1. Introduction
Neuromorphic hardware is a promising intelligent hardware in next-generation information and communication technologies [
1,
2]. In particular, memristor-based neuromorphic hardware imitates the event-driven characteristics of biological neural networks and the paradigm of processing in memory, which not only greatly reduces hardware power consumption but also improves the real-time performance of information processing [
3,
4]. Therefore, it distinguishes itself from traditional artificial intelligence hardware and has become a popular research field. However, the application of neuromorphic hardware always faces reliability issues [
5,
6], and it is necessary to take more action on its reliability design.
To support computation, biological neurons must modulate within an acceptable dynamic range; always-active or never-active neurons convey no information [
7,
8] and even cause pathological states such as neurological dysfunction and cell death in the nervous system [
8,
9]. In order to achieve this goal, the biological brain always manages to compensate for changes in neuron activity caused by internal and external disturbances, thereby ensuring the robustness of the nervous system function. A large amount of theoretical and experimental evidence [
10,
11] demonstrates that homeostatic inhibitory plasticity is one of the crucial mechanisms to promote the robustness of neuronal function. This mechanism can balance Hebbian plasticity and resist external environmental disturbances and local injury by regulating inhibitory synapses to maintain sparse homeostatic fire neuronal activity [
12,
13,
14]. Consequently, this paper draws on the biological presynaptic homeostatic inhibitory plasticity mechanism to construct a synaptic weight learning circuit and provides a new opportunity for designing reliable neuromorphic hardware.
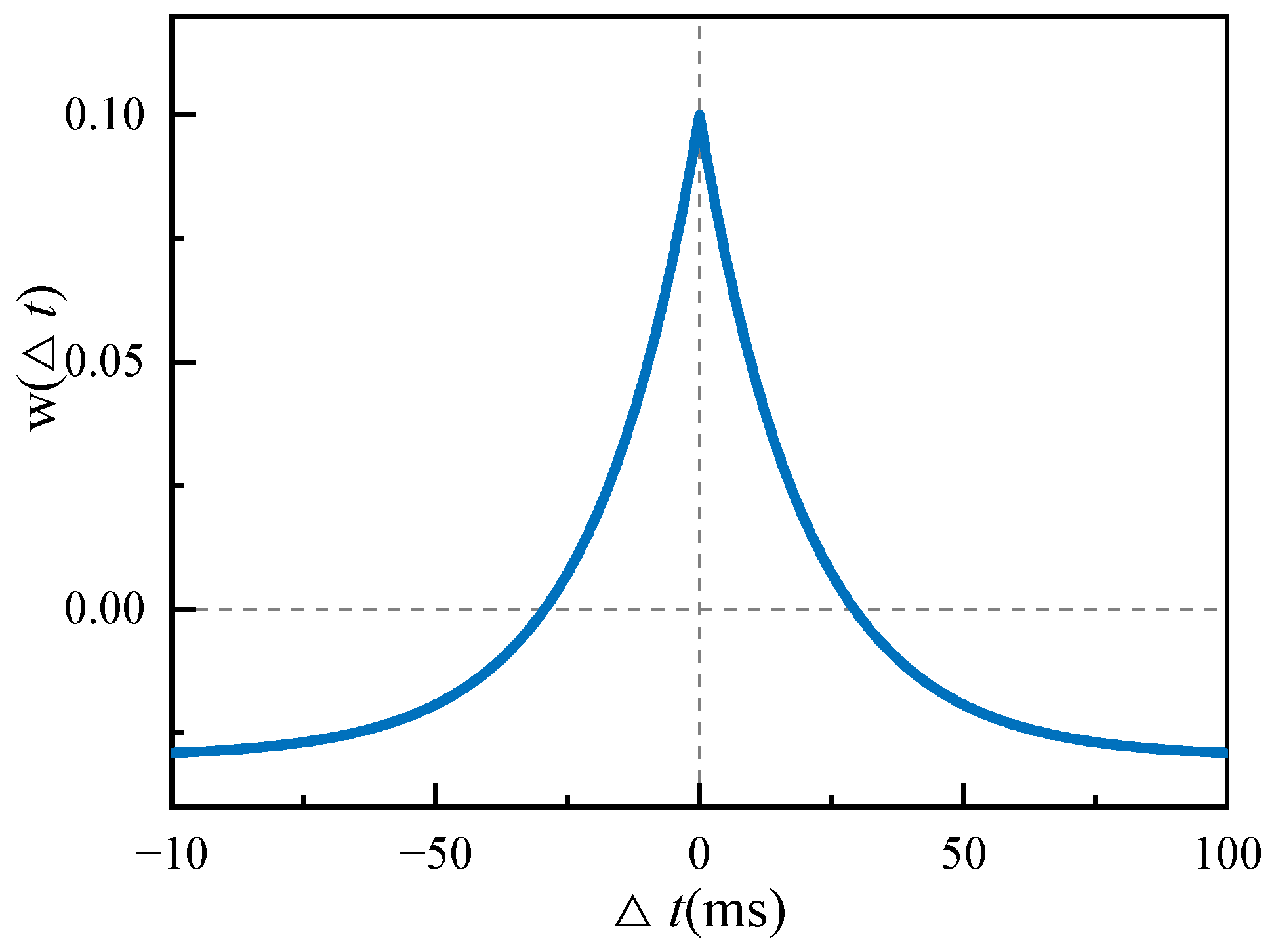
Homeostatic inhibitory plasticity is a spike-timing-dependent plasticity (STDP) mechanism. The mechanism describes an inhibitory synaptic weight change rule: the synaptic weight change
decreases with an increase in |∆
t| and finally stabilizes to a negative value, as shown in
Figure 1. ∆
t =
tpre tpost is the relative time interval of the pre- and postsynaptic spikes.
The homeostatic inhibition plasticity rule can be expressed as follows:
where
A+ and
A− refer to the maximum learning rate;
and
refer to the time constant;
is the minimum value of inhibitory synaptic enhancement,
in
Figure 1; and
α is the homeostatic factor, which determines the homeostatic firing rate of postsynaptic neurons.
In recent years, many studies have been devoted to implementing STDP rule circuits based on memristive synapses. Methods of implementation are typically classified into two categories. The first category implements the STDP mechanism using a single memristive synapse device [
15,
16,
17]. This STDP rule implementation largely depends on the threshold characteristics of the memristor and the shape of the neural action potential waveform. Although this implementation is structurally simple, its adjustment accuracy is low. It must work with extremely precise neuronal circuits that output the proper shape and consistency of neural action potential waveforms. However, designing such a neuronal circuit is not an easy task. The second category accomplishes the STDP mechanism by combining a memristive synapse and an STDP control circuit [
18,
19,
20]. This STDP implementation mainly relies on the STDP control circuit to generate a square wave signal with different pulse widths that modulates the memristive synaptic conductance. Since the pulse width of the square wave signal is precisely adjustable, this implementation approach may achieve accurate adjustment of the memristive synaptic conductance and has also become a popular method for implementing the STDP rules. Related papers [
19,
20] using the second method have achieved the antisymmetric Hebbian learning rule, the antisymmetric anti-Hebbian learning rule, the symmetric Hebbian learning rule, and the symmetric anti-Hebbian learning rule. However, to the best of our knowledge, no circuits that implement the homeostatic inhibitory plasticity rule have been reported in currently published papers.
