Inhibitory Effect of Ionizing Radiation on Echinococcus granulosus Hydatid Cyst

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Collection
2.3. Radiation Procedure
2.4. LD50 Assay
2.5. Light and Electron Microscopy
2.6. Immunoblotting
2.7. Caspase-3 Activity Assay
2.8. MtDNA Damage Assay
2.9. Statistical Analysis
3. Results
3.1. Effect of Ionizing Radiation on Hydatic Cysts Survival
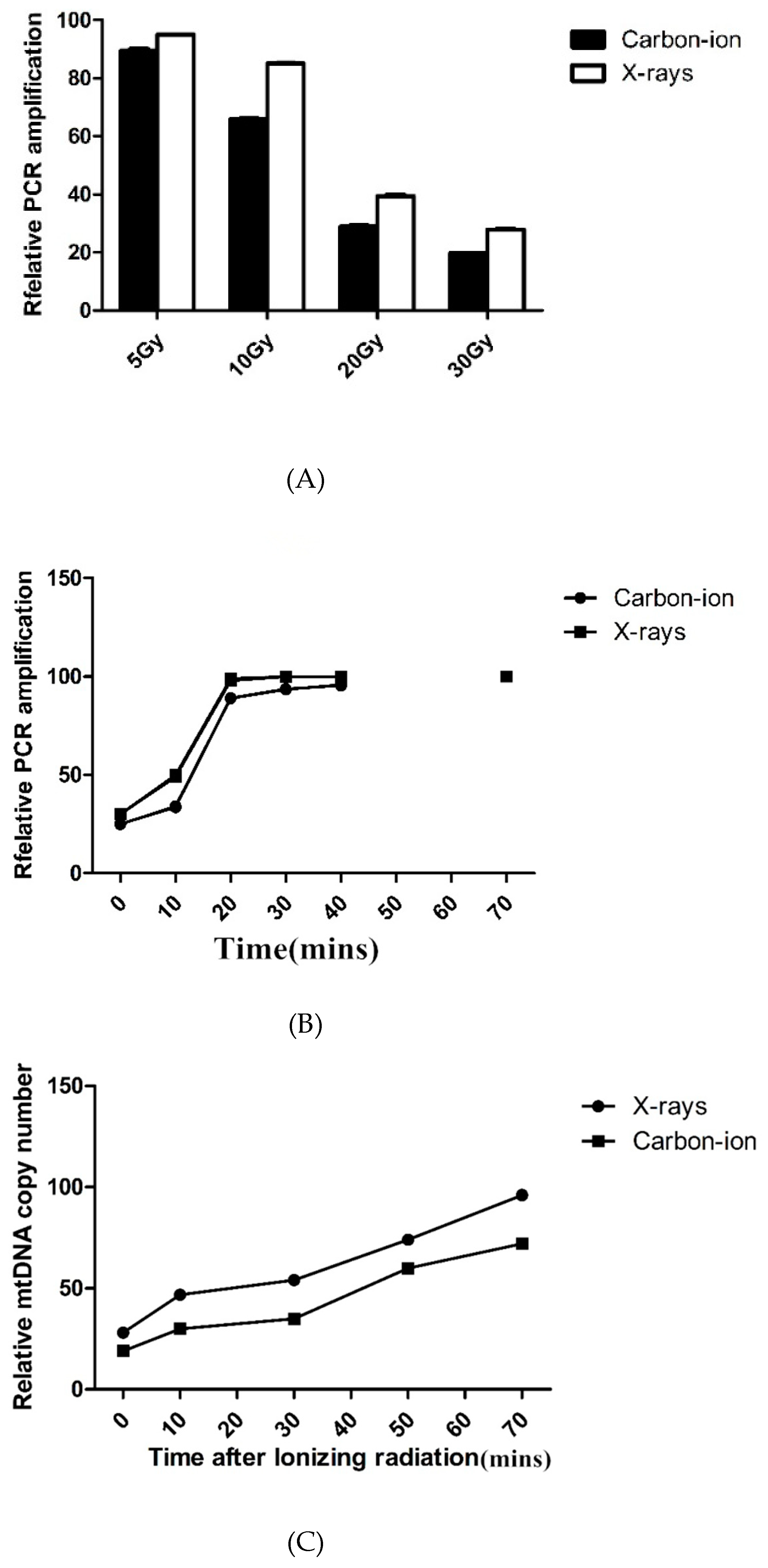
3.2. MtDNA Damage Repair and mtDNA Copy Number in Irradiated Hydatid Cysts
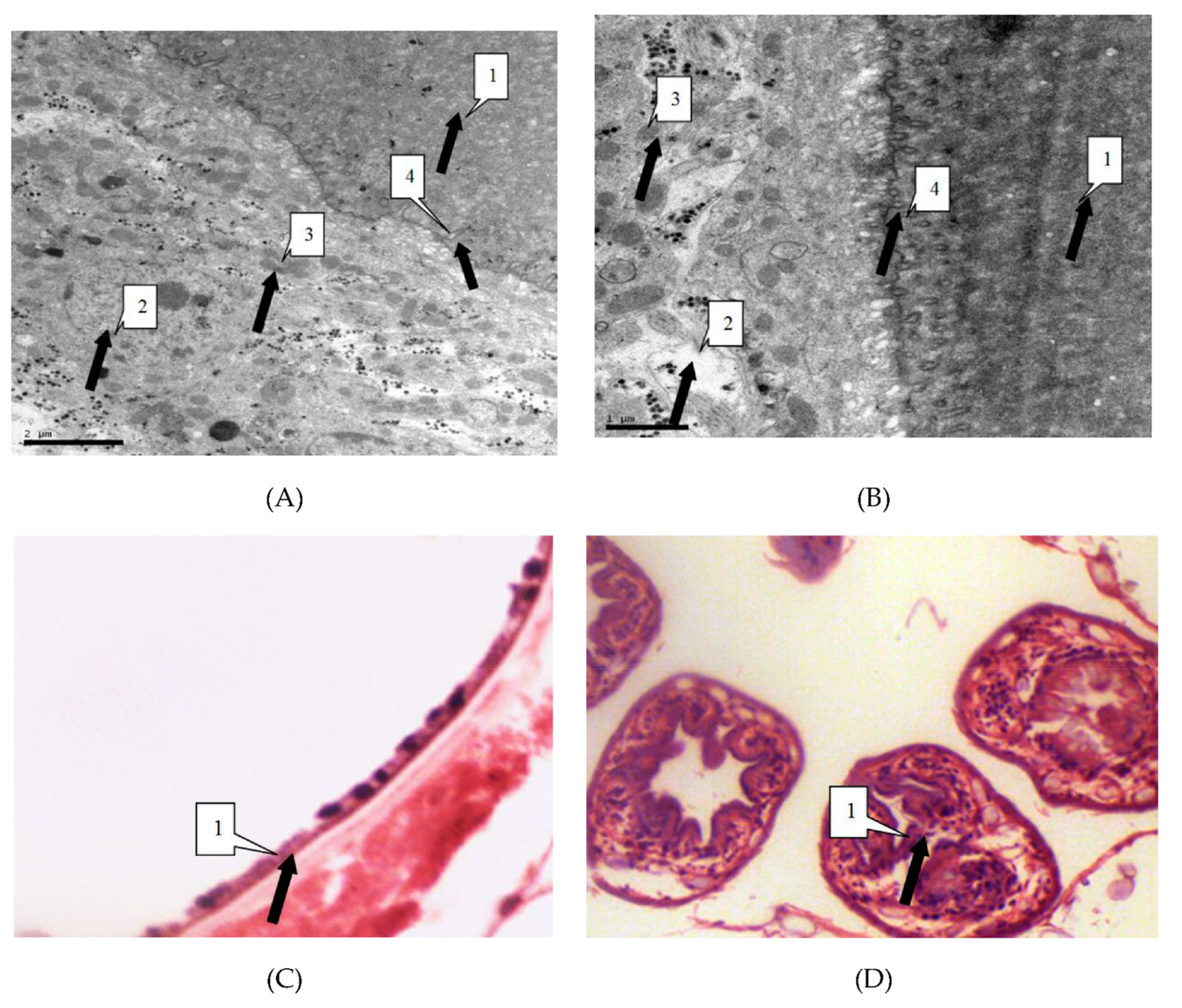
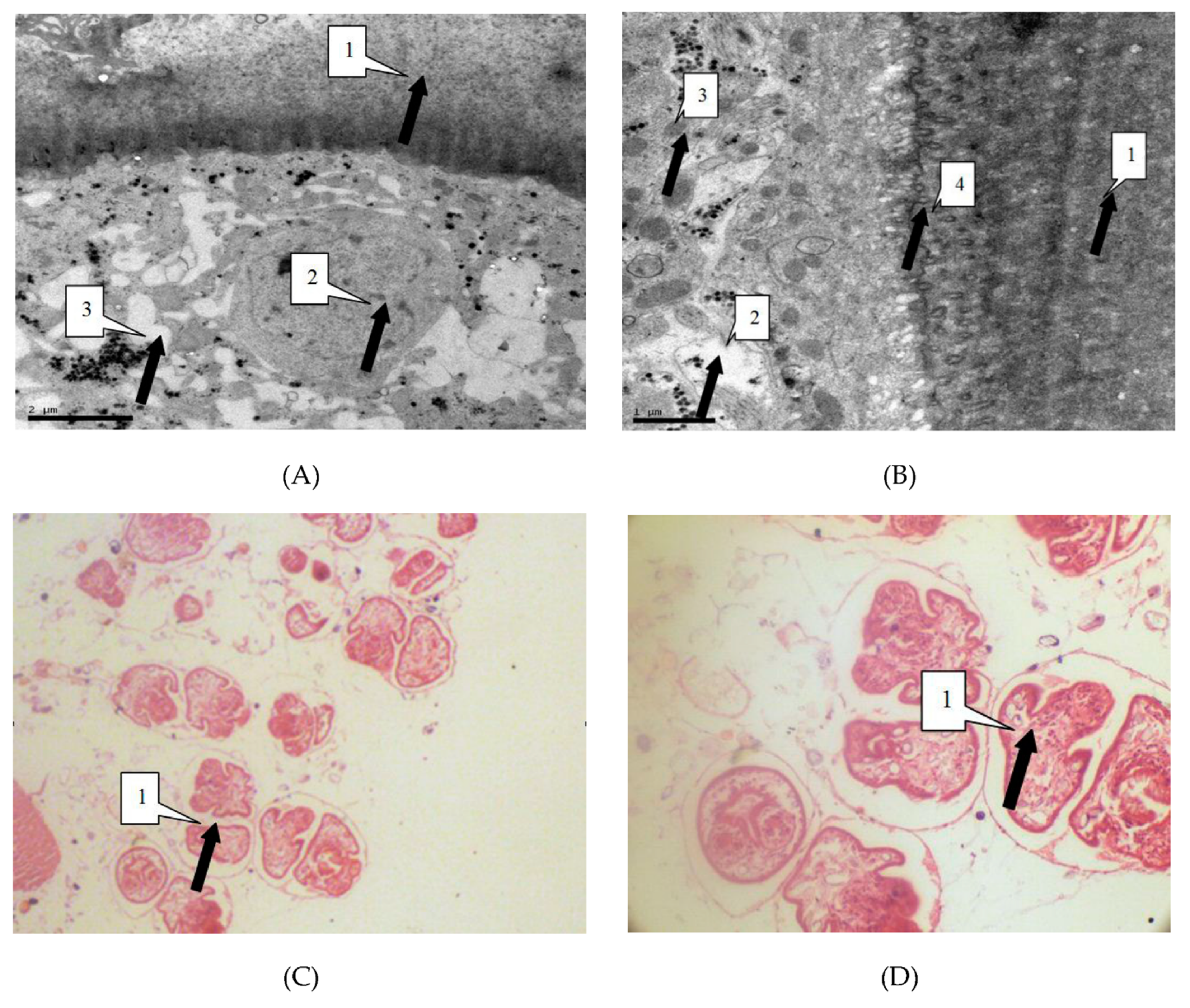
3.3. Morphological Alterations of Irradiated Parasites
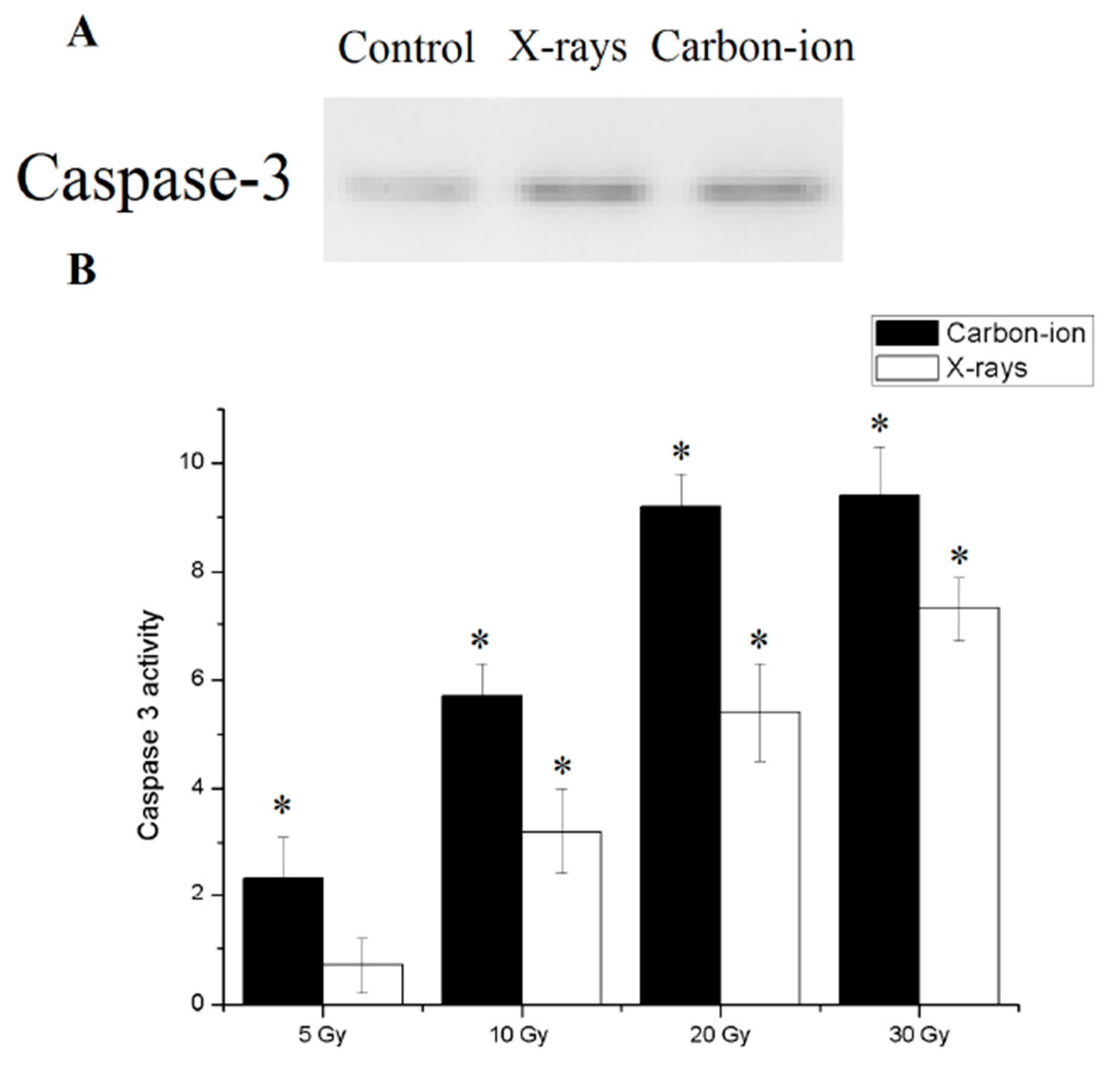
3.4. Radiation Induced Hydatid Cysts Apoptosis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eckert, J.; Deplazes, P. Biological, epidemiological, and clinical aspects of echinococcosis, a zoonosis of increasing concern. J. Clin. Microbiol. Rev. 2004, 17, 107–135. [Google Scholar] [CrossRef]
- Bao, Y.X.; Zhang, Y.F.; Ni, Y.Q.; Xie, Z.R.; Qi, H.Z.; Mao, R.; Yang, Y.G.; Wen, H. Effect of 6 MeVradiotherapy on secondary Echinococcus multilocularis infection in rats. Chin. J. Parasitol. Parasit. Dis. 2011, 29, 127–129. [Google Scholar]
