
Streptomyces spp. Isolated from Rosa davurica Rhizome for Potential Cosmetic Application


 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Identification of Strain THA-663
2.3. Liquid–Liquid Extraction (LLE)
2.4. Identification of Components by UPLC-Qtof-MS
2.5. DPPH and ABTS Radical Scavenging Activity
2.6. Antibacterial Activity
2.7. MIC and MBC
2.8. Cell Culture and Treatment
2.9. Cell Viability
2.10. Reactive Oxygen Species (ROS) Scavenging Activity
2.11. Enzyme-Linked Immunosorbent Assay (ELISA)
2.12. Reverse Transcription Polymerase Chain Reaction (RT-PCR) Analysis
2.13. Western Blot Analysis
2.14. Statistical Analysis
3. Results
3.1. Molecular Phylogenetic Identification
3.2. Antimicrobial Activity of THA-663S on Methicillin-Resistant Staphylococcus aureus (MRSA)
3.3. Identification of Components in THA-663S by UPLC-Qtof-MS
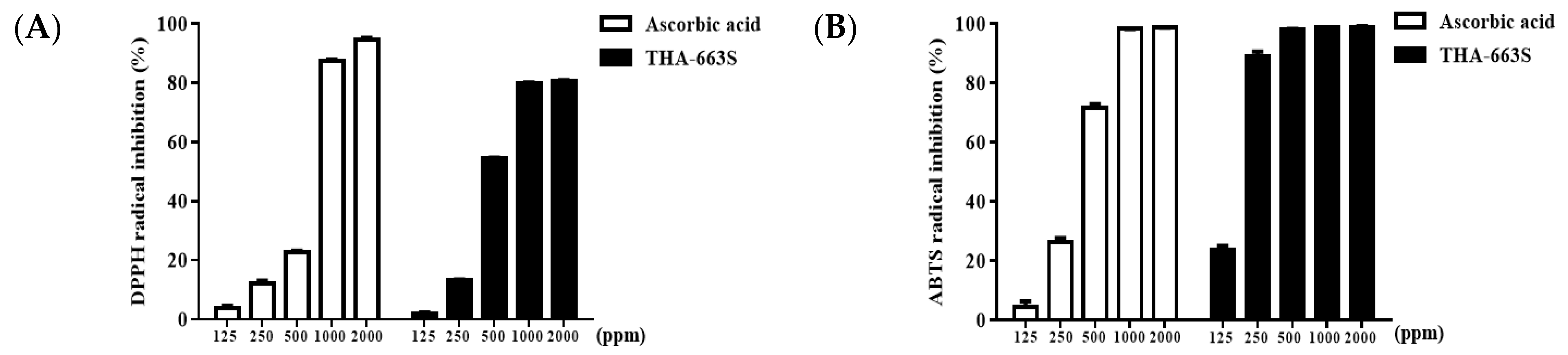
3.4. DPPH and ABTS Radical Scavenging Activity
3.5. MIC and MBC
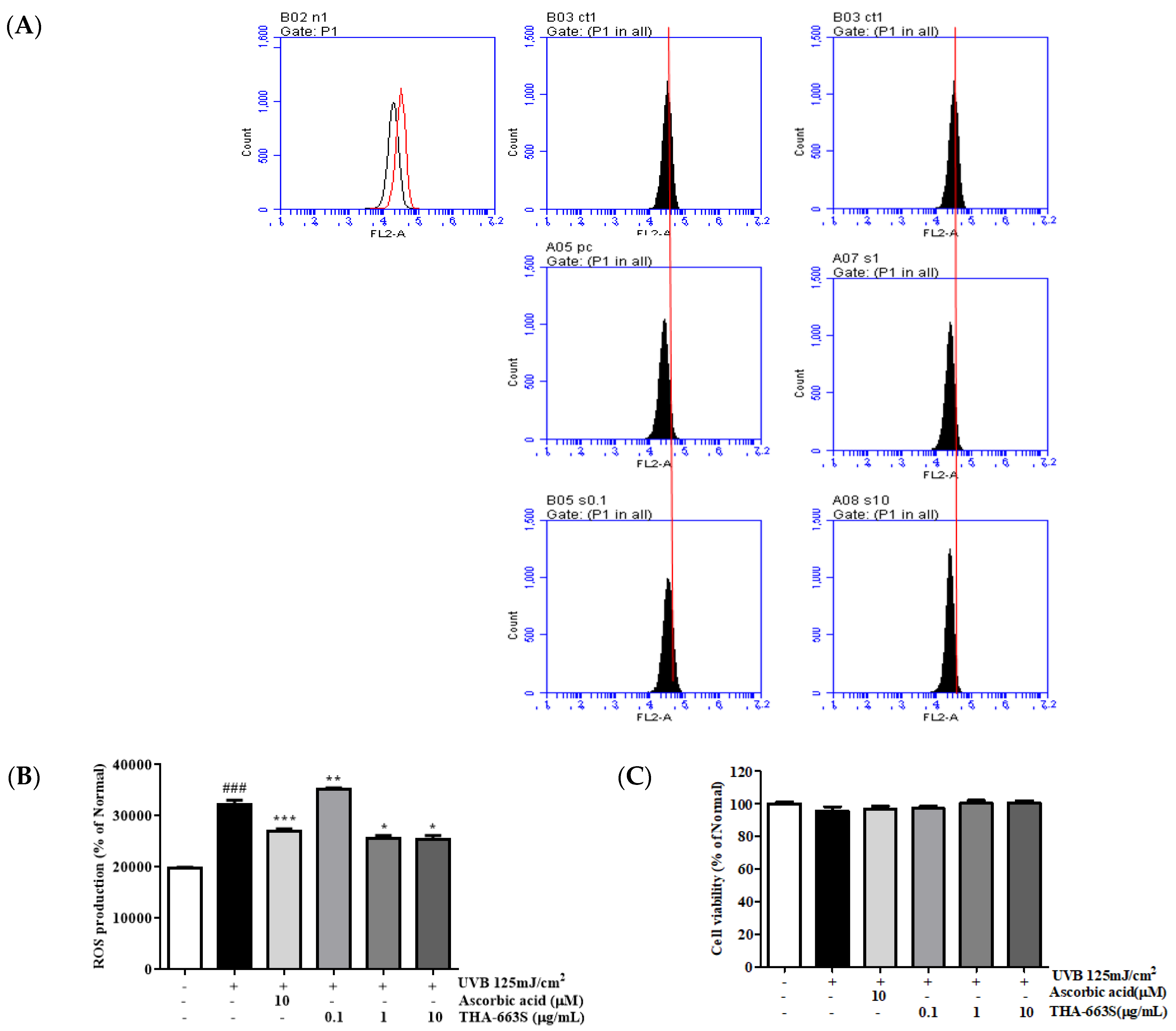
3.6. Effect of THA-663S on Cell Viability and Cytoprotective
3.7. Effect of THA-663S on MMP-1 and MMP-3 Secretion in UVB-Irradiated HaCaT Cells
3.8. Effect of THA-663S on MMP-1, TGF-β1, and Procollagen Type I mRNA Expression in UVB-Irradiated HaCaT Cells
3.9. Effect of THA-663S on Protein Expression of MAPK/AP-1 Signaling Pathway
3.10. Effect of THA-663S on Protein Expression of TGF-β/Smad Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Makrantonaki, E.; Zouboulis, C.C. Skin anti-aging strategies. Derm.-Endocrinol. 2012, 4, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Chambers, E.S.; Vukmanovic-Stejic, M. Skin barrier immunity and ageing. Immunology 2020, 160, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jager, T.L.; Cockrell, A.E.; Du Plessis, S.S. Ultraviolet Light Induced Generation of Reactive Oxygen Species. Adv. Exp. Med. Biol. 2017, 996, 15–23. [Google Scholar] [PubMed]
- Kwon, K.-R.; Alam, M.B.; Park, J.-H.; Kim, T.-H.; Lee, S.-H. Attenuation of UVB-Induced Photo-Aging by Polyphenolic-Rich Spatholobus Suberectus Stem Extract via Modulation of MAPK/AP-1/MMPs Signaling in Human Keratinocytes. Nutrients 2019, 11, 1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Uchi, H.; Furue, M. Antioxidant cinnamaldehyde attenuates UVB-induced photoaging. J. Dermatol. Sci. 2019, 96, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.-J.; Ji, Y.; Jang, Y.-P.; Choung, S.-Y. Acer tataricum subsp. ginnala Inhibits Skin Photoaging via Regulating MAPK/AP-1, NF-κB, and TGFβ/Smad Signaling in UVB-Irradiated Human Dermal Fibroblasts. Molecules 2021, 26, 662. [Google Scholar] [CrossRef]
