
Cryptomphalus aspersa Eggs Extract Potentiates Human Epidermal Stem Cell Regeneration and Amplification

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
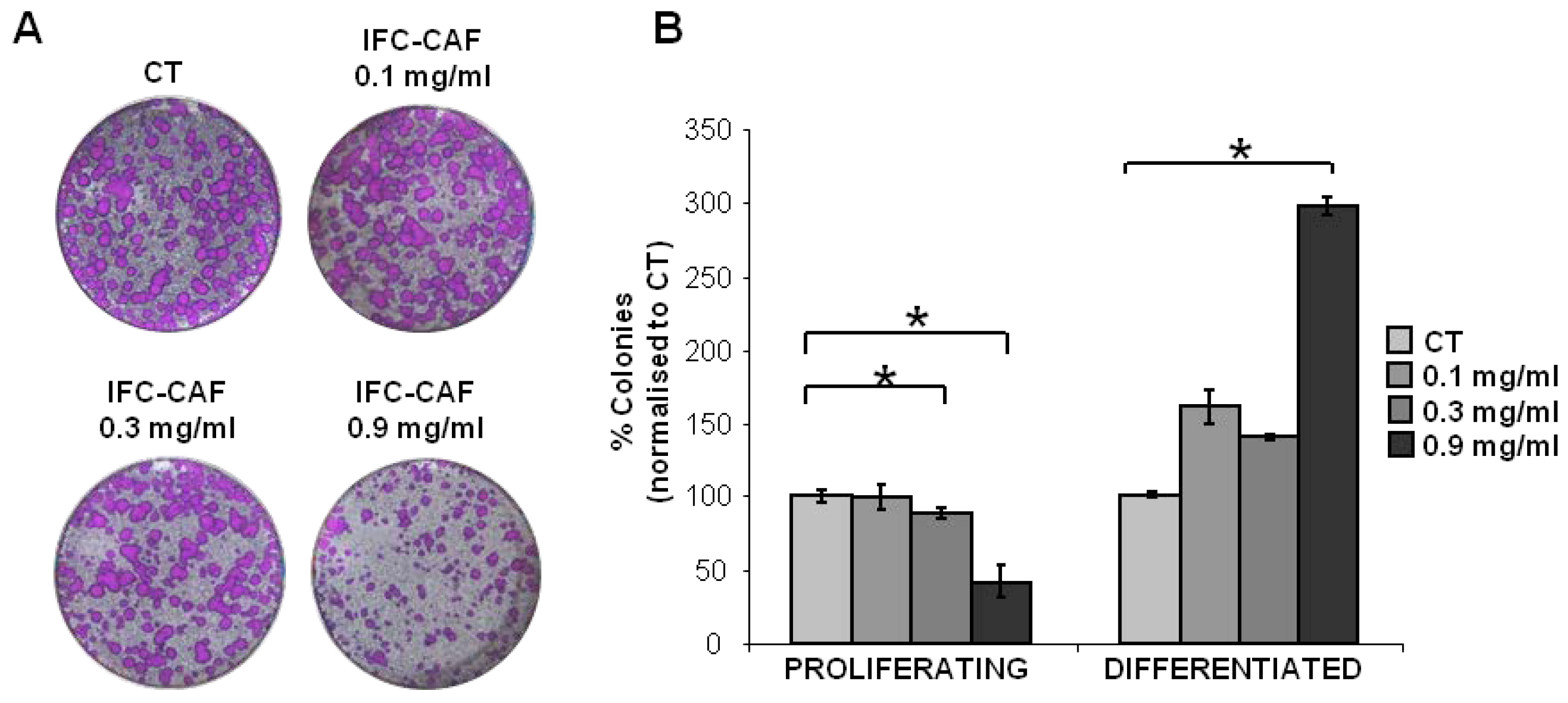
2.1. Evaluation of IFC-CAF® Impact on Primary Keratinocyte Proliferation
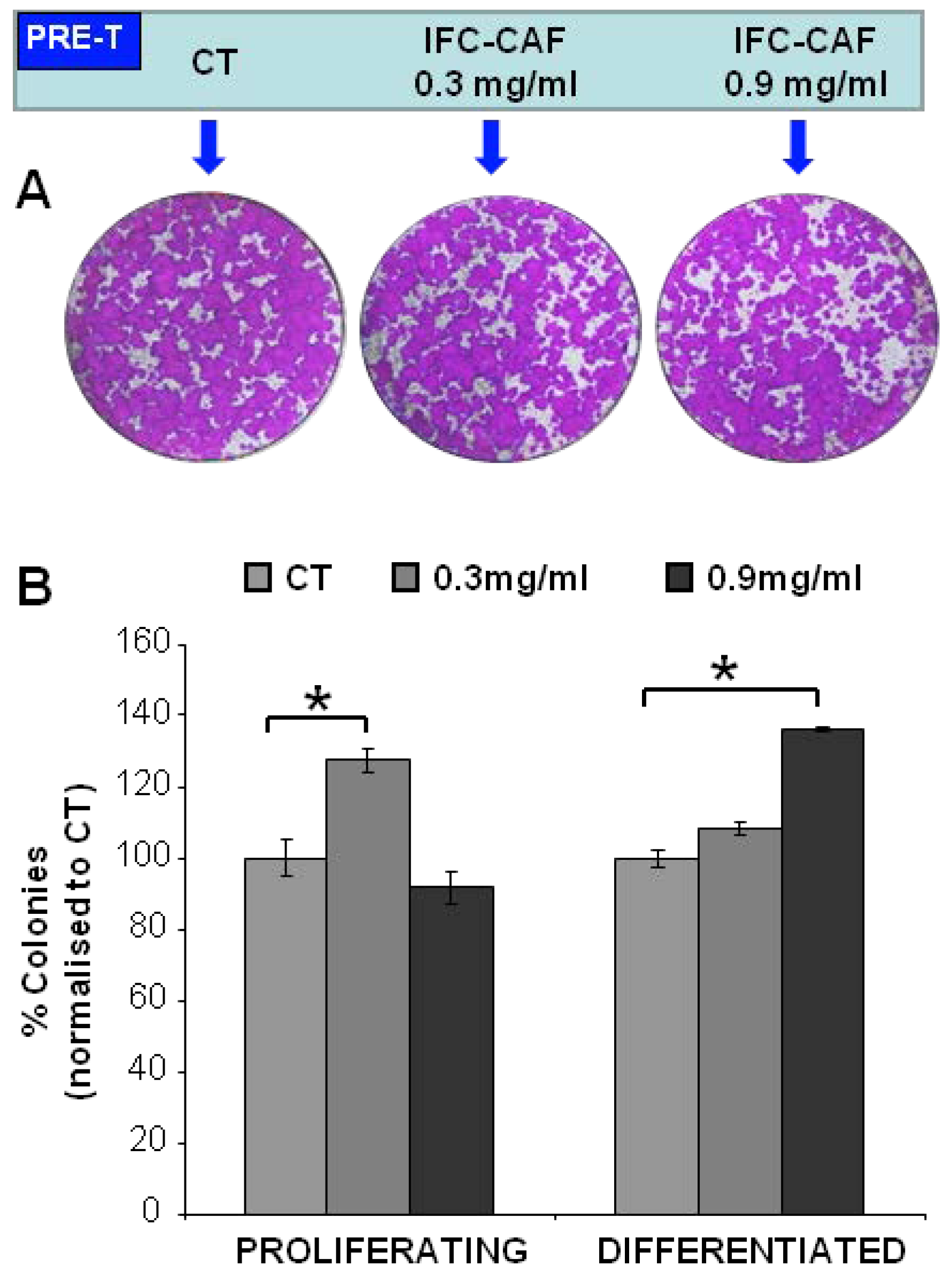
2.2. IFC-CAF® Promotes Epidermal Formation In Vitro
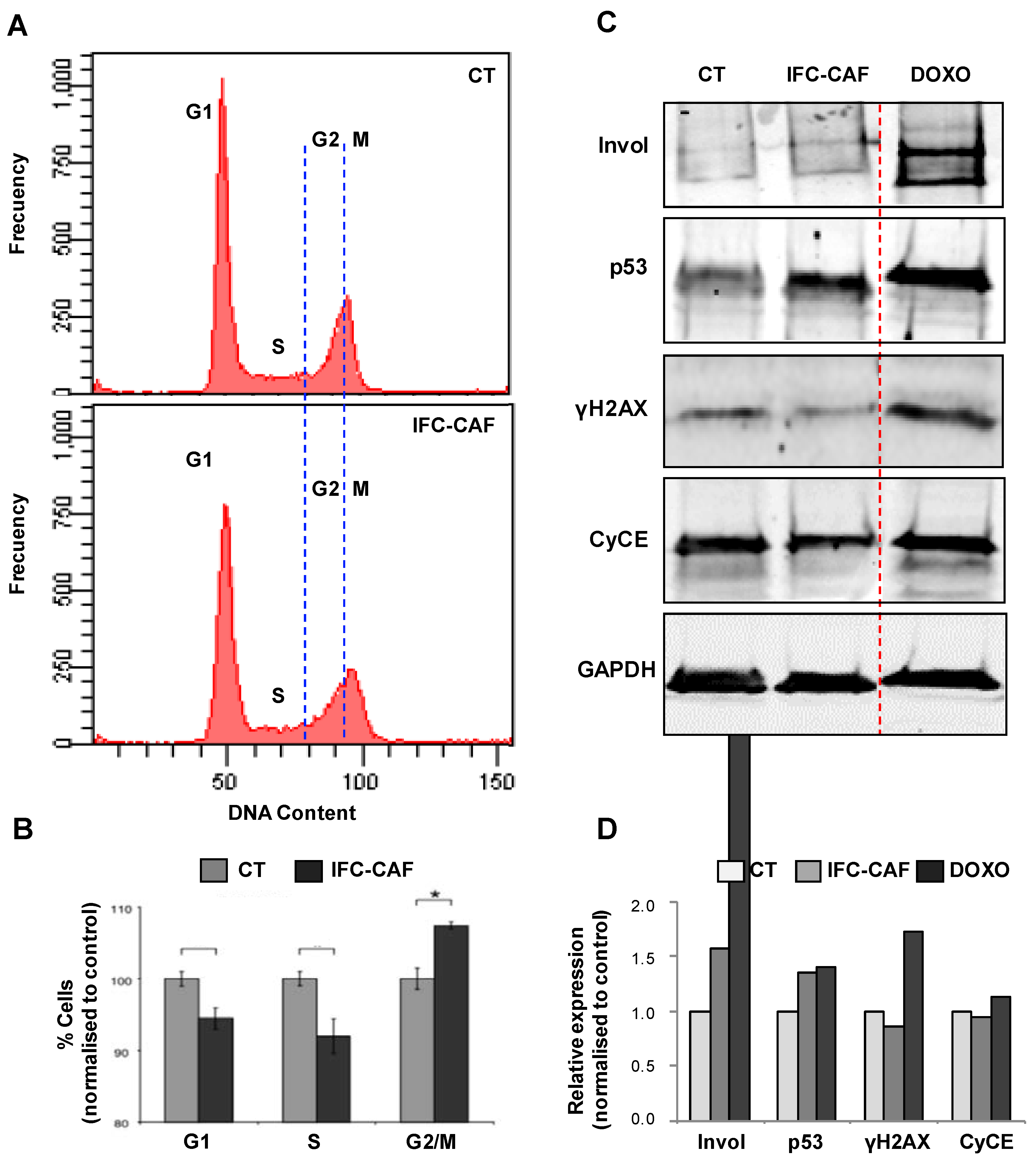
2.3. IFC-CAF® Slows Progression through the Keratinocyte G2/M Cell Cycle Transition and Activates Genome Integrity Surveillance Pathways
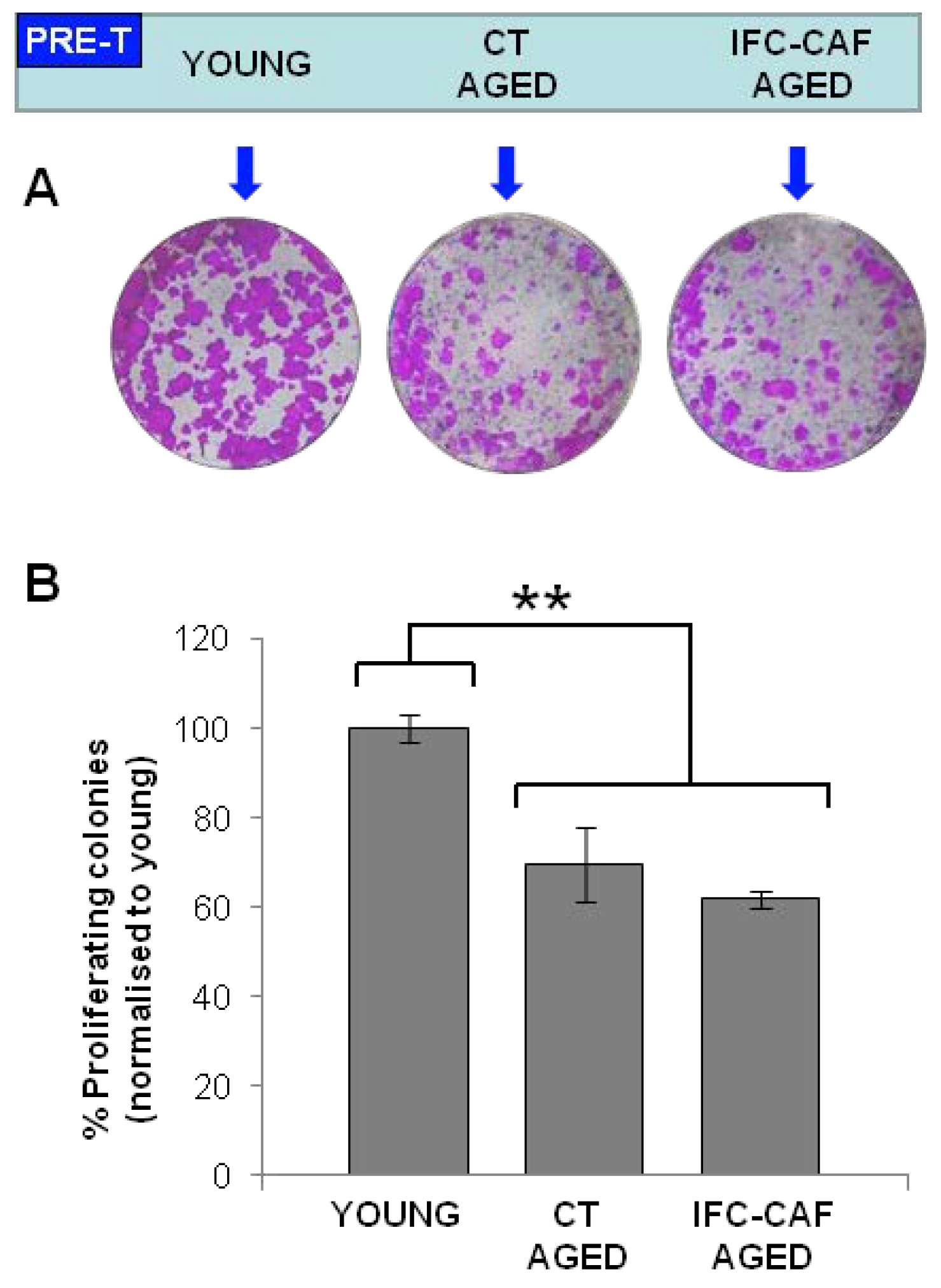
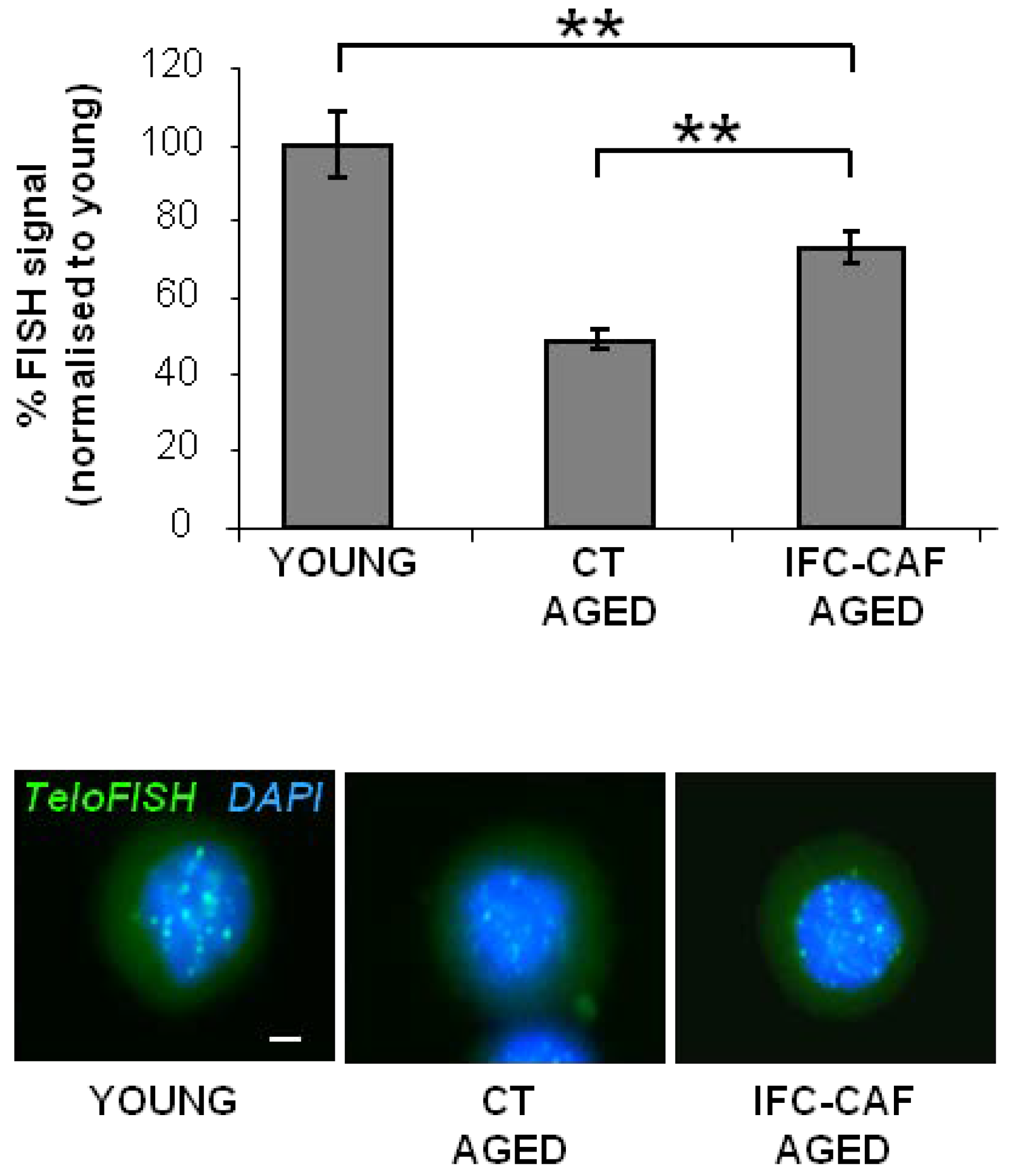
2.4. IFC-CAF® Improves Telomere Maintenance
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Antibodies
4.4. Flow Cytometry
4.5. Confocal Microscopy
4.6. Whole-Cell Extracts and Western Blotting
4.7. Replicative Aging of Keratinocytes
4.8. Telomere Length Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sundaram, H. The mechanisms and potential impact of stem cell activation in skin rejuvenation: An evidence-based analysis. J. Drugs Dermatol. 2017, 16, 378–384. [Google Scholar] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Otin, C.; Galluzzi, L.; Freije, J.M.P.; Madeo, F.; Kroemer, G. Metabolic Control of Longevity. Cell 2016, 166, 