- Bao, Y.X.; Mao, R.; Qi, H.Z.; Zhang, Y.F.; Ni, Y.Q.; Xie, Z.R.; Aziguli, T.; Wen, H. X-ray irradiationagainst Echinococcus multilocularis protoscoleces in vitro. Chin. J. Parasitol. Parasit. Dis. 2011, 29, 208–211. [Google Scholar]
- Ulger, S.; Barut, H.; Tunc, M.; Aydin, E.; Aydinkarahaliloglu, E.; Gokcek, A.; Karaoğlanoğlu, N. Radiationtherapy for resistant sternal hydatid disease. Strahlenther. Onkol. 2013, 189, 508–509. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.; Murikami, K. Preliminary neoplasms of the liver. Results of radiation therapy. Cancer 1960, 13, 714–720. [Google Scholar] [CrossRef]
- Stillwagon, G.B.; Order, S.E.; Guse, C.; Klein, J.L.; Leichner, P.K.; Leibel, S.A.; Fishman, E.K. 194hepatocellular cancers treated by radiation and chemotherapy combinations: Toxicity and response: A Radiation Therapy Oncology Group Study. Int. J. Radiat. Oncol. Biol. Phys. 1989, 17, 1223–1229. [Google Scholar] [CrossRef]
- Movsas, B.; Raffin, T.A.; Epstein, A.H.; Link, C.J., Jr. Pulmonary radiation injury. Chest 1997, 111, 1061–1076. [Google Scholar] [CrossRef] [PubMed]
- Hamada, N.; Imaoka, T.; Masunaga, S.; Ogata, T.; Okayasu, R.; Takahashi, A.; Kato, T.A.; Kobayashi, Y.; Ohnishi, T.; Ono, K.; et al. Recent advances in the biology of heavy-ion cancer therapy. J. Radiat. Res. 2010, 51, 365–383. [Google Scholar] [CrossRef] [PubMed]