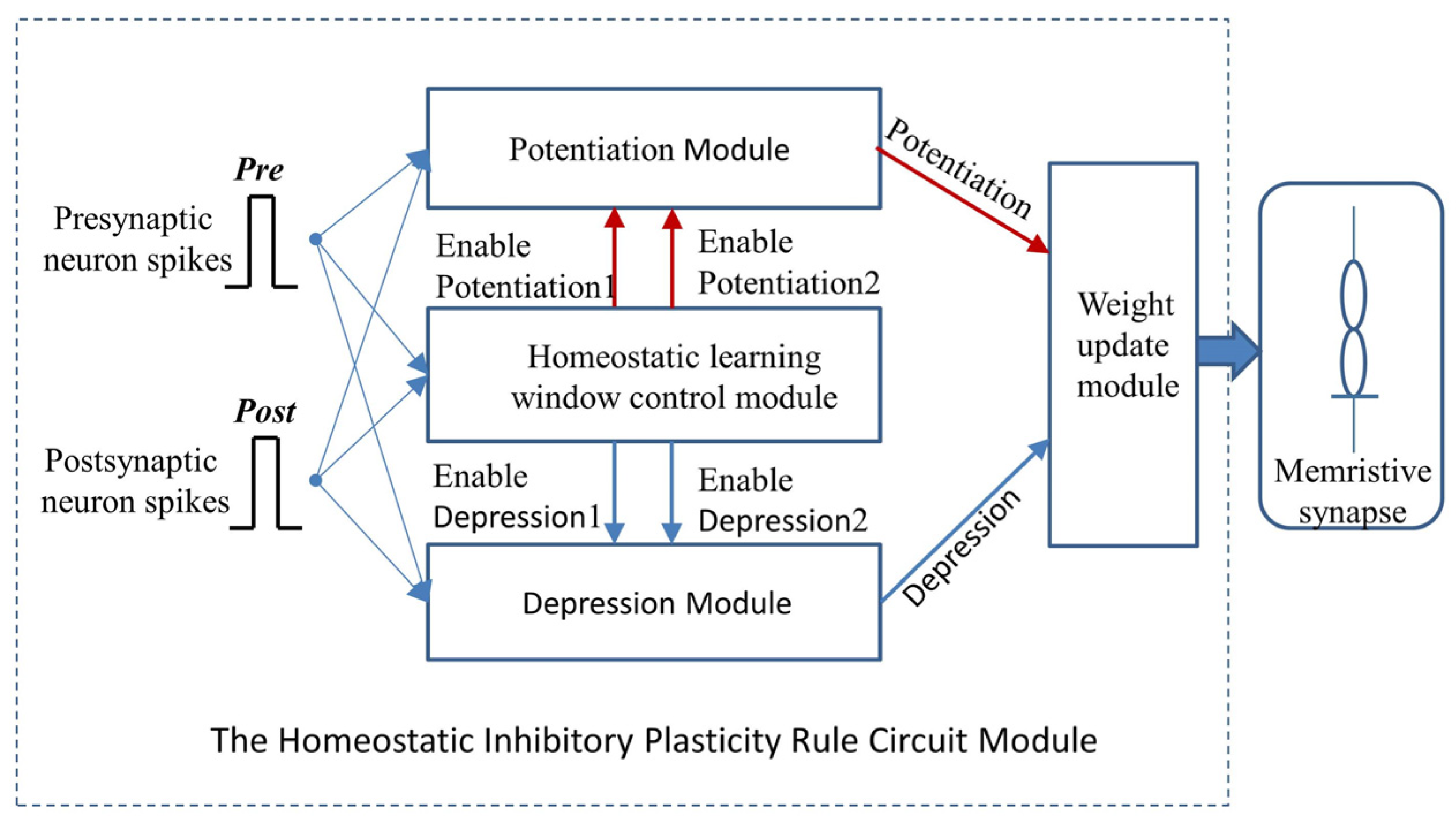
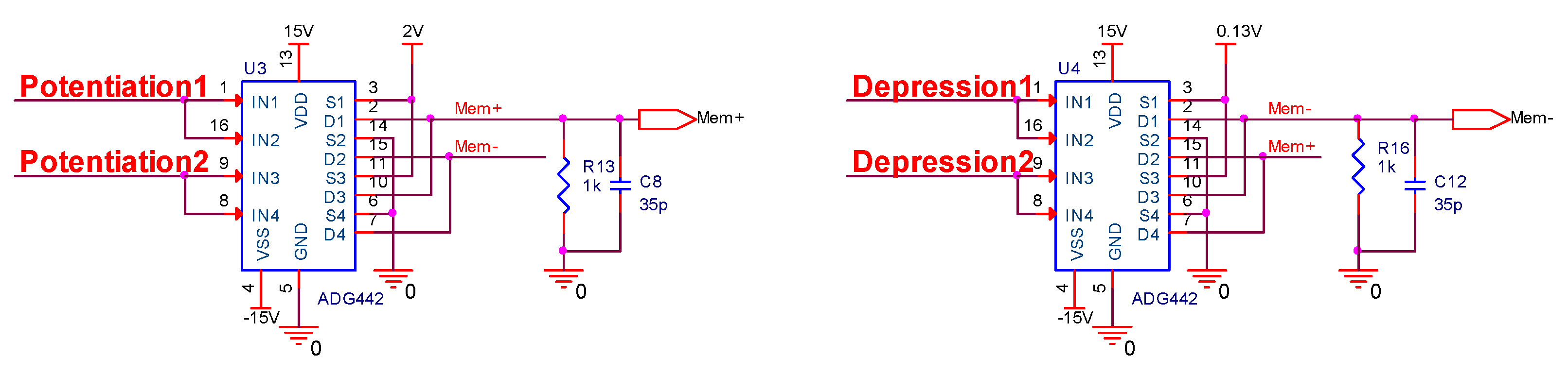
In this research, a homeostatic inhibitory plasticity rule circuit with a memristive synapse is built based on the second STDP implementation approach and the pulse width and amplitude modulation characteristics of memristive conductance, and its performance is verified in PSpice
©. The remainder of this paper is organized as follows: The memristor model applied to the synaptic device is introduced in
Section 2.
Section 3 describes the implementation procedures for the homeostatic inhibitory plasticity circuit.
Section 4 analyzes and discusses the PSpice
© simulation results of the proposed system. The last
Section 5 provides the conclusions.
2. The Memristor Model Used in Synaptic Device
The Knowm self-directed channel (SDC) memristor was the first commercially available memristor fabricated by Knowm Inc. [
21,
22]. Its material stack is a metal ion-conducting device, which relies on Ag+ movement into channels within the active layer to change the device’s resistance. Therefore, the electrical performance of the Knowm memristor is more stable than that of conventional metal oxide memristors [
23]. As a result, it was selected as a potential candidate for the synaptic device. The mathematical model of the Knowm memristor can be represented by Equations (2)–(4).
where
x is the state variable of the memristor;
VON and
VOFF are the thresholds of the memristor ON state and OFF state, respectively;
β is the temperature parameter;
τ is the time constant of the memristor;
Gm is the conductance of the memristor; and
i and
v are the current and voltage through the memristor, respectively. According to our previous research [
24], the
i-
v characteristics of the Knowm memristor can be better simulated when the parameters are set to the following values:
RON = 5.88 kΩ,
ROFF = 44.02 KΩ,
VON = 0.37 V,
VOFF = 0.17 V, and
β = 38.46.
