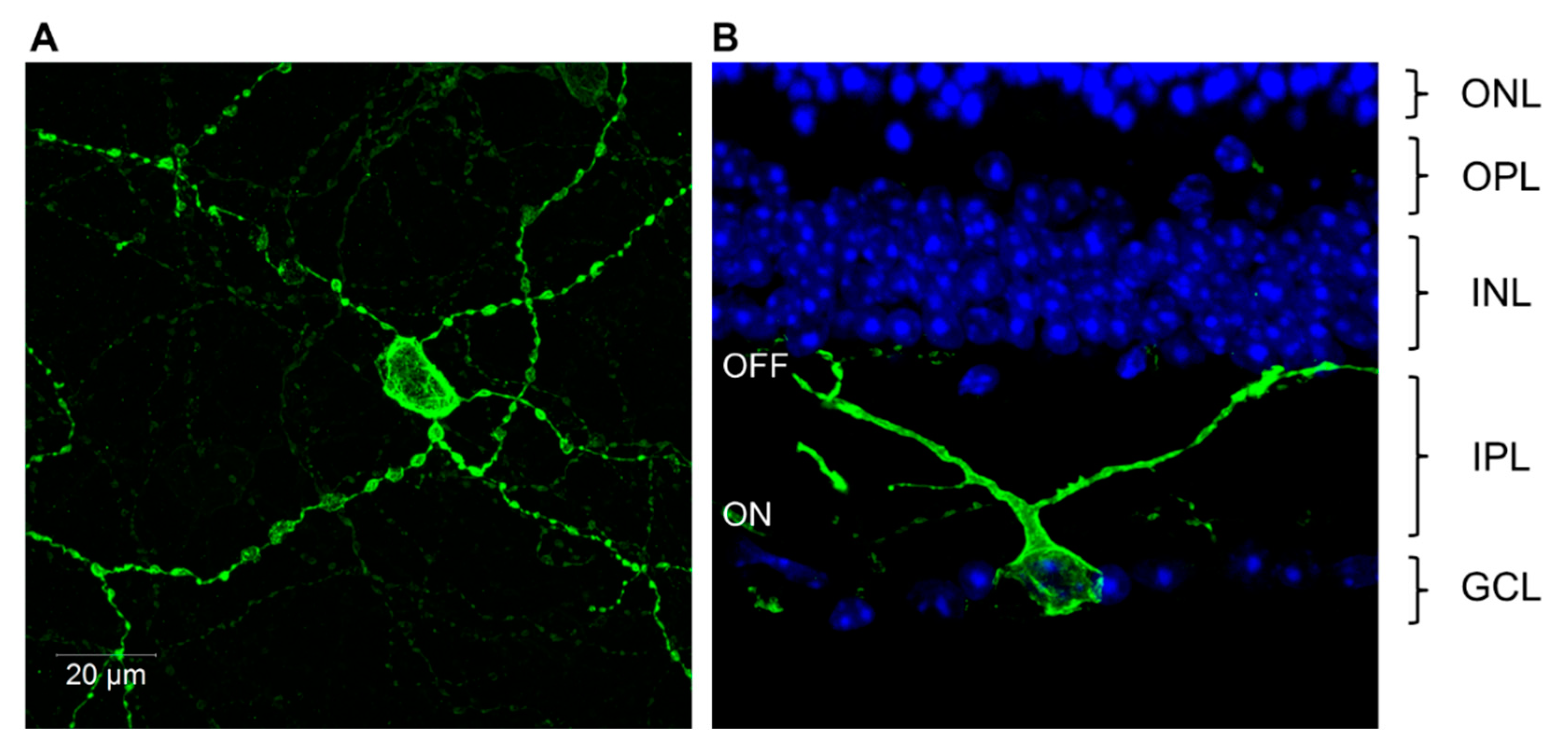
2.2. Identification of Photosensitive Retinal Ganglion Cells (pRGCs)
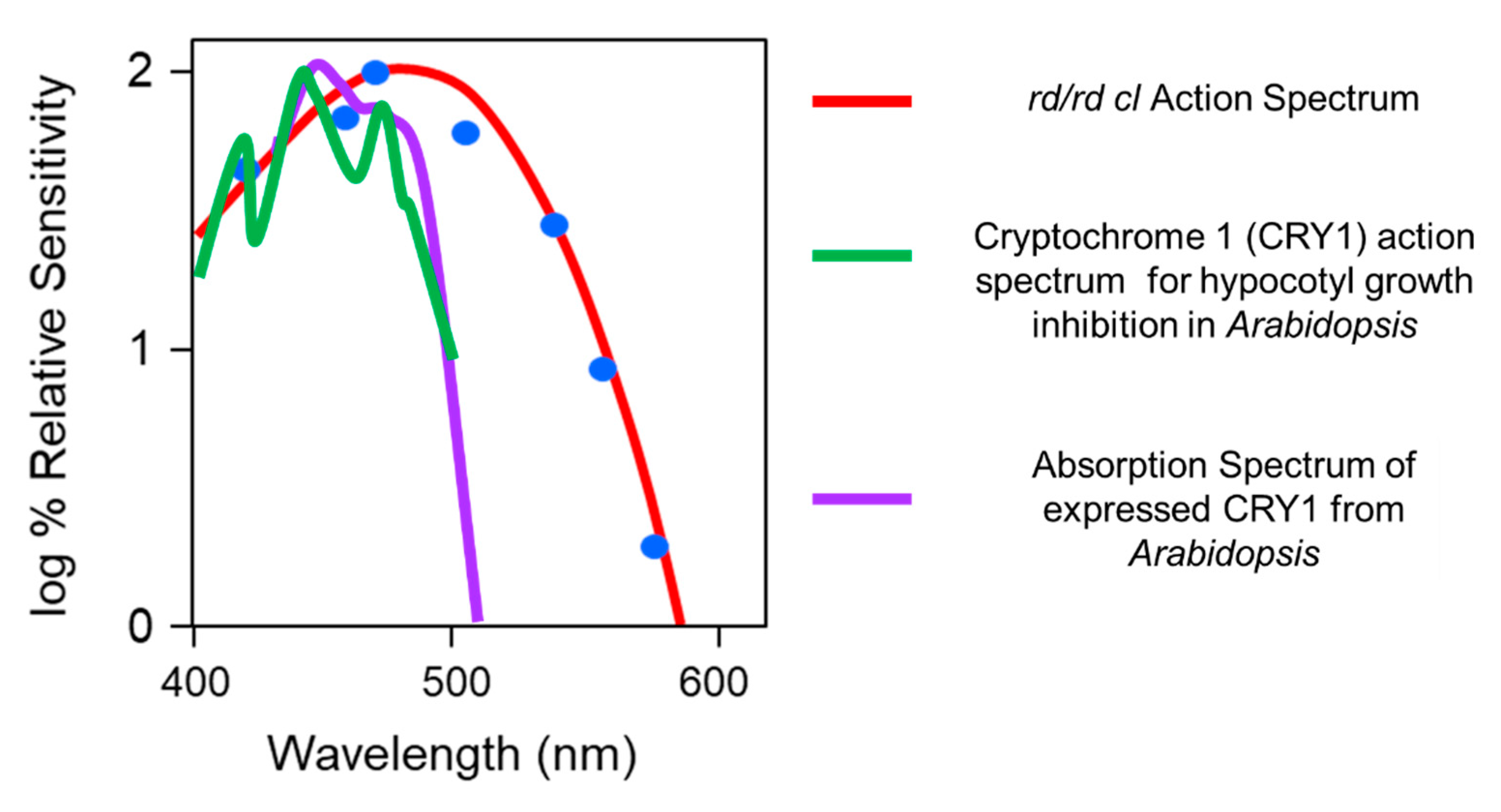
2.3. Defining the Photopigment of the pRGCs Using Action Spectroscopy
2.4. The Identification of Melanopsin (OPN4)
2.5. Melanopsin Expression Studies
2.6. OPN4 and pRGC Complexity
2.7. Diversity of Melanopsin Light Responses
2.8. Rod, Cone, pRGC Interactions at the Level of the SCN
2.9. The Intensity, Duration and Spectrum of Effective Light Stimuli—Ecological Relevance
2.10. Key Conclusions from Studies on Mice
- Light at twilight (dawn and dusk) is the key “zeitgeber” for the entrainment of circadian rhythms.
- The precise form of the phase response curve (PRC) varies but broadly light at dusk delays the clock (start activity later), whilst light at dawn advances the clock (start activity earlier).
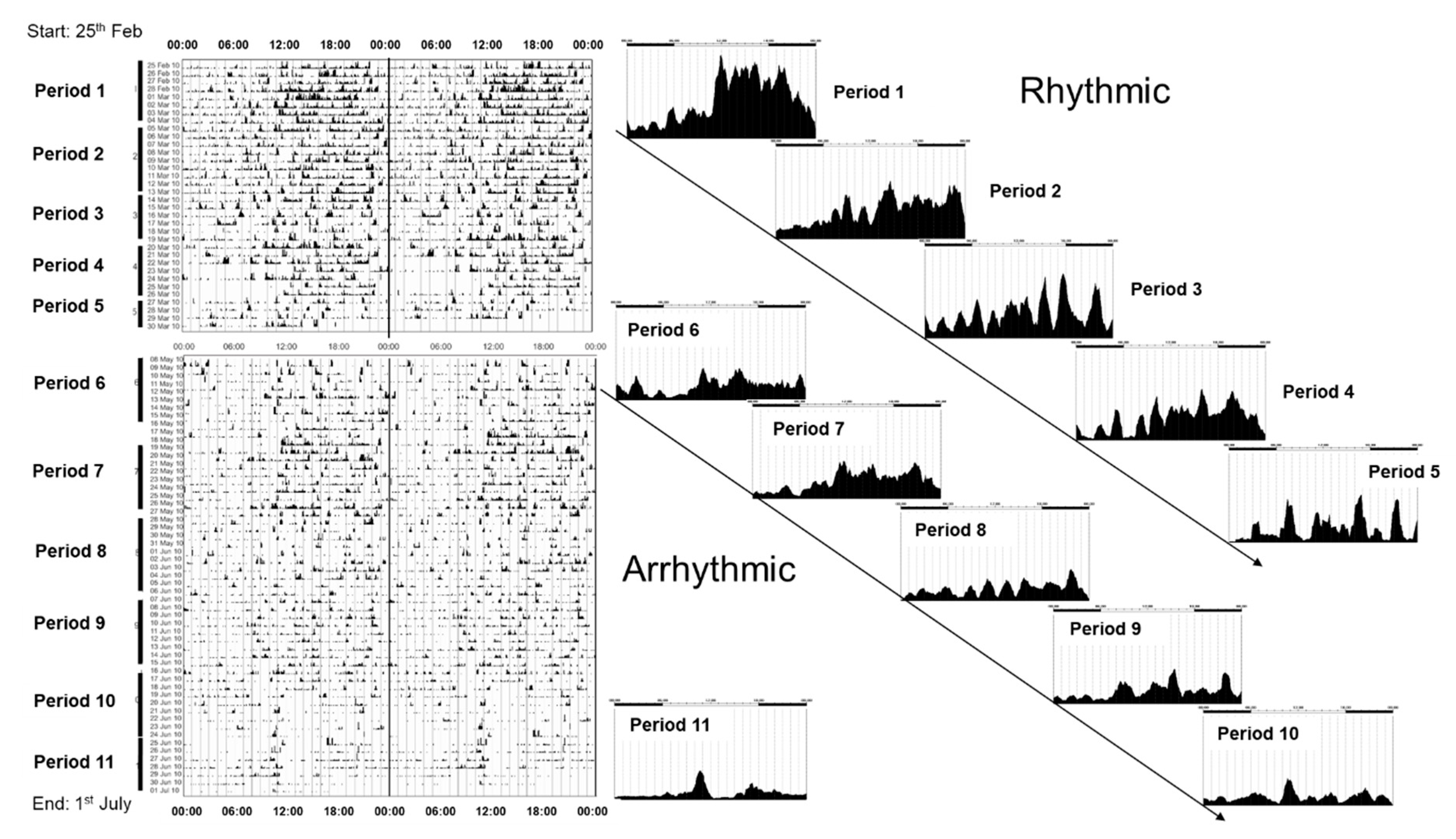
- There is a suggestion that the PRCs of mice and humans differ with regard to the possession of a “dead zone.” However, methodological differences, especially the duration of the light pulses used, may account for these inconsistencies.
