Elevated Gene Expression of Interleukin-32 Isoforms Alpha, Beta, Gamma, and Delta in the Peripheral Blood of Chronic Psoriatic Patients


Abstract
:1. Introduction
2. Material and Methods
2.1. Patient Recruitment
2.2. Preparation of PBMCs, Reverse Transcription, and Real-Time Quantitative PCR
2.3. Statistical Analysis
3. Results
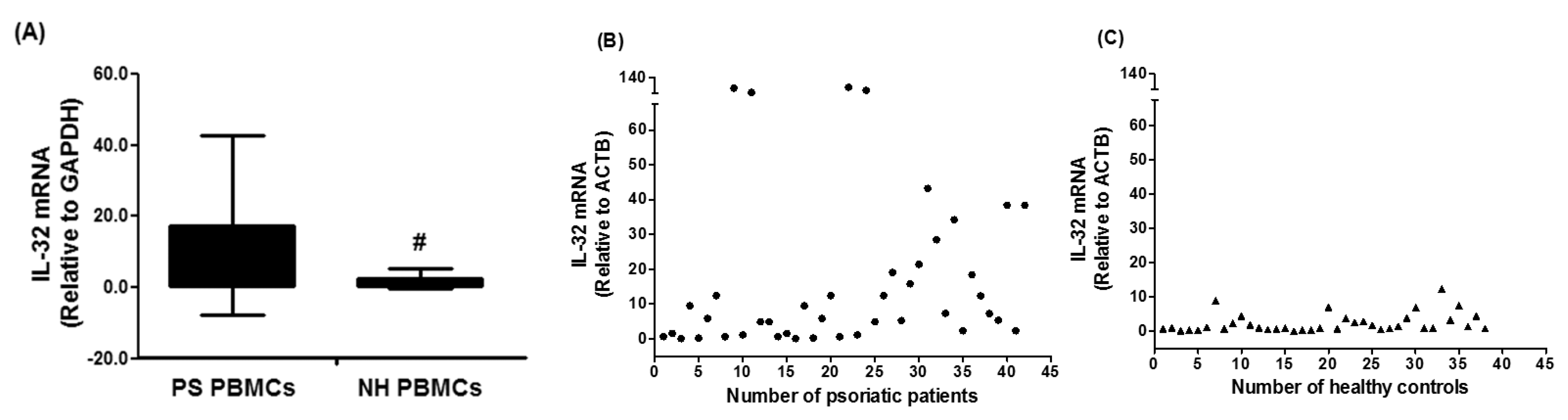
3.1. Gene Expression of IL-32 in PBMCs of Chronic Psoriatic Patients
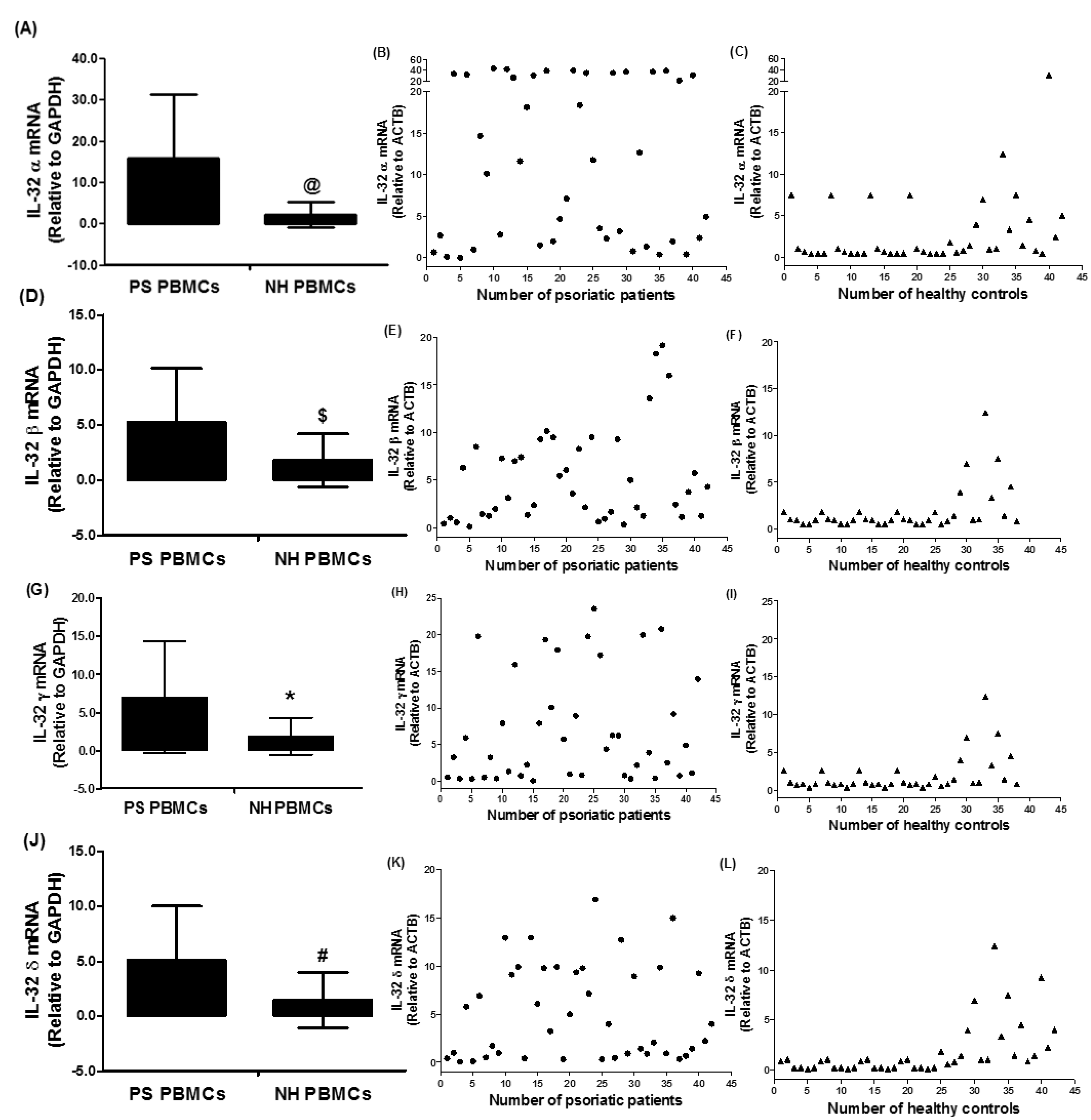
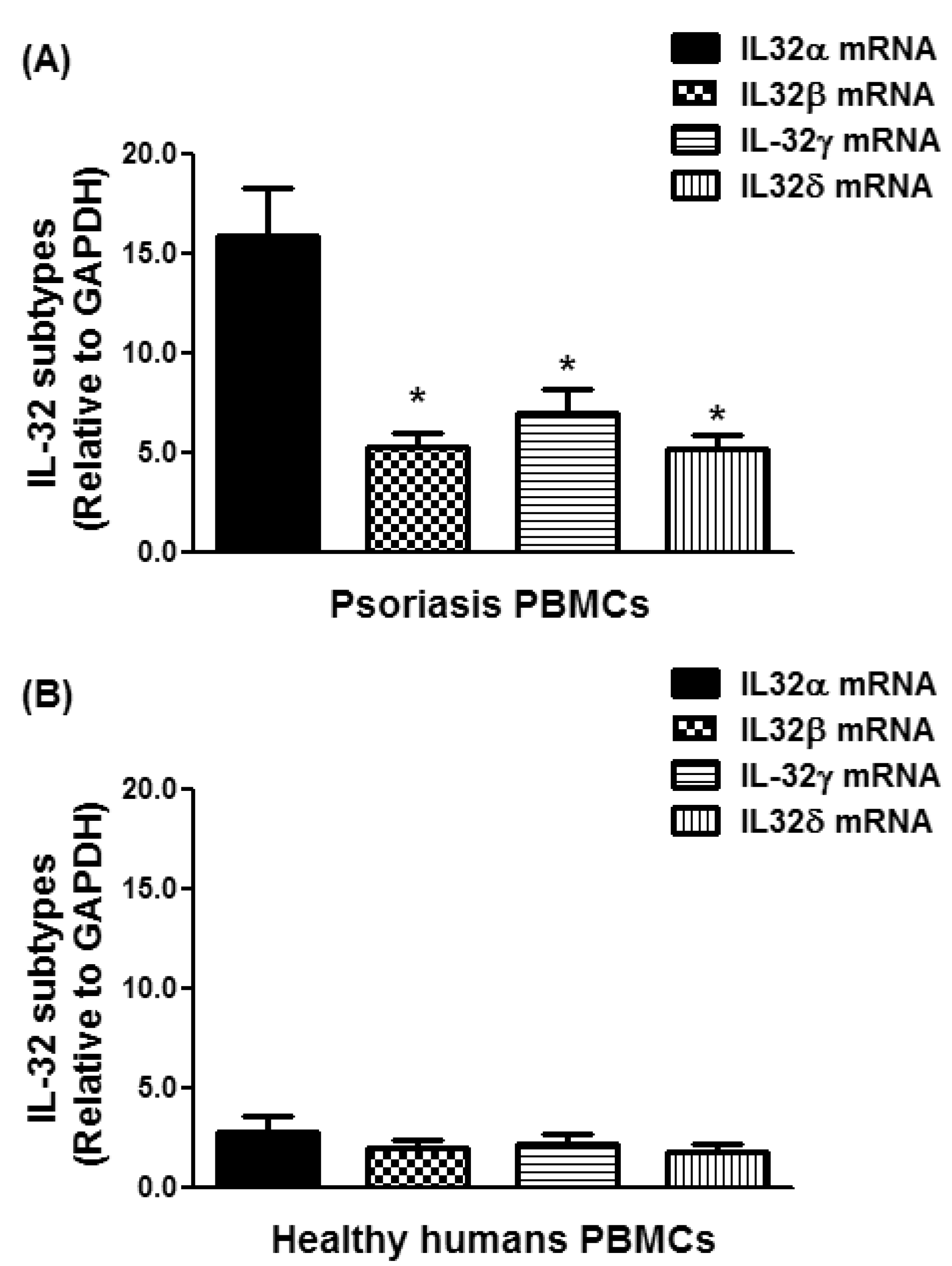
3.2. Expression of IL-32 Isoforms α, β, γ, and δ in PBMCs of Chronic Psoriatic Patients
3.3. Overexpression of IL-32α in Psoriatic Patients
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Patel, N.; Nadkarni, A.; Cardwell, L.A.; Vera, N.; Frey, C.; Patel, N.; Feldman, S.R. Psoriasis, Depression, and Inflammatory Overlap: A Review. Am. J. Clin. Dermatol. 2017, 18, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Raychaudhuri, S.K.; Maverakis, E.; Raychaudhuri, S.P. Diagnosis and classification of psoriasis. Autoimmun. Rev. 2014, 13, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.B.; Jerome, D.; Yeung, J. Diagnosis and management of psoriasis. Can. Fam. Phys. 2017, 63, 278–285. [Google Scholar]
- Wick, M.R. Psoriasiform dermatitides: A brief review. Semin. Diagn. Pathol. 2017, 34, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Tobin, A.M.; Kirby, B. TNF alpha inhibitors in the treatment of psoriasis and psoriatic arthritis. Biol. Drugs 2005, 19, 47–57. [Google Scholar]
- Kircik, L.H.; Del Rosso, J.Q. Anti-TNF agents for the treatment of psoriasis. J. Drugs Dermatol. 2009, 8, 546–559. [Google Scholar] [PubMed]
- Villadsen, L.S.; Schuurman, J.; Beurskens, F.; Dam, T.N.; Dagnaes-Hansen, F.; Skov, L.; Rygaard, J.; Voorhorst-Ogink, M.M.; Gerritsen, A.F.; van Dijk, M.A.; et al. Resolution of psoriasis upon blockade of IL-15 biological activity in a xenograft mouse model. J. Clin. Investig. 2003, 112, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Fallen, R.S.; Lima, H.C. Cytokine-based therapy in psoriasis. Clin. Rev. Allergy Immunol. 2013, 44, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Baliwag, J.; Barnes, D.H.; Johnston, A. Cytokines in psoriasis. Cytokine 2015, 73, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Kouris, A.; Pistiki, A.; Katoulis, A.; Georgitsi, M.; Giatrakou, S.; Papadavid, E.; Netea, M.G.; Stavrianeas, N.; Giamarellos-Bourboulis, E. Proinflammatory cytokine responses in patients with psoriasis. Eur. Cytokine Netw. 2014, 25, 63–68. [Google Scholar] [PubMed]