As illustrated in
Figure 2, the PSpice
© model is based on the mathematical model of the Knowm memristor. In
Figure 2a, an extra floating XSV pin was added to the model symbol to facilitate its monitoring of the conductance change in the memristor. The voltage at the XSV pin represents the memristor state variable x.
The full listing of the PSpice
© model of the Knowm memristor is shown in
Appendix A.
4. PSpice© Simulation Results and Discussion
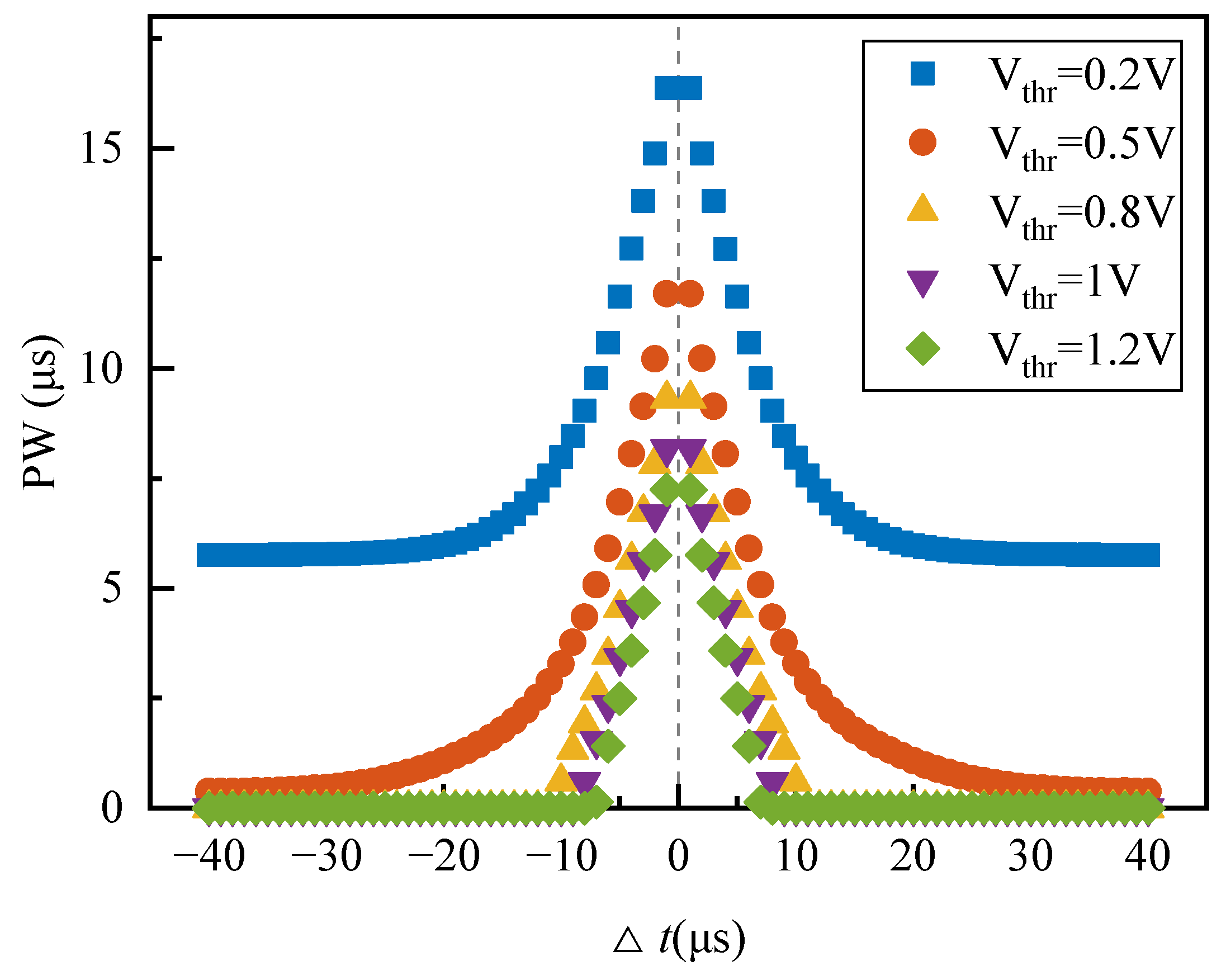
This section presents the PSpice© simulation results of the homeostatic inhibitory plasticity rule circuit with a Knowm memristive synapse. First, the functionality of the proposed circuit is verified by traversing the Δt and applying continuous pre- and postsynaptic pulses. Second, the PSpice© simulation results are discussed.
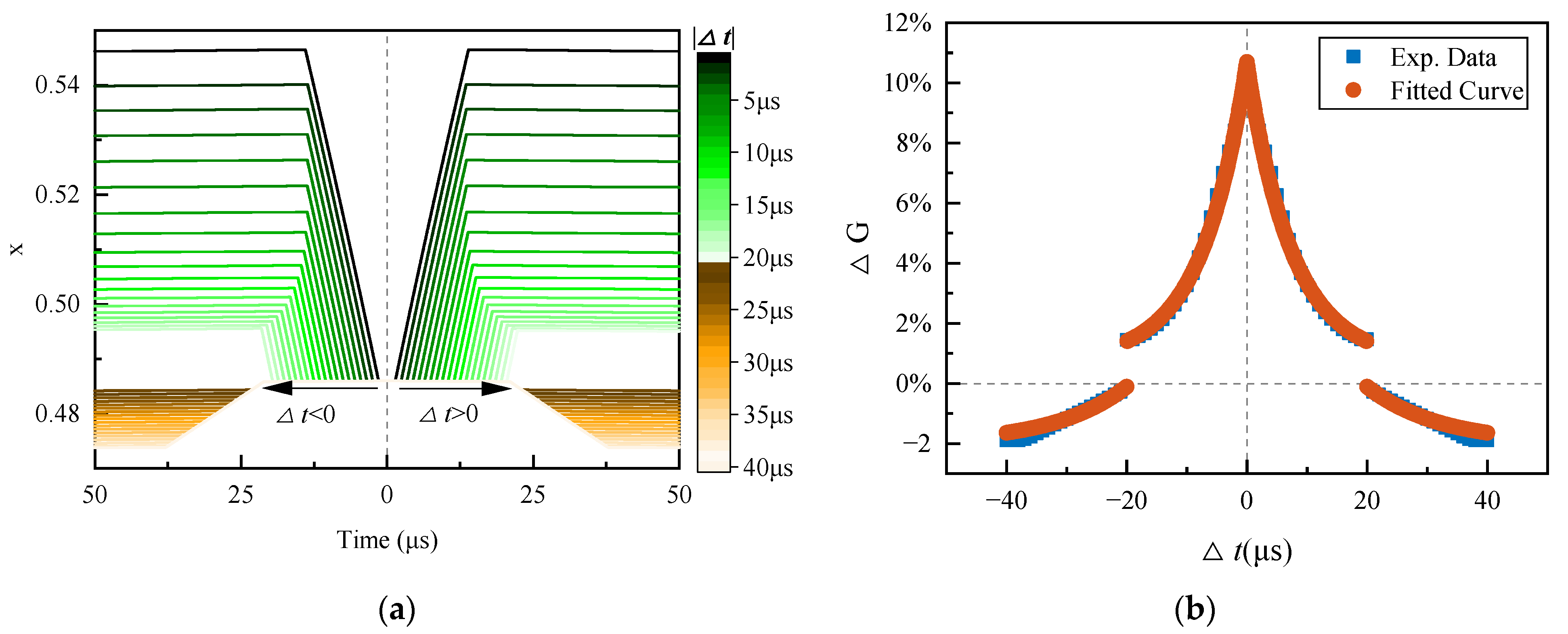
4.1. Traversing the Δt
Set
Vthr = 0.5 V and
Vthr_Depression = 0.6 V. The traversal range of ∆
t is (−40, 40) µs, and the traversal step of ∆
t is 1 µs. Since negative parameters cannot be simulated in PSpice
©, the situations of ∆
t < 0 and ∆
t > 0 are simulated separately with a 50 µs simulation time. The waveform of the state variable
x of the memristor is obtained, as illustrated in
Figure 12a, by monitoring the voltage at the XSV pin of the memristor
V(XSV), since
V(XSV) represents the memristor state variable x. The plot to the left of the dotted line at Time = 0 is the change in
x when ∆
t > 0, while the plot to the right represents the change in
x when ∆
t <0. When |∆
t| ≤ 20 µs,
x increases, and as |∆
t| decreases,
x increases more; when |∆
t| > 20 µs,
x decreases, and as |∆
t| increases,
x decreases more, and then remains constant.