802–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.C.; Ji, J.A.; Jiang, Z.Y.; You, Q.D. The Keap1–Nrf2–ARE Pathway As a Potential Preventive and Therapeutic Target: An Update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef]
- Brewer, J.W.; Diehl, J.A. PERK mediates cell-cycle exit during the mammalian unfolded protein response. Proc. Natl. Acad. Sci. USA 2000, 97, 12625–12630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorrio, S.; Rodríguez-Luna, A.; Delgado-Wicke, P.; Mascaraque, M.; Gallego, M.; Pérez-Davó, A.; González, S.; Juarranz, Á. Protective effect of the aqueous extract of Deschampsia antarctica (EDAFENCE®) on skin cells against blue light emitted from digital devices. Int. J. Mol. Sci. 2020, 21, 988. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Espin, A.; Morel, E.; Juarranz, A.; Guerrero, A.; Gonzalez, S.; Jimenez, A.; Sevilla, F. An Extract from the Plant Deschampsia antarctica Protects Fibroblasts from Senescence Induced by Hydrogen Peroxide. Oxid. Med. Cell. Longev. 2017, 2017, 2694945. [Google Scholar] [CrossRef] [Green Version]
- Milani, M.; Hashtroody, B.; Piacentini, M.; Celleno, L. Skin protective effects of an antipollution, antioxidant serum containing Deschampsia antartica extract, ferulic acid and vitamin C: A controlled single-blind, prospective trial in women living in urbanized, high air pollution area. Clin. Cosmet. Investig. Dermatol. 