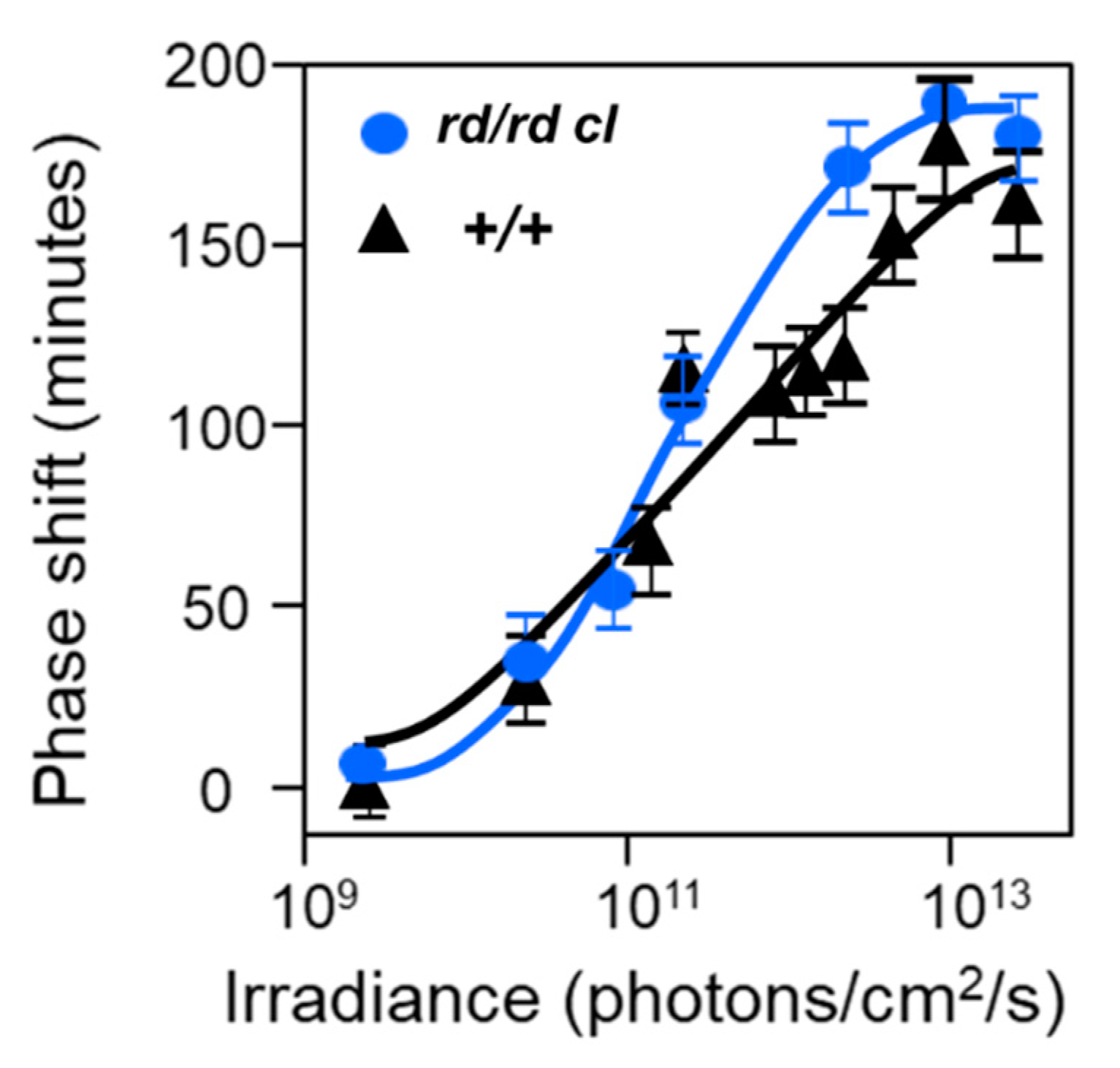
- The thresholds for entrainment vary between mouse strains (Table 1) and illustrate the point that there is variation in circadian photosensitivities within a single species.
- Mice lacking rods and cones show normal circadian entrainment. This finding demonstrated for the first time the existence of a “3rd ocular photoreceptor” within the mammalian eye.
- The 3rd ocular photoreceptor is based upon a network of photosensitive retinal ganglion cells (pRGCs).
- In addition to circadian entrainment multiple irradiance, detection tasks are mediated by the pRGCs.
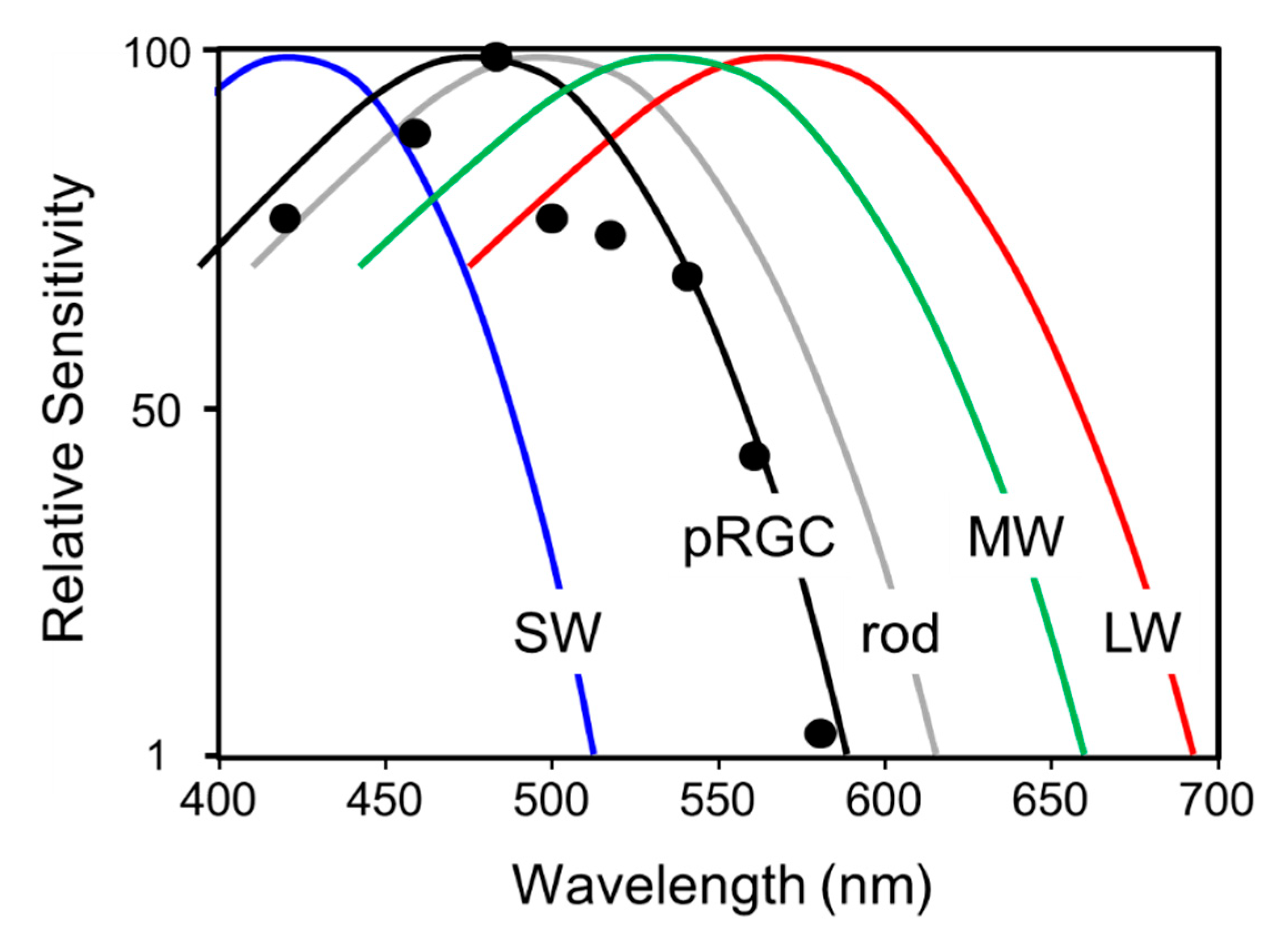
- The photopigment of the pRGCs is melanopsin (OPN4) and has a peak spectral sensitivity in the “blue” part of the spectrum with a λmax close to 480 nm.
- There are at least five different types of pRGCs based upon their anatomy and levels of melanopsin expression. The electrical properties of the pRGCs also vary and in some limited cases specific electrical responses can be linked to a specific pRGC sub-type.
- The single Opn4 gene is alternately spliced, and the long and short isoforms are expressed at different levels in the pRGCs. This adds to the complexity of pRGC signaling.
- Phototransduction in pRGCs results in cellular depolarization and is very different from rod and cone phototransduction which leads to cellular hyperpolarization. Key details regarding pRGC phototransduction remain un-resolved.
- It remains unclear which sub-classes of the pRGCs project to different target regions of the brain and which pRGCs regulate specific behavioral and physiological responses.
- Rods (λmax~498 nm) and cones (M Cone λmax~508 nm; UVS~360 nm) do not project directly to the to pRGCs but modify their endogenous light response via the activation of inner retinal neurons.
- The sensory task of dawn/dusk (twilight) detection is complex in terms of: (1) the light signal itself (irradiance and wavelength); (2) individual exposure to the light signal; and (3) and individual responses to the light signal.
- It seems very likely that rods, cones and pRGCs interact to measure and integrate both the irradiance and wavelength of light at twilight to entrain the circadian system.
- The working hypothesis is that there is an integration of light signals within the pRGCs such that the rods are employed for dim light detection; cones are used for the detection of higher light intensities and for the integration of intermittent light exposure; and the pRGCs provide information regarding bright light over longer durations of exposure.
3. The Effects of Light on the Human Circadian System
3.1. Identifying the 3rd Retinal Photoreceptor in Humans
3.2. The Intensity, Duration and Spectrum of Effective Light Stimuli
3.3. The Impact of Different Light Stimuli on Circadian Entrainment
3.3.1. Field Studies and Natural Light Exposure
3.3.2. Quantitative Measures of Circadian Responses under Artificial Light Stimuli
3.3.3. Quantitative Measures of Circadian Responses Using Colored Light Stimuli
3.3.4. Exposure to Light-Emitting Electronic Devices
3.4. The Impact of Light History
3.5. The Impact of Age on Circadian Photosensitivity
3.6. Key Conclusions from Studies on Humans
- Light at twilight (dawn and dusk) is the key zeitgeber for the entrainment of human circadian rhythms and humans show different phases of entrainment under artificial (usually phase delayed) vs. natural light (usually phase advanced).
- The precise form of the human phase response curve (PRC) is debated but broadly light at dusk delays the clock (starting activity and sleep later the next day), whilst light at dawn advances the clock (start activity and sleep earlier the next day).
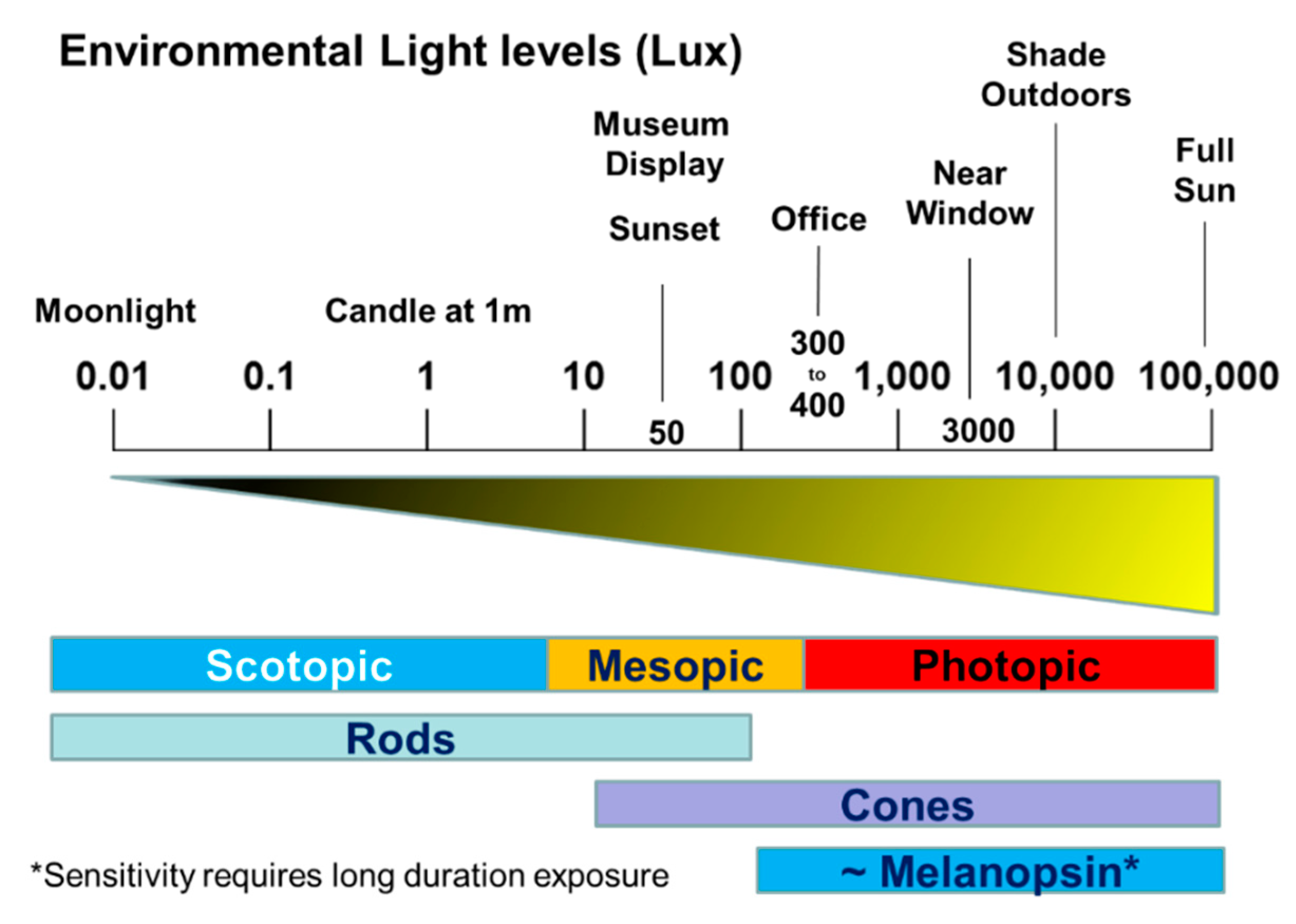
- Compared to mice, humans require light stimuli of a high irradiance (>100’s lux) and of a long duration (>30 min) to achieve entrainment, but the precise irradiance/duration relationships have yet to be defined.