The conductance of the memristor at any time can be calculated from the relationship between the state variable and the conductance (Equation (3)). Assuming G
0 is the initial conductance of the memristor, and G is the updated conductance, the change in the memristive synapse conductance (i.e., the change in synaptic weight) can be calculated as follows:
As shown in
Figure 12b, the blue symbols represent the change in memristive synaptic conductance with ∆
t. Using the biological homeostatic inhibition plasticity learning rule (Equation (1)) to fit the PSpice
© experimental data, it is determined that when
A+ =
A− = 0.1,
= 7.5 µs,
= 12 µs,
, and α = 0.02, Equation (1) can fit the PSpice
© experimental data well (shown in orange symbols in
Figure 12b). It is indicated that the circuit achieves a homeostatic inhibitory plasticity learning rule similar to a biological one.
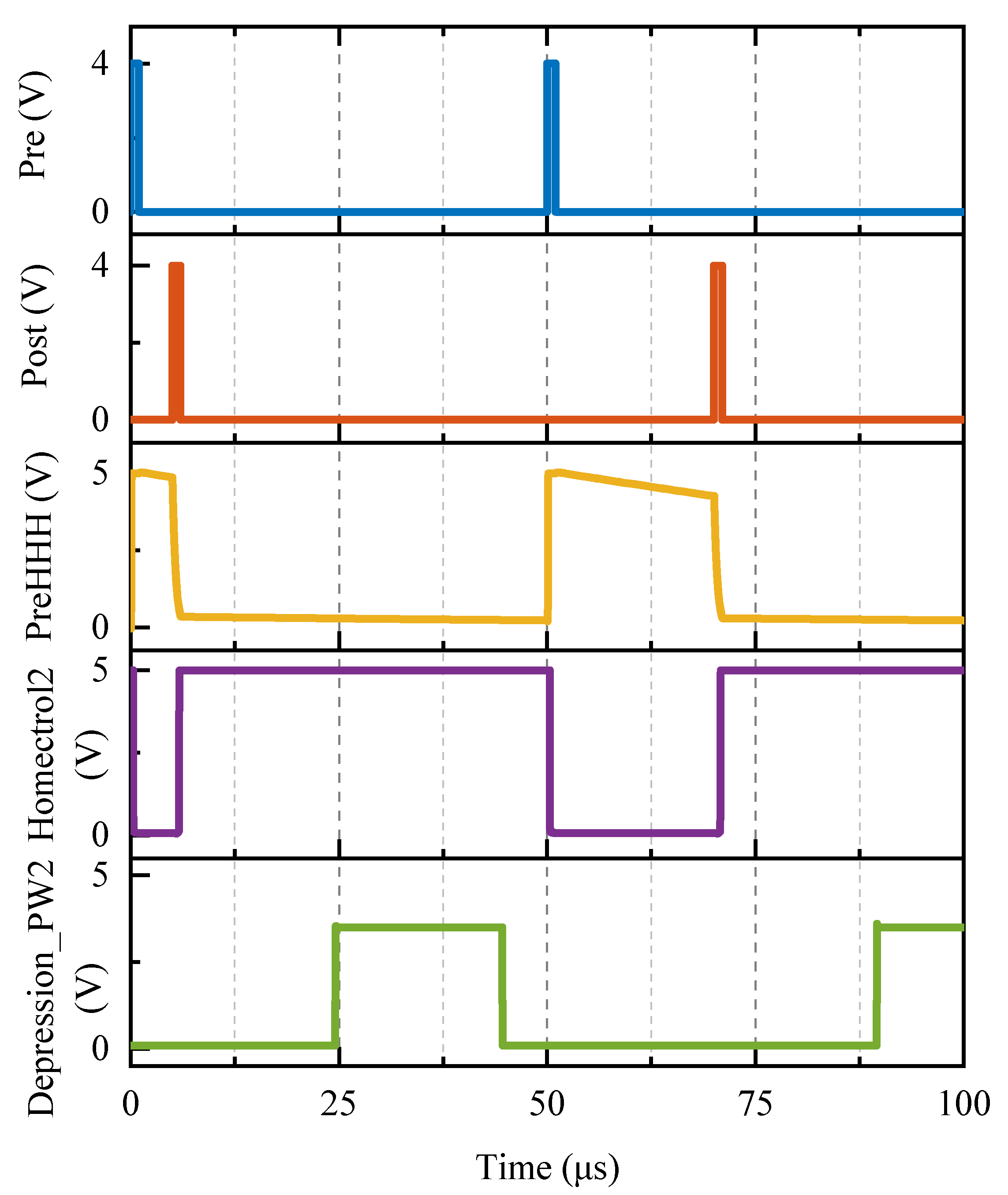
4.2. Applying Continuous Pre- and Postsynaptic Pulses
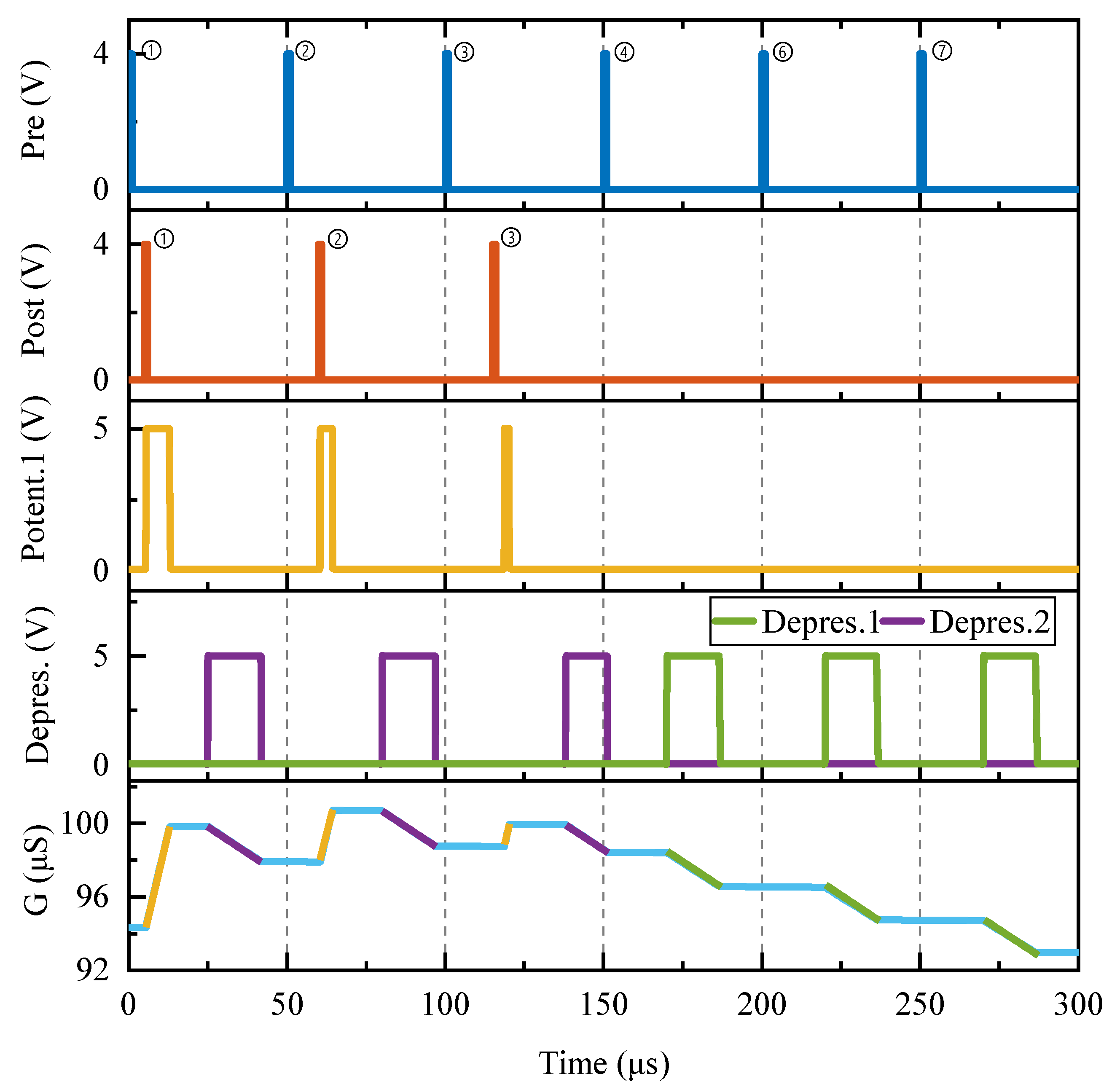
To further verify the function of the homeostatic inhibitory plasticity rule circuit, pre- and postsynaptic neuronal pulses were continuously applied to the proposed circuit at various time intervals.
Figure 13 illustrates the responses of the proposed circuit to continuous pre- and postsynaptic pulses. The time-domain waveforms of
Pre,
Post,
Potentiation1,
Depression1, and
Depression2, and memristive synaptic conductance are depicted from top to bottom. A
Pre is applied every 50 µs, followed by a
Post with a 5 µs, 10 µs, and 18 µs delay relative to the Nos. ➀–➂
Pre, respectively.
First, all
Posts lag behind
Pre by less than 20 µs, causing the enhancement module to output
Potentiation1 and the memristive conductance to increase. The longer the
Post lags behind the
Pre, the narrower the PW of
Potentiation1, reducing the rise in memristive conductance relative to its original value (shown in
Figure 13 by the yellow line superimposed on the conductance change curve). This conclusion is supported by the ∆
t > −20 µs condition depicted in
Figure 12b. Second, Nos. ➁–➃
Pre follows behind Nos. ➀–➂
Post by 45 µs, 40 µs, and 32 µs, all of which are greater than 20 µs. As a result, the depression module generates the depression signal