2019, 12, 393–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gombau, L.; García, F.; Lahoz, A.; Fabre, M.; Roda-Navarro, P.; Majano, P.; Alonso-Lebrero, J.L.; Pivel, J.P.; Castell, J.V.; Gomez-Lechon, M.J.; et al. Polypodium leucotomos extract: Antioxidant activity and disposition. Toxicol. In Vitro 2006, 20, 464–471. [Google Scholar] [CrossRef]
- Gonzalez, S.; Gilaberte, Y.; Philips, N.; Juarranz, A. Fernblock, a Nutriceutical with Photoprotective Properties and Potential Preventive Agent for Skin Photoaging and Photoinduced Skin Cancers. Int. J. Mol. Sci. 2011, 12, 8466–8475. [Google Scholar] [CrossRef] [Green Version]
- Brieva, A.; Philips, N.; Tejedor, R.; Guerrero, A.; Pivel, J.P.; Alonso-Lebrero, J.L.; Gonzalez, S. Molecular basis for the regenerative properties of a secretion of the mollusk Cryptomphalus aspersa. Skin Pharmacol. Physiol. 2008, 21, 15–22. [Google Scholar] [CrossRef]
- Alameda, M.T.; Morel, E.; Parrado, C.; González, S.; Juarranz, Á. Cryptomphalus aspersa mollusc egg extract promotes regenerative effects in human dermal papilla stem cells. Int. J. Mol. Sci. 2017, 18, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espada, J.; Matabuena, M.; Salazar, N.; Lucena, S.; Kourani, O.; Carrasco, E.; Calvo, M.; Rodriguez, C.; Reyes, E.; Gonzalez, S.; et