- Humans lacking rods and cones show normal circadian entrainment. This finding demonstrated for the first time the existence of a “3rd ocular photoreceptor” within the human eye.
- On the basis of similarities with mice, the 3rd ocular photoreceptor appears to be based upon a network of photosensitive retinal ganglion cells (pRGCs).
- In addition to circadian entrainment, multiple irradiance detection tasks are mediated by the pRGCs in humans (e.g., alertness, pupil constriction, melatonin suppression).
- An action spectrum in a rodless/coneless individual suggests that the photopigment of the pRGCs is based upon melanopsin with a λmax close to 480 nm. Such results contradict earlier studies suggesting that the λmax was close to 460 nm.
- Emerging anatomical results show that there are multiple types of pRGCs in the human retina. There is no knowledge regarding the function or projections of the different pRGCs.
- There is anatomical evidence that rods (λmax~498 nm); SWS cones (λmax~420 nm); MWS cones (λmax~534 nm); and LWS cones (λmax~564 nm) communicate with the pRGCs via intermediate neurons of the retina. Studies in the primate retina show that these photoreceptors modify the endogenous light responses of the pRGC.
- The sensory thresholds and spectral sensitivities of the different photopigments overlap. As a result, the use of monochromatic light to selectively stimulate a specific photoreceptor channel is not possible. However, the more recent use of silent substitution approaches does provide a possible way forward [271]. Nevertheless, most studies suggest that long duration exposure to “blue” light is the most effective stimulus for entrainment.
- On the basis of behavioral studies it seems very likely that rods, cones and pRGCs interact to measure and integrate both the irradiance and wavelength of light at twilight to entrain the circadian system.
- The working hypothesis, with significant extrapolation from mouse studies, is that there is an integration of light signals within the pRGCs such that the rods are employed for dim light detection; cones are used for the detection of higher light intensities and for the integration of intermittent light exposure; and the pRGCs provide information regarding bright light over extended periods of exposure.
- An individual’s age, prior light history and genetics modify how light defines the phase of entrainment.
4. Future Experiments Relating to Entrainment of the Human Circadian System
4.1. Measuring Effectiveness
- Tau (τ) = the intrinsic period of the clock;
- T = the period of the entraining light/dark cycle;
- φΔ = phase shift needed for τ = T.
4.2. Defining the Optimum Duration, Irradiance, Wavelength and Timing of Artificial Light Stimuli
4.3. Experimental Options and Future Approaches
Author Contributions
Funding
Conflicts of Interest
References
- Pittendrigh, C.S. Temporal organization: Reflections of a Darwinian clock-watcher. Annu. Rev. Physiol. 1993, 55, 16–54. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.H.; Takahashi, J.S. Molecular components of the mammalian circadian clock. Hum. Mol. Genet. 2006, 15, R271–R277. [Google Scholar] [CrossRef]
- Daan, S.; Pittendrigh, C.S. A functional analysis of circadian pacemakers in nocturnal rodents. II. The variabilityof phase response curves. J. Comp. Physiol. 1976, 106, 253–266. [Google Scholar] [CrossRef]
- Khalsa, S.B.; Jewett, M.E.; Cajochen, C.; Czeisler, C.A. A phase response curve to single bright light pulses in human subjects. J. Physiol. 2003, 549, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Pittendrigh, C.S. Circadian rhythms and the circadian organization of living systems. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 159–184. [Google Scholar] [CrossRef] [PubMed]
- Dumont, M.; Carrier, J. Daytime sleep propensity after moderate circadian phase shifts induced with bright light exposure. Sleep 1997, 20, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewett, M.E.; Rimmer, D.W.; Duffy, J.F.; Klerman, E.B.; Kronauer, R.E.; Czeisler, C.A. Human circadian pacemaker is sensitive to light throughout subjective day without evidence of transients. Am. J. Physiol. 1997, 273, R1800–R1809. [Google Scholar] [CrossRef] [Green Version]
- Duffy, J.F.; Dijk, D.J. Getting through to circadian oscillators: Why use constant routines? J. Biol. Rhythm. 2002, 17, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Beersma, D.G.; Daan, S. Strong or weak phase resetting by light pulses in humans? J. Biol. Rhythm. 1993, 8, 340–347. [Google Scholar] [CrossRef]
- Czeisler, C.A.; Kronauer, R.E.; Allan, J.S.; Duffy, J.F.; Jewett, M.E.; Brown, E.N.; Ronda, J.M. Bright light induction of strong (type 0) resetting of the human circadian pacemaker. Science 1989, 244, 1328–1333. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Steiner, R.; Blattner, P.; Oelhafen, P.; Gotz, T.; Cajochen, C. Non-visual effects of light on melatonin, alertness and cognitive performance: Can blue-enriched light keep us alert? PLoS ONE 2011, 6, e16429. [Google Scholar] [CrossRef]
- Mrosovsky, N. Masking: History, definitions, and measurement. Chronobiol. Int. 1999, 16, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Shand, J.; Foster, R.G. The extra-retinal photoreceptors of non-mammalian vertebrates. In Adaptive Mechanisms in the Ecology of Vision; Archer, S., Djamgoz, M., Loew, E.R., Eds.; Kluwer Academic Publishers: Berlin, Germany, 1999; pp. 197–222. [Google Scholar]
- Foster, R.G.; Provencio, I. The regulation of vertebrate biological clocks by light. In Adaptive Mechanisms in the Ecology of Vision; Archer, S., Djamgoz, M., Loew, E.R., Eds.; Chapman and Hall: London, UK, 1999; pp. 223–243. [Google Scholar]
- Halford, S.; Pires, S.S.; Turton, M.; Zheng, L.; Gonzalez-Menendez, I.; Davies, W.L.; Peirson, S.N.; Garcia-Fernandez, J.M.; Hankins, M.W.; Foster, R.G. VA opsin-based photoreceptors in the hypothalamus of birds. Curr. Biol. 2009, 19, 1396–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, W.I.; Turton, M.; Peirson, S.N.; Follett, B.K.; Halford, S.; Garcia-Fernandez, J.M.; Sharp, P.J.; Hankins, M.W.; Foster, R.G. Vertebrate ancient opsin photopigment spectra and the avian photoperiodic response. Biol. Lett. 2012, 8, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.G.; Bellingham, J. Opsins and melanopsins. Curr. Biol. 2002, 12, 243–544. [Google Scholar] [CrossRef] [Green Version]
- Soni, B.G.; Foster, R.G. A novel and ancient vertebrate opsin. FEBS Lett. 1997, 406, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Soni, B.G.; Philp, A.R.; Knox, B.E.; Foster, R.G. Novel retinal photoreceptors. Nature 1998, 394, 27–28. [Google Scholar] [CrossRef]
- Roenneberg, T.; Foster, R.G. Twilight times: Light and the circadian system. Photochem. Photobiol. 1997, 66, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.G.; Helfrich-Forster, C. The regulation of circadian clocks by light in fruitflies and mice. Philos. Trans. R. Soc. Lond B Biol. Sci. 2001, 356, 1779–1789. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.E.; Takahashi, J.S. Sensitivity and integration in a visual pathway for circadian entrainment in the hamster (Mesocricetus auratus). J. Physiol. 1991, 439, 115–145. [Google Scholar] [CrossRef]
- Nelson, R.J.; Zucker, I. Absence of extraocular photoreception in diurnal and nocturnal rodents exposed to direct sunlight. Comp. Biochem. Physiol. Part A Physiol. 1981, 69, 145–148. [Google Scholar] [CrossRef]
- Foster, R.G.; Menaker, M. Circadian photoreception in mammals and other vertebrates. In Light and Biological Rhythms in Man; Wetterberg, L., Ed.; Pergamon: Oxford, UK, 1993; pp. 73–91. [Google Scholar]
- Gerkema, M.P.; Davies, W.I.; Foster, R.G.; Menaker, M.; Hut, R.A. The nocturnal bottleneck and the evolution of activity patterns in mammals. Proc. Biol. Sci. 2013, 280, 20130508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Deguchi, T. Entrainment of the circadian rhythms of blinded infant rats by nursing mothers. Physiol. Behav. 1983, 31, 373–378. [Google Scholar] [CrossRef]
- Foster, R.G.; Provencio, I.; Hudson, D.; Fiske, S.; De Grip, W.; Menaker, M. Circadian photoreception in the retinally degenerate mouse (rd/rd). J. Comp. Physiol. A 1991, 169, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Cassone, V.M. Circadian organization and photoreception in an Australian dasyurid marsupial (Sminthopsis macroura). J. Biol. Rhythm. 1987, 2, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.G. Keeping an eye on the time: The Cogan Lecture. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1286–1298. [Google Scholar]
- Moore, R.Y.; Lenn, N.J. A retinohypothalamic projection in the rat. J. Comp. Neurol. 1972, 146, 1–14. [Google Scholar] [CrossRef]
- Stephan, F.K.; Zucker, I. Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc. Natl. Acad. Sci. USA 1972, 69, 1583–1586. [Google Scholar] [CrossRef] [Green Version]
- Pittler, S.J.; Baehr, W. Identification of a nonsense mutation in the rod photoreceptor cGMP phosphodiesterase beta-subunit gene of the rd mouse. Proc. Natl. Acad. Sci. USA 1991, 88, 8322–8326. [Google Scholar] [CrossRef] [Green Version]
- Provencio, I.; Wong, S.; Lederman, A.B.; Argamaso, S.M.; Foster, R.G. Visual and circadian responses to light in aged retinally degenerate mice. Vis. Res. 1994, 34, 1799–1806. [Google Scholar] [CrossRef]
- Ebihara, S.; Tsuji, K. Entrainment of the circadian activity rhythm to the light cycle: Effective light intensity for a Zeitgeber in the retinal degenerate C3H mouse and the normal C57BL mouse. Physiol. Behav. 1980, 24, 523–527. [Google Scholar] [CrossRef]
- Yoshimura, T.; Ebihara, S. Decline of circadian photosensitivity associated with retinal degeneration in CBA/J-rd/rd mice. Brain Res. 1998, 779, 188–193. [Google Scholar] [CrossRef]
- Yoshimura, T.; Nishio, M.; Goto, M.; Ebihara, S. Differences in circadian photosensitivity between retinally degenerate CBA/J mice (rd/rd) and normal CBA/N mice (+/+). J. Biol. Rhythm. 1994, 9, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, E.M.; Linder, C.C.; Sargent, E.E.; Davisson, M.T.; Mobraaten, L.E.; Sharp, J.J. Genetic variation among 129 substrains and its importance for targeted mutagenesis in mice. Nat. Genet. 1997, 16, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Peirson, S.N.; Brown, L.A.; Pothecary, C.A.; Benson, L.A.; Fisk, A.S. Light and the laboratory mouse. J. Neurosci. Methods 2018, 300, 26–36. [Google Scholar] [CrossRef]
- Argamaso-Hernan, S. Light-evoked behaviour in mice with inherited retinal degeneration: An analysis of circadian photoentrainment. In Department of Biology; University of Virginia: Charlottesville, VA, USA, 1996. [Google Scholar]
- David-Gray, Z.K.; Janssen, J.W.; DeGrip, W.J.; Nevo, E.; Foster, R.G. Light detection in a ‘blind’ mammal. Nat. Neurosci. 1998, 1, 655–656. [Google Scholar] [CrossRef]
- Foster, R.G.; Argamaso, S.; Coleman, S.; Colwell, C.S.; Lederman, A.; Provencio, I. Photoreceptors regulating circadian behavior: A mouse model. J. Biol. Rhythm. 1993, 8 (Suppl. P), S17–S23. [Google Scholar]
- Sun, H.; Macke, J.P.; Nathans, J. Mechanisms of spectral tuning in the mouse green cone pigment. Proc. Natl. Acad. Sci. USA 1997, 94, 8860–8865. [Google Scholar] [CrossRef] [Green Version]