- Munshi, A.; Ramesh, R. Mitogen-Activated Protein Kinases and Their Role in Radiation Response. Genes Cancer 2013, 4, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Karin, M. The Regulation of AP-1 Activity by Mitogen-activated Protein Kinases. J. Biol. Chem. 1995, 270, 16483–16486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, S.J.; Bowden, G.T. Ultraviolet B regulation of transcription factor families: Roles of nuclear factor-kappa B (NF-κB) and activator protein-1 (AP-1) in UVB-induced skin carcinogenesis. Curr. Cancer Drug Targets 2007, 7, 325–334. [Google Scholar] [CrossRef]
- Li, N.; Karin, M. Ionizing radiation and short wavelength UV activate NF-κB through two distinct mechanisms. Proc. Natl. Acad. Sci. USA 1998, 95, 13012–13017. [Google Scholar] [CrossRef]
- He, T.; Quan, T.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Oxidative exposure impairs TGF-β pathway via reduction of type II receptor and SMAD3 in human skin fibroblasts. Age 2014, 36, 9623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, T.; He, T.; Voorhees, J.J.; Fisher, G.J. Ultraviolet Irradiation Blocks Cellular Responses to Transforming Growth Factor-β by Down-regulating Its Type-II Receptor and Inducing Smad7. J. Biol. Chem. 2001, 276, 26349–26356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omura, S.; Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Takahashi, C.; Shinose, M.; Takahashi, Y.; Horikawa, H.; Nakazawa, H.; Osonoe, T.; et al. Genome sequence of an industrial microorganism Streptomyces avermitilis: Deducing the ability of producing secondary metabolites. Proc. Natl. Acad. Sci. USA 2001, 98, 12215–12220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bĕhal, V. Bioactive products from Streptomyces. Adv. Appl. Microbiol. 2000, 47, 113–156. [Google Scholar]
- Laube, S.; Farrell, A.M. Bacterial Skin Infections in the Elderly. Drugs Aging 2002, 19, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Lee, H.M.; Oh, S.; Zheng, S.; Bellere, A.D.; Kim, M.; Choi, J.; Kim, M.; Yu, D.; Yi, T.H. Rosa davurica inhibits skin photoaging via regulating MAPK/AP-1, NF-κB, and Nrf2/HO-1 signaling in UVB-irradiated HaCaTs. Photochem. Photobiol. Sci. 2022; Advance online publication. [Google Scholar] [CrossRef]
