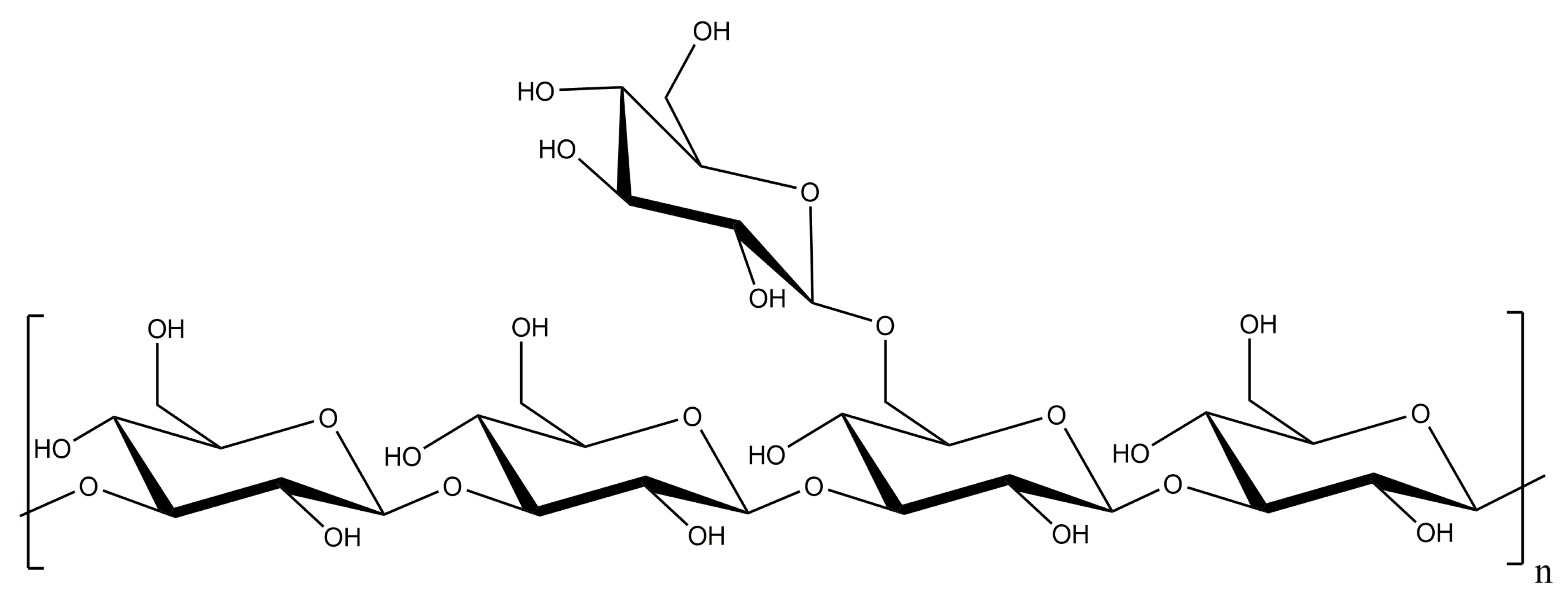
Laminarin Effects, a β-(1,3)-Glucan, on Skin Cell Inflammation and Oxidation

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Treatment
2.3. Cell Viability Assay
2.4. Cell Cytotoxicity Assay
2.5. Hyaluronic Acid (HA) Dosage
2.6. Type I Procollagen Dosage
2.7. Intracellular Reactive Oxygen Species (ROS) Assay
2.8. Effect of Laminarin with Cells—Glycans Interaction Study
2.9. Effect of Laminarin with Cells—Carbohydrate Receptor Study
2.10. Cytokine Secretions Study
2.11. Statistic Analysis
3. Results
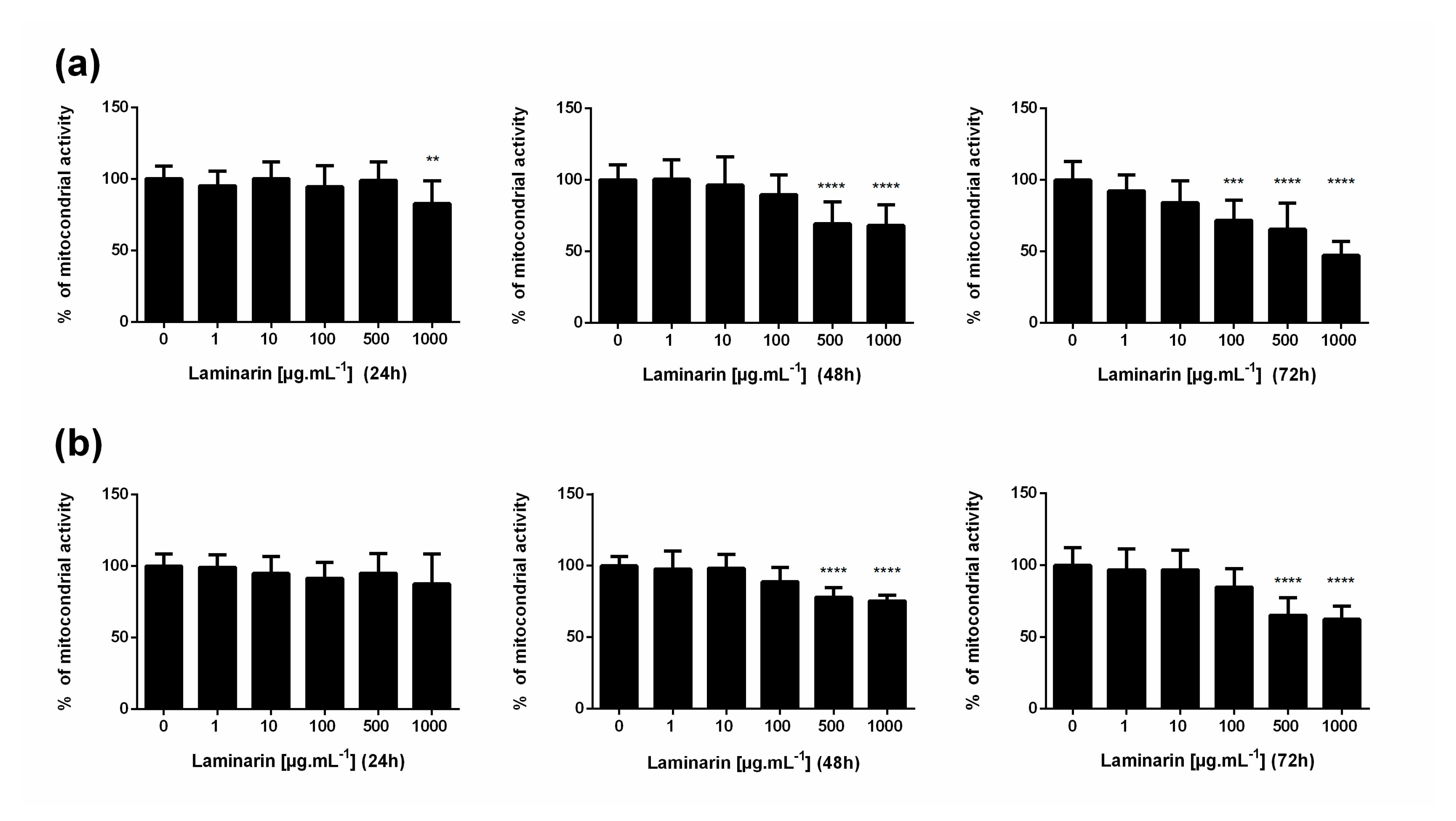
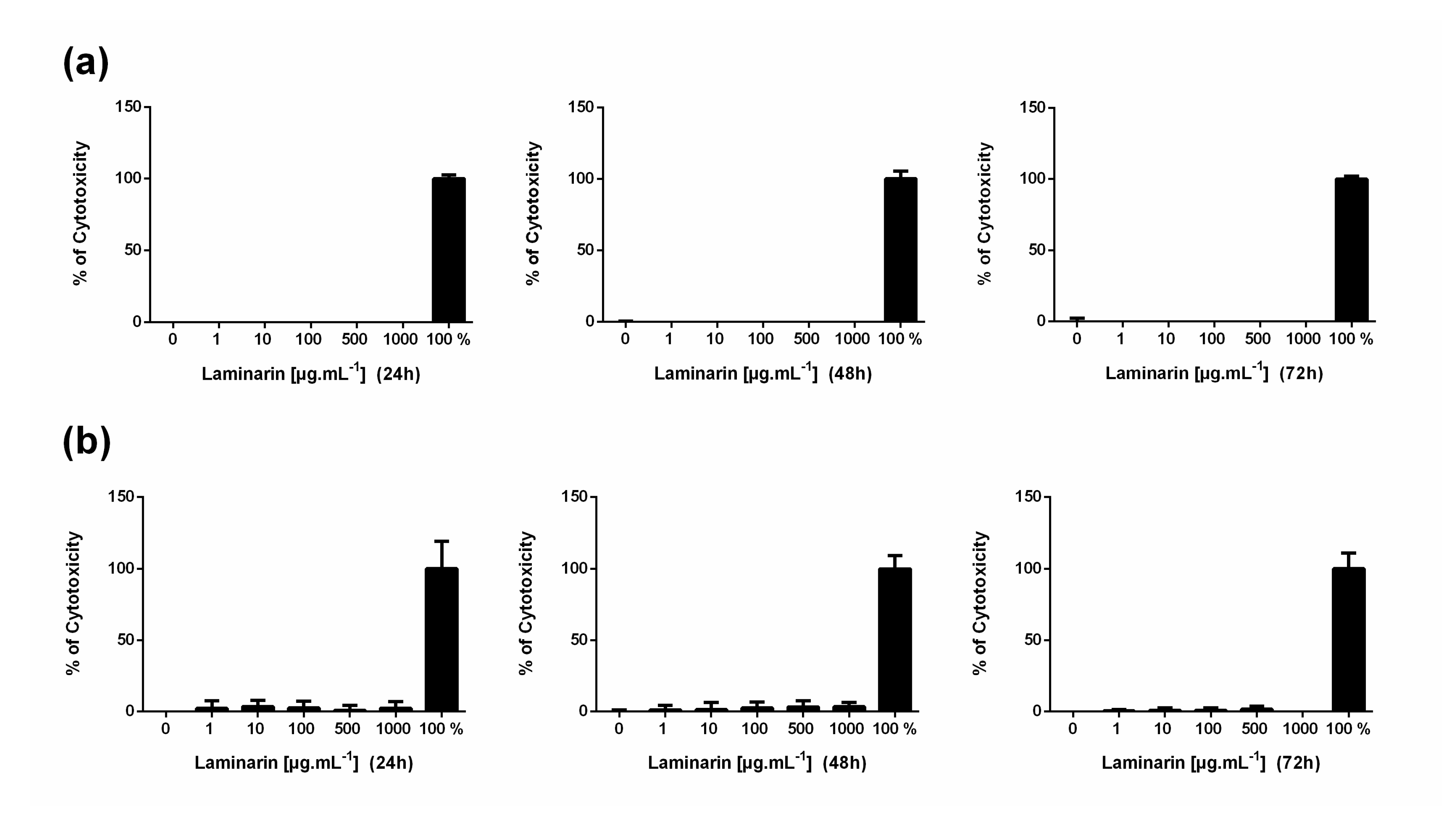
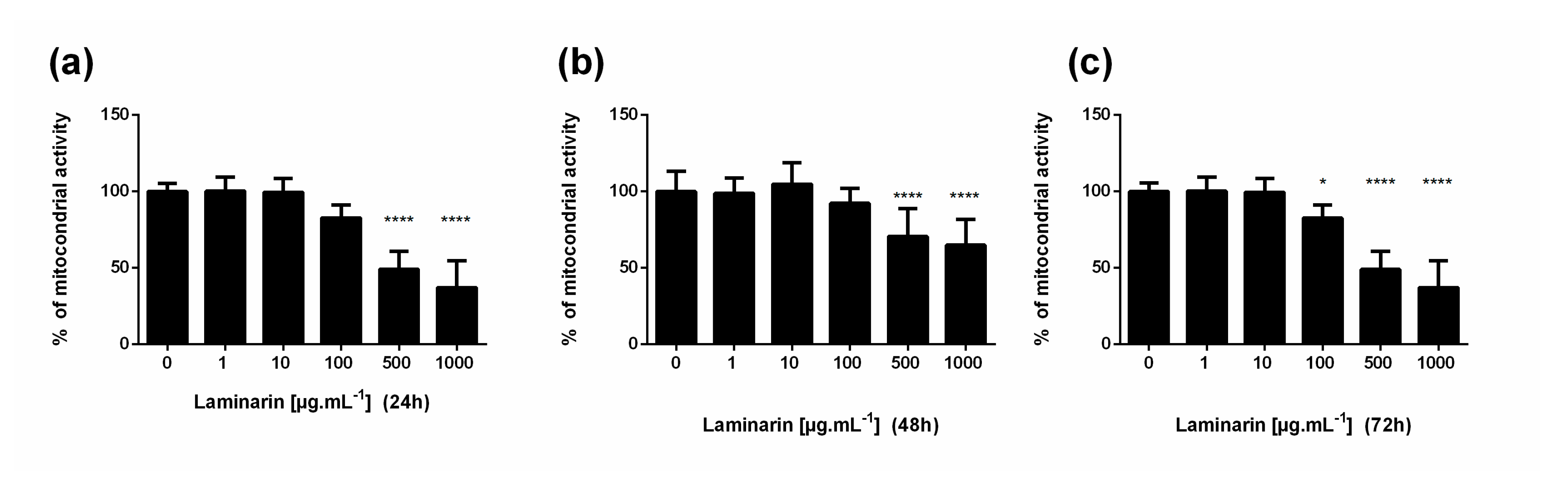
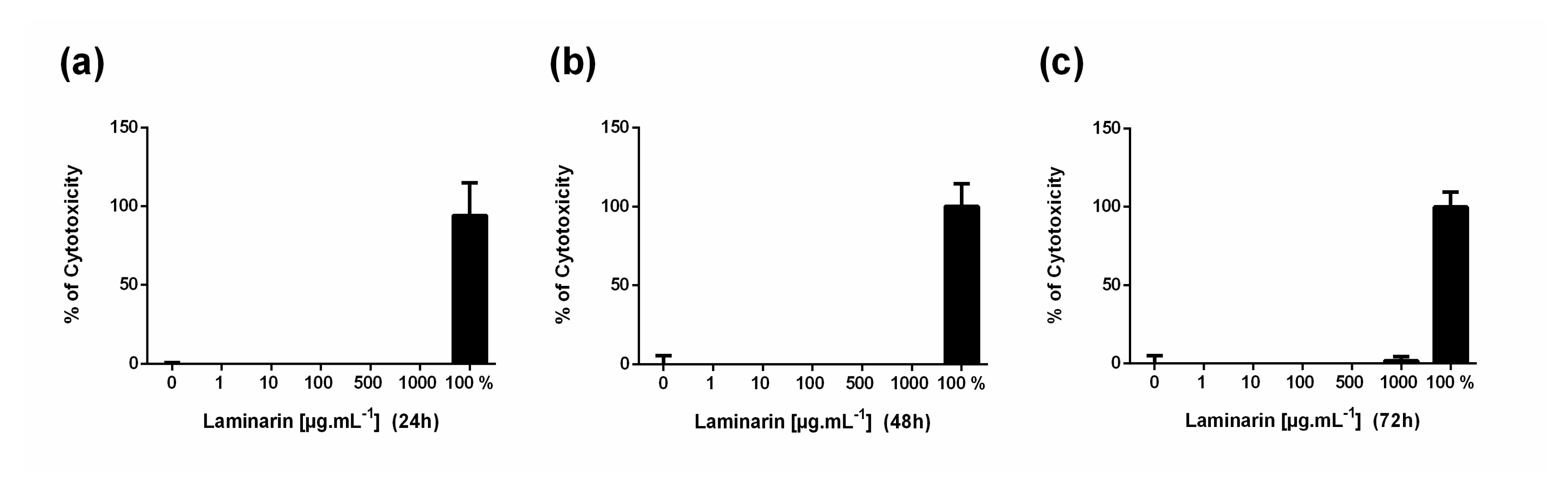
3.1. High Concentration of Laminarin Reduces Cellular Metabolic Activity without Cytolytic Activity
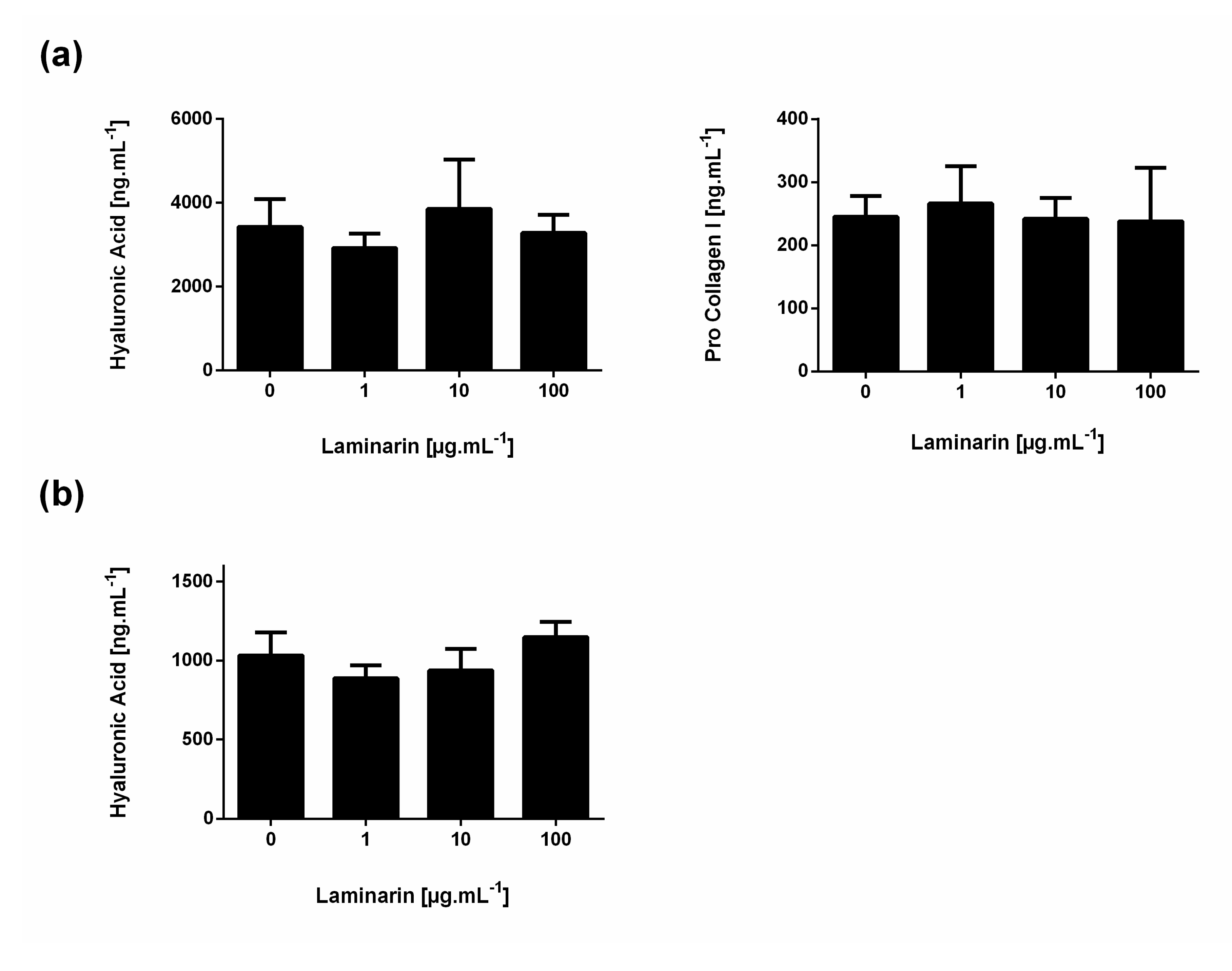
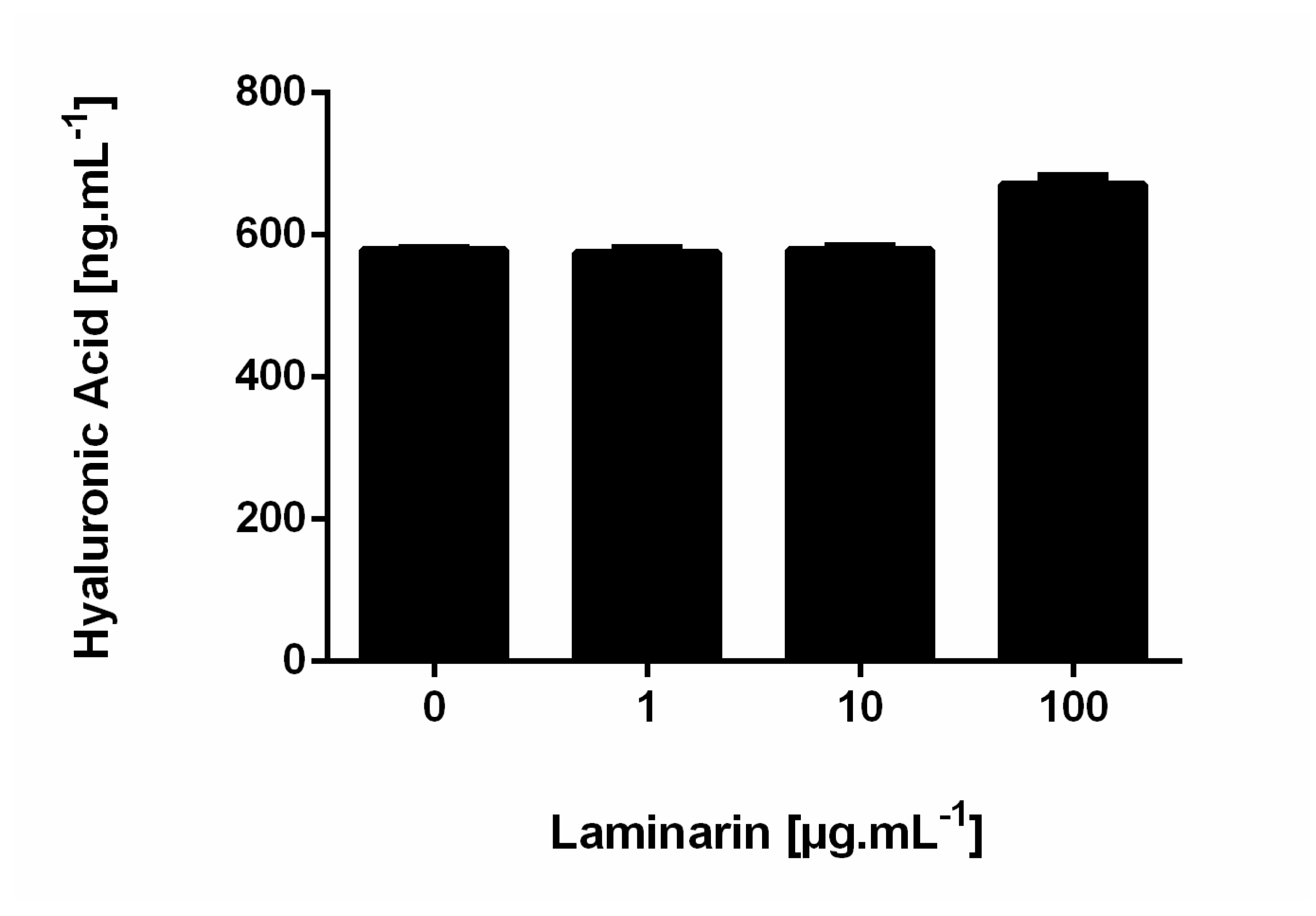
3.2. Influence of Laminarin on Matrix Deposition
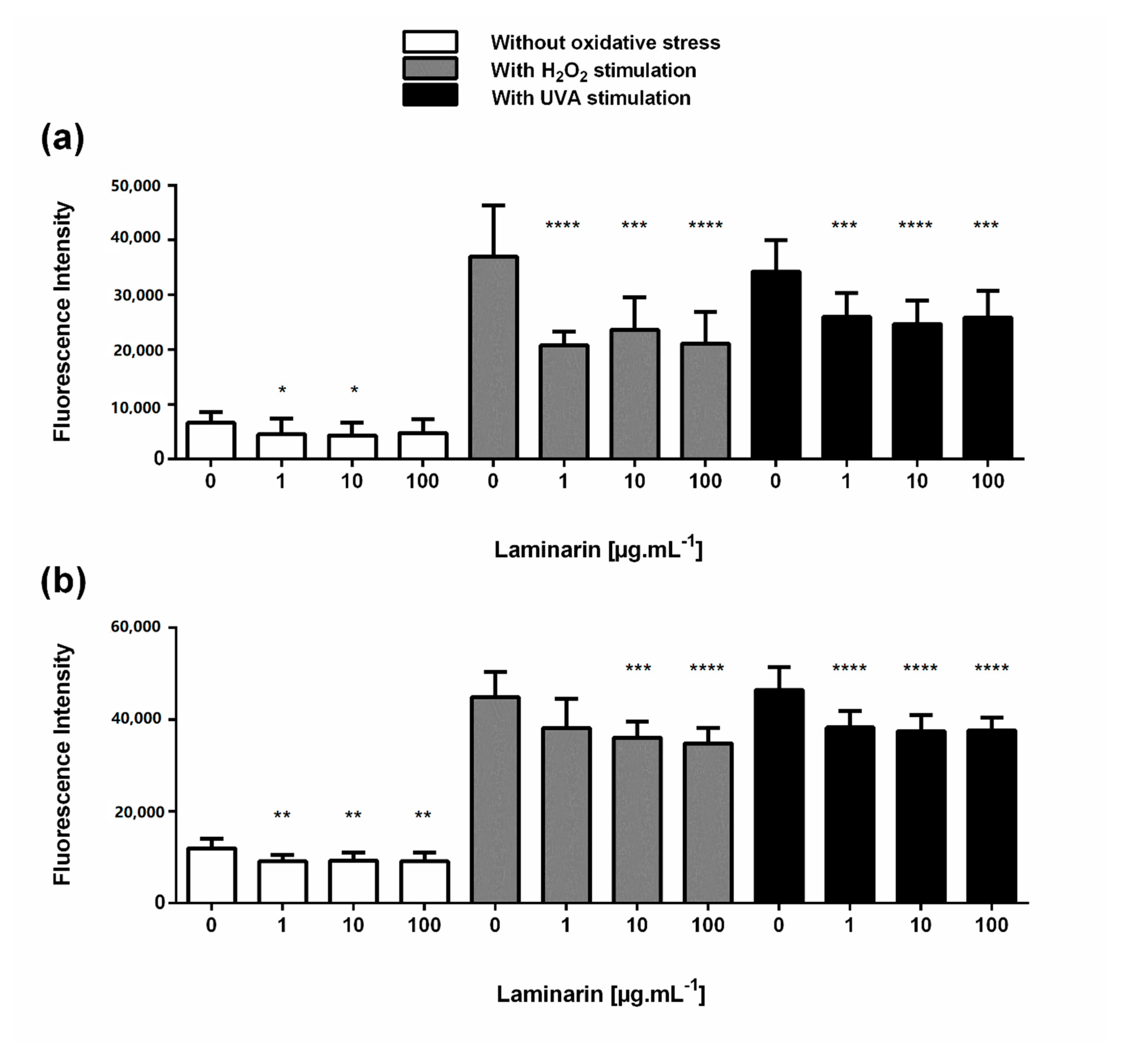
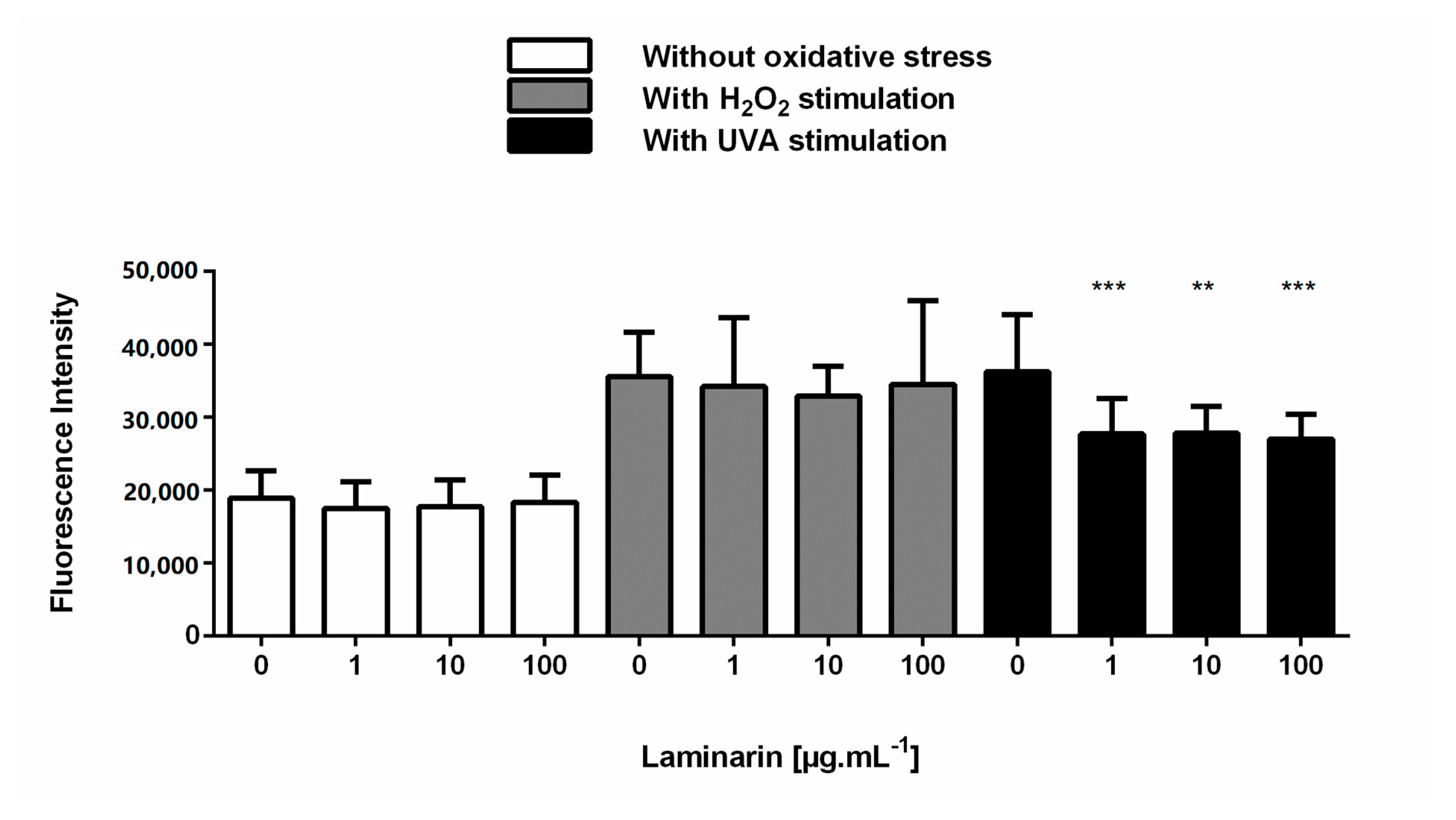
3.3. Laminarin Reduced Oxidative Stress on Skin Cells
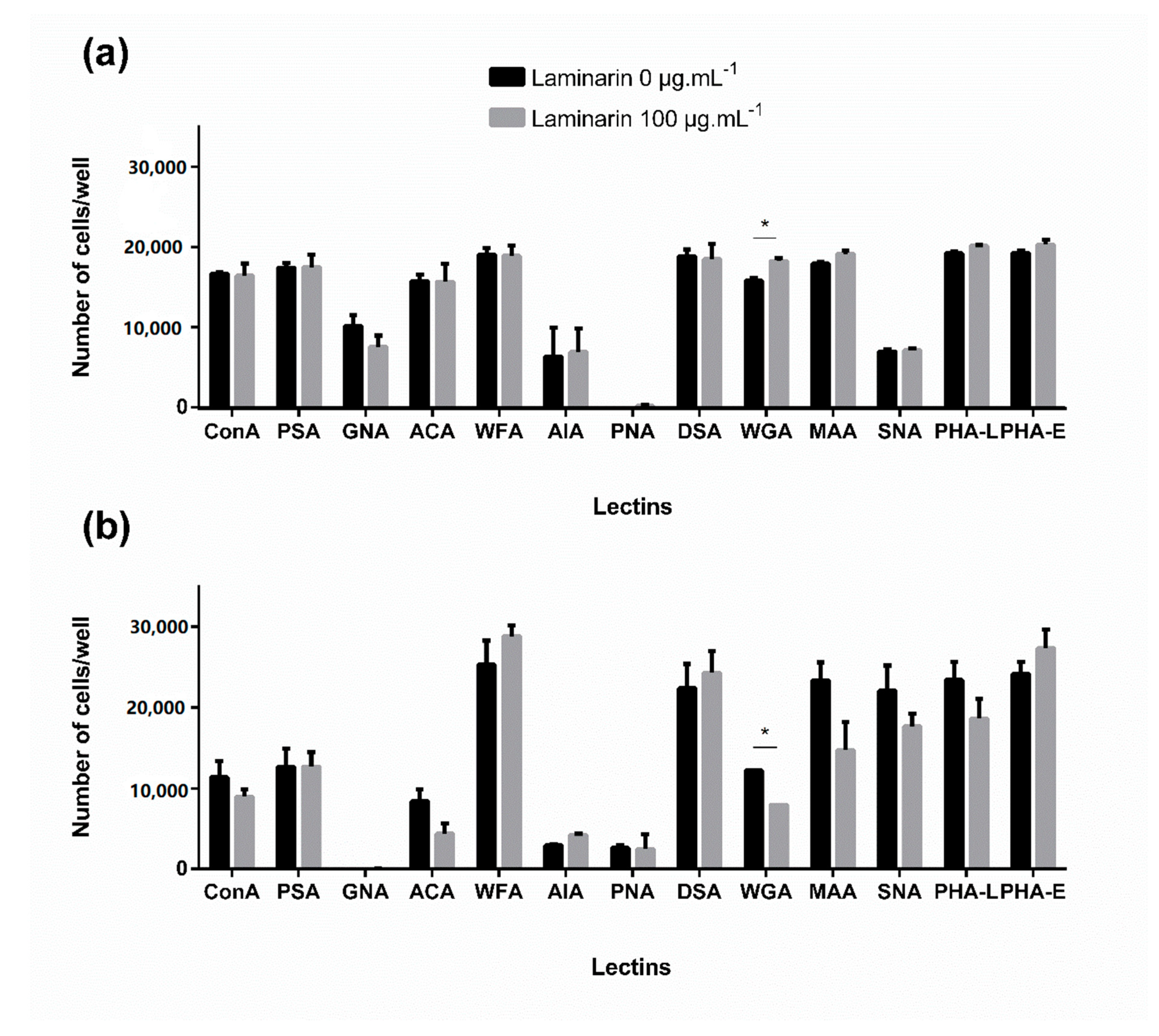
3.4. Effect of Laminarin on Cells—Glycan Interaction Study
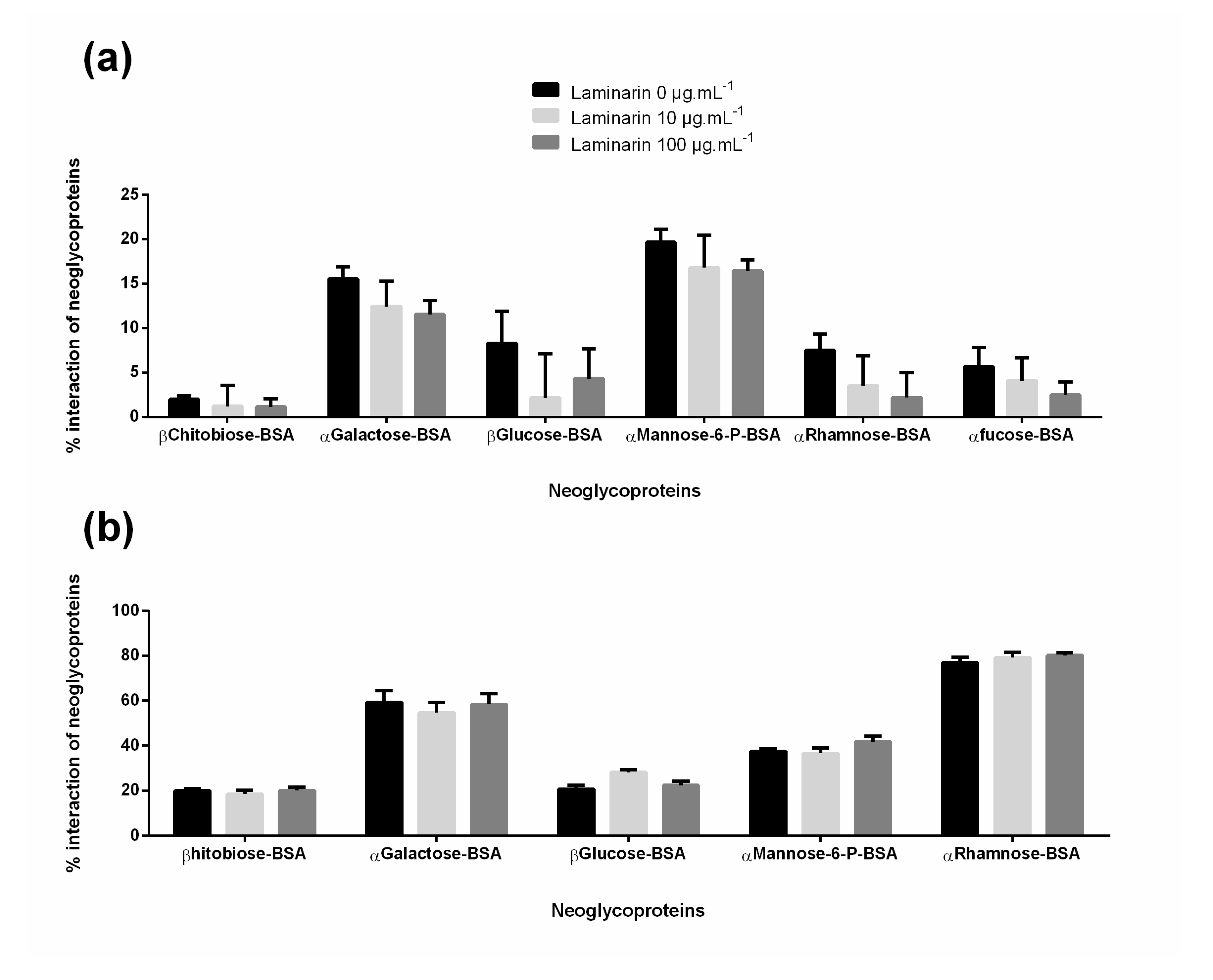
3.5. Effect of Laminarin with Cells—Carbohydrates Receptors Study
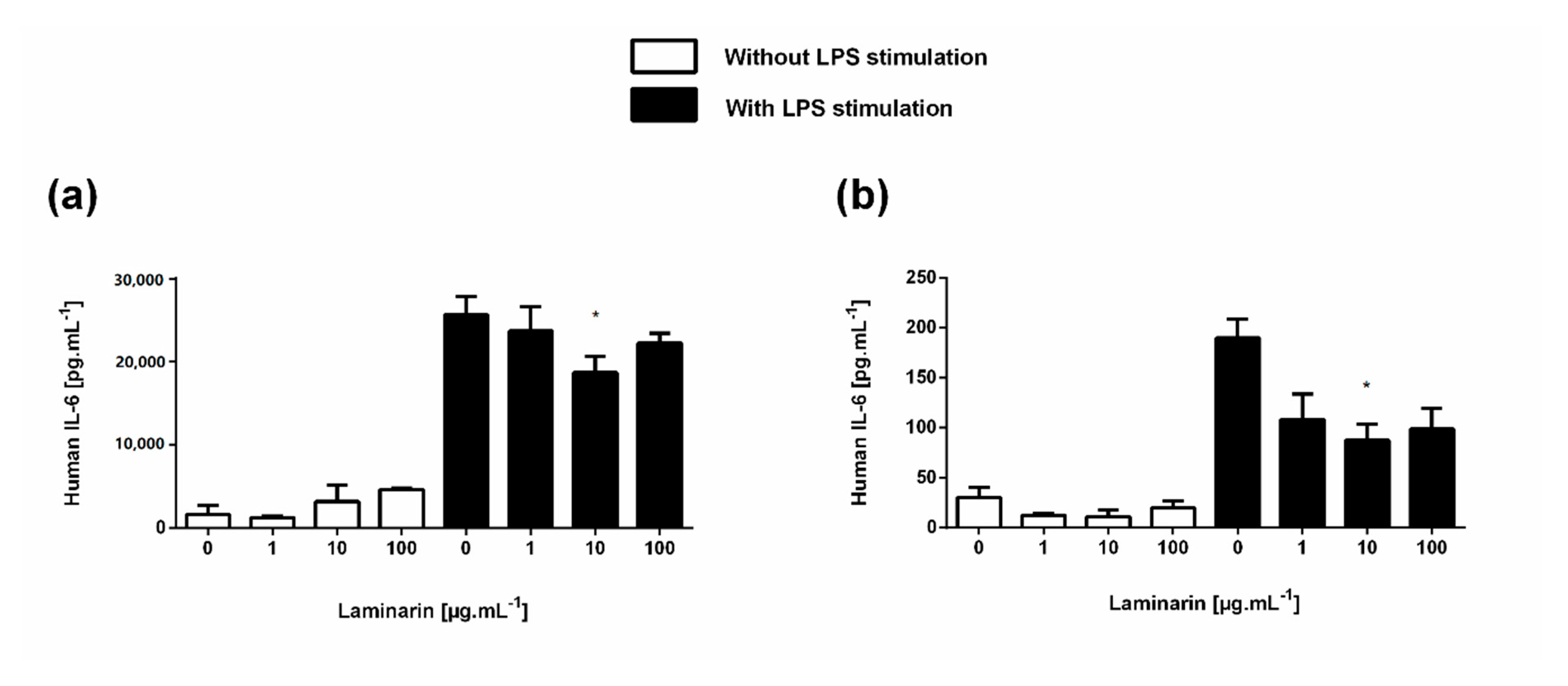
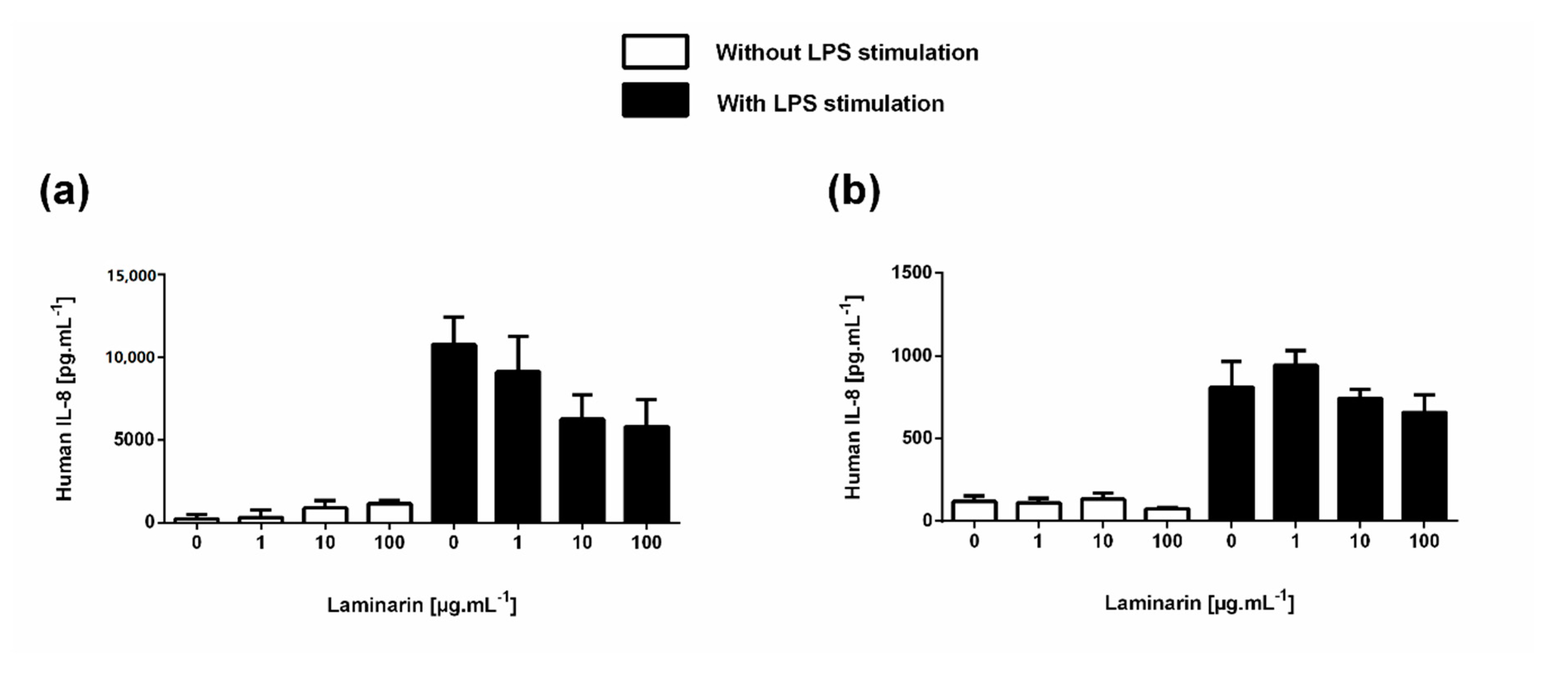
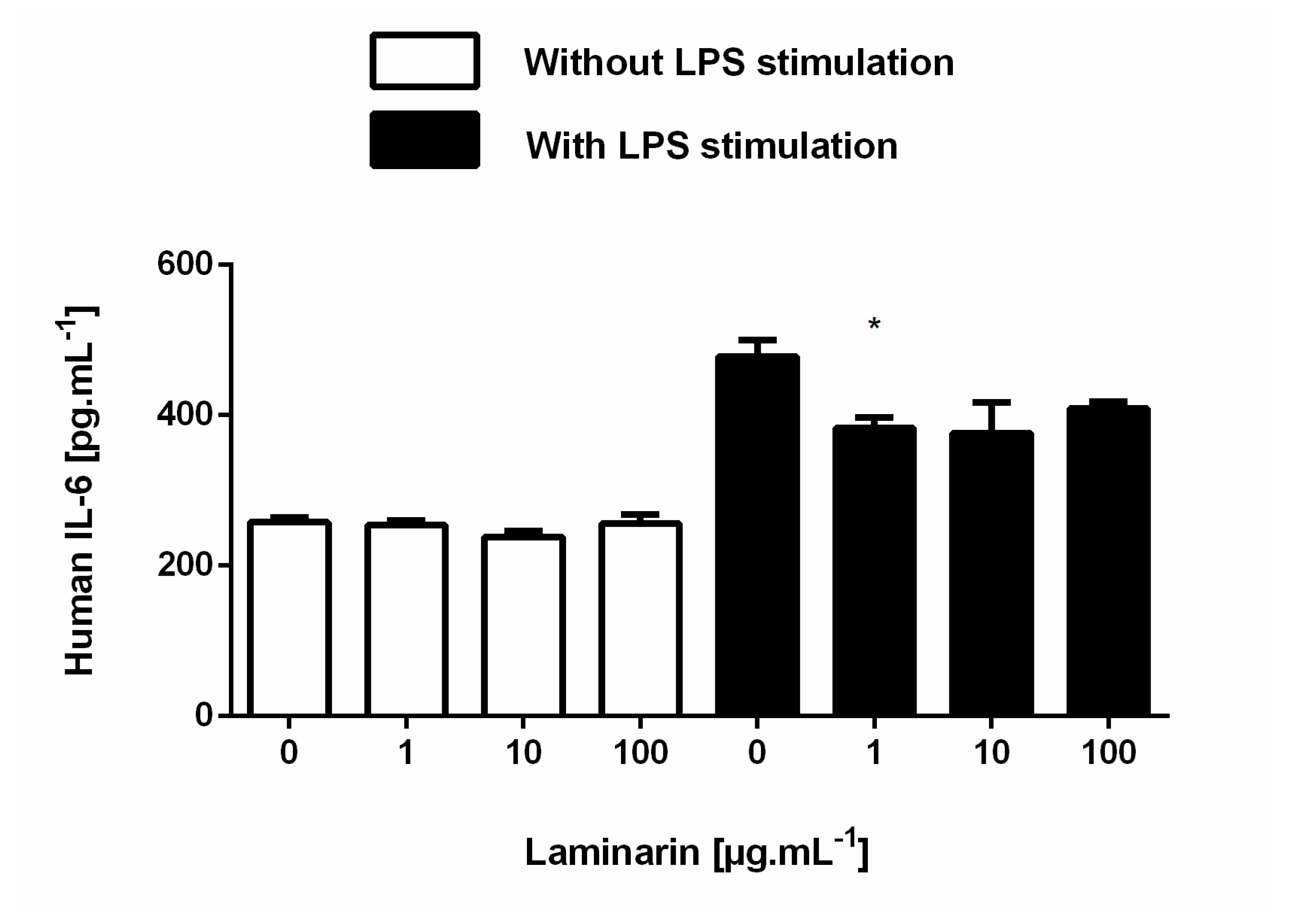
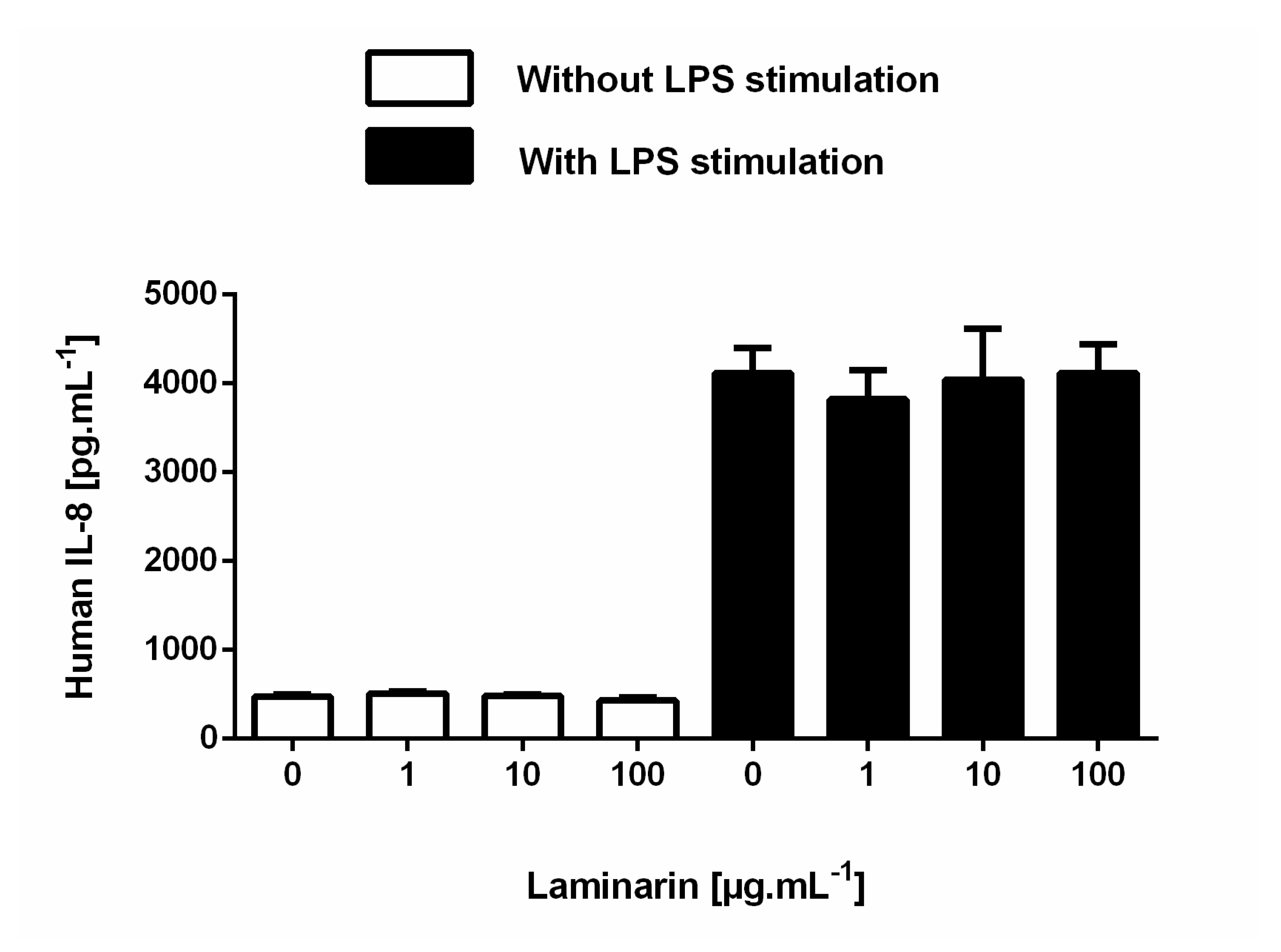
3.6. Laminarin Reduced IL-6 Secretion in Inflammatory Skin Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A






References
- Ruocco, N.; Costantini, S.; Guariniello, S.; Costantini, M. Polysaccharides from the marine environment with pharmacological, cosmeceutical and nutraceutical potential. Molecules 2016, 21, 551. [Google Scholar] [CrossRef]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Ferrières, V.; Legentil, L.; Sylla, B.; Descroix, K.; Nugier-Chauvin, C.; Daniellou, R. Contribution of Biocatalysis to the Synthesis of β-(1, 3)-Glucans. Biol. Chem. Beta Glucan 2013, 2, 102. [Google Scholar]
- Zhu, F.; Du, B.; Xu, B. A critical review on production and industrial applications of beta-glucans. Food Hydrocoll. 2016, 52, 275–288. [Google Scholar] [CrossRef]
- Zeković, D.B.; Kwiatkowski, S.; Vrvić, M.M.; Jakovljević, D.; Moran, C.A. Natural and modified (1→3)-β-D-glucans in health promotion and disease alleviation. Crit. Rev. Biotechnol. 2005, 25, 205–230. [Google Scholar] [CrossRef] [PubMed]