- Dinarello, C.A.; Kim, S.H. IL-32, a novel cytokine with a possible role in disease. Ann. Rheum. Dis. 2006, 65 (Suppl. 3), iii61–iii614. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Lewis, E.C.; Azam, T.; Joosten, L.A.; Jaekal, J.; Bae, S.Y.; Dinarello, C.A.; Kim, S.H. Interleukin-32 induces the differentiation of monocytes into macrophage-like cells. Proc. Natl. Acad. Sci. USA 2008, 105, 3515–3520. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Han, S.Y.; Azam, T.; Yoon, D.Y.; Dinarello, C.A. Interleukin-32: A cytokine and inducer of TNFalpha. Immunity 2005, 22, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, J.C.; Heinhuis, B.; Gomes, R.S.; Damen, M.S.; Real, F.; Mortara, R.A.; Keating, S.T.; Dinarello, C.A.; Joosten, L.A.; Ribeiro-Dias, F. Cytokines and microbicidal molecules regulated by IL-32 in THP-1-derived human macrophages infected with New World Leishmania species. PLoS Negl. Trop. Dis. 2017, 11, e0005413. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhang, S.; Pan, X.; Cao, H.; Huang, X.; Xu, Q.; Zhong, H.; Peng, X. TIMP-1 expression induced by IL-32 is mediated through activation of AP-1 signal pathway. Int. Immunopharmacol. 2016, 38, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.T.; Son, D.J.; Lee, C.K.; Yoon, D.Y.; Lee, D.H.; Park, M.H. Interleukin 32, inflammation and cancer. Pharmacol. Ther. 2017, 174, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Kim, J.A.; Park, J.H.; Park, I.H.; Han, I.H.; Lee, H.M. Toll-like receptor 4-mediated expression of interleukin-32 via the c-Jun N-terminal kinase/protein kinase B/cyclic adenosine monophosphate response element binding protein pathway in chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2016, 6, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- El-Far, M.; Kouassi, P.; Sylla, M.; Zhang, Y.; Fouda, A.; Fabre, T.; Goulet, J.P.; van Grevenynghe, J.; Lee, T.; Singer, J.; et al. Investigators of the Canadian HIV+ Slow Progressor Cohort. Proinflammatory isoforms of IL-32 as novel and robust biomarkers for control failure in HIV-infected slow progressors. Sci. Rep. 2016, 6, 22902. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Sun, Z.; Li, X.; Leng, C.; Zhang, L.; Fu, X.; Li, L.; Zhang, X.; Chang, Y.U.; Nan, F.; et al. Expression and clinical significance of cyclooxygenase-2 and interleukin-32 in primary gastric B-cell lymphoma. Oncol. Lett. 2016, 11, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Azam, T.; Ferwerda, G.; Girardin, S.E.; Walsh, M.; Park, J.S.; Abraham, E.; Kim, J.M.; Yoon, D.Y.; Dinarello, C.A.; et al. IL-32 synergizes with nucleotide oligomerization domain (NOD) 1 and NOD2 ligands for IL-1beta and IL-6 production through a caspase 1-dependent mechanism. Proc. Natl. Acad. Sci. USA 2005, 102, 16309–16314. [Google Scholar] [CrossRef] [PubMed]
- Joosten, L.A.; Netea, M.G.; Kim, S.H.; Yoon, D.Y.; Oppers-Walgreen, B.; Radstake, T.R.; Barrera, P.; van de Loo, F.A.; Dinarello, C.A.; van den Berg, W.B. IL-32, a proinflammatory cytokine in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2006, 103, 3298–3303. [Google Scholar] [CrossRef] [PubMed]
- Yousif, N.G.; Al-Amran, F.G.; Hadi, N.; Lee, J.; Adrienne, J. Expression of IL-32 modulates NF-κB and p38 MAP kinase pathways in human esophageal cancer. Cytokine 2013, 61, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, K.E.; Cheon, S.; Song, J.H.; Houh, Y.; Kim, T.S.; Gil, M.; Lee, K.J.; Kim, S.; Kim, D.; et al. Interleukin-32α induces migration of human melanoma cells through downregulation of E-cadherin. Oncotarget 2016, 7, 65825–65836. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, S.; Su, J.; Chu, G.; You, H.; Chen, Z.; Sun, H.; Chen, B.; Zhou, M. Interleukin-32α inactivates JAK2/STAT3 signaling and reverses interleukin-6-induced epithelial-mesenchymal transition, invasion, and metastasis in pancreatic cancer cells. OncoTargets Ther. 2016, 9, 4225–4237. [Google Scholar] [CrossRef] [PubMed]
