Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Paederia foetida L. Extract
2.2. Cell Culture
2.3. Measurement of Cell Viability
2.4. Analysis of Cell Melanin Content
2.5. Analysis of Intracellular Tyrosinase Activity
2.6. Preparation of Western Blot Analysis
2.7. Preparation of Primary Skin Irritation Test
2.8. Significance Test through Statistical Analysis
3. Results and Discussion
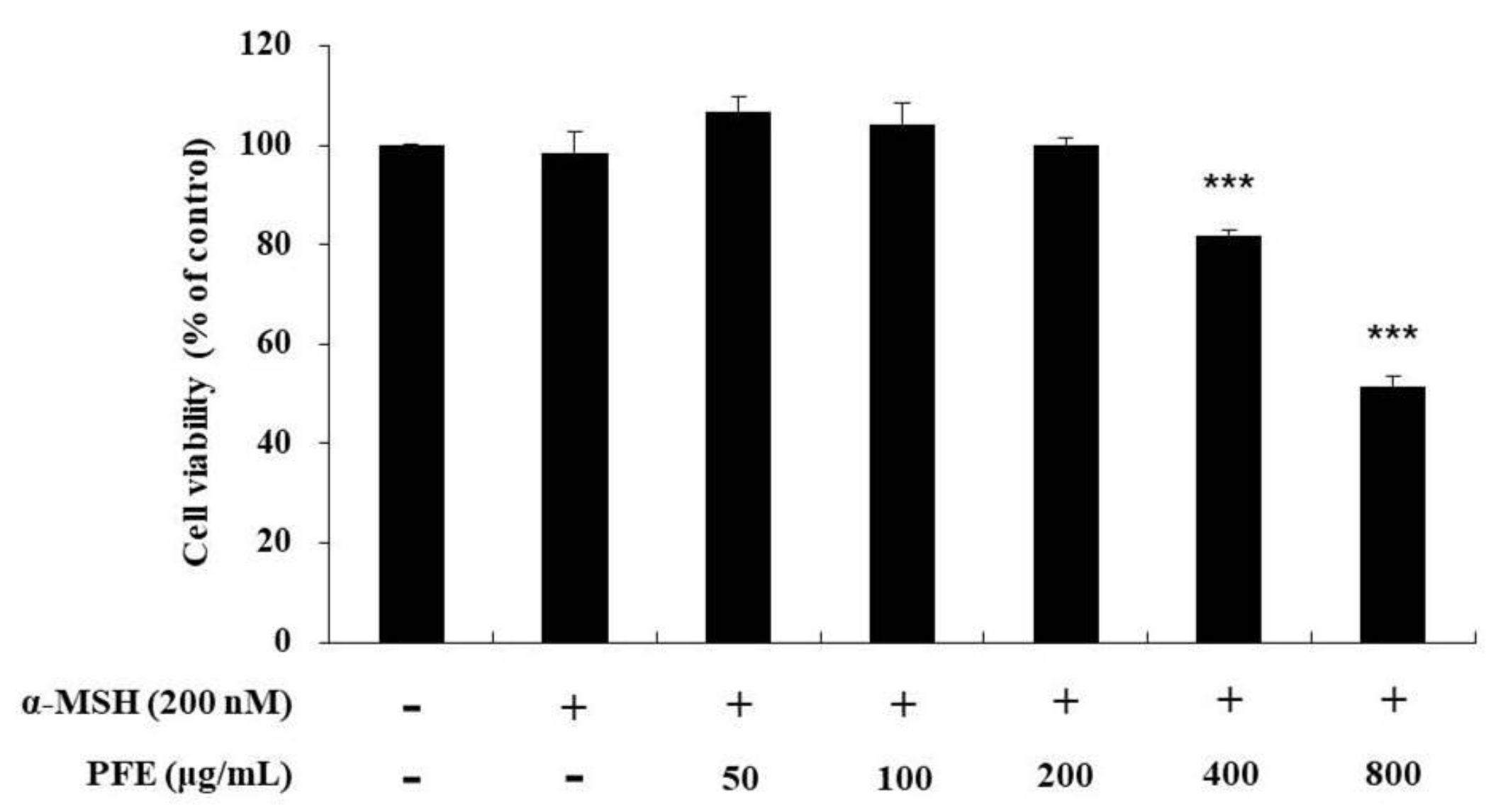
3.1. Effect of PFE on Cell Viabilty
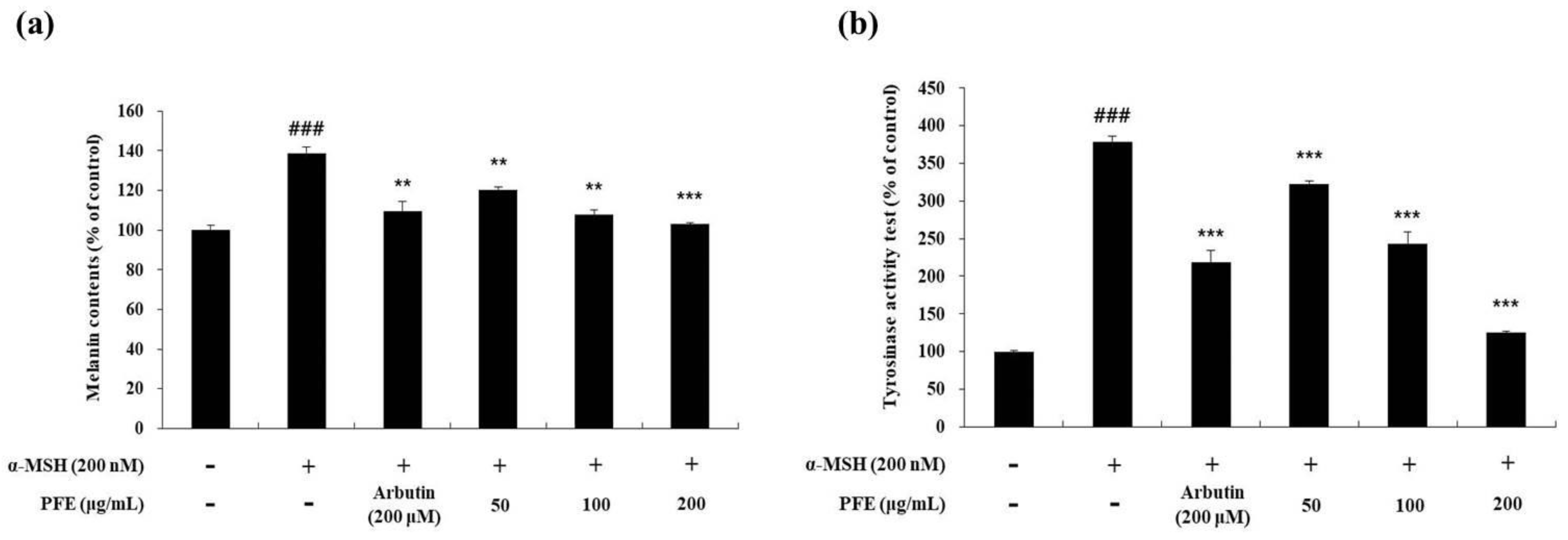
3.2. Melanin Content and Intracellular Tyrosinase Inhibitory Activity by PFE Treatment
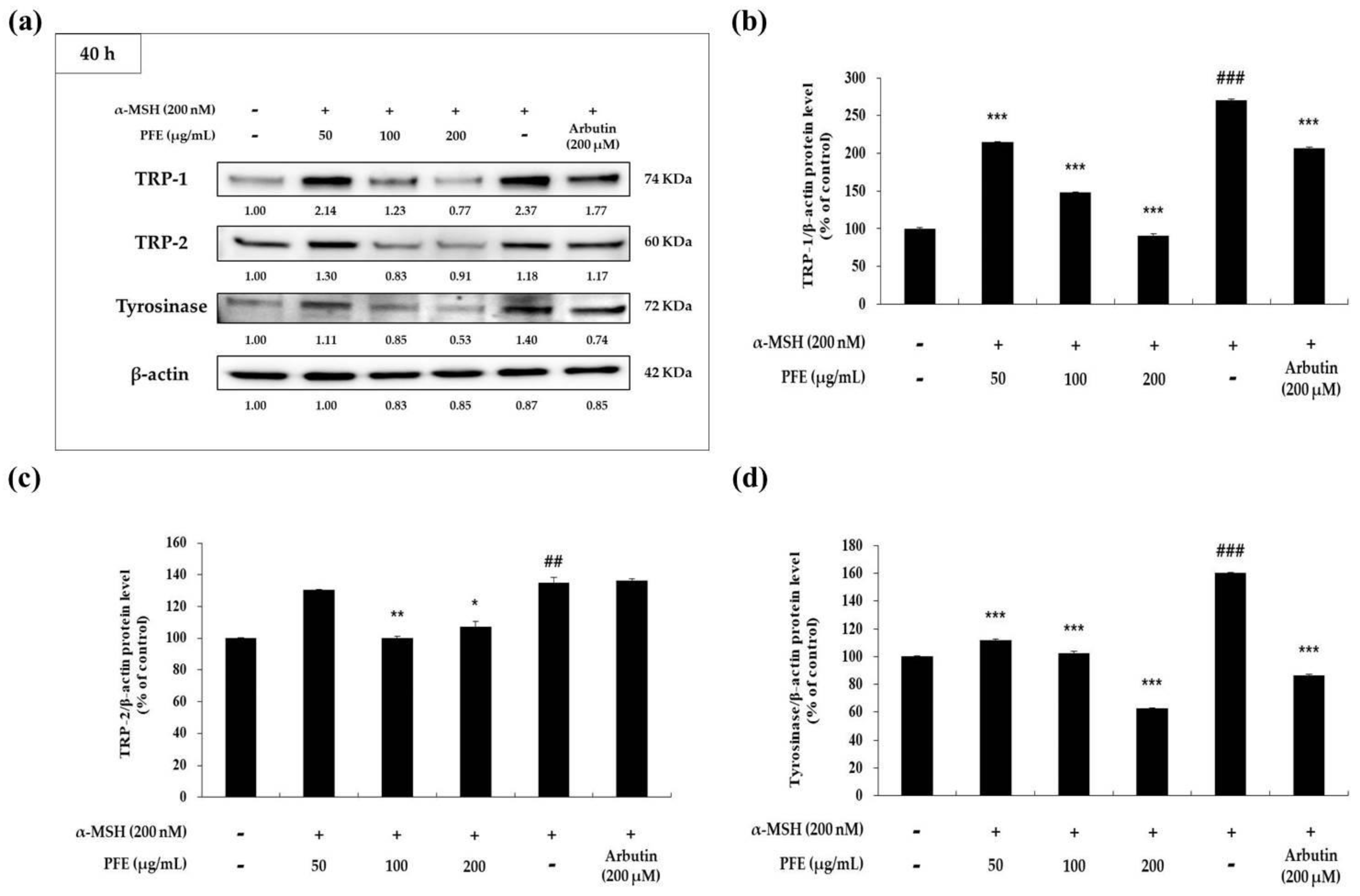
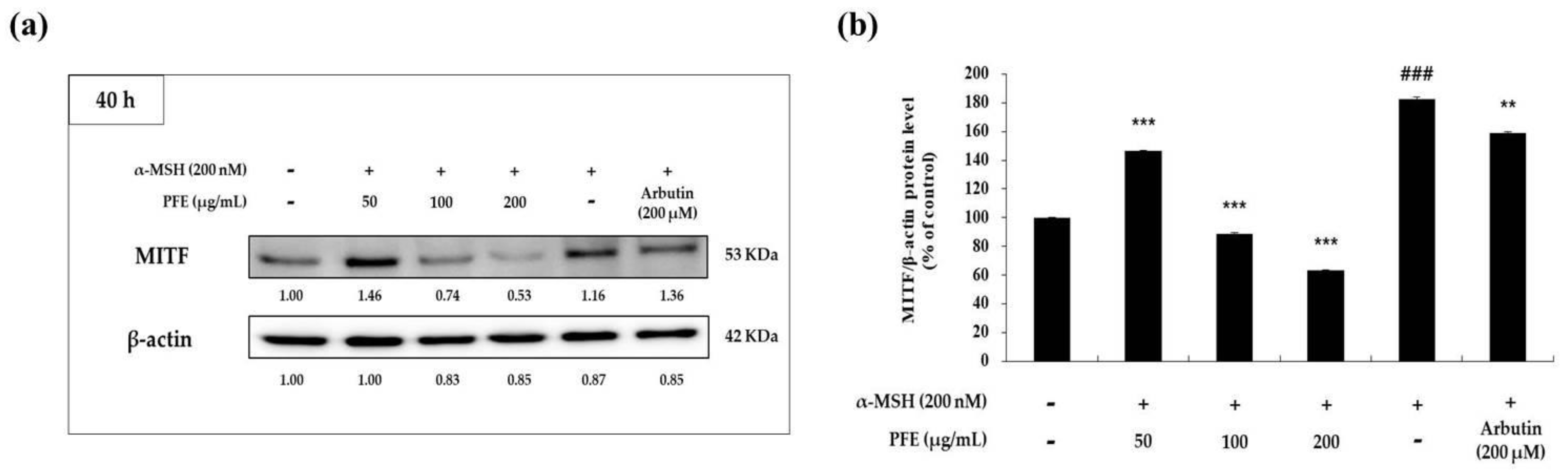
3.3. Downregulation of PFE on Melanogenic Enzymes and MITF Expression
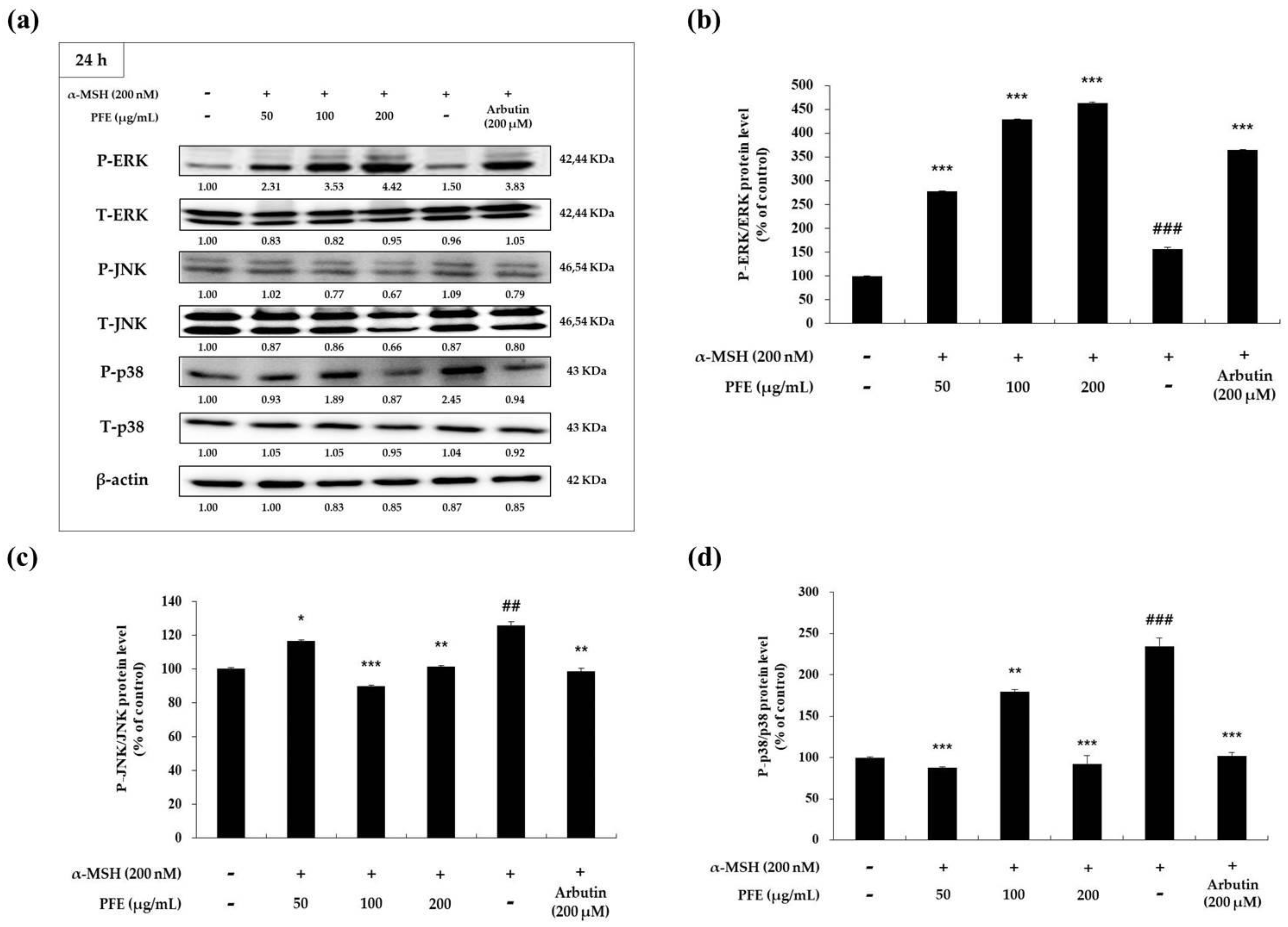
3.4. Effect of PFE on MAPK Phosphorylation
3.5. Primary Skin Irritation Test of PFE
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hearing, V.J. Biogenesis of pigment granules: A sensitive way to regulate melanocyte function. J. Dermatol. Sci. 2005, 37, 3–14. [Google Scholar] [CrossRef]
- Bonaventure, J.; Domingues, M.J.; Larue, L. Cellular and molecular mechanisms controlling the migration of melanocytes and melanoma cells. Pigment Cell Melanoma Res. 2013, 26, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, T.; Yamaguchi, Y.; Batzer, J.; Coelho, S.G.; Zmudzka, B.Z.; Miller, S.A.; Wolber, R.; Beer, J.Z.; Hearing, V.J. Mechanisms of skin tanning in different racial/ethnic groups in response to ultraviolet radiation. J. Investig. Dermatol. 2005, 124, 1326–1332. [Google Scholar] [CrossRef] [Green Version]