- Flärdh, K.; Buttner, M.J. Streptomyces morphogenetics: Dissecting differentiation in a filamentous bacterium. Nat. Rev. Microbiol. 2009, 7, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Davoodbasha, M.; Edachery, B.; Nooruddin, T.; Lee, S.-Y.; Kim, J.-W. An evidence of C16 fatty acid methyl esters extracted from microalga for effective antimicrobial and antioxidant property. Microb. Pathog. 2018, 115, 233–238. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Arifiyanto, A.; Surtiningsih, T.; Ni’Matuzahroh; Fatimah; Agustina, D.; Alami, N.H. Antimicrobial activity of biosurfactants produced by actinomycetes isolated from rhizosphere of Sidoarjo mud region. Biocatal. Agric. Biotechnol. 2020, 24, 101513. [Google Scholar] [CrossRef]
- Sahin, N. Antimicrobial activity of Streptomyces species against mushroom blotch disease pathogen. J. Basic Microbiol. 2005, 45, 64–71. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Sarkar, F.H. Signaling mechanism(s) of reactive oxygen species in Epithelial-Mesenchymal Transition reminiscent of cancer stem cells in tumor progression. Curr. Stem Cell Res. Ther. 2010, 5, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verrecchia, F.; Mauviel, A. Transforming growth factor-beta signaling through the Smad pathway: Role in extracellular matrix gene expression and regulation. J. Investig. Dermatol. 2002, 118, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chater, K.F. Streptomyces inside-out: A new perspective on the bacteria that provide us with antibiotics. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Gromkowska-Kępka, K.J.; Puścion-Jakubik, A.; Markiewicz-Żukowska, R.; Socha, K. The impact of ultraviolet radiation on skin photoaging—Review of in vitro studies. J. Cosmet. Dermatol. 2021, 20, 3427–3431. [Google Scholar] [CrossRef] [PubMed]
- Brandner, J.; Zorn-Kruppa, M.; Yoshida, T.; Moll, I.; Beck, L.; De Benedetto, A. Epidermal tight junctions in health and disease. Tissue Barriers 2014, 3, e974451. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.; Hwang, E.; Ngo, H.T.T.; Seo, S.A.; Yi, T.H. Sambucus nigra L. ameliorates UVB-induced photoaging and inflammatory response in human skin keratinocytes. Cytotechnology 2019, 71, 1003–1017. [Google Scholar] [CrossRef]
- Yang, M.; Huang, C.Z. Mitogen-activated protein kinase signaling pathway and invasion and metastasis of gastric cancer. World J. Gastroenterol. 2015, 21, 11673–11679. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Hwang, E.; Ngo, H.T.T.; Lin, P.; Gao, W.; Liu, Y.; Yi, T.-H. Antiphotoaging Effect of Prunus yeonesis Blossom Extract via Inhibition of MAPK/AP-1 and Regulation of the TGF-βI/Smad and Nrf2/ARE Signaling Pathways. Photochem. Photobiol. 2018, 94, 725–732. [Google Scholar] [CrossRef]