al. Cryptomphalus aspersa mollusc eggs extract promotes migration and prevents cutaneous ageing in keratinocytes and dermal fibroblasts in vitro. Int. J. Cosmet. Sci. 2015, 37, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Draelos, Z.D. The Role of a Natural Mollusk Egg-Derived Ingredient in Facial Appearance. J. Drugs Dermatol. 2017, 16, 678–681. [Google Scholar] [PubMed]
- García-Honduvilla, N.; Cifuentes, A.; Ortega, M.A.; Delgado, A.; González, S.; Bujan, J.; Alvarez-Mon, M. High Sensitivity of Human Adipose Stem Cells to Differentiate into Myofibroblasts in the Presence of C. aspersa Egg Extract. Stem Cells Int. 2017, 2017, 9142493. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.H.; Watt, F.M. Separation of human epidermal stem cells from transit amplifying cells on the basis of differences in integrin function and expression. Cell 1993, 73, 713–724. [Google Scholar] [CrossRef]
- Rheinwald, J.G. Human epidermal keratinocyte cell culture and xenograft systems: Applications in the detection of potential chemical carcinogens and the study of epidermal transformation. Prog. Clin. Biol. Res. 1989, 298, 113–125. [Google Scholar] [PubMed]
- Bayerl, C.; Taake, S.; Moll, I.; Jung, E.G. Characterization of sunburn cells after exposure to ultraviolet light. Photodermatol. Photoimmunol. Photomed. 1995, 11, 149–154. [Google Scholar] [CrossRef]
