Oxygen Deficient (OD) Combustion and Metabolism: Allometric Laws of Organs and Kleiber’s Law from OD Metabolism?


Abstract
:1. Introduction and Background
- Specifically, when body mass increases across mammals, why are all fk values negative {Equation (7)}? Why do various organs and tissues have different dk and fk values? [17].
- The allometric size relationship is somehow ‘programmed’ into cells although the factors that let them know whether they are in a small or large organism are still unknown [25].
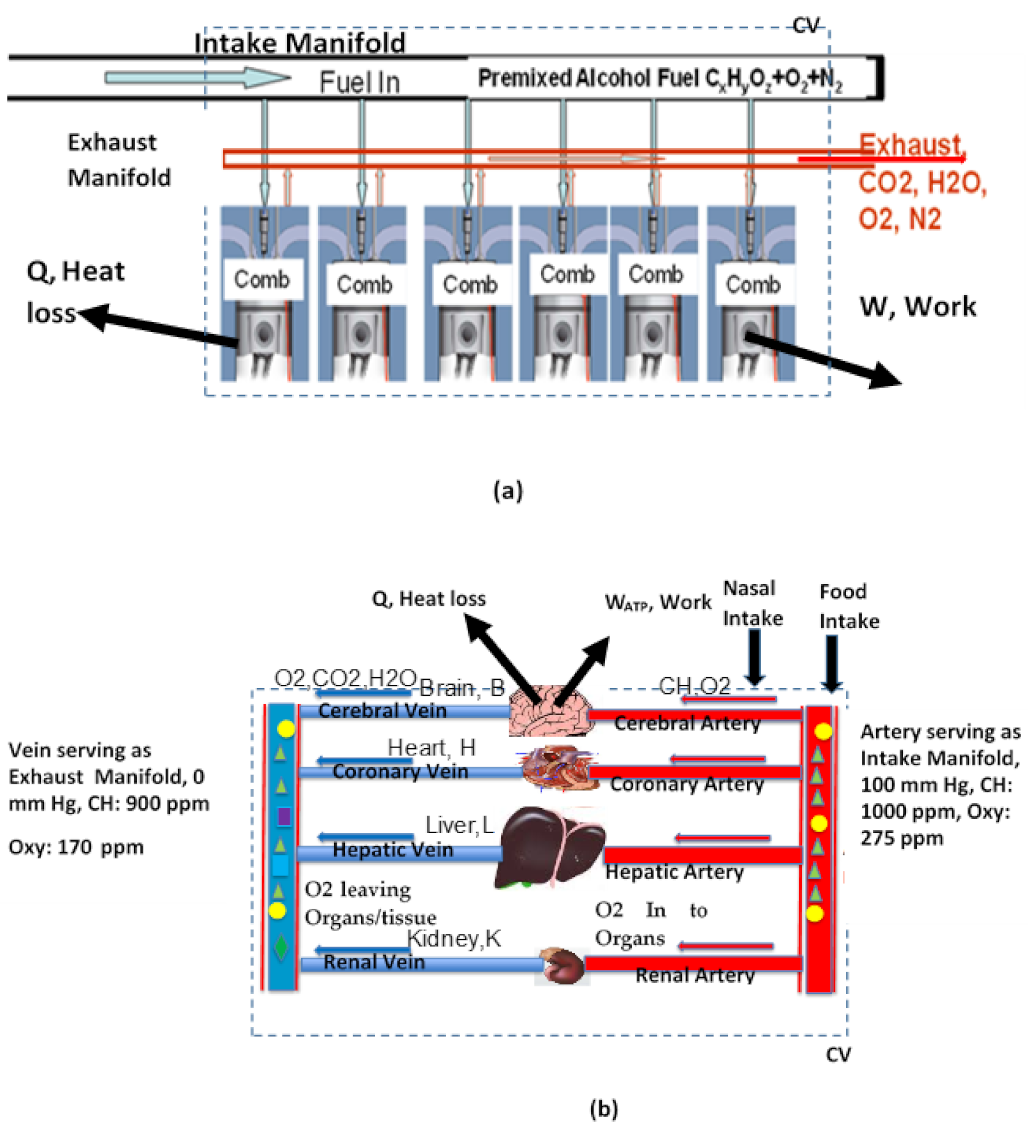
2. Literature Review of Group or Oxygen Deficient Combustion (ODC) in Engineering and Oxygen Deficient Metabolism (ODM) in Biology
2.1. Oxidation of Carbon
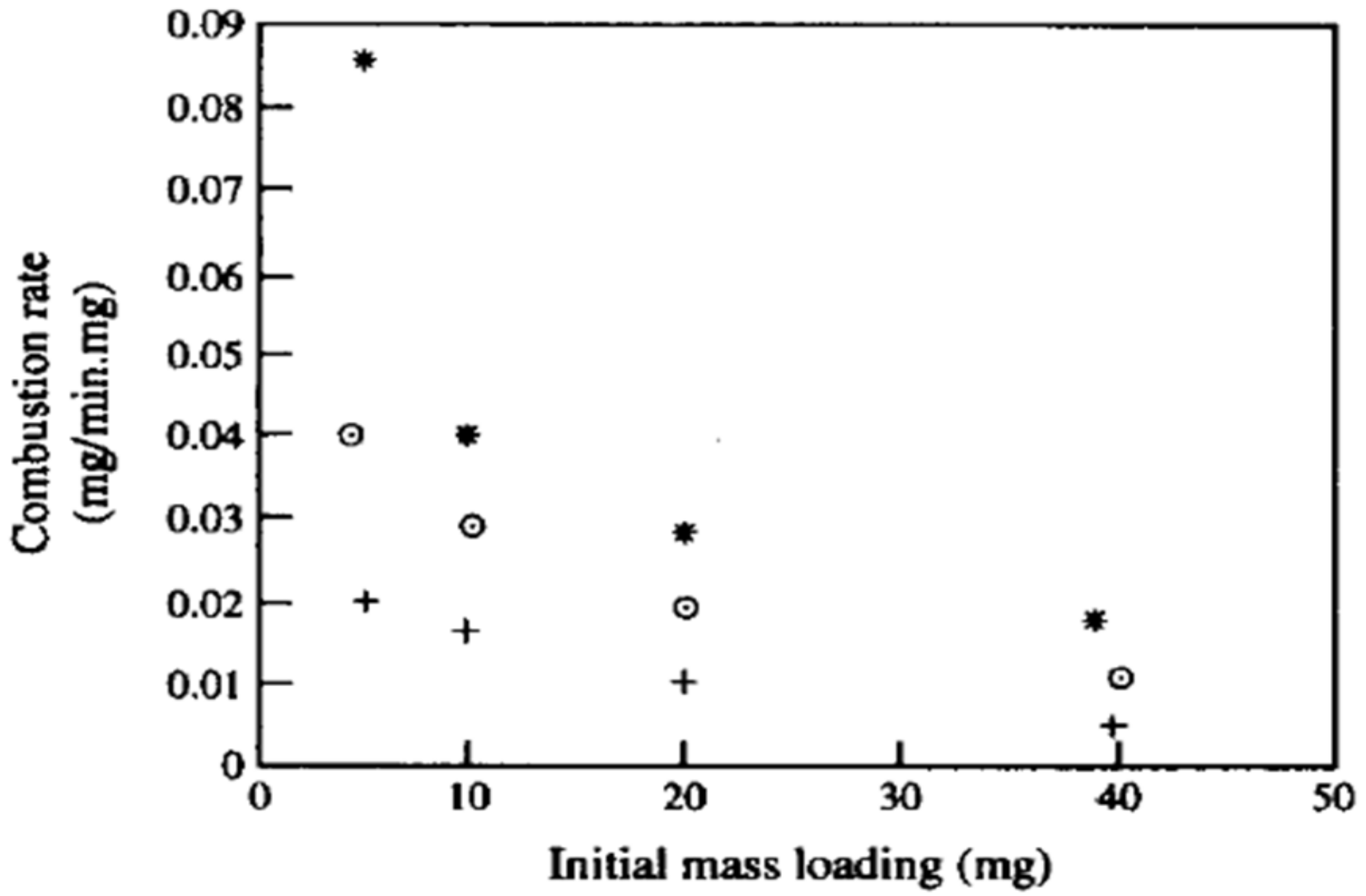
2.2. Group Combustion Or ODC
2.3. Oxidation of Nutrients in BS
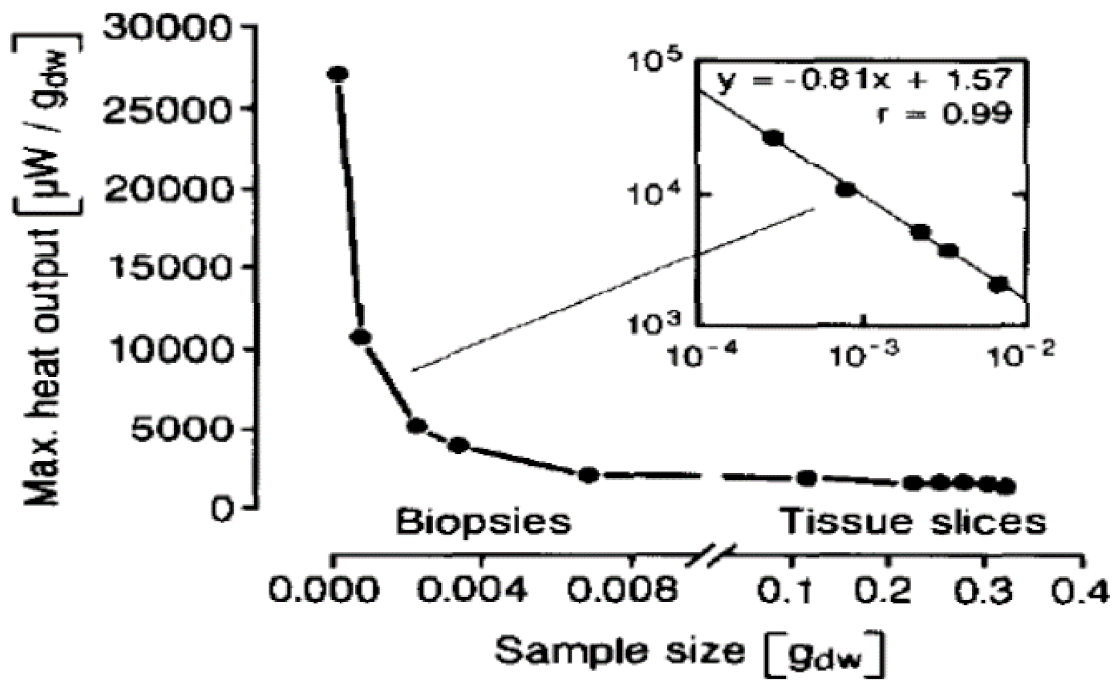
2.4. Oxidation Models for Organs
2.5. Oxygen Deficient Metabolism (ODM) in Organs
2.6. Organ Mass Based Allometry (OMA)
3. Objectives
4. Materials and Methods
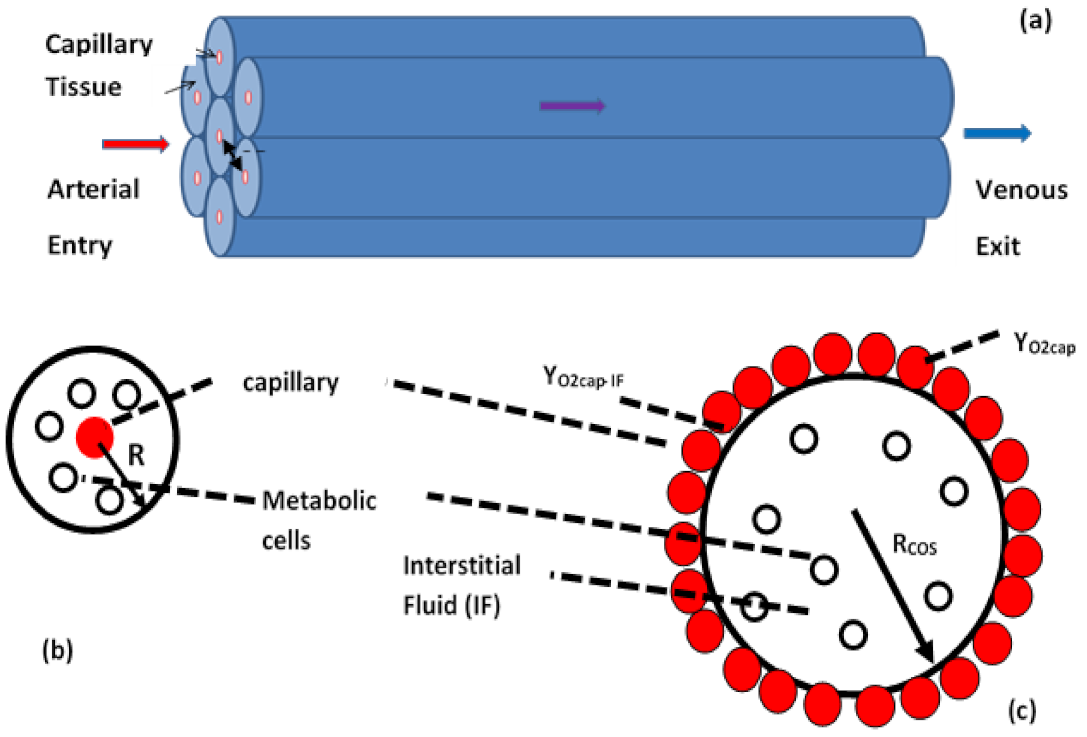
Governing Equations and Boundary Conditions for the ODM Model of Cell Clouds
5. Results