- McCall, M.A.; Gregg, R.G.; Merriman, K.; Goto, Y.; Peachey, N.S.; Stanford, L.R. Morphological and physiological consequences of the selective elimination of rod photoreceptors in transgenic mice. Exp. Eye Res. 1996, 63, 35–50. [Google Scholar] [CrossRef]
- Freedman, M.S.; Lucas, R.J.; Soni, B.; von Schantz, M.; Munoz, M.; David-Gray, Z.; Foster, R. Regulation of mammalian circadian behavior by non-rod, non-cone, ocular photoreceptors. Science 1999, 284, 502–504. [Google Scholar] [CrossRef]
- Lucas, R.J.; Freedman, M.S.; Munoz, M.; Garcia-Fernandez, J.M.; Foster, R.G. Regulation of the mammalian pineal by non-rod, non-cone, ocular photoreceptors. Science 1999, 284, 505–507. [Google Scholar] [CrossRef]
- Trejo, L.J.; Cicerone, C.M. Retinal sensitivity measured by the pupillary light reflex in RCS and albino rats. Vis. Res. 1982, 22, 1163–1171. [Google Scholar] [CrossRef] [Green Version]
- Lucas, R.J.; Douglas, R.H.; Foster, R.G. Characterization of an ocular photopigment capable of driving pupillary constriction in mice. Nat. Neurosci. 2001, 4, 621–626. [Google Scholar] [CrossRef]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by retinal ganglion cells that set the circadian clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [Green Version]
- Sekaran, S.; Foster, R.G.; Lucas, R.J.; Hankins, M.W. Calcium imaging reveals a network of intrinsically light-sensitive inner-retinal neurons. Curr. Biol. 2003, 13, 1290–1298. [Google Scholar] [CrossRef]
- Quattrochi, L.E.; Stabio, M.E.; Kim, I.; Ilardi, M.C.; Michelle Fogerson, P.; Leyrer, M.L.; Berson, D.M. The M6 cell: A small-field bistratified photosensitive retinal ganglion cell. J. Comp. Neurol. 2019, 527, 297–311. [Google Scholar] [CrossRef] [Green Version]
- Peirson, S.N.; Thompson, S.; Hankins, M.W.; Foster, R.G. Mammalian photoentrainment: Results, methods, and approaches. Methods Enzymol. 2005, 393, 697–726. [Google Scholar]
- Hattar, S.; Lucas, R.J.; Mrosovsky, N.; Thompson, S.; Douglas, R.H.; Hankins, M.W.; Lem, J.; Biel, M.; Hofmann, F.; Foster, R.G.; et al. Melanopsin and rod-cone photoreceptive systems account for all major accessory visual functions in mice. Nature 2003, 424, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Sonoda, T.; Schmidt, T.M. M1 Intrinsically Photosensitive Retinal Ganglion Cells Integrate Rod and Melanopsin Inputs to Signal in Low Light. Cell Rep. 2019, 29, 3349–3355 e2. [Google Scholar] [CrossRef] [Green Version]
- Milner, E.S.; Do, M.T.H. A Population Representation of Absolute Light Intensity in the Mammalian Retina. Cell 2017, 171, 865–876 e16. [Google Scholar] [CrossRef] [Green Version]
- Vitaterna, M.H.; Selby, C.P.; Todo, T.; Niwa, H.; Thompson, C.; Fruechte, E.M.; Hitomi, K.; Thresher, R.J.; Ishikawa, T.; Miyazaki, J.; et al. Differential regulation of mammalian period genes and circadian rhythmicity by cryptochromes 1 and 2. Proc. Natl. Acad. Sci. USA 1999, 96, 12114–12119. [Google Scholar] [CrossRef] [Green Version]
- Sancar, A. Cryptochrome: The second photoactive pigment in the eye and its role in circadian photoreception. Annu. Rev. Biochem. 2000, 69, 31–67. [Google Scholar] [CrossRef] [Green Version]
- Van Gelder, R.N.; Sancar, A. Cryptochromes and inner retinal non-visual irradiance detection. Novartis Found. Symp. 2003, 253, 31–42. [Google Scholar] [PubMed]
- Lucas, R.J.; Foster, R.G. Photoentrainment in mammals: A role for cryptochrome? J. Biol. Rhythm. 1999, 14, 4–10. [Google Scholar] [CrossRef]
- Ahmad, M.; Grancher, N.; Heil, M.; Black, R.C.; Giovani, B.; Galland, P.; Lardemer, D. Action spectrum for cryptochrome-dependent hypocotyl growth inhibition in Arabidopsis. Plant Physiol. 2002, 129, 774–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dacey, D.M.; Liao, H.W.; Peterson, B.B.; Robinson, F.R.; Smith, V.C.; Pokorny, J.; Yau, K.W.; Gamlin, P.D. Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project to the LGN. Nature 2005, 433, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Provencio, I.; Jiang, G.; De Grip, W.J.; Hayes, W.P.; Rollag, M.D. Melanopsin: An opsin in melanophores, brain, and eye. Proc. Natl. Acad. Sci. USA 1998, 95, 340–345. [Google Scholar] [CrossRef] [Green Version]
- Bellingham, J.; Whitmore, D.; Philp, A.R.; Wells, D.J.; Foster, R.G. Zebrafish melanopsin: Isolation, tissue localisation and phylogenetic position. Brain Res. Mol. Brain Res. 2002, 107, 128–136. [Google Scholar] [CrossRef]
- Provencio, I.; Rodriguez, I.R.; Jiang, G.; Hayes, W.P.; Moreira, E.F.; Rollag, M.D. A novel human opsin in the inner retina. J. Neurosci. 2000, 20, 600–605. [Google Scholar] [CrossRef] [Green Version]
- Semo, M.; Munoz Llamosas, M.; Foster, R.G.; Jeffery, G. Melanopsin (Opn4) positive cells in the cat retina are randomly distributed across the ganglion cell layer. Vis. Neurosci. 2005, 22, 111–116. [Google Scholar] [CrossRef]
- Pires, S.S.; Shand, J.; Bellingham, J.; Arrese, C.; Turton, M.; Peirson, S.; Foster, R.G.; Halford, S. Isolation and characterization of melanopsin (Opn4) from the Australian marsupial Sminthopsis crassicaudata (fat-tailed dunnart). Proc. Biol. Sci. 2007, 274, 2791–2799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provencio, I.; Cooper, H.M.; Foster, R.G. Retinal projections in mice with inherited retinal degeneration: Implications for circadian photoentrainment. J. Comp. Neurol. 1998, 395, 417–439. [Google Scholar] [CrossRef]
- Lucas, R.J.; Hattar, S.; Takao, M.; Berson, D.M.; Foster, R.G.; Yau, K.W. Diminished pupillary light reflex at high irradiances in melanopsin-knockout mice. Science 2003, 299, 245–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruby, N.F.; Brennan, T.J.; Xie, X.; Cao, V.; Franken, P.; Heller, H.C.; O’Hara, B.F. Role of melanopsin in circadian responses to light. Science 2002, 298, 2211–2213. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Sato, T.K.; Castrucci, A.M.; Rollag, M.D.; DeGrip, W.J.; Hogenesch, J.B.; Provencio, I.; Kay, S.A. Melanopsin (Opn4) requirement for normal light-induced circadian phase shifting. Science 2002, 298, 2213–2216. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.; Provencio, I.; Tu, D.C.; Pires, S.S.; Rollag, M.D.; Castrucci, A.M.; Pletcher, M.T.; Sato, T.K.; Wiltshire, T.; Andahazy, M.; et al. Melanopsin is required for non-image-forming photic responses in blind mice. Science 2003, 301, 525–527. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, Y.; Sancar, A. Vitamin B2-based blue-light photoreceptors in the retinohypothalamic tract as the photoactive pigments for setting the circadian clock in mammals. Proc. Natl. Acad. Sci. USA 1998, 95, 6097–6102. [Google Scholar] [CrossRef] [Green Version]
- Newman, L.A.; Walker, M.T.; Brown, R.L.; Cronin, T.W.; Robinson, P.R. Melanopsin forms a functional short-wavelength photopigment. Biochemistry 2003, 42, 12734–12738. [Google Scholar] [CrossRef]
- Melyan, Z.; Tarttelin, E.E.; Bellingham, J.; Lucas, R.J.; Hankins, M. Addition of human melanopsin renders mammalian cells photoresponsive. Nature 2005, 433, 741–745. [Google Scholar] [CrossRef]
- Qiu, X.; Kumbalasiri, T.; Carlson, S.M.; Wong, K.Y.; Krishna, V.; Provencio, I.; Berson, D.M. Induction of photosensitivity by heterologous expression of melanopsin. Nature 2005, 433, 745–749. [Google Scholar] [CrossRef]
- Panda, S.; Nayak, S.K.; Campo, B.; Walker, J.R.; Hogenesch, J.B.; Jegla, T. Illumination of the melanopsin signaling pathway. Science 2005, 307, 600–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mure, L.S.; Rieux, C.; Hattar, S.; Cooper, H.M. Melanopsin-dependent nonvisual responses: Evidence for photopigment bistability in vivo. J. Biol. Rhythm. 2007, 22, 411–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emanuel, A.J.; Do, M.T. Melanopsin tristability for sustained and broadband phototransduction. Neuron 2015, 85, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyanagi, M.; Kubokawa, K.; Tsukamoto, H.; Shichida, Y.; Terakita, A. Cephalochordate melanopsin: Evolutionary linkage between invertebrate visual cells and vertebrate photosensitive retinal ganglion cells. Curr. Biol. 2005, 15, 1065–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrosovsky, N.; Hattar, S. Impaired masking responses to light in melanopsin-knockout mice. Chronobiol. Int. 2003, 20, 989–999. [Google Scholar] [CrossRef]
- Lupi, D.; Oster, H.; Thompson, S.; Foster, R.G. The acute light-induction of sleep is mediated by OPN4-based photoreception. Nat. Neurosci. 2008, 11, 1068–1073. [Google Scholar]
- Hubbard, J.; Ruppert, E.; Gropp, C.M.; Bourgin, P. Non-circadian direct effects of light on sleep and alertness: Lessons from transgenic mouse models. Sleep Med. Rev. 2013, 17, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Altimus, C.M.; Guler, A.D.; Villa, K.L.; McNeill, D.S.; Legates, T.A.; Hattar, S. Rods-cones and melanopsin detect light and dark to modulate sleep independent of image formation. Proc. Natl. Acad. Sci. USA 2008, 105, 19998–20003. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.W.; Hannibal, J.; Hagiwara, G.; Colas, D.; Ruppert, E.; Ruby, N.F.; Heller, H.C.; Franken, P.; Bourgin, P. Melanopsin as a sleep modulator: Circadian gating of the direct effects of light on sleep and altered sleep homeostasis in Opn4(-/-) mice. PLoS Biol. 2009, 7, e1000125. [Google Scholar] [CrossRef]
- Pilorz, V.; Tam, S.K.; Hughes, S.; Pothecary, C.A.; Jagannath, A.; Hankins, M.W.; Bannerman, D.M.; Lightman, S.L.; Vyazovskiy, V.V.; Nolan, P.M.; et al. Melanopsin Regulates Both Sleep-Promoting and Arousal-Promoting Responses to Light. PLoS Biol. 2016, 14, e1002482. [Google Scholar] [CrossRef] [Green Version]
- Milosavljevic, N.; Cehajic-Kapetanovic, J.; Procyk, C.A.; Lucas, R.J. Chemogenetic Activation of Melanopsin Retinal Ganglion Cells Induces Signatures of Arousal and/or Anxiety in Mice. Curr. Biol. 2016, 26, 2358–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, D.C.; Fogerson, P.M.; Lazzerini Ospri, L.; Thomsen, M.B.; Layne, R.M.; Severin, D.; Zhan, J.; Singer, J.H.; Kirkwood, A.; Zhao, H.; et al. Light Affects Mood and Learning through Distinct Retina-Brain Pathways. Cell 2018, 175, 71–84.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semo, M.; Gias, C.; Ahmado, A.; Sugano, E.; Allen, A.E.; Lawrence, J.M.; Tomita, H.; Coffey, P.J.; Vugler, A.A. Dissecting a role for melanopsin in behavioural light aversion reveals a response independent of conventional photoreception. PLoS ONE 2010, 5, e15009. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Wu, V.; Donovan, M.; Majumdar, S.; Renteria, R.C.; Porco, T.; Van Gelder, R.N.; Copenhagen, D.R. Melanopsin-dependent light avoidance in neonatal mice. Proc. Natl. Acad. Sci. USA 2010, 107, 17374–17378. [Google Scholar] [CrossRef] [Green Version]
- LeGates, T.A.; Altimus, C.M.; Wang, H.; Lee, H.K.; Yang, S.; Zhao, H.; Kirkwood, A.; Weber, E.T.; Hattar, S. Aberrant light directly impairs mood and learning through melanopsin-expressing neurons. Nature 2012, 491, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Van Diepen, H.C.; Foster, R.G.; Meijer, J.H. A colourful clock. PLoS Biol. 2015, 13, e1002160. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.E.; Storchi, R.; Martial, F.P.; Bedford, R.A.; Lucas, R.J. Melanopsin Contributions to the Representation of Images in the Early Visual System. Curr. Biol. 2017, 27, 1623–1632 e4. [Google Scholar] [CrossRef] [Green Version]