- Tabassum, N.; Hamdani, M. Plants used to treat skin diseases. Pharmacogn. Rev. 2014, 8, 52–60. [Google Scholar] [CrossRef] [Green Version]
- D’Ischia, M.; Wakamatsu, K.; Cicoira, F.; Mauro, E.D.; Garcia-Borron, J.C.; Commo, S.; Galvan, I.; Ghanem, G.; Kenzo, K.; Meredith, P.; et al. Melanins and melanogenesis: From pigment cells to human health and technological applications. Pigment Cell Melanoma Res. 2015, 28, 520–544. [Google Scholar] [CrossRef] [Green Version]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Otręba, M.; Buszman, E.; Miliński, M.; Wrześniok, D. Hypomelanoses transmitted from generation to generation. Postepy Hig Med Dosw 2014, 68, 1081–1090. [Google Scholar] [CrossRef]
- Otręba, M.; Miliński, M.; Buszman, E.; Wrześniok, D.; Beberok, A. Hereditary hypomelanocytoses: The role of PAX3, SOX10, MITF, SNAI2, KIT, EDN3 and EDNRB genes. Postepy Hig. Med. Dosw. 2013, 67, 1109–1118. [Google Scholar] [CrossRef]
- Miyamura, Y.I.; Coelho, S.G.; Wolber, R.; Miller, S.A.; Wakamatsu, K.; Zmudzka, B.Z.; Ito, S.; Smuda, C.; Passeron, T.; Choi, W. Regulation of human skin pigmentation and responses to ultraviolet radiation. Pigment Cell Res. 2007, 20, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Camera, E.; Picardo, M. Chemical and instrumental approaches to treat hyperpigmentation. Pigment Cell Res. 2003, 16, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Sugumaran, M. Reactivities of quinone methides versus o-quinones in catecholamine metabolism and eumelanin biosynthesis. Int. J. Mol. Sci. 2016, 17, 1576. [Google Scholar] [CrossRef] [Green Version]