5.1. Equilibrium Levels of Oxygen in Arterial Blood
5.2. Oxygen Profiles in Cell Clouds
5.3. Effectiveness Factor (ηeff)
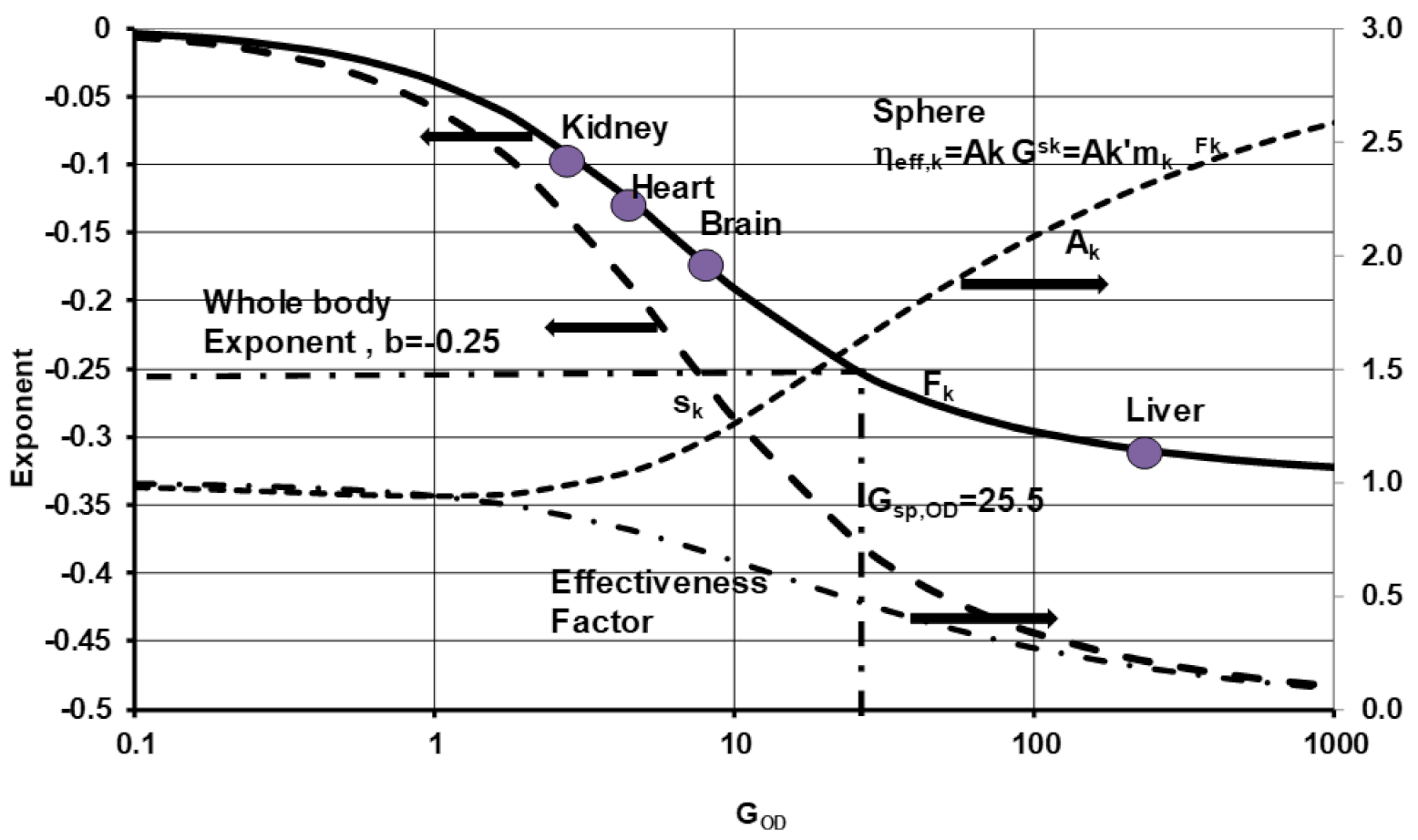
5.4. Allometric Laws Derived from Effectiveness Factor Charts
5.5. Validation with Experimental Data for Allometric Exponents
5.6. Whole Body Allometric Law for Metabolic Rate
5.7. GOD Number Estimation of Vital Organs
5.8. Incipient ODC or Group Combustion and Incipient ODM
6. Conclusions
- The dimensionless number (G) used in combustion science was modified to GOD for the application to OD metabolisms in organs.
- The rationale for negative exponents (Fk) in the allometric law for SMR of vital organs, , is provided through the adoption of group combustion or OD theory from combustion science within biology. The exponent Fk satisfies the inequality for COS-O2 model. Thus, the allometric law for an organ is shown to be an empirical approximation of effectiveness chart.
- The negative exponent b’ in the allometric law for the SMR of the whole body satisfies the inequality with b’ = −1/4 (Kleiber’s law). The negative exponent b’ = −1/4 is due to distributed sizes of various organs with differing values for Fk.
