Are Steroid Hormones Dysregulated in Autistic Girls?

Abstract
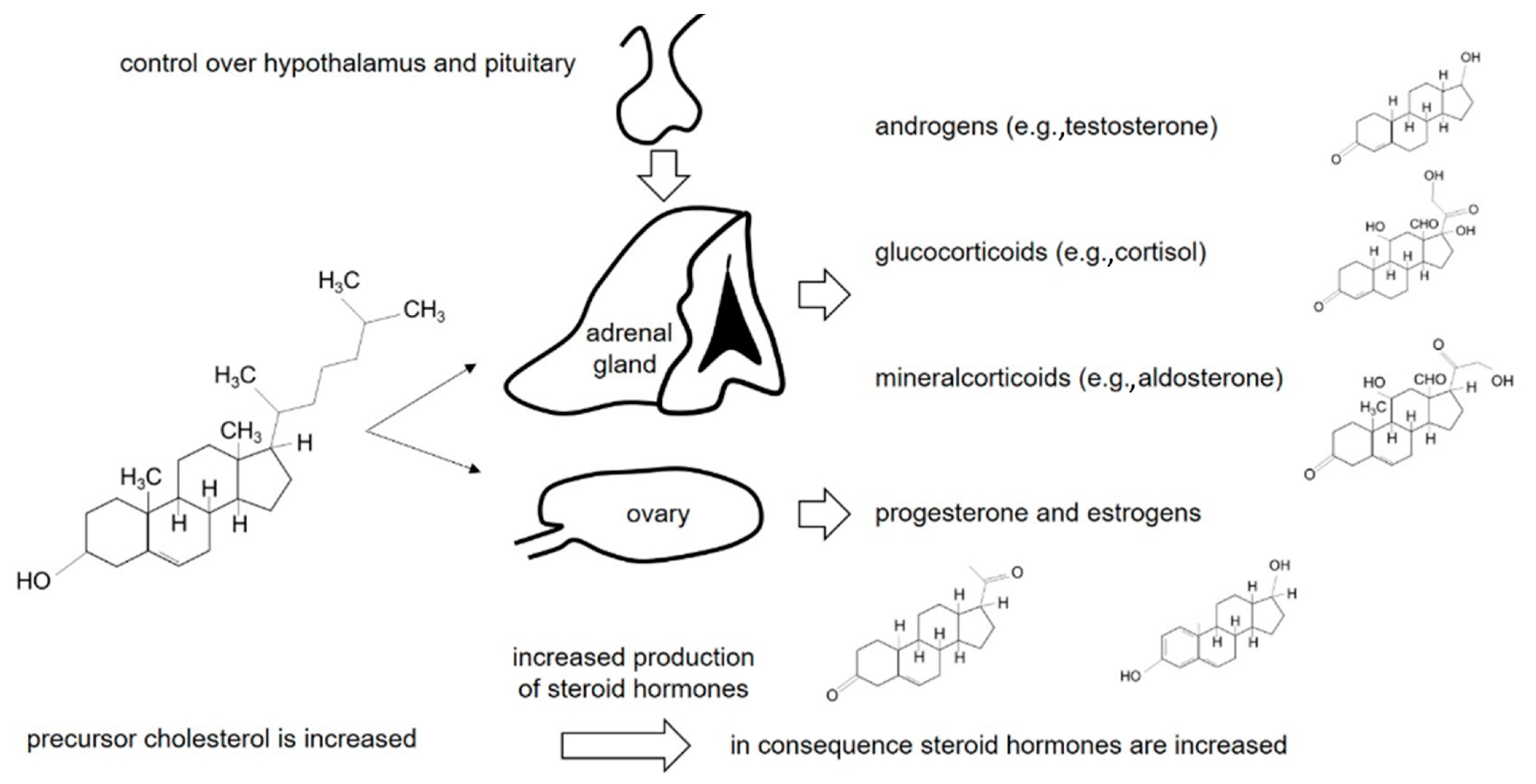
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Methods
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Spratt, E.; Nicholas, J.S.; Brady, K.T.; Carpenter, L.; Hatcher, C.R.; Meekins, K.A.; Furlanetto, R.W.; Charles, J.M. Enhanced Cortisol Response to Stress in Children in Autism. J. Autism Dev. Disord. 2012, 42, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Hollocks, M.; Howlin, P.; Papadopoulos, A.S.; Khondoker, M.; Simonoff, E. Differences in HPA-axis and heart rate responsiveness to psychosocial stress in children with autism spectrum disorders with and without co-morbid anxiety. Psychoneuroendocrinology 2014, 46, 32–45. [Google Scholar] [CrossRef]
- Jacobson, L. Hypothalamic-Pituitary-Adrenocortical Axis: Neuropsychiatric Aspects. Compr. Physiol. 2014, 4, 715–738. [Google Scholar] [PubMed]
- Taylor, J.L.; Corbett, B.A. A review of rhythm and responsiveness of cortisol in individuals with autism spectrum disorders. Psychoneuroendocrinology 2014, 49, 207–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tani, P.; Lindberg, N.; Matto, V.; Appelberg, B.; Wendt, T.N.-V.; Von Wendt, L.; Porkka-Heiskanen, T. Higher plasma ACTH levels in adults with Asperger syndrome. J. Psychosom. Res. 2005, 58, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.; Gillberg, C. The Autisms, 4th ed.; Oxford University Press: New York, NY, USA, 2012. [Google Scholar]
- Gillberg, C.; Fernell, E.; Kočovská, E.; Minnis, H.; Bourgeron, T.; Thompson, L.; Allely, C.S. The role of cholesterol metabolism and various steroid abnormalities in autism spectrum disorders: A hypothesis paper. Autism Res. 2017, 10, 1022–1044. [Google Scholar] [CrossRef]
- Gillberg, C. Debate and Argument: Is Autism a Pervasive Developmental Disorder? J. Child Psychol. Psychiatry 1991, 32, 1169–1170. [Google Scholar] [CrossRef]
- Bejerot, S.; Eriksson, J. Sexuality and Gender Role in Autism Spectrum Disorder: A Case Control Study. PLoS ONE 2014, 9, e87961. [Google Scholar] [CrossRef]