- Choi, H.-J.; Alam, M.B.; Baek, M.-E.; Kwon, Y.-G.; Lim, J.-Y.; Lee, S.-H. Protection against UVB-Induced Photoaging by Nypa fruticans via Inhibition of MAPK/AP-1/MMP-1 Signaling. Oxid. Med. Cell. Longev. 2020, 2020, 2905362. [Google Scholar] [CrossRef]
- Woo, Y.K.; Park, J.; Ryu, J.H.; Cho, H.J. The anti-inflammatory and anti-apoptotic effects of advanced anti-inflammation composition (AAIC) in heat shock-induced human HaCaT keratinocytes. J. Cosmet. Dermatol. 2020, 19, 2114–2124. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.Y.; Kim, S.N.; Chae, J.H.; Hyun, S.S.; Seo, S.Y.; Lee, Y.S.; Lee, K.O.; Kim, S.H.; Lee, Y.S.; Jeong, J.M.; et al. Syntheses and anti-MRSA activities of the C3 analogs of mansonone F, a potent anti-bacterial sesquiterpenoid: Insights into its structural requirements for anti-MRSA activity. Bioorg. Med. Chem. Lett. 2004, 14, 4519–4523. [Google Scholar] [CrossRef] [PubMed]
- Fortuna, C.G.; Bonaccorso, C.; Bulbarelli, A.; Caltabiano, G.; Rizzi, L.; Goracci, L.; Musumarra, G.; Pace, A.; Piccionello, A.P.; Guarcello, A.; et al. New linezolid-like 1,2,4-oxadiazoles active against Gram-positive multiresistant pathogens. Eur. J. Med. Chem. 2013, 65, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cheng, K.; Wang, X.; Yin, H. Selection, synthesis, and anti-inflammatory evaluation of the arylidene malonate derivatives as TLR4 signaling inhibitors. Bioorg. Med. Chem. 2012, 20, 6073–6079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, R.M.P. Antihepatotoxic, nephroprotective, and antioxidant activities of phenolic compounds from Satureja macrostema leaves against carbon tetrachloride-induced hepatic damage in mice. Med. Chem. Res. 2013, 22, 1846–1855. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | THA-663 | KACC 14678 2 | KACC 21704 3 |
|---|---|---|---|
| pH range | 5–9 | 5–9 | 5–9 |
| Optimum pH | 6–7 | 6–7 | 6–8 |
| 10 °C | - | W 1 | - |
| 30 °C | + | + | + |
| 37 °C | + | + | + |
| Optimum temperature | 30 | 30 | 37 |
| Maximum NaCl con. (%, w/v) | 4% | 6% | 4% |
| Antibiotics | THA-663 | KACC 14678 1 | KACC 21074 2 |
|---|---|---|---|
| Kanamycin | 1.0 | 1.0 | 1.5 |
| Ampicillin | >256 | >256 | -1) |
| Erythromycin | 1.5 | 0.38 | 0.38 |
| Streptomycin | 0.5 | 0.5 | 0.75 |
| Vancomycin | 0.38 | 0.38 | 0.38 |
| Clindamycin | 48 | 1.5 | 0.25 |
| Gentamycin | 0.38 | 0.25 | 0.38 |
| Tetracycline | 48 | 8 | 32 |
| Chloramphenicol | - | >256 | - |
| Microorganisms | Collection Code No. | THA-663S | AMP 1 | NOR 2 | GEN 3 | NAT 4 |
|---|---|---|---|---|---|---|