- Based on experimental data from Fk, the GOD numbers were estimated for vital organs as 2.8, 4.2, 9, and 240 for the kidney, heart, brain, and liver, respectively. The average GOD for the whole body of all BS with an equivalent organ is about 26 with effectiveness factor of 0.48 (i.e., oxidation of all tissues at 48% capillary oxygen concentration), considering spherical geometries for cell clouds.
- While the hypoxic condition of the whole body is characterized by the saturation percentage, the introduction of the GOD number characterizes OD at organ level.
- Glycolysis is inhibited when there is a high level of ATP within cells and vice versa; thus, reduced O2 concentration results in a reduced ERR per unit volume which results in reduced ATP concentration, which promotes glycolysis pathway [58,59]. Cancer and virus cells, including those of COVID-19 patients, rely on the glycolysis pathway to provide the building blocks for uncontrollable cell growth.
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations/Nomenclature
| Nomenclature | |
| Ak | see Equation (22), pre-exponential factor |
| a, b, b’ | allometric coefficients for metabolism |
| Cch, cell | Characteristic Oxygen Consumption Rate for cell, g/s, {Table 3} |
| Cch, p | Characteristic Oxygen Consumption Rate for carbon particle, g/s, {Table 3} |
| ck | pre-exponent in the allometry relation for mass of organ |
| D | Diffusivity, cm2/s |
| Deff | Effective Diffusion Coefficient, cm2/s |
| dcell | Diameter of cell, cm |
| dk | exponent in allometry relation for mass of organ |
| dp | Diameter of particle, cm |
| D | Diffusion Coefficient of Molecular Oxygen |
| Deff | Effective diffusion Coefficient |
| ek | pre-exponent in the allometry relation for metabolic rate |
| Fk | Allometric exponent, organ mass based {} |
| fk | Allometric exponent, body mass based allometry used in } |
| G | Group combustion number in combustion science {Table 3} |
| GOD | Group metabolism number in biology to indicate extent of Oxygen Deficiency (OD) |
| GOD, kin | Group metabolism number for organ k under kinetics control {= ΨT2 or Thiele Modulus2, |
| Gk, OD, diff | Group metabolism number for organ k under diffusion control, |
| gk, hk | see Equation (25) |
| (Hb) | Hemoglobin concentration on mass basis, g per DL of blood |
| [Hb] | Hemoglobin concentration on mole basis, moles per DL of blood |
| HO2 | Henry’s constant for solubility of O2 in blood |
| HV | Heating Value of fuel or nutrient, J/g fuel |
| HVO2 | Heating Value per unit mass of O2 consumed, J/g O2 |
| Jk | Pre-exponent in allometry relation for OEF |
| kMM | Constant in MM kinetics, Equation (29), = YO2 at half of maximum metabolic rate |
| Kn’ | Equilibrium Constant, (1/mm Hg when n = 1, 1/mm Hg2 n = 2, etc.), n = 1, 2, 3, 4 |
| LMM | Constant in LM kinetics, YO2 at half of maximum oxygen consumption rate |
| mB | Body mass, kg |
| mcl | cloud mass, kg |
| mk, | k th organ mass, kg, Brain, Heart, Kidney, liver etc. |
| mk* | Non-dimensional mass, = {mass of organ k/Mass of kidney} |
| n | number of particle or cells per unit volume, particles/cm3, or cells/cm3 |
| pO2 | Partial pressure of O2, mm of Hg |
| Energy released in the form of heat, W | |
| Energy release rate (ERR) or metabolic rate, W | |
| k, m | Specific Energy Release Rate or SMR (SERR), W/kg of k-th organ |
| m | Specific Energy Release Rate or SMR (SERR), W/kg |
| r | radius |
| rcap | Capillary radius, cm |
| R, Rcl | Cloud radius, cm |
| RQ | Respiratory Quotient, =CO2 moles/O2 moles, Table 1 |
| S, Scap | Capillary surface area, cm2 |
| Scap, m | Specific capillary surface area based on mass, cm2/g of tissue, {=S/ (ρ V)} |
| sk | See equation (22), exponent |
| V | Volume, = cm3 |
| Energy Released in the form of Work or ATP, W | |
| Oxygen Consumption rate, g/s; per unit area, g/(cm2 s), per unit volume, g/(cm3 s) | |
| YO2 | Mass fraction of oxygen, (g of O2/g of mixture) |
| δ | Aerobic shell thickness |
| Greek Symbols | |
| ξ | Non-dimensional radius, r/R |
| ρbl | Density of blood, g/cm3 |
| ψT | Thiele Modulus, |
| ηeff | Effectiveness factor based on capillary-IF interface O2 concentration, = YO2, avg/YO2, cap-IF |
| ηM | metabolic efficiency |
| νO2 | stoichiometric Oxygen |
| Subscripts | |
| A | Alveolar |
| avg | Average |
| a | Arterial |
| B | Body |
| Bl | Blood |
| cap | capillary |
| Cap-IF | interface between capillary and Interstitial Fluid (IF) |
| cl | Cloud |
| eff | effective |
| ext | extinction |
| IF | Interstitial fluid |
| k | organ k, Brain (Br), Heart (H), Kidney (K), liver (L) etc. |
| Leth | Lethal |
| m | per unit mass |
| O2 | oxygen |
| Abbreviations | |
| ATP | Adenosine triphosphate |
| BMA | Body mass based allometry |
| Br, H, K, L, R | Brain, Heart, Kidney, Liver, Residual |
| BS | Biological systems |
| CH | Carbo-hydrate, e.g., glucose |
| COA | Capillary on Axis of Cylinder |
| COS | Capillary on Surface of Cylinder or solid cylinder model |
| ER | Equivalence radio in engineering, O2 used/O2 supplied, ER < 1 dilute mix |
| ERR | Energy Release Rate, W |
| F | Fat |
| GC | Group Combustion |
| ICD | Inter-capillary Distance, cm |
| IF | Interstitial Fluid |
| Iso | isolated |
| LM | Langmuir kinetics |
| MITO | Mitochondria |
| MM | Michaelis Menton kinetics |
| MR | Metabolic rate, W |
| OD | Oxygen Deficiency, oxygen deficient |
| ODC | Oxygen Deficient Combustion |
| ODM | Oxygen deficient metabolism |
| OEF | Oxygen Extraction Fraction (g O2 extracted for metabolism er g O in blood = Equivalence Ratio in Combustion. Science for dilute Combustible Mix) |
| OHb | Oxy-Hemoglobin |
| OMA | Organ mass based allometry |
| OXPHOS | Oxidative phosphorylation |
| P | Protein |
| ppm | parts per million {(g per/g blood) * 106} |
| RBC | Red Blood Cells |
| Sa | Saturation |