Depression2, and the memristive conductance falls, as illustrated by the purple line superimposed on the conductance change curve in
Figure 13. It can be concluded that a shorter time
Pre–
Post delay results in a smaller reduction in synaptic conductance. Furthermore, the memristive conductance decreases by the same amount after Nos. ➁–➂
Pre because the PWs of the depression signals are the same when the time interval between
Pre and
Post is greater than or equal to 40 µs. Finally, there is no more
Post after Nos. ➃–➆
Pre, which corresponds to a situation where the time interval between
Post and
Pre is larger than 40 µs; therefore, the memristive conductance will continue to fall by the same amount (as shown by the superimposed green line part on the conductance change curve in
Figure 13). In conclusion, the aforementioned time-domain variation properties of the memristive conductance are totally consistent with the homeostatic inhibitory plasticity rule.
4.3. Discussion
In this research, a homeostatic inhibitory plasticity rule circuit with a commercial memristor synapse is demonstrated. The above simulation results also allow us to summarize the following points. First, as shown in
Figure 12b, both the learning window and time constant of the proposed circuit are in a microsecond time scale. Thus, the information processing speed of the circuit is approximately one thousand times faster than that of the biological [
10], meeting the urgent requirement for neuromorphic hardware with fast computing speed. Second, the
Vthr40 us and
Vthr20 us control the enable signals of the potentiation and depression modules, as well as the homeostatic learning window. Third, because the maximum pulse width of the potentiation and depression signals determines the maximum change in the memristive conductance, the maximum learning rate for synaptic weight enhancement and depression can be adjusted by modifying the respective values of
Vthr and
Vthr_Depression. In addition, the model fitting experiment revealed that the homeostatic factor α is determined by the minimum reduction in memristive synaptic conductance. Consequently, changing
Vthr_Depression can influence the homeostatic factor α: the bigger the
Vthr_Depression, the smaller α, and according to [
10,
11], the smaller the homeostatic firing rate of the postsynaptic neuron.