- Imada, H.; Kato, H.; Yasuda, S.; Yamada, S.; Yanagi, T.; Kishimoto, R.; Kandatsu, S.; Mizoe, J.E.; Kamada, T.; Yokosuka, O.; et al. Comparison of efficacy and toxicity of short-course carbon ion radiotherapy for hepatocellular carcinoma depending on their proximity to the porta hepatis. Radiother. Oncol. 2010, 96, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Kang, J.; Chen, R.; Mamuti, W.; Wu, G.; Yuan, W.M. Drug-induced apoptosis of Echinococcus granulosus protoscoleces. Parasitol. Res. 2011, 109, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Elissondo, M.C.; Ceballos, L.; Alvarez, L.; Sanchez Bruni, S.; Lanusse, C.; Denegri, G. Flubendazole and ivermectin in vitro combination therapy produces a marked effect on Echinococcus granulosus protoscoleces and metacestodes. Parasitol. Res. 2009, 105, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Yakes, F.M.; Van Houten, B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells followingoxidative stress. Proc. Natl. Acad. Sci. USA. 1997, 94, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Sykora, P.; Wilson, D.M. Repair of persistent strand breaks in the mitochondrial genome. Mech. Ageing. Dev. 2012, 133, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Goto, S.; Kawakatsu, M.; Urata, Y.; Li, T.S. Mitochondrial dysfunction, a probable cause of persistent oxidative stress after exposure to ionizing radiation. Free Radic. Res. 2012, 46, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Paredes, R.; Jimenez, V.; Cabrera, G.; Iraguen, D.; Galanti, N. Apoptosis as apossible mechanism of infertility in Echinococcus granulosus hydatid cysts. J. Cell. Biochem. 2007, 100, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Spotin, A.; Majdi, M.M.; Sankian, M.; Varasteh, A. The study of apoptotic bifunctional effects in relationship between host and parasite in cystic echinococcosis: a new approach to suppression and survival of hydatid cyst. Parasitol. Res. 2012, 110, 1979–1984. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari Amirmajdi, M.; Sankian, M.; Eftekharzadeh Mashhadi, I.; Varasteh, A.; Vahedi, F.; Sadrizadeh, A.; Spotin, A. Apoptosis of human lymphocytes after exposure tohydatid fluid. Iran J. Parasitol. 2011, 6, 9–16. [Google Scholar]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef]