| SERR | Specific Energy Release Rate, W/g |
| SERRM | Specific Energy Release Rate, W/g body mass |
| SMRk | Specific Metabolic Rate (term in Biology for SERR) of organ k, W/g |
| SMRM | Specific Metabolic Rate of whole body, W/g body mass |
Appendix A. Solutions for Oxygen Concentrations with Lethal Volume
Appendix B. Collected Organ Mass and Metabolism Data On 111 Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | M | qL,m | qBr,m | qH,m | qK,m | qR,m | mL | mBr | mH | mK | mR | q Het, | q Het,m | mL * | mBr * | mH * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sorex araneus | 0.0075 | 123 | 43 | 77 | 50 | 3 | 0.00038 | 0.00015 | 0.00011 | 0.00011 | 0.0068 | 0.09 | 11.86 | 6.91 | 2.73 | 2.00 |
| Crocidura russula | 0.0100 | 115 | 42 | 75 | 49 | 3 | 0.00055 | 0.00017 | 0.00008 | 0.00013 | 0.0086 | 0.10 | 10.85 | 8.46 | 2.62 | 1.23 |
| Lasiurus borealis | 0.0140 | 105 | 40 | 72 | 48 | 3 | 0.00035 | 0.00017 | 0.00014 | 0.00011 | 0.013 | 0.10 | 7.17 | 6.36 | 3.09 | 2.55 |
| Lasionycteris noctivagans | 0.0150 | 102 | 39 | 71 | 47 | 3 | 0.00033 | 0.00016 | 0.00016 | 0.00013 | 0.014 | 0.10 | 6.68 | 5.08 | 2.46 | 2.46 |
| Mus musculus | 0.0150 | 102 | 39 | 71 | 47 | 3 | 0.00068 | 0.00036 | 0.00007 | 0.00028 | 0.014 | 0.14 | 9.49 | 4.86 | 2.57 | 0.50 |
| Myodes glareolus | 0.0150 | 101 | 39 | 71 | 47 | 3 | 0.00067 | 0.00035 | 0.0001 | 0.00024 | 0.014 | 0.14 | 9.10 | 5.58 | 2.92 | 0.83 |
| Microtus agrestis | 0.0150 | 101 | 39 | 71 | 47 | 3 | 0.00063 | 0.00039 | 0.00012 | 0.00017 | 0.014 | 0.14 | 8.86 | 7.41 | 4.59 | 1.41 |
| Neomys fodiens | 0.0160 | 101 | 39 | 70 | 47 | 3 | 0.00055 | 0.00025 | 0.00014 | 0.00022 | 0.015 | 0.13 | 8.14 | 5.00 | 2.27 | 1.27 |
| Blarina brevicauda | 0.0180 | 97 | 38 | 69 | 47 | 3 | 0.00093 | 0.00032 | 0.00018 | 0.00021 | 0.016 | 0.17 | 9.64 | 8.86 | 3.05 | 1.71 |
| Apodemus sylvaticus | 0.0180 | 97 | 38 | 69 | 47 | 3 | 0.0011 | 0.00057 | 0.00014 | 0.00026 | 0.016 | 0.19 | 10.70 | 8.46 | 4.38 | 1.08 |
| Microtus | 0.0210 | 93 | 37 | 68 | 46 | 3 | 0.0011 | 0.00058 | 0.00015 | 0.00036 | 0.019 | 0.19 | 8.91 | 6.11 | 3.22 | 0.83 |
| Peromyscus leucopus | 0.0220 | 92 | 37 | 68 | 46 | 3 | 0.0012 | 0.00074 | 0.00015 | 0.0003 | 0.02 | 0.22 | 9.88 | 8.00 | 4.93 | 1.00 |
| Apodemus flavicollis | 0.0250 | 89 | 37 | 67 | 45 | 3 | 0.001 | 0.00061 | 0.00018 | 0.00034 | 0.023 | 0.20 | 7.94 | 5.88 | 3.59 | 1.06 |
| Nyctalus noctula | 0.0260 | 88 | 36 | 66 | 45 | 3 | 0.0005 | 0.00032 | 0.00037 | 0.00013 | 0.024 | 0.15 | 5.76 | 7.69 | 4.92 | 5.69 |
| Microtus arvalis | 0.0270 | 87 | 36 | 66 | 45 | 3 | 0.0019 | 0.00039 | 0.00019 | 0.00055 | 0.024 | 0.28 | 10.31 | 6.91 | 1.42 | 0.69 |
| Mouse | 0.0280 | 86 | 36 | 66 | 45 | 3 | 0.0018 | 0.0005 | 0.00016 | 0.00051 | 0.025 | 0.27 | 9.64 | 7.06 | 1.96 | 0.63 |
| Gerbillus perpallidus | 0.0300 | 85 | 36 | 65 | 45 | 3 | 0.001 | 0.00058 | 0.00013 | 0.00027 | 0.028 | 0.19 | 6.54 | 7.41 | 4.30 | 0.96 |
| Mustela nivalis | 0.0320 | 83 | 35 | 65 | 44 | 3 | 0.0016 | 0.0018 | 0.00036 | 0.00043 | 0.028 | 0.31 | 9.49 | 7.44 | 8.37 | 1.67 |
| Acomys minous | 0.0420 | 77 | 34 | 63 | 43 | 2 | 0.0009 | 0.0009 | 0.00018 | 0.00032 | 0.04 | 0.23 | 5.33 | 5.63 | 5.63 | 1.13 |
| Jaculus jaculus | 0.0480 | 75 | 33 | 62 | 43 | 2 | 0.0011 | 0.0012 | 0.00045 | 0.00029 | 0.045 | 0.27 | 5.67 | 7.59 | 8.28 | 3.10 |
| Rhabdomys pumilio | 0.0500 | 74 | 33 | 61 | 43 | 2 | 0.0018 | 0.0006 | 0.00021 | 0.00041 | 0.047 | 5.96 | 0.00 | 8.78 | 2.93 | 1.02 |
| Talpa europaea | 0.0510 | 73 | 33 | 61 | 43 | 2 | 0.0015 | 0.001 | 0.00031 | 0.00036 | 0.048 | 0.30 | 5.71 | 8.33 | 5.56 | 1.72 |
| Glaucomys volans | 0.0550 | 72 | 33 | 61 | 43 | 2 | 0.0029 | 0.0019 | 0.00056 | 0.00059 | 0.049 | 0.45 | 8.09 | 9.83 | 6.44 | 1.90 |
| Arvicola terrestris | 0.0620 | 70 | 32 | 60 | 42 | 2 | 0.0026 | 0.0011 | 0.00028 | 0.0007 | 0.057 | 0.39 | 6.34 | 7.43 | 3.14 | 0.80 |
| Glis glis | 0.0830 | 64 | 31 | 58 | 41 | 2 | 0.0032 | 0.0015 | 0.00048 | 0.00068 | 0.078 | 0.48 | 5.71 | 9.41 | 4.41 | 1.41 |
| Tamias striatus | 0.1040 | 61 | 30 | 56 | 40 | 2 | 0.0029 | 0.0024 | 0.00066 | 0.00081 | 0.097 | 5.04 | 0.00 | 7.16 | 5.93 | 1.63 |
| Octodon degus | 0.1290 | 57 | 29 | 55 | 40 | 2 | 0.0048 | 0.0019 | 0.00041 | 0.0011 | 0.121 | 4.99 | 0.00 | 8.73 | 3.45 | 0.75 |
| Tupaia glis | 0.1410 | 56 | 29 | 54 | 39 | 2 | 0.0034 | 0.0034 | 0.00117 | 0.0011 | 0.132 | 4.69 | 0.00 | 6.18 | 6.18 | 2.13 |
| Rat | 0.1500 | 55 | 28 | 54 | 39 | 2 | 0.0092 | 0.0023 | 0.0007 | 0.0014 | 0.136 | 6.25 | 0.00 | 13.1 | 3.29 | 1.00 |
| Cebuella | 0.1630 | 54 | 28 | 53 | 39 | 2 | 0.0135 | 0.0044 | 0.00086 | 0.0019 | 0.142 | 1.25 | 0.00 | 14.2 | 4.63 | 0.91 |
| Rattus norvegicus | 0.2100 | 50 | 27 | 52 | 38 | 2 | 0.0092 | 0.0023 | 0.00087 | 0.0015 | 0.196 | 4.75 | 0.00 | 12.3 | 3.07 | 1.16 |
| Cheirogaleus medius | 0.2310 | 49 | 27 | 51 | 38 | 2 | 0.0063 | 0.0028 | 0.00093 | 0.001 | 0.22 | 3.80 | 0.00 | 12.6 | 5.60 | 1.86 |
| Rat | 0.2500 | 48 | 26 | 51 | 37 | 2 | 0.012 | 0.002 | 0.00094 | 0.0021 | 0.233 | 4.73 | 0.00 | 11.4 | 1.90 | 0.90 |
| Mustela erminea | 0.2590 | 48 | 26 | 51 | 37 | 2 | 0.01 | 0.0057 | 0.0025 | 0.0023 | 0.238 | 4.89 | 0.00 | 8.70 | 4.96 | 2.17 |
| Helogale parvula | 0.2600 | 48 | 26 | 51 | 37 | 2 | 0.0111 | 0.0052 | 0.0015 | 0.0025 | 0.24 | 4.84 | 0.00 | 8.88 | 4.16 | 1.20 |
| Sciurus vulgaris | 0.2750 | 47 | 26 | 50 | 37 | 2 | 0.0055 | 0.0063 | 0.0017 | 0.0017 | 0.259 | 3.77 | 0.00 | 6.47 | 7.41 | 2.00 |
| Callithrix jacchus | 0.3120 | 45 | 26 | 49 | 37 | 2 | 0.0178 | 0.0073 | 0.0028 | 0.0029 | 0.281 | 5.57 | 0.00 | 12.3 | 5.03 | 1.93 |
| Saguinus fuscicollis | 0.3300 | 45 | 25 | 49 | 37 | 2 | 0.0144 | 0.0078 | 0.0033 | 0.0019 | 0.303 | 1.60 | 0.00 | 15.2 | 8.21 | 3.47 |
| Rat | 0.3370 | 44 | 25 | 49 | 37 | 2 | 0.008 | 0.0019 | 0.001 | 0.0023 | 0.324 | 1.10 | 0.00 | 6.96 | 1.65 | 0.87 |