- Sylla, B.; Legentil, L.; Saraswat-Ohri, S.; Vashishta, A.; Daniellou, R.; Wang, H.-W.; Vetvicka, V.; Ferrières, V. Oligo-β-(1→3)-glucans: Impact of thio-bridges on immunostimulating activities and the development of cancer stem cells. J. Med. Chem. 2014, 57, 8280–8292. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Cheung, P.C.K. Immunopotentiation and anti-tumor activity of carboxymethylated-sulfated β-(1→3)-d-glucan from Poria cocos. Int. Immunopharmacol. 2010, 10, 398–405. [Google Scholar] [CrossRef]
- Toklu, H.Z.; Sener, G.; Jahovic, N.; Uslu, B.; Arbak, S.; Yeğen, B.C. Beta-glucan protects against burn-induced oxidative organ damage in rats. Int. Immunopharmacol. 2006, 6, 156–169. [Google Scholar] [CrossRef]
- Kim, H.-D.; Cho, H.-R.; Moon, S.-B.; Shin, H.-D.; Yang, K.-J.; Park, B.-R.; Jang, H.-J.; Kim, L.-S.; Lee, H.-S.; Ku, S.-K. Effects of beta-glucan from Aureobasidium pullulans on acute inflammation in mice. Arch. Pharm. Res. 2007, 30, 323–328. [Google Scholar] [CrossRef]
- Vetvicka, V.; Thornton, B.P.; Ross, G.D. Soluble beta-glucan polysaccharide binding to the lectin site of neutrophil or natural killer cell complement receptor type 3 (CD11b/CD18) generates a primed state of the receptor capable of mediating cytotoxicity of iC3b-opsonized target cells. J. Clin. Investig. 1996, 98, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.D.; Taylor, P.R.; Reid, D.M.; Willment, J.A.; Williams, D.L.; Martinez-Pomares, L.; Wong, S.Y.C.; Gordon, S. Dectin-1 is a major beta-glucan receptor on macrophages. J. Exp. Med. 2002, 196, 407–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kougias, P.; Wei, D.; Rice, P.J.; Ensley, H.E.; Kalbfleisch, J.; Williams, D.L.; Browder, I.W. Normal human fibroblasts express pattern recognition receptors for fungal (1→3)-beta-D-glucans. Infect. Immun. 2001, 69, 3933–3938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Berg, L.M.; Zijlstra-Willems, E.M.; Richters, C.D.; Ulrich, M.M.W.; Geijtenbeek, T.B.H. Dectin-1 activation induces proliferation and migration of human keratinocytes enhancing wound re-epithelialization. Cell. Immunol. 2014, 289, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Bian, Z.; Xu, B. Skin Health Promotion Effects of Natural Beta-Glucan Derived from Cereals and Microorganisms: A Review. Phytother. Res. 2014, 28, 159–166. [Google Scholar] [CrossRef]