- Hearing, V.J.; Jimenez, M. Mammalian tyrosinase—the critical regulatory control point in melanocyte pigmentation. Int. J. Biochem. 1987, 19, 1141–1147. [Google Scholar] [CrossRef]
- Jiménez-Cervantes, C.; Solano, F.; Kobayashi, T.; Urabe, K.; Hearing, V.J.; Lozano, J.A.; Garcia-Borrón, J.C. A new enzymatic function in the melanogenic pathway. The 5,6-dihydroxyindole-2-carboxylic acid oxidase activity of tyrosinase-related protein-1 (TRP1). J. Biol. Chem. 1994, 269, 17993–18000. [Google Scholar] [CrossRef]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V.J. A second tyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Xie, H.F.; Tang, Y.; Lin, S.Q.; Li, J.M.; Sun, S.M.; Hu, X.L.; Huang, Y.X.; Shi, W.; Jian, D.G. Protein-coupled estrogen receptor enhances melanogenesis via cAMP-protein kinase (PKA) by upregulating microphthalmia-related transcription factor-tyrosinase in melanoma. J. Steroid Biochem. Mol. Biol. 2017, 165, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Bentley, N.J.; Eisen, T.; Goding, C.R. Melanocyte-specific expression of the human tyrosinase promoter: Activation by the microphthalmia gene product and role of the initiator. Mol. Cell. Biol. 1994, 14, 7996–8006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachtenheim, J.; Borovansky, J. “Transcription physiology” of pigment formation in melanocytes: Central role of MITF. Exp. Dermatol. 