- Sondereker, K.B.; Stabio, M.E.; Renna, J.M. Crosstalk: The diversity of melanopsin ganglion cell types has begun to challenge the canonical divide between image-forming and non-image-forming vision. J. Comp. Neurol. 2020. [Google Scholar] [CrossRef]
- Zaidi, F.H.; Hull, J.T.; Peirson, S.N.; Wulff, K.; Aeschbach, D.; Gooley, J.J.; Brainard, G.C.; Gregory-Evans, K.; Rizzo, J.F., 3rd; Czeisler, C.A.; et al. Short-wavelength light sensitivity of circadian, pupillary, and visual awareness in humans lacking an outer retina. Curr. Biol. 2007, 17, 2122–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, T.M.; Tsujimura, S.; Allen, A.E.; Wynne, J.; Bedford, R.; Vickery, G.; Vugler, A.; Lucas, R.J. Melanopsin-based brightness discrimination in mice and humans. Curr. Biol. 2012, 22, 1134–1141. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.M.; Gias, C.; Hatori, M.; Keding, S.R.; Semo, M.; Coffey, P.J.; Gigg, J.; Piggins, H.D.; Panda, S.; Lucas, R.J. Melanopsin contributions to irradiance coding in the thalamo-cortical visual system. PLoS Biol. 2010, 8, e1000558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, K.E.; Eleftheriou, C.G.; Allen, A.E.; Procyk, C.A.; Lucas, R.J. Melanopsin-derived visual responses under light adapted conditions in the mouse dLGN. PLoS ONE 2015, 10, e0123424. [Google Scholar] [CrossRef] [PubMed]
- Le, T.M.; Alam, N.M.; Chen, S.; Kofuji, P.; Li, W.; Prusky, G.T.; Hattar, S. A role for melanopsin in alpha retinal ganglion cells and contrast detection. Neuron 2014, 82, 781–788. [Google Scholar]
- Allen, A.E.; Storchi, R.; Martial, F.P.; Petersen, R.S.; Montemurro, M.A.; Brown, T.M.; Lucas, R.J. Melanopsin-driven light adaptation in mouse vision. Curr. Biol. 2014, 24, 2481–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ecker, J.L.; Dumitrescu, O.N.; Wong, K.Y.; Alam, N.M.; Chen, S.K.; LeGates, T.; Renna, J.M.; Prusky, G.T.; Berson, D.M.; Hattar, S. Melanopsin-expressing retinal ganglion-cell photoreceptors: Cellular diversity and role in pattern vision. Neuron 2010, 67, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.E.; Martial, F.P.; Lucas, R.J. Form vision from melanopsin in humans. Nat. Commun. 2019, 10, 2274. [Google Scholar] [CrossRef] [Green Version]
- Hankins, M.W.; Hughes, S. Vision: Melanopsin as a novel irradiance detector at the heart of vision. Curr. Biol. 2014, 24, R1055–R1057. [Google Scholar] [CrossRef] [Green Version]
- Barnard, A.R.; Hattar, S.; Hankins, M.W.; Lucas, R.J. Melanopsin regulates visual processing in the mouse retina. Curr. Biol. 2006, 16, 389–395. [Google Scholar] [CrossRef]
- Milosavljevic, N.; Allen, A.E.; Cehajic-Kapetanovic, J.; Lucas, R.J. Chemogenetic Activation of ipRGCs Drives Changes in Dark-Adapted (Scotopic) Electroretinogram. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6305–6312. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.L.; Feng, J.; Zhang, D.Q. Functional integrity and modification of retinal dopaminergic neurons in the rd1 mutant mouse: Roles of melanopsin and GABA. J. Neurophysiol. 2013, 109, 1589–1599. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Q.; Wong, K.Y.; Sollars, P.J.; Berson, D.M.; Pickard, G.E.; McMahon, D.G. Intraretinal signaling by ganglion cell photoreceptors to dopaminergic amacrine neurons. Proc. Natl. Acad. Sci. USA 2008, 105, 14181–14186. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Q.; Belenky, M.A.; Sollars, P.J.; Pickard, G.E.; McMahon, D.G. Melanopsin mediates retrograde visual signaling in the retina. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reifler, A.N.; Chervenak, A.P.; Dolikian, M.E.; Benenati, B.A.; Li, B.Y.; Wachter, R.D.; Lynch, A.M.; Demertzis, Z.D.; Meyers, B.S.; Abufarha, F.S.; et al. All Spiking, Sustained ON Displaced Amacrine Cells Receive Gap-Junction Input from Melanopsin Ganglion Cells. Curr. Biol. 2015, 25, 2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milosavljevic, N.; Storchi, R.; Eleftheriou, C.G.; Colins, A.; Petersen, R.S.; Lucas, R.J. Photoreceptive retinal ganglion cells control the information rate of the optic nerve. Proc. Natl. Acad. Sci. USA 2018, 115, E11817–E11826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renna, J.M.; Weng, S.; Berson, D.M. Light acts through melanopsin to alter retinal waves and segregation of retinogeniculate afferents. Nat. Neurosci. 2011, 14, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Kirkby, L.A.; Feller, M.B. Intrinsically photosensitive ganglion cells contribute to plasticity in retinal wave circuits. Proc. Natl. Acad. Sci. USA 2013, 110, 12090–12095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, K.S.; Renna, J.M.; McNeill, D.S.; Fernandez, D.C.; Keenan, W.T.; Thomsen, M.B.; Ecker, J.L.; Loevinsohn, G.S.; VanDunk, C.; Vicarel, D.C.; et al. A subset of ipRGCs regulates both maturation of the circadian clock and segregation of retinogeniculate projections in mice. Elife 2017, 6, e22861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattar, S.; Kumar, M.; Park, A.; Tong, P.; Tung, J.; Yau, K.W.; Berson, D.M. Central projections of melanopsin-expressing retinal ganglion cells in the mouse. J. Comp. Neurol. 2006, 497, 326–349. [Google Scholar] [CrossRef] [Green Version]
- Baver, S.B.; Pickard, G.E.; Sollars, P.J.; Pickard, G.E. Two types of melanopsin retinal ganglion cell differentially innervate the hypothalamic suprachiasmatic nucleus and the olivary pretectal nucleus. Eur. J. Neurosci. 2008, 27, 1763–1770. [Google Scholar] [CrossRef]
- Schmidt, T.M.; Kofuji, P. Functional and morphological differences among intrinsically photosensitive retinal ganglion cells. J. Neurosci. 2009, 29, 476–482. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.M.; Chen, S.K.; Hattar, S. Intrinsically photosensitive retinal ganglion cells: Many subtypes, diverse functions. Trends Neurosci. 2011, 34, 572–580. [Google Scholar] [CrossRef] [Green Version]
- Hughes, S.; Hankins, M.W.; Foster, R.G.; Peirson, S.N. Melanopsin phototransduction: Slowly emerging from the dark. Prog. Brain Res. 2012, 199, 19–40. [Google Scholar]
- McNeill, D.S.; Sheely, C.J.; Ecker, J.L.; Badea, T.C.; Morhardt, D.; Guido, W.; Hattar, S. Development of melanopsin-based irradiance detecting circuitry. Neural Dev. 2011, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.K.; Badea, T.C.; Hattar, S. Photoentrainment and pupillary light reflex are mediated by distinct populations of ipRGCs. Nature 2011, 476, 92–95. [Google Scholar] [CrossRef] [Green Version]
- Estevez, M.E.; Fogerson, P.M.; Ilardi, M.C.; Borghuis, B.G.; Chan, E.; Weng, S.; Auferkorte, O.N.; Demb, J.B.; Berson, D.M. Form and function of the M4 cell, an intrinsically photosensitive retinal ganglion cell type contributing to geniculocortical vision. J. Neurosci. 2012, 32, 13608–13620. [Google Scholar] [CrossRef] [Green Version]
- Provencio, I.; Rollag, M.D.; Castrucci, A.M. Photoreceptive net in the mammalian retina. This mesh of cells may explain how some blind mice can still tell day from night. Nature 2002, 415, 493. [Google Scholar] [CrossRef]
- Berson, D.M.; Castrucci, A.M.; Provencio, I. Morphology and mosaics of melanopsin-expressing retinal ganglion cell types in mice. J. Comp. Neurol. 2010, 518, 2405–2422. [Google Scholar] [CrossRef] [Green Version]
- Hughes, S.; Watson, T.S.; Foster, R.G.; Peirson, S.N.; Hankins, M.W. Nonuniform distribution and spectral tuning of photosensitive retinal ganglion cells of the mouse retina. Curr. Biol. 2013, 23, 1696–1701. [Google Scholar] [CrossRef] [Green Version]
- Hatori, M.; Le, H.; Vollmers, C.; Keding, S.R.; Tanaka, N.; Buch, T.; Waisman, A.; Schmedt, C.; Jegla, T.; Panda, S. Inducible ablation of melanopsin-expressing retinal ganglion cells reveals their central role in non-image forming visual responses. PLoS ONE 2008, 3, e2451. [Google Scholar] [CrossRef]
- Sonoda, T.; Okabe, Y.; Schmidt, T.M. Overlapping morphological and functional properties between M4 and M5 intrinsically photosensitive retinal ganglion cells. J. Comp. Neurol. 2020, 528, 1028–1040. [Google Scholar] [CrossRef]
- Berg, D.J.; Kartheiser, K.; Leyrer, M.; Saali, A.; Berson, D.M. Transcriptomic Signatures of Postnatal and Adult Intrinsically Photosensitive Ganglion Cells. Eneuro 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Stafford, B.K.; Godin, A.L.; King, W.M.; Wong, K.Y. Photoresponse diversity among the five types of intrinsically photosensitive retinal ganglion cells. J. Physiol. 2014, 592, 1619–1636. [Google Scholar] [CrossRef] [Green Version]
- Tu, D.C.; Zhang, D.; Demas, J.; Slutsky, E.B.; Provencio, I.; Holy, T.E.; Van Gelder, R.N. Physiologic diversity and development of intrinsically photosensitive retinal ganglion cells. Neuron 2005, 48, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mure, L.S.; Vinberg, F.; Hanneken, A.; Panda, S. Functional diversity of human intrinsically photosensitive retinal ganglion cells. Science 2019, 366, 1251–1255. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.; Estevez, M.E.; Berson, D.M. Mouse ganglion-cell photoreceptors are driven by the most sensitive rod pathway and by both types of cones. PLoS ONE 2013, 8, e66480. [Google Scholar] [CrossRef]
- Wong, K.Y.; Dunn, F.A.; Graham, D.M.; Berson, D.M. Synaptic influences on rat ganglion-cell photoreceptors. J. Physiol. 2007, 582, 279–296. [Google Scholar] [CrossRef]
- Belenky, M.A.; Smeraski, C.A.; Provencio, I.; Sollars, P.J.; Pickard, G.E. Melanopsin retinal ganglion cells receive bipolar and amacrine cell synapses. J. Comp. Neurol. 2003, 460, 380–393. [Google Scholar] [CrossRef]
- Pickard, G.E.; Baver, S.B.; Ogilvie, M.D.; Sollars, P.J. Light-induced fos expression in intrinsically photosensitive retinal ganglion cells in melanopsin knockout (opn4) mice. PLoS ONE 2009, 4, e4984. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.M.; Kofuji, P. Differential cone pathway influence on intrinsically photosensitive retinal ganglion cell subtypes. J. Neurosci. 2010, 30, 16262–16271. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.Y. A retinal ganglion cell that can signal irradiance continuously for 10 hours. J. Neurosci. 2012, 32, 11478–11485. [Google Scholar] [CrossRef] [Green Version]
- Perez-Leon, J.A.; Warren, E.J.; Allen, C.N.; Robinson, D.W.; Lane Brown, R. Synaptic inputs to retinal ganglion cells that set the circadian clock. Eur. J. Neurosci. 2006, 24, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.M.; Taniguchi, K.; Kofuji, P. Intrinsic and extrinsic light responses in melanopsin-expressing ganglion cells during mouse development. J. Neurophysiol. 2008, 100, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Reifler, A.N.; Schroeder, M.M.; Jaeckel, E.R.; Chervenak, A.P.; Wong, K.Y. Mechanisms creating transient and sustained photoresponses in mammalian retinal ganglion cells. J. Gen. Physiol. 2017, 149, 335–353. [Google Scholar] [CrossRef] [Green Version]
- Spitschan, M.; Lazar, R.; Yetik, E.; Cajochen, C. No evidence for an S cone contribution to acute neuroendocrine and alerting responses to light. Curr. Biol. 2019, 29, R1297–R1298. [Google Scholar] [CrossRef] [Green Version]