- Gautier, S.; Xhauflaire-Uhoda, E.; Gonry, P.; Piérard, G.E. Chitin–glucan, a natural cell scaffold for skin moisturization and rejuvenation. Int. J. Cosmet. Sci. 2008, 30, 459–469. [Google Scholar] [CrossRef]
- Wang, Y.; Viennet, C.; Jeudy, A.; Fanian, F.; He, L.; Humbert, P. Assessment of the efficacy of a new complex antisensitive skin cream. J. Cosmet. Derm. 2018, 17, 1101–1107. [Google Scholar] [CrossRef]
- Choi, J.-I.; Kim, H.-J.; Kim, J.-H.; Lee, J.-W. Enhanced Biological Activities of Laminarin Degraded by Gamma-Ray Irradiation. J. Food Biochem. 2012, 36, 465–469. [Google Scholar] [CrossRef]
- Yao, X.; Jiang, H.; Liang, S.; Shen, X.; Gao, Q.; Xu, Y.N.; Kim, N.-H. Laminarin enhances the quality of aged pig oocytes by reducing oxidative stress. J. Reprod. Dev. 2018, 64, 489–494. [Google Scholar] [CrossRef]
- Smith, A.J.; Graves, B.; Child, R.; Rice, P.J.; Ma, Z.; Lowman, D.W.; Ensley, H.E.; Ryter, K.T.; Evans, J.T.; Williams, D.L. Immunoregulatory Activity of the Natural Product Laminarin Varies Widely as a Result of Its Physical Properties. J. Immunol. 2018, 200, 788–799. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Bonfim-Mendonça, P.D.S.; Capoci, I.R.G.; Tobaldini-Valerio, F.K.; Negri, M.; Svidzinski, T.I.E. Overview of β-Glucans from Laminaria spp.: Immunomodulation Properties and Applications on Biologic Models. Int. J. Mol. Sci. 2017, 18, 1629. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Xu, L.; Zhang, W.; Cai, Y.; Jang, B.; Oh, J.; Jin, J.-O. Laminarin promotes anti-cancer immunity by the maturation of dendritic cells. Oncotarget 2017, 8, 38554–38567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-H.; Kim, Y.-W.; Kim, H.B.; Lee, B.J.; Lee, D.S. Anti-apoptotic activity of laminarin polysaccharides and their enzymatically hydrolyzed oligosaccharides from Laminaria japonica. Biotechnol. Lett. 2006, 28, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Mouson, A.; Delzenne, N.M. Dietary supplementation with laminarin, a fermentable marine beta (1-3) glucan, protects against hepatotoxicity induced by LPS in rat by modulating immune response in the hepatic tissue. Int. Immunopharmacol. 2007, 7, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, C.J.; O’Doherty, J.V.; Callanan, J.J.; Doyle, D.; Thornton, K.; Sweeney, T. The effect of algal polysaccharides laminarin and fucoidan on colonic pathology, cytokine gene expression and Enterobacteriaceae in a dextran sodium sulfate-challenged porcine model. J. Nutr. Sci. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.-F.; Ji, Y.-B.; Meng, D.-Y. Sulfated modification and anti-tumor activity of laminarin. Exp. Ther. Med. 2013, 6, 1259–1264. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Landemarre, L.; Cancellieri, P.; Duverger, E. Cell surface lectin array: Parameters affecting cell glycan signature. Glycoconj. J. 2013, 30, 195–203. [Google Scholar] [CrossRef]
- Morello, E.; Mallet, A.; Konto-Ghiorghi, Y.; Chaze, T.; Mistou, M.-Y.; Oliva, G.; Oliveira, L.; Di Guilmi, A.-M.; Trieu-Cuot, P.; Dramsi, S. Evidence for the sialylation of PilA, the PI-2a pilus-associated adhesin of Streptococcus agalactiae strain NEM316. PLoS ONE 2015, 10, e0138103. [Google Scholar] [CrossRef] [Green Version]
- Hassani, Z.; Saleh, A.; Turpault, S.; Khiati, S.; Morelle, W.; Vignon, J.; Hugnot, J.-P.; Uro-Coste, E.; Legrand, P.; Delaforge, M. Phostine PST3. 1a targets MGAT5 and inhibits glioblastoma-initiating cell invasiveness and proliferation. Mol. Cancer Res. 2017, 15, 1376–1387. [Google Scholar] [CrossRef] [Green Version]
- Cutolo, G.; Reise, F.; Schuler, M.; Nehmé, R.; Despras, G.; Brekalo, J.; Morin, P.; Renard, P.-Y.; Lindhorst, T.K.; Tatibouët, A. Bifunctional mannoside–glucosinolate glycoconjugates as enzymatically triggered isothiocyanates and FimH ligands. Org. Biomol. Chem. 2018, 16, 4900–4913. [Google Scholar] [CrossRef] [PubMed]
- Fagot, D.; Pham, D.M.; Laboureau, J.; Planel, E.; Guerin, L.; Nègre, C.; Donovan, M.; Bernard, B.A. Crocin, a natural molecule with potentially beneficial effects against skin ageing. Int. J. Cosmet. Sci. 2018, 40, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.M.; Lisowska, E.; Duk, M.; Yang, Z. Lectins as tools in glycoconjugate research. Glycoconj. J. 2009, 26, 899–913. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Duan, E. Fighting against Skin Aging: The Way from Bench to Bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cezar, T.L.C.; Martinez, R.M.; Da Rocha, C.; Melo, C.P.B.; Vale, D.L.; Borghi, S.M.; Fattori, V.; Vignoli, J.A.; Camilios-Neto, D.; Baracat, M.M.; et al. Treatment with maresin 1, a docosahexaenoic acid-derived pro-resolution lipid, protects skin from inflammation and oxidative stress caused by UVB irradiation. Sci. Rep. 2019, 9, 3062. [Google Scholar] [CrossRef]