- Henry, K.A.; Blank, H.M.; Hoose, S.A.; Polymenis, M. The unfolded protein response is not necessary for the G1/S transition, but it is required for chromosome maintenance in Saccharomyces cerevisiae. PLoS ONE 2010, 5, e12732. [Google Scholar] [CrossRef] [Green Version]
- Fragkos, M.; Jurvansuu, J.; Beard, P. H2AX is required for cell cycle arrest via the p53/p21 pathway. Mol. Cell. Biol. 2009, 29, 2828–2840. [Google Scholar] [CrossRef] [Green Version]
- Freije, A.; Molinuevo, R.; Ceballos, L.; Cagigas, M.; Alonso-Lecue, P.; Rodriguez, R.; Menendez, P.; Aberdam, D.; De Diego, E.; Gandarillas, A. Inactivation of p53 in Human Keratinocytes Leads to Squamous Differentiation and Shedding via Replication Stress and Mitotic Slippage. Cell Rep. 2014, 9, 1349–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermeulen, R.; Schymanski, E.L.; Barabasi, A.-L.; Miller, G.W. The exposome and health: Where chemistry meets biology. Science 2020, 367, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aylon, Y.; Oren, M. p53: Guardian of ploidy. Mol. Oncol. 2011, 5, 315–323. [Google Scholar] [CrossRef]
- Molinuevo, R.; Freije, A.; Contreras, L.; Sanz, J.R.; Gandarillas, A. The DNA damage response links human squamous proliferation with differentiation. J. Cell Biol. 2020, 219, e202001063. [Google Scholar] [CrossRef]