| Rat (Wistar) | 0.3900 | 43 | 25 | 48 | 36 | 2 | 0.0143 | 0.0019 | 0.0011 | 0.0028 | 0.37 | 1.44 | 0.00 | 10.2 | 1.36 | 0.79 |
| Sciurus niger | 0.4120 | 42 | 25 | 48 | 36 | 2 | 0.0107 | 0.0075 | 0.0025 | 0.003 | 0.389 | 1.52 | 0.00 | 7.13 | 5.00 | 1.67 |
| Sciurus carolinensis | 0.5960 | 38 | 23 | 46 | 35 | 2 | 0.0164 | 0.0075 | 0.0028 | 0.0032 | 0.566 | 1.94 | 0.00 | 10.3 | 4.69 | 1.75 |
| Saguinus oedipus | 0.6240 | 38 | 23 | 46 | 35 | 2 | 0.0209 | 0.01 | 0.0037 | 0.0031 | 0.586 | 2.21 | 0.00 | 13.5 | 6.45 | 2.39 |
| Mustela putorius | 0.6400 | 37 | 23 | 45 | 35 | 2 | 0.0288 | 0.0104 | 0.0048 | 0.004 | 0.592 | 2.60 | 4.06 | 14.4 | 5.20 | 2.40 |
| Leontopithecus chrysomelas | 0.6420 | 37 | 23 | 45 | 35 | 2 | 0.0189 | 0.0132 | 0.0038 | 0.0041 | 0.602 | 2.27 | 3.54 | 9.22 | 6.44 | 1.85 |
| Guinea pig | 0.8000 | 35 | 22 | 44 | 34 | 2 | 0.027 | 0.0047 | 0.0023 | 0.0056 | 0.76 | 2.49 | 3.11 | 9.64 | 1.68 | 0.82 |
| Potorous tridactylu | 0.8090 | 35 | 22 | 44 | 34 | 2 | 0.0237 | 0.0114 | 0.0048 | 0.0062 | 0.763 | 2.65 | 3.28 | 7.65 | 3.68 | 1.55 |
| Erinaceus europaeus | 0.9500 | 34 | 22 | 43 | 34 | 1 | 0.0496 | 0.0043 | 0.0055 | 0.0089 | 0.881 | 3.59 | 3.78 | 11.2 | 0.97 | 1.24 |
| Sylvilagus floridanus | 0.9720 | 33 | 22 | 43 | 33 | 1 | 0.032 | 0.0079 | 0.0048 | 0.0063 | 0.921 | 3.00 | 3.08 | 10.2 | 2.51 | 1.52 |
| Ondatra zibethicus | 0.9910 | 33 | 22 | 43 | 33 | 1 | 0.026 | 0.0047 | 0.003 | 0.0058 | 0.952 | 2.67 | 2.69 | 8.97 | 1.62 | 1.03 |
| Saimiri boliviensis | 1.0000 | 33 | 22 | 43 | 33 | 1 | 0.0194 | 0.029 | 0.0065 | 0.0067 | 0.941 | 3.14 | 3.13 | 5.79 | 8.66 | 1.94 |
| Martes foina | 1.4100 | 30 | 21 | 41 | 32 | 1 | 0.0349 | 0.019 | 0.0098 | 0.0073 | 1.335 | 3.92 | 2.79 | 9.56 | 5.21 | 2.68 |
| Mephitis | 1.4500 | 30 | 21 | 41 | 32 | 1 | 0.0174 | 0.0098 | 0.006 | 0.0066 | 1.409 | 3.11 | 2.15 | 5.27 | 2.97 | 1.82 |
| Trichosurus vulpecula | 1.5500 | 29 | 20 | 41 | 32 | 1 | 0.0332 | 0.0127 | 0.009 | 0.0135 | 1.482 | 4.03 | 2.60 | 4.92 | 1.88 | 1.33 |
| Martes martes | 1.6000 | 29 | 20 | 41 | 32 | 1 | 0.0379 | 0.0205 | 0.0108 | 0.0088 | 1.525 | 4.29 | 2.68 | 8.61 | 4.66 | 2.45 |
| Cebus apella | 1.7500 | 29 | 20 | 40 | 32 | 1 | 0.0493 | 0.0508 | 0.0134 | 0.0104 | 1.626 | 5.42 | 3.11 | 9.48 | 9.77 | 2.58 |
| Eulemur macaco macaco | 1.8800 | 28 | 20 | 40 | 32 | 1 | 0.0778 | 0.0242 | 0.0091 | 0.0142 | 1.75 | 5.76 | 3.07 | 10.9 | 3.41 | 1.28 |
| Chrotagale owstoni | 1.9600 | 28 | 20 | 40 | 32 | 1 | 0.0441 | 0.0233 | 0.0116 | 0.0128 | 1.868 | 4.99 | 2.53 | 6.89 | 3.64 | 1.81 |
| Vulpes corsac | 2.0800 | 27 | 20 | 40 | 31 | 1 | 0.0356 | 0.0341 | 0.0217 | 0.0088 | 1.975 | 5.33 | 2.56 | 8.09 | 7.75 | 4.93 |
| Lemur catta | 2.0800 | 27 | 20 | 40 | 31 | 1 | 0.0729 | 0.0228 | 0.0117 | 0.0112 | 1.956 | 5.76 | 2.77 | 13.0 | 4.07 | 2.09 |
| Eulemur fulvus fulvus | 2.5000 | 26 | 19 | 39 | 31 | 1 | 0.0434 | 0.0225 | 0.0118 | 0.0095 | 2.413 | 5.33 | 2.12 | 9.14 | 4.74 | 2.48 |
| Felis silvestris | 2.5700 | 26 | 19 | 39 | 31 | 1 | 0.0502 | 0.0381 | 0.0103 | 0.0154 | 2.459 | 5.91 | 2.31 | 6.52 | 4.95 | 1.34 |
| Didelphis virginiana | 2.6300 | 26 | 19 | 38 | 31 | 1 | 0.1573 | 0.0083 | 0.0121 | 0.0229 | 2.433 | 8.33 | 3.17 | 13.7 | 0.72 | 1.06 |
| Aonyx cinerea | 2.6800 | 25 | 19 | 38 | 31 | 1 | 0.1064 | 0.0359 | 0.0151 | 0.0306 | 2.487 | 7.99 | 2.98 | 6.95 | 2.35 | 0.99 |
| Leopardus geoffroyi | 3.1000 | 24 | 18 | 38 | 30 | 1 | 0.0584 | 0.0321 | 0.016 | 0.0307 | 2.963 | 7.12 | 2.30 | 3.80 | 2.09 | 1.04 |
| Lepus europaeus | 3.3400 | 24 | 18 | 37 | 30 | 1 | 0.0904 | 0.0148 | 0.0289 | 0.0185 | 3.186 | 7.85 | 2.35 | 9.77 | 1.60 | 3.12 |
| Dasyprocta punctata | 3.4000 | 24 | 18 | 37 | 30 | 1 | 0.1088 | 0.0228 | 0.0363 | 0.0213 | 3.211 | 8.81 | 2.59 | 10.2 | 2.14 | 3.41 |
| Potos flavus | 3.9200 | 23 | 18 | 37 | 30 | 1 | 0.1657 | 0.0311 | 0.0211 | 0.0144 | 3.688 | 9.83 | 2.51 | 23.0 | 4.32 | 2.93 |
| Dasyprocta azarae | 4.1000 | 23 | 18 | 37 | 30 | 1 | 0.0935 | 0.0238 | 0.0304 | 0.0227 | 3.93 | 8.81 | 2.15 | 8.24 | 2.10 | 2.68 |
| Varecia rubra | 4.2000 | 23 | 18 | 36 | 30 | 1 | 0.0722 | 0.0357 | 0.0181 | 0.0224 | 4.052 | 8.23 | 1.96 | 6.45 | 3.19 | 1.62 |
| Alouatta sara | 4.4000 | 22 | 18 | 36 | 30 | 1 | 0.0812 | 0.0565 | 0.024 | 0.0099 | 4.228 | 8.77 | 1.99 | 16.4 | 11.41 | 4.85 |
| Monkey | 4.5000 | 22 | 17 | 36 | 29 | 1 | 0.11 | 0.042 | 0.023 | 0.021 | 4.304 | 9.49 | 2.11 | 10.5 | 4.00 | 2.19 |
| Martes pennanti | 4.7900 | 22 | 17 | 36 | 29 | 1 | 0.113 | 0.0412 | 0.0274 | 0.0211 | 4.588 | 9.88 | 2.07 | 10.7 | 3.91 | 2.60 |
| Trachypithecus vetulus | 5.0000 | 22 | 17 | 36 | 29 | 1 | 0.09 | 0.072 | 0.0192 | 0.0154 | 4.803 | 9.64 | 1.93 | 11.7 | 9.35 | 2.49 |
| Lutrogale perspicillata | 5.1000 | 21 | 17 | 36 | 29 | 1 | 0.152 | 0.0622 | 0.0485 | 0.0485 | 4.789 | 12.7 | 2.50 | 6.27 | 2.56 | 2.00 |
| Chlorocebus pygerythrus | 5.3000 | 21 | 17 | 35 | 29 | 1 | 0.089 | 0.0808 | 0.0426 | 0.0121 | 5.076 | 10.7 | 2.02 | 14.7 | 13.36 | 7.04 |
| Lutra lutra | 5.3300 | 21 | 17 | 35 | 29 | 1 | 0.255 | 0.0478 | 0.0514 | 0.0611 | 4.91 | 15.2 | 2.85 | 8.35 | 1.56 | 1.68 |
| Proteles cristata | 5.4000 | 21 | 17 | 35 | 29 | 1 | 0.182 | 0.0399 | 0.0906 | 0.0243 | 5.063 | 13.9 | 2.59 | 14.9 | 3.28 | 7.46 |
| Agouti paca | 5.4600 | 21 | 17 | 35 | 29 | 1 | 0.14 | 0.0321 | 0.0176 | 0.0222 | 5.248 | 10.5 | 1.92 | 12.6 | 2.89 | 1.59 |
| Macaca nigra | 5.6000 | 21 | 17 | 35 | 29 | 1 | 0.095 | 0.1052 | 0.0239 | 0.0186 | 5.357 | 10.9 | 1.96 | 10.2 | 11.31 | 2.57 |
| Puma yagouaroundi | 5.9000 | 21 | 17 | 35 | 29 | 1 | 0.116 | 0.043 | 0.0296 | 0.0391 | 5.673 | 11.4 | 1.93 | 5.93 | 2.20 | 1.51 |
| Hylobates concolor | 6.5500 | 20 | 17 | 35 | 29 | 1 | 0.293 | 0.1378 | 0.0582 | 0.0352 | 6.026 | 17.6 | 2.68 | 16.7 | 7.83 | 3.31 |
| Prionailurus viverrinus | 7.3000 | 19 | 16 | 34 | 28 | 1 | 0.16 | 0.0529 | 0.0335 | 0.0559 | 6.998 | 14.0 | 1.92 | 5.72 | 1.89 | 1.20 |