- Puizina-Ivić, N. Skin aging. Acta Dermatovenerol. Alp. Panon. Et Adriat. 2008, 17, 47–54. [Google Scholar]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Woo, S.-Y.; Cho, K.-A.; Cho, M.-S.; Lee, K.H. PD-L1 produced by HaCaT cells under polyinosinic-polycytidylic acid stimulation inhibits melanin production by B16F10 cells. PLoS ONE 2020, 15, e0233448. [Google Scholar] [CrossRef]
- Lehmann, B. HaCaT cell line as a model system for vitamin D3 metabolism in human skin. J. Investig. Dermatol. 1997, 108, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Diez, M.; Santamaría, G.; Ortega, A.D.; Cuezva, J.M. Biogenesis and dynamics of mitochondria during the cell cycle: Significance of 3’UTRs. PLoS ONE 2006, 1, e107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garn, H.; Krause, H.; Enzmann, V.; Drössler, K. An improved MTT assay using the electron-coupling agent menadione. J. Immunol. Methods 1994, 168, 253–256. [Google Scholar] [CrossRef]
- Thom, S.M.; Horobin, R.W.; Seidler, E.; Barer, M.R. Factors affecting the selection and use of tetrazolium salts as cytochemical indicators of microbial viability and activity. J. Appl. Bacteriol. 1993, 74, 433–443. [Google Scholar] [CrossRef]
- Shoemaker, M.; Cohen, I.; Campbell, M. Reduction of MTT by aqueous herbal extracts in the absence of cells. J. Ethnopharmacol. 2004, 93, 381–384. [Google Scholar] [CrossRef]
- Peng, L.; Wang, B.; Ren, P. Reduction of MTT by flavonoids in the absence of cells. Colloids Surf. B Biointerfaces 2005, 45, 108–111. [Google Scholar] [CrossRef]
- Morgan, D.M.L. Tetrazolium (MTT) Assay for Cellular Viability and Activity. In Polyamine Protocols; Morgan, D.M.L., Ed.; Humana Press: Totowa, NJ, USA, 1998; pp. 179–184, Methods in Molecular BiologyTM; ISBN 978-1-59259-565-5. [Google Scholar]
- Park, H.-K.; Kim, I.-H.; Kim, J.; Nam, T.-J. Induction of apoptosis and the regulation of ErbB signaling by laminarin in HT-29 human colon cancer cells. Int. J. Mol. Med. 2013, 32, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Uitto, J. Collagen polymorphism: Isolation and partial characterization of α1(I)-trimer molecules in normal human skin. Arch. Biochem. Biophys. 1979, 192, 371–379. [Google Scholar] [CrossRef]
- Stern, R.; Maibach, H.I. Hyaluronan in skin: Aspects of aging and its pharmacologic modulation. Clin. Dermatol. 2008, 26, 106–122. [Google Scholar] [CrossRef]