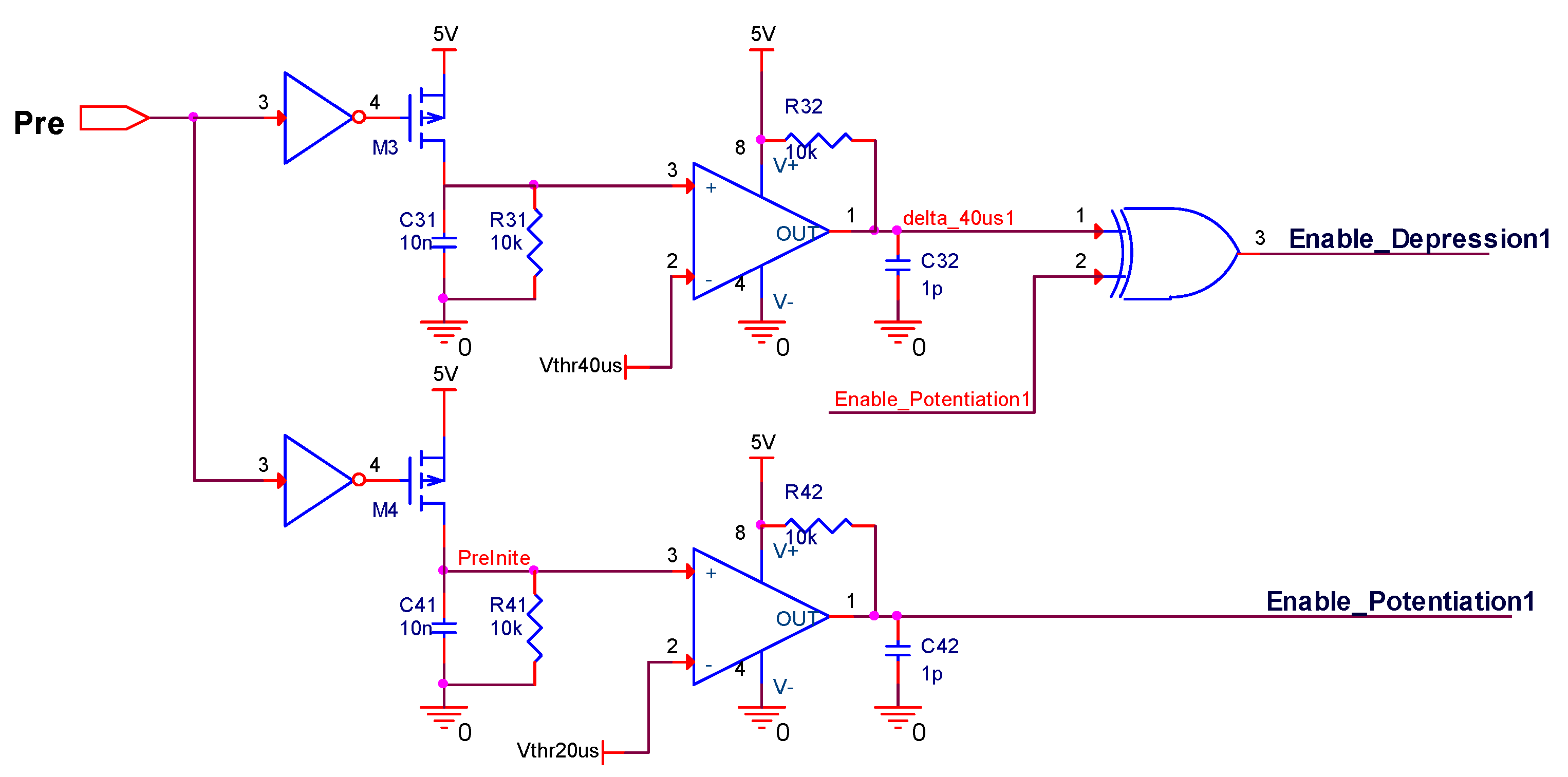
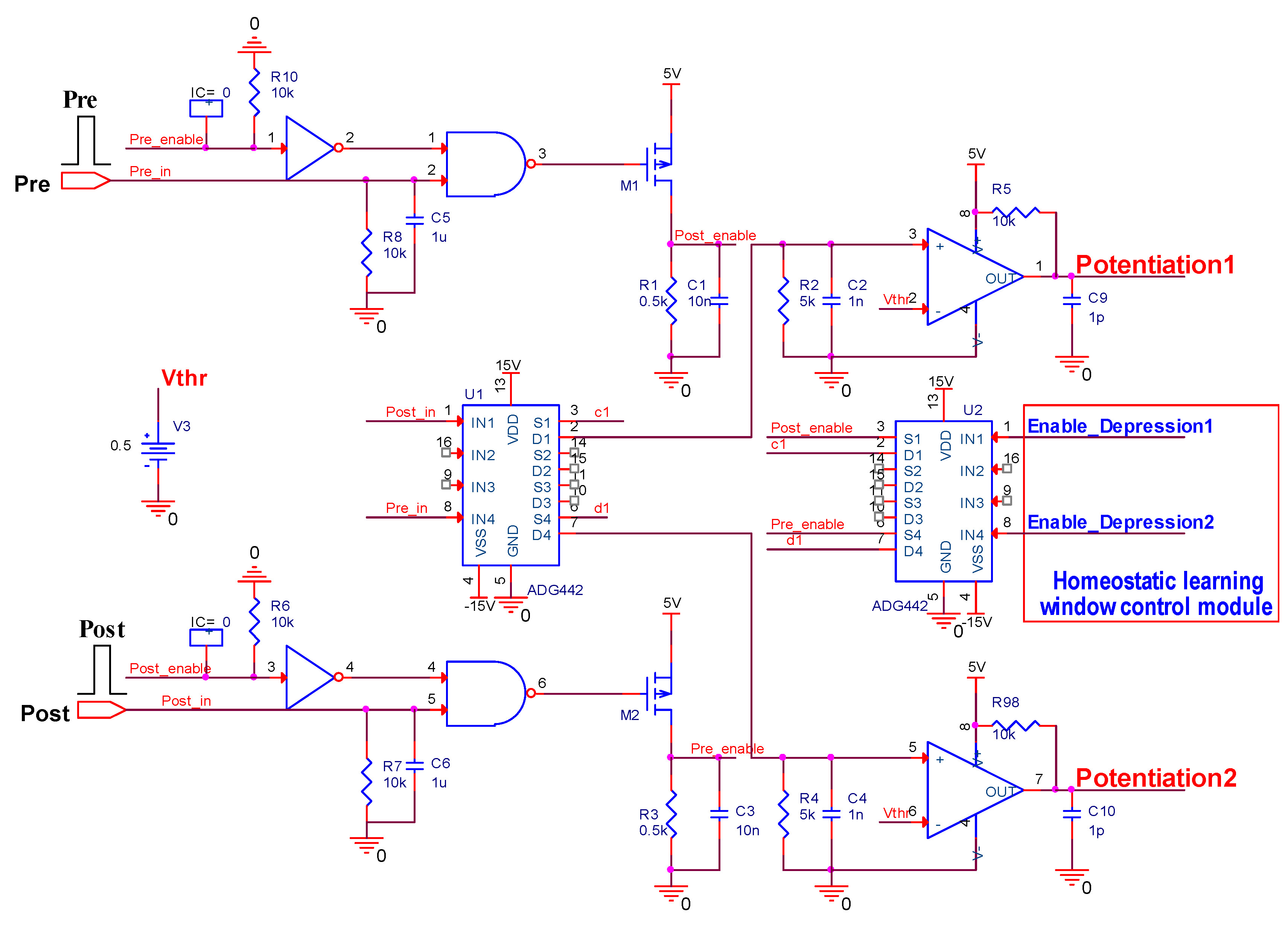
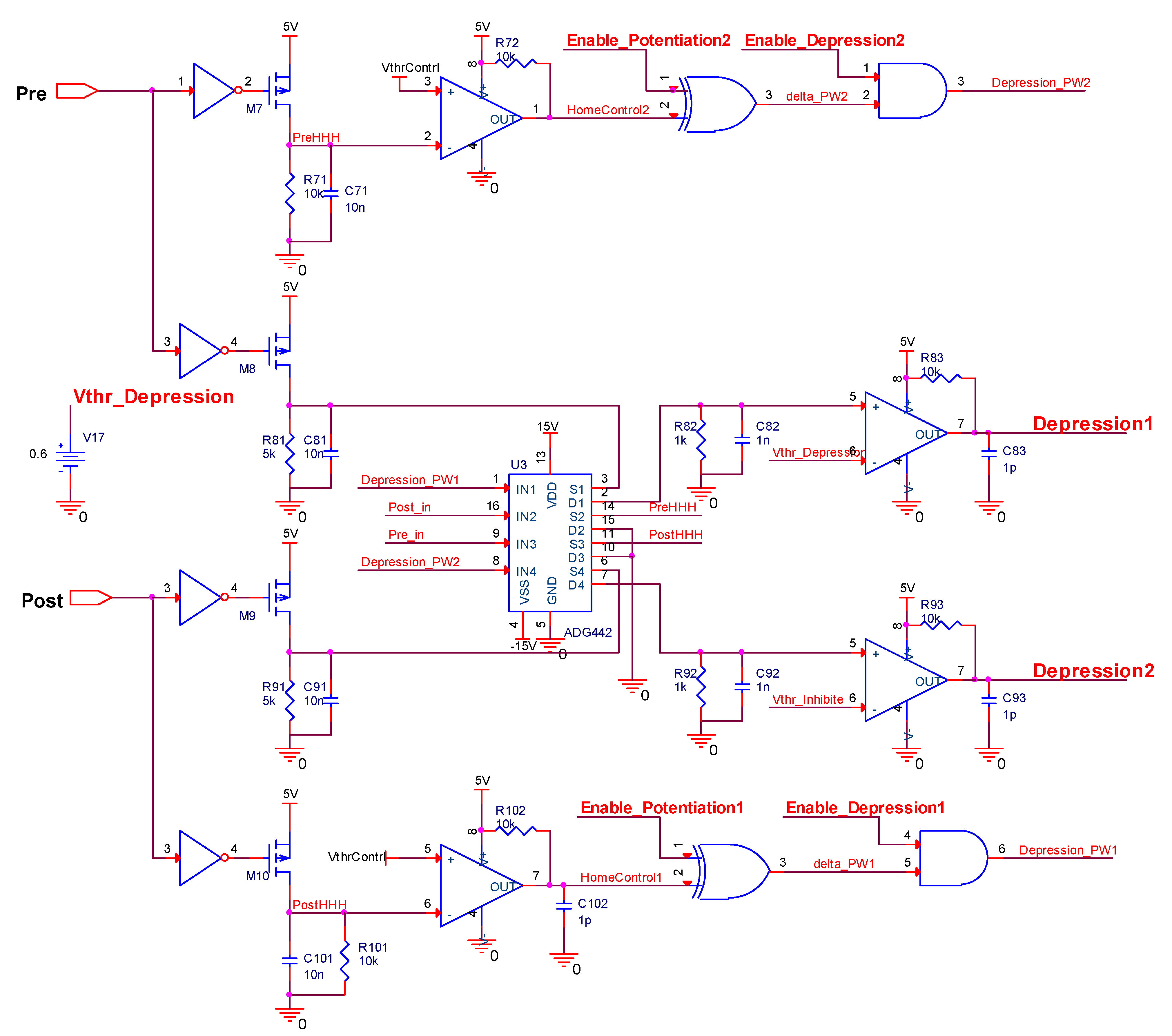
This paper focuses more on the design of a board-level homeostatic inhibitory plasticity rule demonstration circuit. The circuit contains ten MOSFETs, one Knowm memristor, thirty resistors, twenty-eight capacitors, ten inverters, four XOR gates, two AND gates, and ten comparators LM393. Since Pre and Post had high voltages, and comparators, inverters, and gate circuits required high stimulus; together with many large capacitors and resistors, the circuit consumed more power. The total power dissipation of the homeostatic inhibitory plasticity rule circuit estimated using PSpice© was 0.083 W at the displayed power supply. We must admit that this is not a small power dissipation. However, the board-level circuit design in this paper provides an idea for the integrated design of the homeostatic inhibitory plasticity rule circuit. In the nanoscale integrated circuit design field, the input pulse voltage could be appropriately reduced, and the comparator could become smaller and consume less power. Accordingly, the values of the capacitor and resistor used in the design would become smaller, which will be achievable in the near future.