- David-Gray, Z.K.; Cooper, H.M.; Janssen, J.W.; Nevo, E.; Foster, R.G. Spectral tuning of a circadian photopigment in a subterranean ‘blind’ mammal (Spalax ehrenbergi). FEBS Lett. 1999, 461, 343–347. [Google Scholar] [CrossRef] [Green Version]
- David-Gray, Z.K.; Bellingham, J.; Munoz, M.; Avivi, A.; Nevo, E.; Foster, R.G. Adaptive loss of ultraviolet-sensitive/violet-sensitive (UVS/VS) cone opsin in the blind mole rat (Spalax ehrenbergi). Eur. J. Neurosci. 2002, 16, 1186–1194. [Google Scholar] [CrossRef]
- Provencio, I.; Foster, R.G. Circadian rhythms in mice can be regulated by photoreceptors with cone-like characteristics. Brain Res. 1995, 694, 183–190. [Google Scholar] [CrossRef]
- Reifler, A.N.; Chervenak, A.P.; Dolikian, M.E.; Benenati, B.A.; Meyers, B.S.; Demertzis, Z.D.; Lynch, A.M.; Li, B.Y.; Wachter, R.D.; Abufarha, F.S.; et al. The rat retina has five types of ganglion-cell photoreceptors. Exp. Eye Res. 2015, 130, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Sexton, T.J.; Bleckert, A.; Turner, M.H.; Van Gelder, R.N. Type I intrinsically photosensitive retinal ganglion cells of early post-natal development correspond to the M4 subtype. Neural Dev. 2015, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Do, M.T.; Yau, K.W. Adaptation to steady light by intrinsically photosensitive retinal ganglion cells. Proc. Natl. Acad. Sci. USA 2013, 110, 7470–7475. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.Y.; Dunn, F.A.; Berson, D.M. Photoreceptor adaptation in intrinsically photosensitive retinal ganglion cells. Neuron 2005, 48, 1001–1010. [Google Scholar] [CrossRef] [Green Version]
- Emanuel, A.J.; Kapur, K.; Do, M.T.H. Biophysical Variation within the M1 Type of Ganglion Cell Photoreceptor. Cell Rep. 2017, 21, 1048–1062. [Google Scholar] [CrossRef] [Green Version]
- Sodhi, P.; Hartwick, A.T. Adenosine modulates light responses of rat retinal ganglion cell photoreceptors througha cAMP-mediated pathway. J. Physiol. 2014, 592, 4201–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hook, M.J.; Wong, K.Y.; Berson, D.M. Dopaminergic modulation of ganglion-cell photoreceptors in rat. Eur. J. Neurosci. 2012, 35, 507–518. [Google Scholar] [CrossRef]
- Warren, E.J.; Allen, C.N.; Brown, R.L.; Robinson, D.W. The light-activated signaling pathway in SCN-projecting rat retinal ganglion cells. Eur. J. Neurosci. 2006, 23, 2477–2487. [Google Scholar] [CrossRef]
- Hartwick, A.T.; Bramley, J.R.; Yu, J.; Stevens, K.T.; Allen, C.N.; Baldridge, W.H.; Sollars, P.J.; Pickard, G.E. Light-evoked calcium responses of isolated melanopsin-expressing retinal ganglion cells. J. Neurosci. 2007, 27, 13468–13480. [Google Scholar] [CrossRef]
- Sekaran, S.; Lall, G.S.; Ralphs, K.L.; Wolstenholme, A.J.; Lucas, R.J.; Foster, R.G.; Hankins, M.W. 2-Aminoethoxydiphenylborane is an acute inhibitor of directly photosensitive retinal ganglion cell activity in vitro and in vivo. J. Neurosci. 2007, 27, 3981–3986. [Google Scholar] [CrossRef]
- Graham, D.M.; Wong, K.Y.; Shapiro, P.; Frederick, C.; Pattabiraman, K.; Berson, D.M. Melanopsin ganglion cells use a membrane-associated rhabdomeric phototransduction cascade. J. Neurophysiol. 2008, 99, 2522–2532. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Do, M.T.; Riccio, A.; Jiang, Z.; Hsieh, J.; Wang, H.C.; Merbs, S.L.; Welsbie, D.S.; Yoshioka, T.; Weissgerber, P.; et al. Melanopsin signalling in mammalian iris and retina. Nature 2011, 479, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.; Jagannath, A.; Hickey, D.; Gatti, S.; Wood, M.; Peirson, S.N.; Foster, R.G.; Hankins, M.W. Using siRNA to define functional interactions between melanopsin and multiple G Protein partners. Cell. Mol. Life Sci. 2015, 72, 165–179. [Google Scholar] [CrossRef] [Green Version]
- Peirson, S.N.; Oster, H.; Jones, S.L.; Leitges, M.; Hankins, M.W.; Foster, R.G. Microarray analysis and functional genomics identify novel components of melanopsin signaling. Curr. Biol. 2007, 17, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoda, T.; Lee, S.K.; Birnbaumer, L.; Schmidt, T.M. Melanopsin Phototransduction Is Repurposed by ipRGC Subtypes to Shape the Function of Distinct Visual Circuits. Neuron 2018, 99, 754–767.e4. [Google Scholar] [CrossRef] [PubMed]
- Pires, S.S.; Hughes, S.; Turton, M.; Melyan, Z.; Peirson, S.N.; Zheng, L.; Kosmaoglou, M.; Bellingham, J.; Cheetham, M.E.; Lucas, R.J.; et al. Differential expression of two distinct functional isoforms of melanopsin (Opn4) in the mammalian retina. J. Neurosci. 2009, 29, 12332–12342. [Google Scholar] [CrossRef] [Green Version]
- Krupnick, J.G.; Benovic, J.L. The role of receptor kinases and arrestins in G protein-coupled receptor regulation. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 289–319. [Google Scholar] [CrossRef]
- Ritter, S.L.; Hall, R.A. Fine-tuning of GPCR activity by receptor-interacting proteins. Nat. Rev. Mol. Cell Biol. 2009, 10, 819–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishnivetskiy, S.A.; Raman, D.; Wei, J.; Kennedy, M.J.; Hurley, J.B.; Gurevich, V.V. Regulation of arrestin binding by rhodopsin phosphorylation level. J. Biol. Chem. 2007, 282, 32075–32083. [Google Scholar] [CrossRef] [Green Version]
- Somasundaram, P.; Wyrick, G.R.; Fernandez, D.C.; Ghahari, A.; Pinhal, C.M.; Simmonds Richardson, M.; Rupp, A.C.; Cui, L.; Wu, Z.; Brown, R.L.; et al. C-terminal phosphorylation regulates the kinetics of a subset of melanopsin-mediated behaviors in mice. Proc. Natl. Acad. Sci. USA 2017, 114, 2741–2746. [Google Scholar] [CrossRef] [Green Version]
- Blasic, J.R., Jr.; Matos-Cruz, V.; Ujla, D.; Cameron, E.G.; Hattar, S.; Halpern, M.E.; Robinson, P.R. Identification of critical phosphorylation sites on the carboxy tail of melanopsin. Biochemistry 2014, 53, 2644–2649. [Google Scholar] [CrossRef]
- Valdez-Lopez, J.C.; Gulati, S.; Ortiz, E.A.; Palczewski, K.; Robinson, P.R. Melanopsin Carboxy-terminus phosphorylation plasticity and bulk negative charge, not strict site specificity, achieves phototransduction deactivation. PLoS ONE 2020, 15, e0228121. [Google Scholar] [CrossRef] [Green Version]
- Hughes, S.; Welsh, L.; Katti, C.; Gonzalez-Menendez, I.; Turton, M.; Halford, S.; Sekaran, S.; Peirson, S.N.; Hankins, M.W.; Foster, R.G. Differential expression of melanopsin isoforms Opn4L and Opn4S during postnatal development of the mouse retina. PLoS ONE 2012, 7, e34531. [Google Scholar] [CrossRef] [Green Version]
- Jagannath, A.; Hughes, S.; Abdelgany, A.; Pothecary, C.A.; Di Pretoro, S.; Pires, S.S.; Vachtsevanos, A.; Pilorz, V.; Brown, L.A.; Hossbach, M.; et al. Isoforms of Melanopsin Mediate Different Behavioral Responses to Light. Curr. Biol. 2015, 25, 2430–2434. [Google Scholar] [CrossRef] [Green Version]
- van Oosterhout, F.; Fisher, S.P.; van Diepen, H.C.; Watson, T.S.; Houben, T.; VanderLeest, H.T.; Thompson, S.; Peirson, S.N.; Foster, R.G.; Meijer, J.H. Ultraviolet light provides a major input to non-image-forming light detection in mice. Curr. Biol. 2012, 22, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Meijer, J.H.; Watanabe, K.; Schaap, J.; Albus, H.; Detari, L. Light responsiveness of the suprachiasmatic nucleus: Long-term multiunit and single-unit recordings in freely moving rats. J. Neurosci. 1998, 18, 9078–9087. [Google Scholar] [CrossRef] [PubMed]
- Aggelopoulos, N.C.; Meissl, H. Responses of neurones of the rat suprachiasmatic nucleus to retinal illumination under photopic and scotopic conditions. J. Physiol. 2000, 523, 211–222. [Google Scholar] [CrossRef]
- Nakamura, T.J.; Fujimura, K.; Ebihara, S.; Shinohara, K. Light response of the neuronal firing activity in the suprachiasmatic nucleus of mice. Neurosci. Lett. 2004, 371, 244–248. [Google Scholar] [CrossRef]
- Drouyer, E.; Rieux, C.; Hut, R.A.; Cooper, H.M. Responses of suprachiasmatic nucleus neurons to light and dark adaptation: Relative contributions of melanopsin and rod-cone inputs. J. Neurosci. 2007, 27, 9623–9631. [Google Scholar] [CrossRef] [Green Version]
- Groos, G.; Mason, R. Maintained discharge of rat suprachiasmatic neurons at different adaptation levels. Neurosci. Lett. 1978, 8, 59–64. [Google Scholar] [CrossRef]
- Meijer, J.H.; Groos, G.A.; Rusak, B. Luminance coding in a circadian pacemaker: The suprachiasmatic nucleus of the rat and the hamster. Brain Res. 1986, 382, 109–118. [Google Scholar] [CrossRef]
- Brown, T.M.; Wynne, J.; Piggins, H.D.; Lucas, R.J. Multiple hypothalamic cell populations encoding distinct visual information. J. Physiol. 2011, 589, 1173–1194. [Google Scholar] [CrossRef] [PubMed]
- van Diepen, H.C.; Ramkisoensing, A.; Peirson, S.N.; Foster, R.G.; Meijer, J.H. Irradiance encoding in the suprachiasmatic nuclei by rod and cone photoreceptors. FASEB J. 2013, 27, 4204–4212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, L.; Morin, L.P. Absence of normal photic integration in the circadian visual system: Response to millisecond light flashes. J. Neurosci. 2007, 27, 3375–3382. [Google Scholar] [CrossRef] [Green Version]
- Lall, G.S.; Revell, V.L.; Momiji, H.; Al Enezi, J.; Altimus, C.M.; Guler, A.D.; Aguilar, C.; Cameron, M.A.; Allender, S.; Hankins, M.W.; et al. Distinct contributions of rod, cone, and melanopsin photoreceptors to encoding irradiance. Neuron 2010, 66, 417–428. [Google Scholar] [CrossRef]
- Dowling, G.A.; Hubbard, E.M.; Mastick, J.; Luxenberg, J.S.; Burr, R.L.; Van Someren, E.J. Effect of morning bright light treatment for rest-activity disruption in institutionalized patients with severe Alzheimer’s disease. Int. Psychogeriatr. 2005, 17, 221–236. [Google Scholar] [CrossRef]
- Archer, S.N. Adaptive Mechanisms in the Ecology of Vision; Springer: Dordrecht, The Netherlands, 1999; 668p. [Google Scholar]
- Spitschan, M.; Aguirre, G.K.; Brainard, D.H.; Sweeney, A.M. Variation of outdoor illumination as a function of solar elevation and light pollution. Sci. Rep. 2016, 6, 26756. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, L.; Hanna, L.; Mouland, J.; Martial, F.; West, A.; Smedley, A.R.; Bechtold, D.A.; Webb, A.R.; Lucas, R.J.; Brown, T.M. Colour as a signal for entraining the mammalian circadian clock. PLoS Biol. 2015, 13, e1002127. [Google Scholar] [CrossRef] [PubMed]
- Sadun, A.A.; Schaechter, J.D.; Smith, L.E. A retinohypothalamic pathway in man: Light mediation of circadian rhythms. Brain Res. 1984, 302, 371–377. [Google Scholar] [CrossRef]
- Wever, R.A. The Circadian System of Man: Results of Experiments under Temporal Isolation; Springer: New York, NY, USA, 1979. [Google Scholar]
- Aschoff, J.; Fatranska, M.; Giedke, H.; Doerr, P.; Stamm, D.; Wisser, H. Human circadian rhythms in continuous darkness: Entrainment by social cues. Science 1971, 171, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Honma, K.; Honma, S.; Wada, T. Entrainment of human circadian rhythms by artificial bright light cycles. Experientia 1987, 43, 572–574. [Google Scholar] [CrossRef]
- Sack, R.L.; Lewy, A.J.; Blood, M.L.; Keith, L.D.; Nakagawa, H. Circadian rhythm abnormalities in totally blind people: Incidence and clinical significance. J. Clin. Endocrinol. Metab. 1992, 75, 127–134. [Google Scholar]