2010, 19, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Jin, S.H.; Kang, H.Y. LPS induces melanogenesis through p38 MAPK activation in human melanocytes. Arch. Dermatol. Res. 2008, 300, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Widlude, H.R.; Fisher, D.E. Microphthalmia-associated transcription factor: A critical regulator of pigment cell development and survival. Oncogene 2003, 22, 3035–3041. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.; Singh, S.K.; Sarkar, C.; Bera, R.; Ratha, J.; Tobin, D.J.; Bhadra, R. Activation of the Mitf promoter by lipid-stimulated activation of p38-stress signaling to CREB. Pigment Cell Res. 2006, 19, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.C.; Lin, Y.Y.; Yang, S.Y.; Weng, Y.; Tsai, Y. Antimelanogenic effect of c-phycocyanin through modulation of tyrosinase expression by upregulation of ERK and downregulation of p38 MAPK signaling pathways. J. Biomed. Sci. 2011, 18, 74. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.J.; Choi, B.R.; Lee, E.K.; Kim, S.H.; Yi, H.Y.; Park, H.R.; Song, C.H.; Lee, Y.J.; Ku, S.K. Inhibitory Effect of Dried Pomegranate Concentration Powder on Melanogenesis in B16F10 Melanoma Cells; Involvement of p38 and PKA Signaling Pathways. Int. J. Mol. Sci. 2015, 16, 24219–24242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.S.; Sung, J.H.; Lee, S.K. Antimelanogenesis Activity of Hydrolyzed Ginseng Extract (GINST) via Inhibition of JNK Mitogen-activated Protein Kinase in B16F10 Cells. J. Food Sci. 2016, 81, H2085–H2092. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Hong, E.; Choi, M.; Kim, K.; Lee, S. Antioxidant and Skin Whitening Effects of Rhamnus yoshinoi Extracts. Korean J. Food Sci. Anim. Resour. 2010, 42, 750–754. [Google Scholar]