- Meyer, L.J.M.; Stern, R. Age-Dependent Changes of Hyaluronan in Human Skin. J. Investig. Dermatol. 1994, 102, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Voigt, J.; Driver, V.R. Hyaluronic acid derivatives and their healing effect on burns, epithelial surgical wounds, and chronic wounds: A systematic review and meta-analysis of randomized controlled trials. Wound Repair Regen 2012, 20, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Kwansa, A.L.; De Vita, R.; Freeman, J.W. Tensile mechanical properties of collagen type I and its enzymatic crosslinks. Biophys. Chem. 2016, 214, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, A.; Pereira, J.M.; Rioux, L.-E.; Turgeon, S.L.; Beaulieu, M.; Moulin, V.J. Role of seaweed laminaran from Saccharina longicruris on matrix deposition during dermal tissue-engineered production. Int. J. Biol. Macromol. 2015, 75, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Read, S.M.; Currie, G.; Bacic, A. Analysis of the structural heterogeneity of laminarin by electrospray-ionisation-mass spectrometry. Carbohydr. Res. 1996, 281, 187–201. [Google Scholar] [CrossRef]
- Rioux, L.-E.; Turgeon, S.L.; Beaulieu, M. Structural characterization of laminaran and galactofucan extracted from the brown seaweed Saccharina longicruris. Phytochemistry 2010, 71, 1586–1595. [Google Scholar] [CrossRef]
- Moroney, N.C.; O’Grady, M.N.; Lordan, S.; Stanton, C.; Kerry, J.P. Seaweed polysaccharides (laminarin and fucoidan) as functional ingredients in pork meat: An evaluation of anti-oxidative potential, thermal stability and bioaccessibility. Mar. Drugs 2015, 13, 2447–2464. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Liang, B.; Li, M.; Jin, M. Influence of laminarin polysaccahrides on oxidative damage. Int. J. Biol. Macromol. 2011, 48, 63–66. [Google Scholar] [CrossRef]
- Wu, S.-J. Preparation and antioxidant activity of the oligosaccharides derived from Laminaria japonica. Carbohydr. Polym. 2014, 106, 22–24. [Google Scholar] [CrossRef]
- Ramachandran, S.; Rajendra Prasad, N.; Karthikeyan, S. Sesamol inhibits UVB-induced ROS generation and subsequent oxidative damage in cultured human skin dermal fibroblasts. Arch. Dermatol. Res. 2010, 302, 733–744. [Google Scholar] [CrossRef]
- Karran, P.; Brem, R. Protein oxidation, UVA and human DNA repair. DNA Repair 2016, 44, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, R.; Kanimozhi, G.; Prasad, N.R.; Agilan, B.; Ganesan, M.; Srithar, G. Alpha pinene modulates UVA-induced oxidative stress, DNA damage and apoptosis in human skin epidermal keratinocytes. Life Sci. 2018, 212, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Custódio, C.A.; Reis, R.L.; Mano, J.F. Photo-Cross-Linked