- Klein, T.; Martens, H.; Dijk, D.J.; Kronauer, R.E.; Seely, E.W.; Czeisler, C.A. Circadian sleep regulation in the absence of light perception: Chronic non-24-hour circadian rhythm sleep disorder in a blind man with a regular 24-hour sleep-wake schedule. Sleep 1993, 16, 333–343. [Google Scholar] [CrossRef]
- Campbell, S.S.; Murphy, P.J. Extraocular circadian phototransduction in humans. Science 1998, 279, 396–399. [Google Scholar] [CrossRef] [Green Version]
- Foster, R.G. Shedding light on the biological clock. Neuron 1998, 20, 829–832. [Google Scholar] [CrossRef] [Green Version]
- Lindblom, N.; Hatonen, T.; Laakso, M.; Alila-Johansson, A.; Laipio, M.; Turpeinen, U. Bright light exposure of a large skin area does not affect melatonin or bilirubin levels in humans. Biol. Psychiatry 2000, 48, 1098–1104. [Google Scholar] [CrossRef]
- Lindblom, N.; Heiskala, H.; Hatonen, T.; Mustanoja, S.; Alfthan, H.; Alila-Johansson, A.; Laakso, M.L. No evidence for extraocular light induced phase shifting of human melatonin, cortisol and thyrotropin rhythms. Neuroreport 2000, 11, 713–717. [Google Scholar] [CrossRef]
- Yamazaki, S.; Goto, M.; Menaker, M. No evidence for extraocular photoreceptors in the circadian system of the Syrian hamster. J. Biol. Rhythm. 1999, 14, 197–201. [Google Scholar] [CrossRef]
- Wright, K.P., Jr.; Czeisler, C.A. Absence of circadian phase resetting in response to bright light behind the knees. Science 2002, 297, 571. [Google Scholar] [CrossRef] [Green Version]
- Czeisler, C.A.; Shanahan, T.L.; Klerman, E.B.; Martens, H.; Brotman, D.J.; Emens, J.S.; Klein, T.; Rizzo, J.F., 3rd. Suppression of melatonin secretion in some blind patients by exposure to bright light. N. Engl. J. Med. 1995, 332, 6–11. [Google Scholar] [CrossRef]
- Skene, D.J.; Lockley, S.W.; Arendt, J. Melatonin in circadian sleep disorders in the blind. Biol. Signals Recept. 1999, 8, 90–95. [Google Scholar] [CrossRef]
- Matsuyama, T.; Yamashita, T.; Imamoto, Y.; Shichida, Y. Photochemical properties of mammalian melanopsin. Biochemistry 2012, 51, 5454–5462. [Google Scholar] [CrossRef]
- Walker, M.T.; Brown, R.L.; Cronin, T.W.; Robinson, P.R. Photochemistry of retinal chromophore in mouse melanopsin. Proc. Natl. Acad. Sci. USA 2008, 105, 8861–8865. [Google Scholar] [CrossRef] [Green Version]
- Bailes, H.J.; Lucas, R.J. Human melanopsin forms a pigment maximally sensitive to blue light (lambdamax approximately 479 nm) supporting activation of G(q/11) and G(i/o) signalling cascades. Proc. Biol. Sci. 2013, 280, 20122987. [Google Scholar] [CrossRef] [Green Version]
- Hannibal, J.; Hindersson, P.; Ostergaard, J.; Georg, B.; Heegaard, S.; Larsen, P.J.; Fahrenkrug, J. Melanopsin is expressed in PACAP-containing retinal ganglion cells of the human retinohypothalamic tract. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4202–4209. [Google Scholar] [CrossRef] [Green Version]
- Hannibal, J.; Kankipati, L.; Strang, C.E.; Peterson, B.B.; Dacey, D.; Gamlin, P.D. Central projections of intrinsically photosensitive retinal ganglion cells in the macaque monkey. J. Comp. Neurol. 2014, 522, 2231–2248. [Google Scholar] [CrossRef] [Green Version]
- Jusuf, P.R.; Lee, S.C.; Hannibal, J.; Grunert, U. Characterization and synaptic connectivity of melanopsin-containing ganglion cells in the primate retina. Eur. J. Neurosci. 2007, 26, 2906–2921. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, J.; Christiansen, A.T.; Heegaard, S.; Fahrenkrug, J.; Kiilgaard, J.F. Melanopsin expressing human retinal ganglion cells: Subtypes, distribution, and intraretinal connectivity. J. Comp. Neurol. 2017, 525, 1934–1961. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Kumar, C.J.; Merrow, M. The human circadian clock entrains to sun time. Curr. Biol. 2007, 17, R44–R45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kantermann, T.; Juda, M.; Merrow, M.; Roenneberg, T. The human circadian clock’s seasonal adjustment is disrupted by daylight saving time. Curr. Biol. 2007, 17, 1996–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jean-Louis, G.; Kripke, D.F.; Ancoli-Israel, S.; Klauber, M.R.; Sepulveda, R.S. Sleep duration, illumination, and activity patterns in a population sample: Effects of gender and ethnicity. Biol. Psychiatry 2000, 47, 921–927. [Google Scholar] [CrossRef]
- Thorne, H.C.; Jones, K.H.; Peters, S.P.; Archer, S.N.; Dijk, D.J. Daily and seasonal variation in the spectral composition of light exposure in humans. Chronobiol. Int. 2009, 26, 854–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, K.P., Jr.; McHill, A.W.; Birks, B.R.; Griffin, B.R.; Rusterholz, T.; Chinoy, E.D. Entrainment of the human circadian clock to the natural light-dark cycle. Curr. Biol. 2013, 23, 1554–1558. [Google Scholar] [CrossRef] [Green Version]
- Stothard, E.R.; McHill, A.W.; Depner, C.M.; Birks, B.R.; Moehlman, T.M.; Ritchie, H.K.; Guzzetti, J.R.; Chinoy, E.D.; LeBourgeois, M.K.; Axelsson, J.; et al. Circadian Entrainment to the Natural Light-Dark Cycle across Seasons and the Weekend. Curr. Biol. 2017, 27, 508–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcheret, K.; Wald, L.; Fritschi, L.; Gerkema, M.; Gordijn, M.; Merrrow, M.; Rajaratnam, S.M.W.; Rock, D.; Sletten, T.L.; Warman, G.; et al. Chronotype and environmental light exposure in a student population. Chronobiol. Int. 2018, 35, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Honma, K.; Honma, S.; Wada, T. Phase-dependent shift of free-running human circadian rhythms in response to a single bright light pulse. Experientia 1987, 43, 1205–1207. [Google Scholar] [CrossRef] [PubMed]
- Honma, K.; Honma, S.; Nakamura, K.; Sasaki, M.; Endo, T.; Takahashi, T. Differential effects of bright light and social cues on reentrainment of human circadian rhythms. Am. J. Physiol. 1995, 268, R528–R535. [Google Scholar] [CrossRef]
- Zeitzer, J.M.; Dijk, D.J.; Kronauer, R.; Brown, E.; Czeisler, C. Sensitivity of the human circadian pacemaker to nocturnal light: Melatonin phase resetting and suppression. J. Physiol. 2000, 526, 695–702. [Google Scholar] [CrossRef]
- Chang, A.M.; Santhi, N.; St Hilaire, M.; Gronfier, C.; Bradstreet, D.S.; Duffy, J.F.; Lockley, S.W.; Kronauer, R.E.; Czeisler, C.A. Human responses to bright light of different durations. J. Physiol. 2012, 590, 3103–3112. [Google Scholar] [CrossRef] [Green Version]
- Jagannath, A.; Butler, R.; Godinho, S.I.; Couch, Y.; Brown, L.A.; Vasudevan, S.R.; Flanagan, K.C.; Anthony, D.; Churchill, G.C.; Wood, M.J.; et al. The CRTC1-SIK1 pathway regulates entrainment of the circadian clock. Cell 2013, 154, 1100–1111. [Google Scholar] [CrossRef] [Green Version]
- Ho Mien, I.; Chua, E.C.; Lau, P.; Tan, L.C.; Lee, I.T.; Yeo, S.C.; Tan, S.S.; Gooley, J.J. Effects of exposure to intermittent versus continuous red light on human circadian rhythms, melatonin suppression, and pupillary constriction. PLoS ONE 2014, 9, e96532. [Google Scholar] [CrossRef] [Green Version]
- Usui, T.; Kremers, J.; Sharpe, L.T.; Zrenner, E. Response phase of the flicker electroretinogram (ERG) is influenced by cone excitation strength. Vis. Res. 1998, 38, 3247–3251. [Google Scholar] [CrossRef] [Green Version]
- Rimmer, D.W.; Boivin, D.B.; Shanahan, T.L.; Kronauer, R.E.; Duffy, J.F.; Czeisler, C.A. Dynamic resetting of the human circadian pacemaker by intermittent bright light. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1574–R1579. [Google Scholar] [CrossRef]
- Gronfier, C.; Wright, K.P., Jr.; Kronauer, R.E.; Jewett, M.E.; Czeisler, C.A. Efficacy of a single sequence of intermittent bright light pulses for delaying circadian phase in humans. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E174–E181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeitzer, J.M.; Ruby, N.F.; Fisicaro, R.A.; Heller, H.C. Response of the human circadian system to millisecond flashes of light. PLoS ONE 2011, 6, e22078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugh, E.N., Jr. The discovery of the ability of rod photoreceptors to signal single photons. J. Gen. Physiol. 2018, 150, 383–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeitzer, J.M.; Kronauer, R.E.; Czeisler, C.A. Photopic transduction implicated in human circadian entrainment. Neurosci. Lett. 1997, 232, 135–138. [Google Scholar] [CrossRef]
- Brainard, G.C.; Hanifin, J.P.; Greeson, J.M.; Byrne, B.; Glickman, G.; Gerner, E.; Rollag, M.D. Action spectrum for melatonin regulation in humans: Evidence for a novel circadian photoreceptor. J. Neurosci. 2001, 21, 6405–6412. [Google Scholar] [CrossRef] [Green Version]
- Thapan, K.; Arendt, J.; Skene, D.J. An action spectrum for melatonin suppression: Evidence for a novel non-rod, non-cone photoreceptor system in humans. J. Physiol. 2001, 535, 261–267. [Google Scholar] [CrossRef]
- Hankins, M.W.; Lucas, R.J. The primary visual pathway in humans is regulated according to long-term light exposure through the action of a nonclassical photopigment. Curr. Biol. 2002, 12, 191–198. [Google Scholar] [CrossRef]
- Spitschan, M. Melanopsin contributions to non-visual and visual function. Curr. Opin. Behav. Sci. 2019, 30, 67–72. [Google Scholar] [CrossRef]
- Warman, V.L.; Dijk, D.J.; Warman, G.R.; Arendt, J.; Skene, D.J. Phase advancing human circadian rhythms with short wavelength light. Neurosci. Lett. 2003, 342, 37–40. [Google Scholar] [CrossRef]
- Wright, H.R.; Lack, L.C. Effect of light wavelength on suppression and phase delay of the melatonin rhythm. Chronobiol. Int. 2001, 18, 801–808. [Google Scholar] [CrossRef]
- Lockley, S.W.; Brainard, G.C.; Czeisler, C.A. High sensitivity of the human circadian melatonin rhythm to resetting by short wavelength light. J. Clin. Endocrinol. Metab. 2003, 88, 4502–4505. [Google Scholar] [CrossRef]
- Revell, V.L.; Arendt, J.; Terman, M.; Skene, D.J. Short-wavelength sensitivity of the human circadian system to phase-advancing light. J. Biol. Rhythm. 2005, 20, 270–272. [Google Scholar] [CrossRef]
- Souman, J.L.; Tinga, A.M.; Te Pas, S.F.; van Ee, R.; Vlaskamp, B.N.S. Acute alerting effects of light: A systematic literature review. Behav. Brain Res. 2018, 337, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Brainard, G.C.; Sliney, D.; Hanifin, J.P.; Glickman, G.; Byrne, B.; Greeson, J.M.; Jasser, S.; Gerner, E.; Rollag, M.D. Sensitivity of the human circadian system to short-wavelength (420-nm) light. J. Biol. Rhythm. 2008, 23, 379–386. [Google Scholar] [CrossRef] [Green Version]
- McDougal, D.H.; Gamlin, P.D. The influence of intrinsically-photosensitive retinal ganglion cells on the spectral sensitivity and response dynamics of the human pupillary light reflex. Vis. Res. 2010, 50, 72–87. [Google Scholar] [CrossRef] [Green Version]
- Figueiro, M.G.; Wood, B.; Plitnick, B.; Rea, M.S. The impact of light from computer monitors on melatonin levels in college students. Neuroendocrinol. Lett. 2011, 32, 158–163. [Google Scholar]
- Cajochen, C.; Frey, S.; Anders, D.; Spati, J.; Bues, M.; Pross, A.; Mager, R.; Wirz-Justice, A.; Stefani, O. Evening exposure to a light-emitting diodes (LED)-backlit computer screen affects circadian physiology and cognitive performance. J. Appl. Physiol. 2011, 110, 1432–1438. [Google Scholar] [CrossRef] [Green Version]