| Macropus agilis | 7.7000 | 19 | 16 | 34 | 28 | 1 | 0.203 | 0.0308 | 0.0602 | 0.0463 | 7.36 | 15.4 | 1.99 | 8.77 | 1.33 | 2.60 |
| Lontra canadensis | 7.9000 | 19 | 16 | 34 | 28 | 1 | 0.255 | 0.0425 | 0.0541 | 0.0747 | 7.474 | 17.1 | 2.17 | 6.83 | 1.14 | 1.45 |
| Dolichotis patagonum | 8.4300 | 19 | 16 | 34 | 28 | 1 | 0.158 | 0.0365 | 0.0651 | 0.036 | 8.134 | 15.0 | 1.78 | 8.78 | 2.03 | 3.62 |
| Symphalangus syndactylus | 8.5000 | 19 | 16 | 33 | 28 | 1 | 0.294 | 0.143 | 0.0515 | 0.0437 | 7.968 | 18.8 | 2.21 | 13.5 | 6.54 | 2.36 |
| Colobus guereza | 9.7500 | 18 | 16 | 33 | 28 | 1 | 0.171 | 0.0865 | 0.037 | 0.0233 | 9.432 | 15.6 | 1.61 | 14.7 | 7.42 | 3.18 |
| Felis chaus | 9.8000 | 18 | 16 | 33 | 28 | 1 | 0.153 | 0.0497 | 0.0483 | 0.0819 | 9.467 | 16.8 | 1.71 | 3.74 | 1.21 | 1.18 |
| Lynx canadensis | 10.000 | 18 | 16 | 33 | 28 | 1 | 0.158 | 0.0826 | 0.0388 | 0.0549 | 9.666 | 16.5 | 1.65 | 5.76 | 3.01 | 1.41 |
| Dog | 10.000 | 18 | 16 | 33 | 28 | 1 | 0.42 | 0.075 | 0.085 | 0.07 | 9.35 | 22.7 | 2.27 | 12.0 | 2.14 | 2.43 |
| Hystrix indica | 11.300 | 17 | 15 | 32 | 27 | 1 | 0.255 | 0.0407 | 0.0562 | 0.0524 | 10.85 | 18.8 | 1.67 | 9.73 | 1.55 | 2.15 |
| Theropithecus gelada | 11.400 | 17 | 15 | 32 | 27 | 1 | 0.236 | 0.1409 | 0.0772 | 0.038 | 10.91 | 20.3 | 1.78 | 12.4 | 7.42 | 4.06 |
| Pudu puda | 13.000 | 17 | 15 | 32 | 27 | 1 | 0.206 | 0.0616 | 0.0505 | 0.0199 | 12.56 | 18.4 | 1.43 | 20.7 | 6.19 | 5.08 |
| Gazella gazella | 15.000 | 16 | 15 | 31 | 27 | 1 | 0.327 | 0.0793 | 0.12 | 0.0406 | 14.43 | 24.6 | 1.64 | 16.1 | 3.91 | 5.91 |
| Castor fiber | 15.600 | 16 | 15 | 31 | 27 | 1 | 0.345 | 0.0489 | 0.044 | 0.0783 | 15.05 | 23.5 | 1.51 | 8.81 | 1.25 | 1.12 |
| Macaca arctoides | 15.900 | 16 | 15 | 31 | 27 | 1 | 0.241 | 0.118 | 0.061 | 0.05 | 15.4 | 22.9 | 1.44 | 9.64 | 4.72 | 2.44 |
| Lynx lynx | 17.5000 | 15 | 14 | 31 | 26 | 1 | 0.264 | 0.0943 | 0.093 | 0.0795 | 16.97 | 25.6 | 1.47 | 6.64 | 2.37 | 2.34 |
| Capreolus capreolus | 20.000 | 15 | 14 | 30 | 26 | 1 | 0.48 | 0.1 | 0.16 | 0.08 | 19.18 | 32.4 | 1.62 | 12.00 | 2.50 | 4.00 |
| Cuon alpinus | 20.000 | 15 | 14 | 30 | 26 | 1 | 0.346 | 0.116 | 0.158 | 0.0764 | 19.3 | 30.6 | 1.53 | 9.06 | 3.04 | 4.14 |
| Dog | 20.400 | 15 | 14 | 30 | 26 | 1 | 0.447 | 0.096 | 0.153 | 0.092 | 19.6 | 32.2 | 1.58 | 9.72 | 2.09 | 3.33 |
| Mandrillus sphinx | 23.000 | 14 | 14 | 30 | 26 | 1 | 0.331 | 0.168 | 0.076 | 0.0499 | 22.4 | 29.8 | 1.30 | 13.3 | 6.73 | 3.05 |
| Papio hamadryas | 23.300 | 14 | 14 | 30 | 26 | 1 | 0.392 | 0.174 | 0.103 | 0.0803 | 22.5 | 32.5 | 1.39 | 9.76 | 4.33 | 2.57 |
| Zalophus californianus | 34.000 | 13 | 13 | 28 | 25 | 1 | 1.274 | 0.31 | 0.168 | 0.2059 | 32 | 56.2 | 1.65 | 12.4 | 3.01 | 1.63 |
| Hydrochaeris hydrochaeris | 34.000 | 13 | 13 | 28 | 25 | 1 | 0.696 | 0.084 | 0.104 | 0.1035 | 33 | 42.2 | 1.24 | 13.5 | 1.62 | 2.01 |
| Canis lupus chanco | 38.000 | 12 | 13 | 28 | 25 | 1 | 0.971 | 0.14 | 0.303 | 0.2069 | 36.4 | 56.3 | 1.48 | 9.39 | 1.35 | 2.93 |
| Sheep | 52.000 | 11 | 12 | 27 | 24 | 1 | 0.96 | 0.106 | 0.28 | 0.16 | 50.5 | 61.7 | 1.19 | 12.0 | 1.33 | 3.50 |
| Reference women | 58.000 | 10 | 12 | 21 | 21 | 1 | 1.4 | 1.2 | 0.24 | 0.275 | 54.9 | 66.1 | 1.14 | 10.2 | 8.73 | 1.75 |
| Human | 60.000 | 10 | 12 | 21 | 21 | 1 | 1.7 | 1.3 | 0.32 | 0.25 | 56.4 | 72.2 | 1.20 | 13.6 | 10.4 | 2.56 |
| Reference man | 70.000 | 10 | 12 | 21 | 21 | 1 | 1.8 | 1.4 | 0.33 | 0.31 | 66.2 | 80.7 | 1.15 | 11.6 | 9.03 | 2.13 |
| Panthera tigris altaica | 75.000 | 10 | 12 | 26 | 23 | 1 | 1.1 | 0.342 | 0.305 | 0.4246 | 72.8 | 84.8 | 1.13 | 5.2 | 1.61 | 1.44 |
| Hog | 125.00 | 9 | 11 | 24 | 22 | 1 | 1.6 | 0.12 | 0.35 | 0.26 | 123 | 110 | 0.88 | 12.3 | 0.92 | 2.69 |
| Dairy cow | 488.00 | 6 | 9 | 21 | 20 | 1 | 6.46 | 0.4 | 1.88 | 1.16 | 478 | 354 | 0.73 | 11.1 | 0.69 | 3.24 |
| Horse | 600.00 | 6 | 9 | 20 | 20 | 0 | 6.7 | 0.67 | 4.25 | 1.66 | 587 | 458 | 0.76 | 8.1 | 0.81 | 5.12 |
| Steer | 700.00 | 6 | 9 | 20 | 19 | 0 | 5 | 0.5 | 2.3 | 1 | 691 | 434 | 0.62 | 10.0 | 1.00 | 4.60 |
| Elephant | 6650.0 | 3 | 6 | 15 | 16 | 0 | 6.3 | 5.7 | 2.2 | 1.2 | 6635 | 2327 | 0.35 | 10.5 | 9.50 | 3.67 |
References
- Lemprecht, I. Calorimetry and thermodynamics of living systems. Thermochim. Acta 2003, 405, 1–13. [Google Scholar] [CrossRef]
- Wenger, B.C. Chapter 29: The regulation of body temperature. In Medical Physiology: Principles for Clinical Medicine, 3rd ed.; Rhoades, R.A., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2009; p. 816. ISBN 9780781768528. [Google Scholar]
- Bellamy, K.; Laban, K.L.; Barrett, K.E.; Talbot, C.S. x Detection of viruses and body fluids which may contain viruses in the domestic environment. Epidemiol. Infèct. 1998, 121, 673–680. [Google Scholar] [CrossRef]
- Dobson, G.P. A Comparative View on being the right size: Heart design, mitochondrial efficiency and lifespan potentialx. Clin. Exp. Pharmacol. Physiol. Muscle Mech. Energetics 2003, 30, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, K.; Puri, I.; Jog, M. Advanced Thermodynamics Engineering, 2nd ed.; Chapter 14: Biology and Thermodynamics; Taylor and Francis (CRC Press): Boca Raton, FL, USA, 2011. [Google Scholar]
- Singer, D.; Bach, F.; Bretschneider, H.J.; Kuhn, H.-J. Microcalorimetric monitoring of ischemic tissue metabolism: Influence of incubation conditions and experimental animal species. Thermochim. Acta 1991, 187, 55–69. [Google Scholar] [CrossRef]
- Boron Walter, F.; Boulpaep Emile, L. Medical Physiology, 2nd ed.; Saunders Elsevier: New York, NY, USA, 2009. [Google Scholar]
- Horowiz, G.L. Oxygen Concentration of Blood: PO2, Co-Oximetry, and More, Beth Israel Medical Center. 2013. Available online: https://www.aacc.org/~/media/files/meetings-and-events/resources-from-past-events/conferences/2013/professional-practice/april-28/gc_oxygen_concentration_of_blood_apr_28_2013.pdf?la=en (accessed on 1 June 2021).