Laminarin-Based Hydrogels for Biomedical Applications. Biomacromolecules 2016, 17, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.Q.; Ishai-Michaeli, R.; Peretz, T.; Vlodavsky, I. Laminarin sulfate mimics the effects of heparin on smooth muscle cell proliferation and basic fibroblast growth factor-receptor binding and mitogenic activity. J. Cell. Physiol. 1995, 164, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.; Larm, O.; Larsson, K.; Andersson, L.O.; Holmer, E.; Söderström, G. Studies on the blood-anticoagulant activity of sulphated polysaccharides with different uronic acid content. Carbohydr. Polym. 1982, 2, 115–121. [Google Scholar] [CrossRef]
- Tsoni, S.V.; Brown, G.D. β-Glucans and Dectin-1. Ann. N. Y. Acad. Sci. 2008, 1143, 45–60. [Google Scholar] [CrossRef]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; Toscano, M.A. Turning′sweet′on immunity: Galectin–glycan interactions in immune tolerance and inflammation. Nat. Rev. Immunol. 2009, 9, 338–352. [Google Scholar] [CrossRef]
- Kobayashi, M.; Yoshiki, R.; Sakabe, J.; Kabashima, K.; Nakamura, M.; Tokura, Y. Expression of toll-like receptor 2, NOD2 and dectin-1 and stimulatory effects of their ligands and histamine in normal human keratinocytes. Br. J. Dermatol. 2009, 160, 297–304. [Google Scholar] [CrossRef]
- Schafer, Z.T.; Brugge, J.S. IL-6 involvement in epithelial cancers. J. Clin. Investig. 2007, 117, 3660–3663. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, T. INTERLEUKIN-6: From Basic Science to Medicine—40 Years in Immunology. Annu. Rev. Immunol. 2005, 23, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Wallenius, V.; Wallenius, K.; Ahrén, B.; Rudling, M.; Carlsten, H.; Dickson, S.L.; Ohlsson, C.; Jansson, J.-O. Interleukin-6-deficient mice develop mature-onset obesity. Nat. Med. 2002, 8, 75–79. [Google Scholar] [CrossRef]
- Chiu, J.J.; Sgagias, M.K.; Cowan, K.H. Interleukin 6 acts as a paracrine growth factor in human mammary carcinoma cell lines. Clin. Cancer Res. 1996, 2, 215–221. [Google Scholar] [PubMed]
- Conze, D.; Weiss, L.; Regen, P.S.; Bhushan, A.; Weaver, D.; Johnson, P.; Rincón, M. Autocrine production of interleukin 6 causes multidrug resistance in breast cancer cells. Cancer Res. 2001, 61, 8851–8858. [Google Scholar] [PubMed]