- Sanz-Gómez, N.; de Pedro, I.; Ortigosa, B.; Santamaría, D.; Malumbres, M.; de Cárcer, G.; Gandarillas, A. Squamous differentiation requires G2/mitosis slippage to avoid apoptosis. Cell Death Differ. 2020, 27, 2451–2467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, L.K.; Reed, S.I. Cyclin E Deregulation and Genomic Instability. Adv. Exp. Med. Biol. 2017, 1042, 527–547. [Google Scholar]
- Feng, Y.X.; Sokol, E.S.; Del Vecchio, C.A.; Sanduja, S.; Claessen, J.H.L.; Proia, T.A.; Jin, D.X.; Reinhardt, F.; Ploegh, H.L.; Wang, Q.; et al. Epithelial-to-mesenchymal transition activates PERK-eIF2α and sensitizes cells to endoplasmic reticulum stress. Cancer Discov. 2014, 4, 702–715. [Google Scholar] [CrossRef] [Green Version]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef]
- Yeo, S.Y.; Lee, K.W.; Shin, D.; An, S.; Cho, K.H.; Kim, S.H. A positive feedback loop bi-stably activates fibroblasts. Nat. Commun. 2018, 9, 3016. [Google Scholar] [CrossRef]
- Walter, P.; Ron, D. The unfolded protein response: From stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Alvarez, M.; Bakal, C. PERK links the clock and protein stress in cancer. Nat. Cell Biol. 2018, 20, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Dufey, E.; Bravo-San Pedro, J.M.; Eggers, C.; Gonzalez-Quiroz, M.; Urra, H.; Sagredo, A.I.; Sepulveda, D.; Pihán, P.; Carreras-Sureda, A.; Hazari, Y.; et al. Genotoxic stress triggers the activation of IRE1alpha-dependent RNA decay to modulate the DNA damage response. Nat. Commun. 2020, 11, 2401. [Google Scholar] [CrossRef]
- Sanchez-Alvarez, M.; Zhang, Q.; Finger, F.; Wakelam, M.J.O.; Bakal, C. Cell cycle progression is an essential regulatory component of phospholipid metabolism and membrane homeostasis. Open Biol. 2015, 5, 150093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollien, J. Evolution of the unfolded protein response. Biochim. Et Biophys. Acta-Mol. Cell Res. 2013, 1833, 2458–2463. [Google Scholar] [CrossRef] [Green Version]
- Sequeira, S.J.; Ranganathan, A.C.; Adam, A.P.; Iglesias, B.V.; Farias, E.F.; Aguirre-Ghiso, J.A. Inhibition of proliferation by PERK regulates mammary acinar morphogenesis and tumor formation. PLoS ONE 2007, 2, e615. [Google Scholar]
- Sanz-Gómez, N.; Freije, A.; Gandarillas, A. Keratinocyte Differentiation by Flow Cytometry. Methods Mol. Biol. 2020, 2109, 83–92. [Google Scholar] [PubMed]