- Atkins, P.; de Paula, J. Physical Chemistry, 7th ed.; W H Freeman Company: New York, NY, USA, 2002. [Google Scholar]
- Available online: http://www.scymed.com/en/smnxpr/prgdb276.htm (accessed on 2 June 2021).
- Elnara, M.; Negri Bruna, M.P.; Luciana, K.M.; Carlos, V.P.; Shari, A.E.; Marcelo, A.F.; Elbio, A.D.; Daniel, D. Heparin therapy improving hypoxia in COVID-19 patients—A case series. MedRxiv 2020. [Google Scholar] [CrossRef]
- Miller, J. Oxygen Transport and Application of Oxygen Deficient Group Combustion within Vital Organ, MS, Mech Engg; Texas A&M University: College Station, TX, USA, 2014. [Google Scholar]
- Annamalai, K. Respiratory quotient (Rq), exhaust gas analyses, CO2 Emission and applications in automobile engineering. Adv. Automob. Eng. 2013, 2. [Google Scholar] [CrossRef] [Green Version]
- Annamalai, K.; Thanapal, S.; Ranjan, D. Ranking renewable and fossil fuels on global warming potential using respiratory quotient (RQ) Concept. J. Combust. 2018, 2018, 1270708. [Google Scholar] [CrossRef] [Green Version]
- Popovic, M. Thermodynamic properties of microorganisms: Determination and analysis of enthalpy, entropy, and Gibbs free energy of biomass, cells and colonies of 32 microorganism species. Heliyon 2019, 5, e0195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, D.K.; DiMaio, V.J. Normal organ weights in men: Part II—The brain, lungs, liver, spleen, and kidneys. Am. J. Forensic. Med. Pathol. 2012, 33, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; O’Connor, T.P.; Heshka, S.; Heymsfield, S.B. The reconstruction of Kleiber’s law at the organ-tissue level. J. Nutr. 2011, 131, 2967–2970. [Google Scholar] [CrossRef] [Green Version]
- Kleiber, M. Body size and metabolism. Hilgardia 1932, 6, 315–353. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ying, Z.; Bosy-Westphal, A.; Zhang, J.; Schautz, B.; Later, W. Specific metabolic rates of major organs and tissues across adulthood: Evaluation by mechanistic model of resting energy expenditure. Am. J. Clin. Nutr. 2010, 92, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, K.; Nanda, A. Biological aging and life span based on entropy stress via organ and mitochondrial metabolic loading. Entropy 2017, 19, 566. [Google Scholar] [CrossRef]
- Elia, M. Organ and tissue contribution to metabolic rate. In Energy Metabolism: Tissue Determinants and Cellular Corollaries; Raven Press, Ltd.: New York, NY, USA, 1992; pp. 61–79. [Google Scholar]
- Krebs, A.H. Body size and tissue respiration. Biochem. Biophys. Acta 1950, 4, 249–269. [Google Scholar] [CrossRef]
- Porter, R.K. Allometry of mammalian cellular oxygen consumption. Cell. Mol. Life Sci. 2001, 58, 815–822. [Google Scholar] [CrossRef]
- Gutierrez, W.R. xSite Model of Allometric Scaling and Fractal Distribution Networks of Organs 2019. Available online: https://arxiv.org/pdf/q-bio/0404039 (accessed on 26 February 2019).
- Singer, D. Size relationship of metabolic rate: Oxygen availability as the missing link between structure and function? Review. Thermochim. Acta 2006, 446, 20–28. [Google Scholar] [CrossRef]
- Annamalai, K.; Ryan, W.; Dhanapalan, S. Interactive processes in gasification and combustion-III: Coal particle arrays, streams and clouds. J. Prog. Energy Combust. Sci. 1994, 20, 487–618. [Google Scholar] [CrossRef]
- Chiu, H.; Liu, T. Group combustion of liquid droplets. Combust. Sci. Technol. 1977, 17, 127–140. [Google Scholar] [CrossRef]
- Sirignano, W.A. Fluid Dynamics and Transport of Droplets and Sprays, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; ISBN 9781107428003. [Google Scholar]
- Annamalai, K.; Puri, I.K. Combustion Science and Engineering; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2007. [Google Scholar]
- Gilot, P.; Bonnefoy, F.; Marcuccilli, F.; Prado, G. Determination of kinetic data for soot oxidation. modeling of competition between oxygen diffusion and reaction during thermogravimetric analysis. Combust. Flame 1993, 95, 87–100. [Google Scholar] [CrossRef]
- Forster, R.E. Is Oxygen Essentail Nutrient? Annu. Rev. Nutr. 1993, 13, 383–403. [Google Scholar] [CrossRef] [PubMed]
- Trayhun, P. Oxygen—A Critical, but Overlooked, Nutrient. Front. Nutr. Hypothesis Theory Artic. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Li, H.; Chen, J.; Qian, Q. Lactic Acid: No Longer an Inert and End Product of Glycolysis. Physiology 2017, 32, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, L.; Supuran, C.T.; Alfarouk, K.O. The Warburg Effect and the Hallmarks of Cancer. Anticancer. Agents Med. Chem. 2017, 17, 164–170. [Google Scholar] [CrossRef]
- Seyfried, T.N.; Shelton, L.M. Cancer as a metabolic disease. Nutr. Metab. 2010, 7, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Robert, J.; Gillies, R.J.; Robey, J.; Gatenby, R.A. Causes and Consequences of Increased Glucose Metabolism of Cancers. J. Uclear Med. 2008, 49, 24S–42S. [Google Scholar]
- Lázaro, L. The warburg effect: Why and how do cancer cells activate glycolysis in the presence of oxygen? Anticancer. Agents Med. Chem. 2008, 8, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A. The number and distribution of capillaries in muscles with calculations of the oxygen pressure head necessary for supplying the tissue. J. Physiol. 1919, 52, 409–415. [Google Scholar] [CrossRef]
- Daniel, G. Theoretical Models of Microvascular Oxygen Transport to Tissue. Microcirculation 2008, 15, 795–811. [Google Scholar] [CrossRef] [Green Version]
- Spalding, D.B. A Theory of Inflammability Limits and Flame-Quenching. Proc. R. Soc. Lond. Ser. A 1957, 240, 83–100. [Google Scholar]
- Piiper, P.; Scheid, J. Cross-sectional PO2 distributions in Krogh cylinder and solid cylinder models. Respir Physiol. 1986, 64, 241–251. [Google Scholar] [CrossRef]
- McMurtrey, R.J. Analytic Models of Oxygen and Nutrient Diffusion, Metabolism Dynamics, and Architecture Optimization in Three-Dimensional Tissue Constructs with Applications and Insights in Cerebral Organoidsvol. Tissue Eng. Part C Methods 2016, 22, 221–249. [Google Scholar] [CrossRef] [Green Version]
- Erekinsca, M.; Silver, I.A. ATP and Brain Function. J. Blood Flow Metab. 1989, 9, 2–19. [Google Scholar]
- Available online: https://www.health.harvard.edu/blog/the-hidden-long-term-cognitive-effects-of-covid-2020100821133 (accessed on 2 June 2021).
- Cavezzi ATroiani, E.; Corrao, S. COVID-19: Hemoglobin, iron, and hypoxia beyond inflammation. A narrative review. Clin. Pract. 2020, 10, 1271. [Google Scholar]
- Frontera, J.A.; Sabadia, S.; Lalchan, R.; Fang, T.; Flusty, B.; Millar-Vernetti, P.; Galetta, S. A Prospective Study of Neurologic Disorders in Hospitalized Patients with COVID-19 in New York City. J. Neurol. 2021, 96, e575–e586. [Google Scholar] [CrossRef] [PubMed]
- Granchi, C.; Bertini, S.; Macchia, M.; Minutolo, F. Inhibitors of lactate dehydrogenase isoforms and their therapeutic potentials. Curr. Med. Chem. 2010, 17, 672–697. [Google Scholar] [CrossRef]
- Melkonian, E.A.; Schury, M.P. Biochemistry, Anaerobic Glycolysis. [Updated 21 August 2019]. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK546695/ (accessed on 2 June 2021).