- Shukla, Y.N.; Lloyd, H.A.; Morton, J.F.; Kapadia, G. Iridoid glycosides and other constituents of Paederia foetida. Phytochemistry 1976, 15, 1989–1990. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, Y.; Han, T.; Zheng, C.; Qin, L. A phytochemical, pharmacological and clinical profile of Paederia foetida and P. scandens. Nat. Prod. Commun. 2014, 9, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanda, S.; Sarethy, I.P.; De, B.; Singh, K. Paederia foetida—A promishing ethnomedicinal tribal plant of Northeast India. J. Forestry Res. 2013, 24, 801–808. [Google Scholar] [CrossRef]
- Pradhan, N.; Parbin, S.; Kausar, C.; Kar, S.; Mawatwal, S.; Das, L.; Deb, M.; Sengupta, D.; Dhiman, R.; Patra, S.K. Paederia foetida induces anticancer activity by modulating chromatin modification enzymes and altering pro-inflammatory cytokine gene expression in human prostate cancer cells. Food Chem. Toxicol. 2019, 130, 161–173. [Google Scholar] [CrossRef]
- Afroz, S.; Alamgir, M.; Khan, M.T.H.; Jabbar, S.; Nahar, N.; Choudhuri, M.S.K. Antidiarrhoeal activity of the ethanol extract of Paederia foetida Linn. (Rubiaceae). J. Ethnopharmacol. 2006, 105, 125–130. [Google Scholar] [CrossRef]
- Kumar, V.; Al-Abbasi, F.A.; Ahmed, D.; Verma, A.; Mujeeb, M.; Anwar, F. Paederia foetida Linn. Paederia foetida Linn. inhibits adjuvant induced arthritis by suppression of PGE(2) and COX-2 expression via nuclear factor-κB. Food Funct. 2015, 6, 1652–1666. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.; Deb, L.; Tiwari, R.K.; Singh, K.; Ahmad, S. Gastroprotective mechanism of Paederia foetida Linn. (Rubiaceae)—A popular edible plant used by the tribal community of North-East India. BMC Complement. Altern. Med. 2015, 13, 304. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.M.; Son, Y.O.; Lee, S.A.; Jeon, Y.M.; Lee, J.C. Quercetin Inhibits α-MSH-stimulated Melanogenesis in B16F10 Melanoma Cells. Phytother. Res. 2011, 25, 1166–1173. [Google Scholar] [CrossRef]
- Seong, Z.K.; Lee, S.Y.; Poudel, A.; Oh, S.R.; Lee, H.K. Constituents of Cryptotaenia japonica Inhibit Melanogenesis via CREB- and MAPK-Associated Signaling Pathways in Murine B16 Melanoma Cells. Molecules 2016, 21, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.Y.; Won, K.J.; Hwang, D.I.; Park, S.M.; Kim, B.; Lee, H.M. Chemical Composition, Antioxidant and Anti-melanogenic Activities of Essential Oils from Chrysanthemum boreale Makino at Different Harvesting Stages. Chem. Biodivers. 2018, 15. [Google Scholar] [CrossRef] [PubMed]
- Truong, X.T.; Park, S.H.; Lee, Y.G.; Jeong, H.Y.; Moon, J.H.; Jeon, T.I. Protocatechuic Acid from Pear Inhibits Melanogenesis in Melanoma Cells. Int. J. Mol. Sci. 2017, 18, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.S.; Cha, S.B.; Park, M.C.; Park, S.A.; Kim, H.S.; Woo, W.H.; Mun, Y.J. Scopoletin Stimulates Melanogenesis via cAMP/PKA Pathway and Partially p38 Activation. Biol. Pharm. Bull. 2017, 40, 2068–2074. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Description of Clinical Observation |
|---|---|
| 0 (−) | No signs of inflammation, normal skin |
| 0.5 (±) | Doubtful or slight reaction |