- Joosten, L.A.; Heinhuis, B.; Netea, M.G.; Dinarello, C.A. Novel insights into the biology of interleukin-32. Cell. Mol. Life Sci. 2013, 70, 3883–3892. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Dias, F.; SaarGomes, R.; de LimaSilva, L.L.; DosSantos, J.C.; Joosten, L.A. Interleukin 32: A novel player in the control of infectious diseases. J. Leukoc. Biol. 2017, 101, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Gasiuniene, E.; Lavinskiene, S.; Sakalauskas, R.; Sitkauskiene, B. Levels of IL-32 in Serum, Induced Sputum Supernatant, and Bronchial Lavage Fluid of Patients with Chronic Obstructive Pulmonary Disease. COPD 2016, 13, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, N.; Otsuka, A.; Nonomura, Y.; Nakashima, C.; Honda, T.; Kabashima, K. Decrease of serum IL-32 level in patients with atopic dermatitis after cyclosporine treatment. J. Eur. Acad. Dermatol. Venereol. 2017, 31, e449–e450. [Google Scholar] [CrossRef] [PubMed]
- Thomi, R.; Yerly, D.; Yawalkar, N.; Simon, D.; Schlapbach, C.; Hunger, R.E. Interleukin-32 is highly expressed in lesions of hidradenitis suppurativa. Br. J. Dermatol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, B.; Zhao, Y.; Yu, X.; Liu, Y.; Zhang, L. Association of Plasma IL-32 Levels and Gene Polymorphisms with Systemic Lupus Erythematosus in Chinese Han Population. Dis. Markers 2016, 2016, 2460206. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Conti, P.; Vasiadi, M.; Alysandratos, K.D.; Tagen, M.; Kalogeromitros, D.; Kourelis, T.; Gregoriou, S.; Makris, M.; Stavrianeas, N.G.; et al. IL-32 is increased along with tryptase in lesional psoriatic skin and is up-regulated by substance P in human mast cells. Eur. J. Dermatol. 2010, 20, 865–867. [Google Scholar] [PubMed]
- Al-Shobaili, H.A.; Ahmed, A.A.; Rasheed, Z. Recognition of oxidized albumin and thyroid antigens by psoriasis autoantibodies. A possible role of reactive-oxygen-species induced epitopes in chronic plaque psoriasis. Saudi Med. J. 2015, 36, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Alzolibani, A.A.; Rasheed, Z.; Bin Saif, G.; Al-Dhubaibi, M.S.; Al Robaee, A.A. Altered expression of intracellular Toll-like receptors in peripheral blood mononuclear cells from patients with alopecia areata. BBA Clin. 2016, 5, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Al-Shobaili, H.A.; Rasheed, N.; Mahmood, A.; Khan, M.I. MicroRNA-26a-5p regulates the expression of inducible nitric oxide synthase via activation of NF-κB pathway in human osteoarthritis chondrocytes. Arch. Biochem. Biophys. 