- Taniguchi, K.; Wu, L.; Grivennikov, S.I.; De Jong, P.R.; Lian, I.; Yu, F.-X.; Wang, K.; Ho, S.B.; Boland, B.S.; Chang, J.T.; et al. A gp130-Src-YAP module links inflammation to epithelial regeneration. Nature 2015, 519, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arranz-Valsero, I.; Soriano-Romaní, L.; García-Posadas, L.; López-García, A.; Diebold, Y. IL-6 as a corneal wound healing mediator in an in vitro scratch assay. Exp. Eye Res. 2014, 125, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.Z.; Stevenson, A.W.; Prêle, C.M.; Fear, M.W.; Wood, F.M. The Role of IL-6 in Skin Fibrosis and Cutaneous Wound Healing. Biomedicines 2020, 8, 101. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Short Name | Common Name | Glycan Structures Specificities |
|---|---|---|
| ConA | Concanavalin Agglutinin | Manα6(Manα3)Man |
| PSA | Pisum Sativum Agglutinin | Manα6(Manα3)Man |
| GNA | Galanthus Nivalis Agglutinin | Manα6(Manα3)Man |
| ACA | Amaranthus Caudatus Agglutinin | Galβ3GalNAc |
| WFA | Wisteria Floribunda Agglutinin | GalNAcα6Gal > GalNAcα3GalNAc > GalNAc |
| PNA | Peanut Agglutinin | Galβ3GalNAc |
| AIA | Autocarpus Intergrifolia Agglutinin | Galβ3GalNAc |
| DSA | Datura Stramonium Agglutinin | GlcNAcβ4GlcNAc |
| WGA | Wheat Germ Agglutinin | GlcNAcGlcNAcβ4 |
| MAA | Maackia Amurensis Agglutinin | Neu5Ac3Gal4GalNAc |
| SNA | Sambucus Nigra Agglutinin | Neu5Ac6Gal/GalNAc |
| PHA-L | Phaseolus Vulgaris Agglutinin | Complex glycans |
| PHA-E | Phaseolus Vulgaris Agglutinin | Complex glycans |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozanne, H.; Toumi, H.; Roubinet, B.; Landemarre, L.; Lespessailles, E.; Daniellou, R.; Cesaro, A. Laminarin Effects, a β-(1,3)-Glucan, on Skin Cell Inflammation and Oxidation. Cosmetics 2020, 7, 66. https://doi.org/10.3390/cosmetics7030066
Ozanne H, Toumi H, Roubinet B, Landemarre L, Lespessailles E, Daniellou R, Cesaro A. Laminarin Effects, a β-(1,3)-Glucan, on Skin Cell Inflammation and Oxidation. Cosmetics. 2020; 7(3):66. https://doi.org/10.3390/cosmetics7030066
Chicago/Turabian StyleOzanne, Hélène, Hechmi Toumi, Benoît Roubinet, Ludovic Landemarre, Eric Lespessailles, Richard Daniellou, and Annabelle Cesaro. 2020. "Laminarin Effects, a β-(1,3)-Glucan, on Skin Cell Inflammation and Oxidation" Cosmetics 7, no. 3: 66. https://doi.org/10.3390/cosmetics7030066
APA StyleOzanne, H., Toumi, H., Roubinet, B., Landemarre, L., Lespessailles, E., Daniellou, R., & Cesaro, A. (2020). Laminarin Effects, a β-(1,3)-Glucan, on Skin Cell Inflammation and Oxidation. Cosmetics, 7(3), 66. https://doi.org/10.3390/cosmetics7030066