- Wood, B.; Rea, M.S.; Plitnick, B.; Figueiro, M.G. Light level and duration of exposure determine the impact of self-luminous tablets on melatonin suppression. Appl. Ergon. 2013, 44, 237–240. [Google Scholar] [CrossRef]
- Chang, A.M.; Aeschbach, D.; Duffy, J.F.; Czeisler, C.A. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc. Natl. Acad. Sci. USA 2015, 112, 1232–1237. [Google Scholar] [CrossRef] [Green Version]
- Kazemi, R.; Alighanbari, N.; Zamanian, Z. The effects of screen light filtering software on cognitive performance and sleep among night workers. Health Promot. Perspect. 2019, 9, 233–240. [Google Scholar] [CrossRef]
- Meijer, J.H.; Rusak, B.; Ganshirt, G. The relation between light-induced discharge in the suprachiasmatic nucleus and phase shifts of hamster circadian rhythms. Brain Res. 1992, 598, 257–263. [Google Scholar] [CrossRef]
- Owen, J.; Arendt, J. Melatonin suppression in human subjects by bright and dim light in antarctica: Time and season-dependent effects. Neurosci. Lett. 1992, 137, 181–184. [Google Scholar] [CrossRef]
- Nelson, D.E.; Takahashi, J.S. Integration and saturation within the circadian photic entrainment pathway of hamsters. Am. J. Physiol. 1999, 277, R1351–R1361. [Google Scholar] [CrossRef] [PubMed]
- Refinetti, R. Dark adaptation in the circadian system of the mouse. Physiol. Behav. 2001, 74, 101–107. [Google Scholar] [CrossRef]
- Hebert, M.; Martin, S.K.; Lee, C.; Eastman, C.I. The effects of prior light history on the suppression of melatonin by light in humans. J. Pineal Res. 2002, 33, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.A.; Schoen, M.W.; Czeisler, C.A. Adaptation of human pineal melatonin suppression by recent photic history. J. Clin. Endocrinol. Metab. 2004, 89, 3610–3614. [Google Scholar] [CrossRef]
- Chang, A.-M.; Scheer, F.A.; Czeisler, C.A. Adaptation of the human circadian system by prior light history. Sleep 2008, 31, A45–A46. [Google Scholar]
- Cole, R.J.; Kripke, D.F.; Wisbey, J.; Mason, W.J.; Gruen, W.; Hauri, P.J.; Juarez, S. Seasonal variation in human illumination exposure at two different latitudes. J. Biol. Rhythm. 1995, 10, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Hebert, M.; Dumont, M.; Paquet, J. Seasonal and diurnal patterns of human illumination under natural conditions. Chronobiol. Int. 1998, 15, 59–70. [Google Scholar] [CrossRef]
- Scheuermaier, K.; Laffan, A.M.; Duffy, J.F. Light exposure patterns in healthy older and young adults. J. Biol. Rhythm. 2010, 25, 113–122. [Google Scholar] [CrossRef]
- Kawinska, A.; Dumont, M.; Selmaoui, B.; Paquet, J.; Carrier, J. Are modifications of melatonin circadian rhythm in the middle years of life related to habitual patterns of light exposure? J. Biol. Rhythm. 2005, 20, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Scheuermaier, K.; Laffan, A.M.; Duffy, J.F. Light exposure patterns in healthy older people living in New England, USA. J. Sleep Res. 2006, 15, 94. [Google Scholar]
- Kalmbach, D.A.; Schneider, L.D.; Cheung, J.; Bertrand, S.J.; Kariharan, T.; Pack, A.I.; Gehrman, P.R. Genetic Basis of Chronotype in Humans: Insights from Three Landmark GWAS. Sleep 2017, 40. [Google Scholar] [CrossRef] [PubMed]
- Fabbian, F.; Zucchi, B.; De Giorgi, A.; Tiseo, R.; Boari, B.; Salmi, R.; Cappadona, R.; Gianesini, G.; Bassi, E.; Signani, F.; et al. Chronotype, gender and general health. Chronobiol. Int. 2016, 33, 863–882. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Lombardi, D.A.; Marucci-Wellman, H.; Roenneberg, T. Chronotypes in the US—Influence of age and sex. PLoS ONE 2017, 12, e0178782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, G.; Nolan, P.M.; Peirson, S.N. Reciprocal interactions between circadian clocks and aging. Mamm. Genome 2016, 27, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Wolfson, A.R.; Carskadon, M.A. Sleep schedules and daytime functioning in adolescents. Child Dev. 1998, 69, 875–887. [Google Scholar] [CrossRef]
- Laberge, L.; Petit, D.; Simard, C.; Vitaro, F.; Tremblay, R.E.; Montplaisir, J. Development of sleep patterns in early adolescence. J. Sleep Res. 2001, 10, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Giannotti, F.; Cortesi, F.; Sebastiani, T.; Ottaviano, S. Circadian preference, sleep and daytime behaviour in adolescence. J. Sleep Res. 2002, 11, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Wolfson, A.R.; Carskadon, M.A.; Acebo, C.; Seifer, R.; Fallone, G.; Labyak, S.E.; Martin, J.L. Evidence for the validity of a sleep habits survey for adolescents. Sleep 2003, 26, 213–216. [Google Scholar] [CrossRef]
- Roenneberg, T.; Kuehnle, T.; Pramstaller, P.P.; Ricken, J.; Havel, M.; Guth, A.; Merrow, M. A marker for the end of adolescence. Curr. Biol. 2004, 14, R1038–R1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, E.M.; Mindell, J.A. Sleep and risk-taking behavior in adolescents. Behav. Sleep Med. 2005, 3, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.J.; Acebo, C.; Carskadon, M.A. Sleep, circadian rhythms, and delayed phase in adolescence. Sleep Med. 2007, 8, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Van den Bulck, J. Television viewing, computer game playing, and Internet use and self-reported time to bed and time out of bed in secondary-school children. Sleep 2004, 27, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Gradisar, M.; Wolfson, A.R.; Harvey, A.G.; Hale, L.; Rosenberg, R.; Czeisler, C.A. The sleep and technology use of Americans: Findings from the National Sleep Foundation’s 2011 Sleep in America poll. J. Clin. Sleep Med. 2013, 9, 1291–1299. [Google Scholar] [CrossRef]
- Carskadon, M.A.; Acebo, C.; Jenni, O.G. Regulation of adolescent sleep: Implications for behavior. Ann. N. Y. Acad. Sci. 2004, 1021, 276–291. [Google Scholar] [CrossRef]
- Carskadon, M.A.; Acebo, C.; Richardson, G.S.; Tate, B.A.; Seifer, R. An approach to studying circadian rhythms of adolescent humans. J. Biol. Rhythm. 1997, 12, 278–289. [Google Scholar] [CrossRef]
- Hagenauer, M.H.; Perryman, J.I.; Lee, T.M.; Carskadon, M.A. Adolescent changes in the homeostatic and circadian regulation of sleep. Dev. Neurosci. 2009, 31, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Crowley, S.J.; Cain, S.W.; Burns, A.C.; Acebo, C.; Carskadon, M.A. Increased Sensitivity of the Circadian System to Light in Early/Mid-Puberty. J. Clin. Endocrinol. Metab. 2015, 100, 4067–4073. [Google Scholar] [CrossRef]
- Duffy, J.F.; Zeitzer, J.M.; Czeisler, C.A. Decreased sensitivity to phase-delaying effects of moderate intensity light in older subjects. Neurobiol. Aging 2007, 28, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Cuthbertson, F.M.; Peirson, S.N.; Wulff, K.; Foster, R.G.; Downes, S.M. Blue light-filtering intraocular lenses: Review of potential benefits and side effects. J. Cataract Refract. Surg. 2009, 35, 1281–1297. [Google Scholar] [CrossRef] [PubMed]
- Alexander, I.; Cuthbertson, F.M.; Ratnarajan, G.; Safa, R.; Mellington, F.E.; Foster, R.G.; Downes, S.M.; Wulff, K. Impact of cataract surgery on sleep in patients receiving either ultraviolet-blocking or blue-filtering intraocular lens implants. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4999–5004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najjar, R.P.; Chiquet, C.; Teikari, P.; Cornut, P.L.; Claustrat, B.; Denis, P.; Cooper, H.M.; Gronfier, C. Aging of non-visual spectral sensitivity to light in humans: Compensatory mechanisms? PLoS ONE 2014, 9, e85837. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Benloucif, S.; Reid, K.J.; Weintraub, S.; Kennedy, N.; Wolfe, L.F.; Zee, P.C. Phase-shifting response to light in older adults. J. Physiol. 2014, 592, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Spitschan, M.; Woelders, T. The Method of Silent Substitution for Examining Melanopsin Contributions to Pupil Control. Front Neurol. 2018, 9, 941. [Google Scholar] [CrossRef] [PubMed]
- Aschoff, J.; Hoffmann, K.; Pohl, H.; Wever, R. Re-entrainment of circadian rhythms after phase-shifts of the Zeitgeber. Chronobiologia 1975, 2, 23–78. [Google Scholar]
- Czeisler, C.A.; Duffy, J.F.; Shanahan, T.L.; Brown, E.N.; Mitchell, J.F.; Rimmer, D.W.; Ronda, J.M.; Silva, E.J.; Allan, J.S.; Emens, J.S.; et al. Stability, precision, and near-24-hour period of the human circadian pacemaker. Science 1999, 284, 2177–2181. [Google Scholar] [CrossRef]
- Patterson, S.S.; Kuchenbecker, J.A.; Anderson, J.R.; Neitz, M.; Neitz, J. A Color Vision Circuit for Non-Image-Forming Vision in the Primate Retina. Curr. Biol. 2020, 30, 1269–1274.e2. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) The Percentage of Animals Entrained to 12L:12:D in C57 Wildtype and C3H rd/rd Mice. | |||||
| Strain | 100 lux | 10.0 lux | 1.00 lux | 0.10 lux | 0.01 lux |
| C57 wildtype | 100 (9) | 100 (9) | 87.5 (8) | 85.7 (7) | 83.3 (6) |
| C3H rd/rd | 100 (12) | 100 (18) | 31.5 (19) | 0 (17) | - |
| (B) The Percentage of Animals Entrained to 12L:12:D in C57 Wildtype, C3H rd/rd and C3H +/+ Mice. | |||||
| Strain | 100 lux | 10.0 lux | 1.00 lux | 0.10 lux | 0.01 lux |
| C3H wildtype | 100 (28) | 100 (8) | 50 (10) | 12.5 (8) | 0 (8) |
| C3H rd/rd | 100 (27) | 100 (7) | 100 (8) | 12.5 (8) | 0 (10) |
| (C) The Percentage of Animals Entrained to 16L:8D in C3H Wildtype and C3H rd/rd Mice. | |||||
| Strain | 100 lux | 10.0 lux | 1.00 lux | 0.10 lux | 0.01 lux |
| C57 wildtype | 100 (12) | 100 (14) | 100 (14) | 100 (10) | 75 (8) |
| C3H rd/rd | 100 (16) | 100 (11) | 93.8 (16) | 23.5 (17) | 5.6 (18) |
| C3H wildtype | 100 (4) | 100 (2) | 31.5 (2) | 0 (4) | 0 (4) |
| Variation in the Stimulus (Intensity, Duration, Wavelength) | |
| Channel/Signal Noise | Fluctuations in the light signal. e.g., Cloud cover; daylength/season. |
| Environmental Noise | Extraneous light signals. e.g., Starlight; moonlight; artificial lightning. |
| Receptor Noise | Molecular noise of the receptor pathway. e.g., Variation in external temperature; individual gene polymorphisms in the receptor pathway |
| Variation in exposure & response to the stimulus (Type of Activity, Light History, Age, Time of Day) | |
| Sensory Adaptation | Changing receptor thresholds. e.g., Receptor habituation; changes in pupil size; ocular pigment migration; circadian gated responses. |
| Behavioral Noise | Behavioral state. e.g., type and time spent in work vs home vs recreational environment. |
| Developmental Noise | Stage of development. e.g., Altered behavioral, physiological, biochemical responses with age; impact of disease. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foster, R.G.; Hughes, S.; Peirson, S.N. Circadian Photoentrainment in Mice and Humans. Biology 2020, 9, 180. https://doi.org/10.3390/biology9070180
Foster RG, Hughes S, Peirson SN. Circadian Photoentrainment in Mice and Humans. Biology. 2020; 9(7):180. https://doi.org/10.3390/biology9070180
Chicago/Turabian StyleFoster, Russell G., Steven Hughes, and Stuart N. Peirson. 2020. "Circadian Photoentrainment in Mice and Humans" Biology 9, no. 7: 180. https://doi.org/10.3390/biology9070180
APA StyleFoster, R. G., Hughes, S., & Peirson, S. N. (2020). Circadian Photoentrainment in Mice and Humans. Biology, 9(7), 180. https://doi.org/10.3390/biology9070180