- Schulz-Ertner, D.; Nikoghosyan, A.; Thilmann, C.; Haberer, T.; Jakel, O.; Karger, C.; Kraft, G.; Wannenmacher, M.; Debus, J. Results of carbon ion radiotherapy in 152 patients. Int. J. Radiat. Oncol. Biol. Phys. 2004, 58, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Tsujii, H.; Mizoe, J.E.; Kamada, T.; Baba, M.; Kato, S.; Kato, H.; Tsuji, H.; Yamada, S.; Yasuda, S.; Ohno, T.; et al. Overview of clinical experiences on carbon ion radiotherapy at NIRS. Radiother. Oncol. 2004, 73 (Suppl. S2), S41–S49. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Gui, W.; Zhang, Y.; Mo, G.; Li, D.; Chong, S. Inhibitory Effect of Ionizing Radiation on Echinococcus granulosus Hydatid Cyst. Diseases 2019, 7, 23. https://doi.org/10.3390/diseases7010023
Zhao Y, Gui W, Zhang Y, Mo G, Li D, Chong S. Inhibitory Effect of Ionizing Radiation on Echinococcus granulosus Hydatid Cyst. Diseases. 2019; 7(1):23. https://doi.org/10.3390/diseases7010023
Chicago/Turabian StyleZhao, Yumin, Weifeng Gui, Yishu Zhang, Gang Mo, Dayu Li, and Shigui Chong. 2019. "Inhibitory Effect of Ionizing Radiation on Echinococcus granulosus Hydatid Cyst" Diseases 7, no. 1: 23. https://doi.org/10.3390/diseases7010023
APA StyleZhao, Y., Gui, W., Zhang, Y., Mo, G., Li, D., & Chong, S. (2019). Inhibitory Effect of Ionizing Radiation on Echinococcus granulosus Hydatid Cyst. Diseases, 7(1), 23. https://doi.org/10.3390/diseases7010023