| S. aureus 285 | CCARM 0204 | 1.31 ± 0.4% | - | - | 1.1 ± 0.1% | - |
| S. aureus 503 | CCARM 0205 | 1.36 ± 0.3% | - | - | 1.02 ± 0.2% | - |
| S. aureus | CCARM 3855 | 1.02 ± 0.4% | 1.39 ± 0.1% | - | - | - |
| S. aureus | CCARM 3089 | 1.01 ± 0.2% | - | - | - | - |
| No. | Component Name | Observed RT (min) | Formula | Observed m/z | Mass Error (mDa) | Adducts |
|---|---|---|---|---|---|---|
| 1 | 3,6,9-Trimethyl-benzo[de]chromene | 1.29 | C15H14O | 233.0940 | 0.3 | +Na |
| 2 | 2-(4-ethoxybenzylidene)malonic acid | 2.25 | C11H10O5 | 223.0598 | −0.3 | +H |
| 3 | N-[[3-[3-fluoro-4-(3-methyl-1,2,4-oxadiazol-5-yl)phenyl]-2-oxo-1,3-oxazolidin-5-yl]methyl]acetamide | 3.58 | C16H18N2O6 | 335.1226 | −1.2 | +H |
| 4 | 1H-Indole, 3-(1E)-1-propen-1-yl- | 3.80 | C11H11N | 158.0964 | 0.0 | +H |
| 5 | Kenganthranol A | 4.17 | C25H28O5 | 431.1810 | −1.9 | +Na |
| 6 | [(2-Methoxyphenyl) carbamoyl]methyl 3-chloroadamantane-1-carboxylate | 6.56 | C20H24ClNO4 | 378.1452 | −1.5 | +H |
| 7 | 7-[(2S,3R,4S,5R,6R)-3,5-dihydroxy-6-(hydroxymethyl)-4-[(2S,3R,4R,5R,6S)-3,4,5-trihydroxy-6-methyloxan-2-yl]oxyoxan-2-yl]oxy-5-hydroxy-3,6-dimethoxy-2-(4-methoxyphenyl)chromen-4-one | 9.88 | C30H36O16 | 653.2083 | 0.7 | +H |
| Gram | Microorganisms | Collection Code No. | MIC | MBC |
|---|---|---|---|---|
| Gram-positive | S. aureus 285 | CCARM 0204 | 2.5 | 5 |
| S. aureus 503 | CCARM 0205 | 2.5 | 5 | |
| S. aureus | CCARM 3855 | 1.25 | 5 | |
| S. aureus | CCARM 3089 | 5 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, S.; Oh, S.; Fang, M.; Bellere, A.D.; Jung, J.; Nguyen, T.T.M.; Jeong, J.; Yi, T.-H. Streptomyces spp. Isolated from Rosa davurica Rhizome for Potential Cosmetic Application. Cosmetics 2022, 9, 126. https://doi.org/10.3390/cosmetics9060126
Zheng S, Oh S, Fang M, Bellere AD, Jung J, Nguyen TTM, Jeong J, Yi T-H. Streptomyces spp. Isolated from Rosa davurica Rhizome for Potential Cosmetic Application. Cosmetics. 2022; 9(6):126. https://doi.org/10.3390/cosmetics9060126
Chicago/Turabian StyleZheng, Shengdao, Sarang Oh, Minzhe Fang, Arce Defeo Bellere, Jeyong Jung, Trang Thi Minh Nguyen, Jeehaeng Jeong, and Tae-Hoo Yi. 2022. "Streptomyces spp. Isolated from Rosa davurica Rhizome for Potential Cosmetic Application" Cosmetics 9, no. 6: 126. https://doi.org/10.3390/cosmetics9060126
APA StyleZheng, S., Oh, S., Fang, M., Bellere, A. D., Jung, J., Nguyen, T. T. M., Jeong, J., & Yi, T. -H. (2022). Streptomyces spp. Isolated from Rosa davurica Rhizome for Potential Cosmetic Application. Cosmetics, 9(6), 126. https://doi.org/10.3390/cosmetics9060126