- Achtstaetter, T.; Hatzfeld, M.; Quinlan, R.A.; Parmelee, D.C.; Franke, W.W. Separation of cytokeratin polypeptides by gel electrophoretic and chromatographic techniques and their identification by immunoblotting. Methods Enzymol. 1986, 134, 355–371. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
San Juan, L.; de Pedro, I.; Rodríguez-Luna, A.; Villalba, M.; Guerrero, A.; González, S.; Gandarillas, A. Cryptomphalus aspersa Eggs Extract Potentiates Human Epidermal Stem Cell Regeneration and Amplification. Cosmetics 2022, 9, 2. https://doi.org/10.3390/cosmetics9010002
San Juan L, de Pedro I, Rodríguez-Luna A, Villalba M, Guerrero A, González S, Gandarillas A. Cryptomphalus aspersa Eggs Extract Potentiates Human Epidermal Stem Cell Regeneration and Amplification. Cosmetics. 2022; 9(1):2. https://doi.org/10.3390/cosmetics9010002
Chicago/Turabian StyleSan Juan, Lucía, Isabel de Pedro, Azahara Rodríguez-Luna, María Villalba, Antonio Guerrero, Salvador González, and Alberto Gandarillas. 2022. "Cryptomphalus aspersa Eggs Extract Potentiates Human Epidermal Stem Cell Regeneration and Amplification" Cosmetics 9, no. 1: 2. https://doi.org/10.3390/cosmetics9010002
APA StyleSan Juan, L., de Pedro, I., Rodríguez-Luna, A., Villalba, M., Guerrero, A., González, S., & Gandarillas, A. (2022). Cryptomphalus aspersa Eggs Extract Potentiates Human Epidermal Stem Cell Regeneration and Amplification. Cosmetics, 9(1), 2. https://doi.org/10.3390/cosmetics9010002