2016, 594, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Rasheed, N.; Al-Shobaili, H.A. Epigallocatechin-3-O-gallate up-regulates microRNA-199a-3p expression by down-regulating the expression of cyclooxygenase-2 in stimulated human osteoarthritis chondrocytes. J. Cell. Mol. Med. 2016, 20, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Haqqi, T.M. Endoplasmic reticulum stress induces the expression of COX-2 through activation of eIF2α, p38-MAPK and NF-κB in advanced glycation end products stimulated human chondrocytes. Biochim. Biophys. Acta 2012, 1823, 2179–2189. [Google Scholar] [CrossRef] [PubMed]
- Piskin, G.; Tursen, U.; Sylva-Steenland, R.M.; Bos, J.D.; Teunissen, M.B. Clinical improvement in chronic plaque-type psoriasis lesions after narrow-band UVB therapy is accompanied by a decrease in the expression of IFN-gamma inducers–IL-12, IL-18 and IL-23. Exp. Dermatol. 2004, 13, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Asadullah, K.; Sterry, W.; Stephanek, K.; Jasulaitis, D.; Leupold, M.; Audring, H.; Volk, H.D.; Docke, W.D. IL-10 is a key cytokine in psoriasis. Proof of principle by IL-10 therapy: A new therapeutic approach. J. Clin. Investig. 1998, 101, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Sa, S.M.; Valdez, P.A.; Wu, J.; Jung, K.; Zhong, F.; Hall, L.; Kasman, I.; Winer, J.; Modrusan, Z.; Danilenko, D.M.; et al. The effects of IL-20 subfamily cytokines on reconstituted human epidermis suggest potential roles in cutaneous innate defense and pathogenic adaptive immunity in psoriasis. J. Immunol. 2007, 178, 2229–2240. [Google Scholar] [CrossRef] [PubMed]
- Rutz, S.; Wang, X.; Ouyang, W. The IL-20 subfamily of cytokines–from host defence to tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.C.; Liang, W.G.; Chen, F.W.; Hsu, J.H.; Yang, J.J.; Chang, M.S. IL-19 induces production of IL-6 and TNF-alpha and results in cell apoptosis through TNF-alpha. J. Immunol. 2002, 169, 4288–4297. [Google Scholar] [CrossRef] [PubMed]