| 1 (+) | Slight erythema |
| 2 (++) | Moderate erythema, with or without partial edema or papules |
| 3 (+++) | Moderate erythema with diffuse edema |
| 4 (++++) | Intense erythema with diffuse edema and the presence of vesicles |
| Range of Response | Inference |
|---|---|
| 0 ≤ * R < 0.02 | No irritation |
| 0.02 ≤ R < 0.25 | Low irritation |
| 0.25 ≤ R < 1 | Slight irritation |
| 1 ≤ R < 2.5 | Moderate irritation |
| 2.5 ≤ R | Severe irritation |
| NO. | PFE (1%) | Blank | ||||
|---|---|---|---|---|---|---|
| Reaction | Reaction | |||||
| 24 h * | 48 h | 72 h | 24 h * | 48 h | 72 h | |
| Grade | 0.5 | 0 | 0 | 0 | 0 | 0 |
| Total number of observations | 96 | 96 | ||||
| Irritation Index | 0.005 | 0 | ||||
| Judgment | No irritation | - | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, Y.C.; Lee, J.N.; Kim, B.S.; Hyun, C.-G. Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation. Cosmetics 2021, 8, 22. https://doi.org/10.3390/cosmetics8010022
Chung YC, Lee JN, Kim BS, Hyun C-G. Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation. Cosmetics. 2021; 8(1):22. https://doi.org/10.3390/cosmetics8010022
Chicago/Turabian StyleChung, You Chul, Jung No Lee, Bong Seok Kim, and Chang-Gu Hyun. 2021. "Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation" Cosmetics 8, no. 1: 22. https://doi.org/10.3390/cosmetics8010022
APA StyleChung, Y. C., Lee, J. N., Kim, B. S., & Hyun, C. -G. (2021). Anti-Melanogenic Effects of Paederia foetida L. Extract via MAPK Signaling-Mediated MITF Downregulation. Cosmetics, 8(1), 22. https://doi.org/10.3390/cosmetics8010022