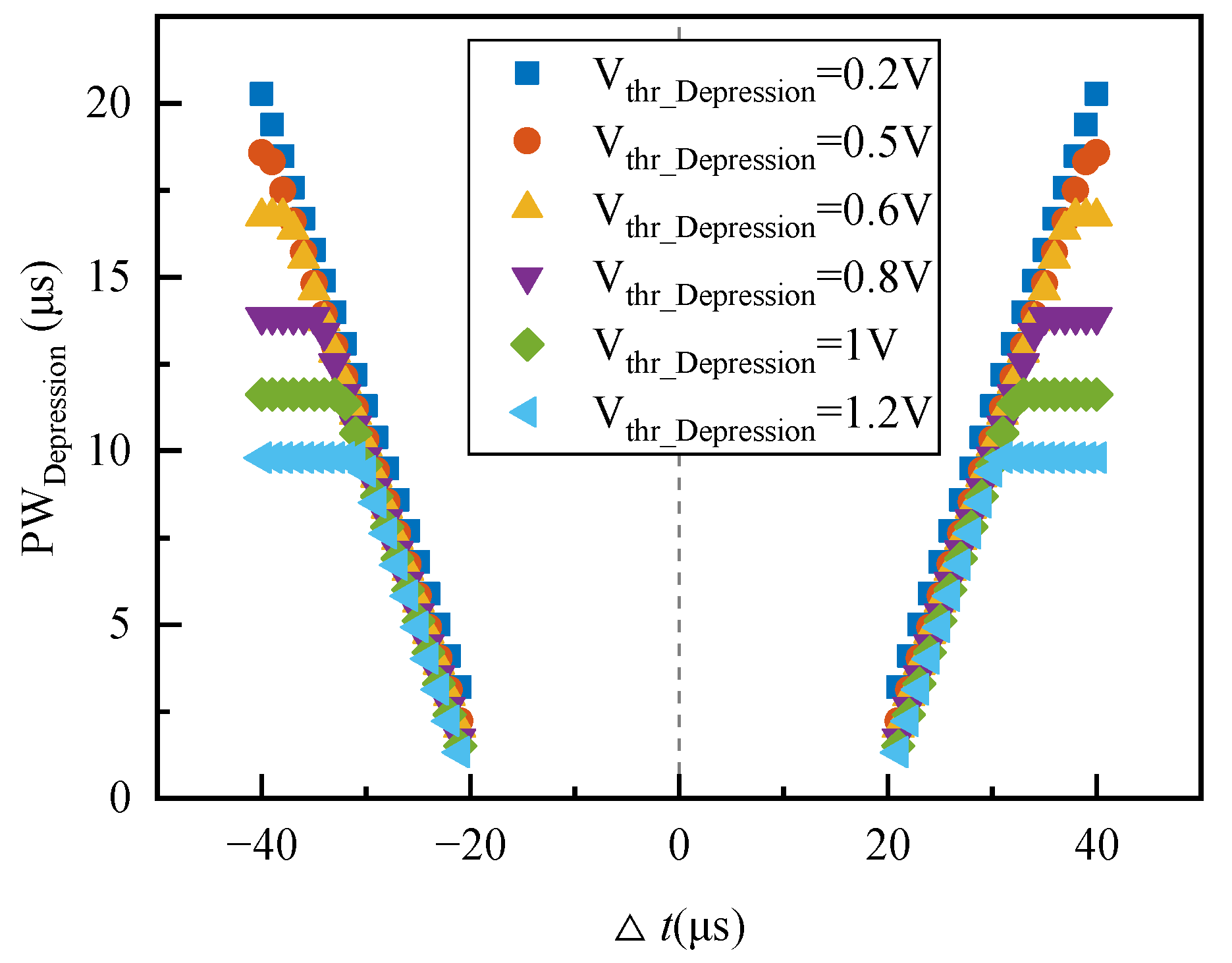
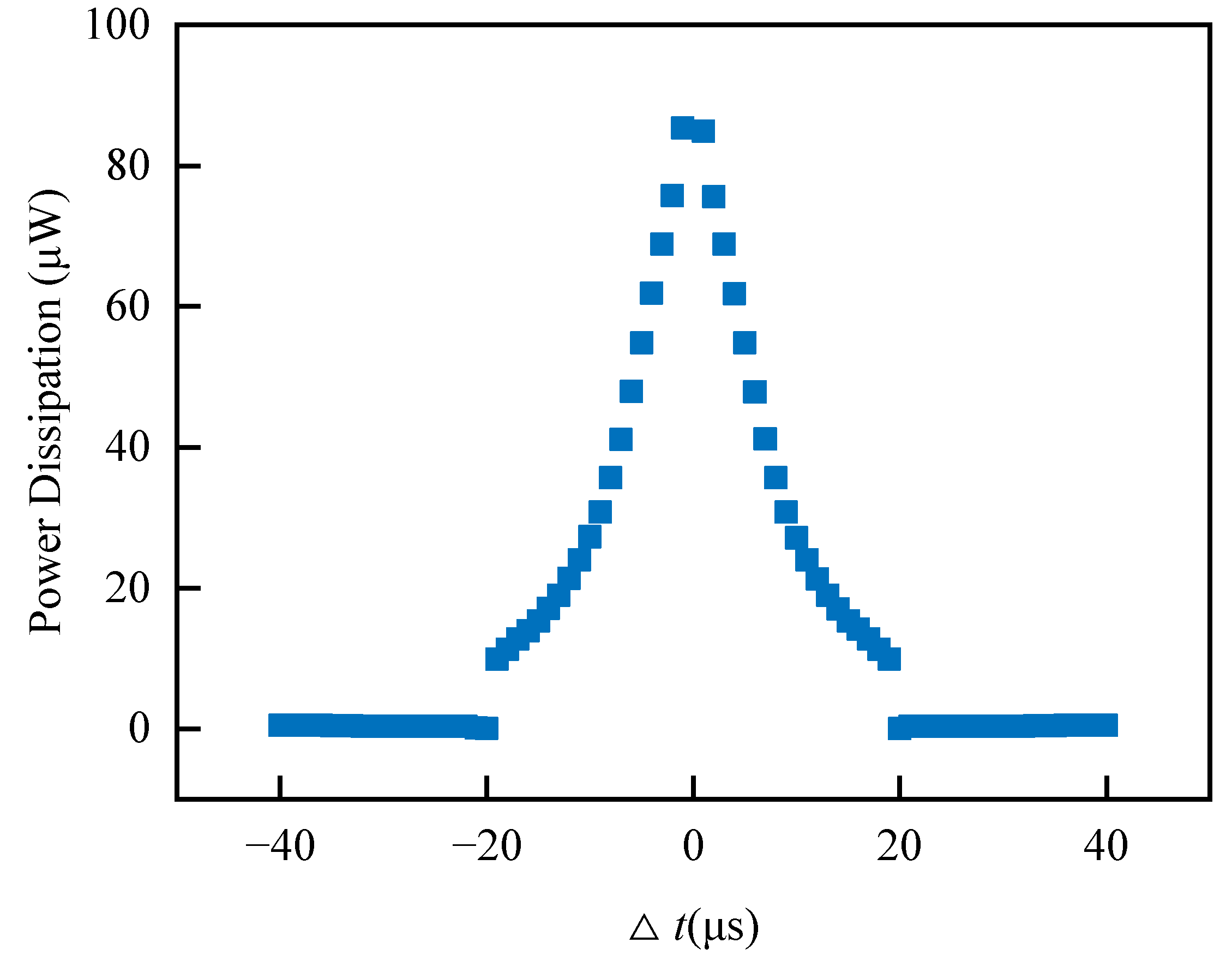
Regardless of the energy consumption of the peripheral control circuit of the memristive synapse, the power consumed by the memristive synapse depends upon the initial state of the memristive conductance, the magnitude of the memristive conductance change, and whether the memristive conductance is increasing or decreasing.
Figure 14 depicts the energy consumed by the memristive synapse as a function of the temporal difference between pre- and postsynaptic spikes with the memristive conductance initialized to 94.32 μS. Each point represents the result of a simulation of a single pair of pre- and postsynaptic spikes. The average power consumption for increasing and decreasing synaptic weights is about 35 μW and 0.34 μW, respectively. The Knowm memristive synapse consumes an average of 17.2 μW of power and 0.86 μJ of energy per spike for a spike rate of 20 MHz. Although the energy consumption of this synapse is greater than that of synapses in [
20,
25], achieving the homeostatic inhibitory plasticity rule for the first time is very meaningful based on a commercial memristor. It also suggests that reducing its power consumption will be one of the following tasks we will implement.
5. Conclusions
In this study, a fast homeostatic inhibitory plasticity rule circuit with a commercial memristor synapse is proposed and verified in PSpice© for the first time. The simulation results indicate that the circuit successfully implements the homeostatic inhibitory plasticity rule, and its time scale is one thousand times quicker than biology. This meets the requirements of neuromorphic hardware for fast computing speed. Moreover, the proposed circuit is promising for a wide range of applications with an adjustable steady-state learning window, learning rate, and steady-state factor. This research offers the groundwork for developing neuromorphic technology with homeostatic firing rates, similar to those of the biological nervous system. In addition, we believe that implementing the homeostatic inhibitory plasticity rule in the design of neuromorphic hardware can improve its reliability. This hypothesis will be verified in our upcoming study.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}