- Stenderup, K.; Rosada, C.; Worsaae, A.; Clausen, J.T.; NormanDam, T. Interleukin-20 as a target in psoriasis treatment. Ann. N. Y. Acad. Sci. 2007, 1110, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.L.; Liang, S.; Li, J.; Napierata, L.; Brown, T.; Benoit, S.; Senices, M.; Gill, D.; Dunussi-Joannopoulos, K.; Collins, M.; et al. IL-22 is required for Th17 cell-mediated pathology in a mouse model of psoriasis-like skin inflammation. J. Clin. Investig. 2008, 118, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.D.; Bae, S.Y.; Hong, J.W.; Azam, T.; Dinarello, C.A.; Her, E.; Choi, W.S.; Kim, B.K.; Lee, C.K.; Yoon, D.Y.; et al. Identification of the most active interleukin-32 isoform. Immunology 2009, 126, 535–542. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Sense | Anti-Sense |
|---|---|---|---|
| IL-32 | NM_004221 | 5′TCGCGGAGGTGGGTTTC3′ | 5′AAAACGGACTAATACGGCAACAG-3′ |
| IL-32 α | NM_001012633.1 | 5′GCTGGAGGACGACTTCAAAGA3′ | 5′GGGCTCCGTAGGACTTGTCA3′ |
| IL-32 β | NM_001012631.1 | 5′CAGTGGAGCTGGGTCATCTCA3′ | 5′GGGCCTTCAGCTTCTTCATGTCATCA3′ |
| IL-32 γ | NM_001012635.1 | 5′AGGCCCGAATGGTAATGCT3′ | 5′CCACAGTGTCCTCAGTGTCACA3′ |
| IL-32 δ | NM_001012636.1 | 5′TCTGTCTCTCTCGGGTCCTCTCT3′ | 5′TGTCTCCAGGTAGCCCTCTTTG3′ |
| GAPDH | NM_002046 | 5′TCGACAGTCAGCCGCATCTTCTTT3′ | 5′ACCAAATCCGTTGACTCCGACCTT3′ |
| ACTB | NM_001101 | 5′-AGAGCTACGAGCTGCCTGAC-3′ | 5′-AGCACTGTGTTGGCGTACAG-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Shobaili, H.A.; Rasheed, Z. Elevated Gene Expression of Interleukin-32 Isoforms Alpha, Beta, Gamma, and Delta in the Peripheral Blood of Chronic Psoriatic Patients. Diseases 2018, 6, 21. https://doi.org/10.3390/diseases6010021
Al-Shobaili HA, Rasheed Z. Elevated Gene Expression of Interleukin-32 Isoforms Alpha, Beta, Gamma, and Delta in the Peripheral Blood of Chronic Psoriatic Patients. Diseases. 2018; 6(1):21. https://doi.org/10.3390/diseases6010021
Chicago/Turabian StyleAl-Shobaili, Hani A., and Zafar Rasheed. 2018. "Elevated Gene Expression of Interleukin-32 Isoforms Alpha, Beta, Gamma, and Delta in the Peripheral Blood of Chronic Psoriatic Patients" Diseases 6, no. 1: 21. https://doi.org/10.3390/diseases6010021
APA StyleAl-Shobaili, H. A., & Rasheed, Z. (2018). Elevated Gene Expression of Interleukin-32 Isoforms Alpha, Beta, Gamma, and Delta in the Peripheral Blood of Chronic Psoriatic Patients. Diseases, 6(1), 21. https://doi.org/10.3390/diseases6010021