- Singer, D.; Schunck, O.; Bach, F.; Kuhn, H.J. Size effects on metabolic rate in cell, tissue, and body calorimetry. Thermochim. Acta 1995, 251, 227–240. [Google Scholar] [CrossRef]
- Kapteijn, F.; Marin, G.B.; Moulijn, J.A. Catalytic Reaction Engineering. In Catalysis: An Integrated Approach; Elsevier: New York, NY, USA, 1999; ISBN 9780444829634. [Google Scholar]
- Wang, Z.; Zhang, J.; Ying, Z.; Heymsfield, S.B. Organ-Tissue Level Model of Resting Energy Expenditure Across Mammals: New Insights into Kleiber’s Law. Int. Sch. Res. Netw. ISRN Zool. 2012, 9. [Google Scholar] [CrossRef] [Green Version]
- Dawson, T.H. Scaling laws for capillary vessels of mammals at rest and in exercise. Proc. R. Soc. Lond. B 2003, 270, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Hulbert, A.J. A Sceptics View: Kleiber’s Law or the 3/4 Rule is neither a Law nor a Rule but Rather an Empirical Approximation. Systems 2014, 2, 186–202. [Google Scholar] [CrossRef] [Green Version]
- Ahulwalia, A. Allometric Scaling In-Vitro, Scientific Reports. 2017. Available online: www.nature.com/scientificreports (accessed on 1 July 2021). [CrossRef] [PubMed]
- Zheng, J. Energy metabolism of cancer: Glycolysis versus oxidative phosphorylation (Review). Oncol. Lett. 2012, 4, 1151–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.V. Links between metabolism and cancer. Genes Dev. 2012, 26, 877–890. Available online: www.genesdev.org (accessed on 1 July 2021). [CrossRef] [PubMed] [Green Version]
- Thomas, F.; Nesse, R.M.; Gatenby, R.; Gidoi, C.; Renaud, F.; Roche, B.; Ujvari, B. Evolutionary Ecology of Organs: A Missing Link in Cancer Development? Trends Cancer 2016, 2, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Epstein, T.; Gatenby, R.A.; Joel SBrown, J.S. Fluctuations in energy demand. PLoS ONE 2017. [Google Scholar] [CrossRef] [Green Version]
- Cooper, G.M. The Cell: A Molecular Approach, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2000. [Google Scholar]












| Fuel | O2/Fuel Stoich. Ratio, νO2, st | Air/fuel Mass Ratio | RQ a | HV (Per Fuel) b | HVO2 (Per O2) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Molar | Mass | (kJ/mol) | (kJ/g) | (kJ/mol) | (kJ/g) | (kJ/L) c | |||
| gasoline (CH2.46) | 1.62 | 3.58 | 7.71 | 0.6 | 698.5 | 48.3 | 431.2 | 13.5 | 17.6 |
| methane (CH4) | 2 | 4 | 9.52 | 0.5 | 890.3 | 55.5 | 445.2 | 13.9 | 18.2 |
| coal-dry, ash-free d (CH0.714O0.18) | 1.09 | 2.23 | 6.22 | 0.9 | 465.2 | 29.8 | 427.4 | 13.4 | 17.4 |
| cattle manure-dry, ash free d (CH1.78O0.64N0.08S0.014) | 1.1 | 1.42 | 5.43 | 0.9 | 530.2 | 20.0 | 431.0 | 14.0 | 18.3 |
| glucose e (C6H12O6) | 6 | 1.07 | 28.56 | 1.0 | 2813 | 15.6 | 468.9 | 14.7 | 19.1 |
| Fat (C16H32O2) | 23 | 2.87 | 12.39 | 0.7 | 10,015 | 39.13 | 436.3 | 13.6 | 17.8 |
| protein d (C4.57H9.03N1.27 O2.25S0.046) | 5.7 | 1.54 | 9.61 | 0.8 | 2718 | 22.8 | 476.7 | 14.9 | 19.5 |
| Organ | Density, ρk, g/cc | ck, kg | dk | ek f | fk g | mk in kg for 85 kg Human | Ek | Fk | qk,m for 85 kg Human | Jk (Assumed YO2in = 300 ppm) [20] | Lk [20] | OEFk for 84 kg Human |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Brain (Br) | 1.036 | 0.011 | 0.76 | 21.62 | 0.14 | 0.32 | 9.42 | −0.184 | 0.044 | 0.524 | −0.084 | 0.37 |
| Heart (H) | 1.06 | 0.006 | 0.98 | 43.11 | 0.12 | 0.47 | 23.04 | −0.122 | 0.15 | 0.257 | −0.146 | 0.48 |
| Kidneys (K)h | 1.05 | 0.007 | 0.85 | 33.41 | 0.08 | 0.31 | 20.94 | −0.094 | 0.11 | 0.0863 | −0.004 | 0.085 |
| Liver (L) | 1.06 | 0.0330 | 0.87 | 33.11 | 0.27 | 1.57 | 11.49 | −0.310 | 0.19 | 1.558 | −0.256 | 0.52 |
| Residual Mass (R)- | 0.939 | 1.01 | 1.45 | 0.17 | 83.44 | 1.44 | −0.168 | 0.19 | - | - |
| # | Geometry | Carbon Dust Cloud | Cell Clouds-COS-O2 |
|---|---|---|---|
| 1 | O2 consumption rate per particle or per cell | ||
| 2 | Characteristic O2 consumption rate per particle (Cch,p) or Characteristic O2 consumption rate per cell (Cch,cell) | , kLM, Langmuir Constant YO2<<kLM First order Langmuir (LM) kinetics | YO2<<kMM First order Michaelis Menten (MM) kinetics |
| 3 | Bulk phase within cloud | Gas of very low density (order of 1 g/cm3) compared to carbon particles of density 1300 g/cm3. | Interstitial fluid (IF) with similar densities for cell and fluid |
| 4 | Conservation Equations (dimensional) | ||
| 5 | Conservation Equations (non-dimensional) | ||
| 6 | Dimensionless G number | , first order kinetics | ,, first order kinetics |
| 7 | Boundary Conditions with lethal volume | - | YO2 = YO2, cap-IF at r = RdYO2/dr = 0 at r = rleth (Figure 7b) |
| 8 | Boundary Conditions without lethal volume | YO2 = YO2, cl at r = R dYO2/dr = 0 at r = 0 | YO2 = YO2, cap-IF at r = R dYO2/dr = 0 at r = 0 (Figure 7a) |
| 9 | Oxygen Profiles with lethal Volume, , | - | See Appendix A (Equations (A2) and (A3)) |
| 10 | Oxygen Profiles without lethal Volume, , [26] | ||
| 11 | Oxygen Mass Fraction at core {without lethal volume}, , [26] | ||
| 12 | Incipient Group/OD Combustion 1 | YO2,0→YO2ext, solve for Ginc | YO2,0→YO2,leth, solve for GOD,inc |
| 13 | Effectiveness factor, ηeff = (YO2avg/YO2cap-IF) at G, [26], without lethal volume | , 1-(G/15)→1 as G→0 , SERR ∝ mcl F, Fk→ −1/3, G > 100 | SERR ∝ mk Fk {biology}, Fk →0 (isometric law) as G→0 ηeff →, Fk→ −1/3, GOD > 100 |
| # | Allometric Constants | Kidneys | Brain | Heart | Liver | Residual |
|---|---|---|---|---|---|---|
| 1 | gk,6 = (ck,6/cK,6) | 1 | 1.71 | 3.14 | 9.43 | 268.29 |
| 2 | hk,6 = (dk,6-dK,6) | 0 | 0.13 | −0.09 | 0.02 | 0.16 |
| 3 | mk,6* = (mk,6/mK,6) | 1 | 3.3–2.1 | 1.6–3.1 | 9.3–10.3 | - |
| 4 | Fk | −0.094 | −0.184 | −0.122 | −0.31 | - |
| 5 | GOD,k from Fk (6 species) | 2.8 | 11 | 4.2 | 240 | |
| 6 | Median mk* (6 species) with ck and dk for 6 species | 1 | 2.66 | 2.28 | 9.82 | 384.91 |
| 7 | Effectiveness factor or YO2 avg/YO2cap-IF | 0.85 | 0.63 | 0.8 | 0.18 |
| ## | Allometric Constants | Kidneys | Brain | Heart | Liver | Residual |
|---|---|---|---|---|---|---|
| 1 | ik,111 | 0.211 | 0.249 | 0.233 | 0.947 | 1.364 |
| 2 | jk,111 | 0.7441 | 0.8137 | 0.6446 | 0.6046 | 0.8402 |
| 3 | ck,111 = ik,111/ek,6 {ck,6} | 0.0063 (0.007) | 0.011 (0.011) | 0.0058 (0.006) | 0.029 (0.033) | 0.94 (0.939) |
| 4 | dk,111 = jk,111 − fk,6 {dk,6} | 0.83 (0.85) | 0.79 (0.76) | 0.93 (0.98) | 0.8723 (0.87) | 1.01 (1.01) |
| 5 | Fk,111 (Fk,6) | −0.101 (−0.098) | −0.181 (−0.187) | −0.126 (−0.121) | −0.307 (−0.308) | −0.166 (−0.165) |
| 6 | gk,111 = (ck,111/cK,111) | 1 | 3.42 | 1.84 | 9.07 | 298.05 |
| 7 | hk,111 = (dk,111 − dK,111) | 0 | −0.041 | 0.10 | 0.045 | 0.18 |
| 8 | mk,111* = (mk,111/mK,111) | 1 | 3.5–2.8 | 1.7–2.9 | 8.8–11.2 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annamalai, K. Oxygen Deficient (OD) Combustion and Metabolism: Allometric Laws of Organs and Kleiber’s Law from OD Metabolism? Systems 2021, 9, 54. https://doi.org/10.3390/systems9030054
Annamalai K. Oxygen Deficient (OD) Combustion and Metabolism: Allometric Laws of Organs and Kleiber’s Law from OD Metabolism? Systems. 2021; 9(3):54. https://doi.org/10.3390/systems9030054
Chicago/Turabian StyleAnnamalai, Kalyan. 2021. "Oxygen Deficient (OD) Combustion and Metabolism: Allometric Laws of Organs and Kleiber’s Law from OD Metabolism?" Systems 9, no. 3: 54. https://doi.org/10.3390/systems9030054
APA StyleAnnamalai, K. (2021). Oxygen Deficient (OD) Combustion and Metabolism: Allometric Laws of Organs and Kleiber’s Law from OD Metabolism? Systems, 9(3), 54. https://doi.org/10.3390/systems9030054