- Posserud, M.B.; Lundervold, A.J.; Gillberg, C. Autistic features in a total population of 7–9-year-old children assessed by the ASSQ (Autism Spectrum Screening Questionnaire). J. Child Psychology Psychiatry 2006, 47, 167–175. [Google Scholar] [CrossRef]
- Crider, A.; Thakkar, R.; Ahmed, A.O.; Pillai, A. Dysregulation of estrogen receptor beta (ERβ), aromatase (CYP19A1), and ER co-activators in the middle frontal gyrus of autism spectrum disorder subjects. Mol. Autism 2014, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Wing, L. Sex ratios in early childhood autism and related conditions. Psychiatry Res. 1981, 5, 129–137. [Google Scholar] [CrossRef]
- Bitsika, V.; Sharpley, C.; Sweeney, J.A.; McFarlane, J.R. HPA and SAM axis responses as correlates of self- vs parental ratings of anxiety in boys with an Autistic Disorder. Physiol. Behav. 2014, 127, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kočovská, E.; Fernell, E.; Billstedt, E.; Minnis, H.; Gillberg, C. Vitamin D and autism: Clinical review. Res. Dev. Disabil. 2012, 33, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Kocovska, E.; Andosdottir, G.; Weihe, P.; Halling, J.; Fernell, E.; Stóra, T.; Biskupstø, R.; Gillberg, I.; Shea, R.L.; Billstedt, E.; et al. Vitamin d in the general population of young adults with autism in the faroe islands. J. Autism Dev. Disord. 2014, 44, 2996–3005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernell, E.; Bejerot, S.; Westerlund, J.; Miniscalco, C.; Simila, H.; Eyles, D.W.; Gillberg, C.; Humble, M.B. Autism spectrum disorder and low vitamin D at birth: A sibling control study. Mol. Autism 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Hamza, R.T.; Hewedi, D.H.; Ismail, M.A. Basal and Adrenocorticotropic Hormone Stimulated Plasma Cortisol Levels Among Egyptian Autistic Children: Relation to Disease Severity. Ital. J. Pediatr. 2010, 36, 71. [Google Scholar] [CrossRef] [Green Version]
- Brosnan, M.; Turner-Cobb, J.; Munro-Naan, Z.; Jessop, D. Absence of a normal Cortisol Awakening Response (CAR) in adolescent males with Asperger Syndrome (AS). Psychoneuroendocrinology 2009, 34, 1095–1100. [Google Scholar] [CrossRef]
- Marinović-Ćurin, J.; Marinović-Terzić, I.; Bujas-Petković, Z.; Zekan, L.; Škrabić, V.; Đogaš, Z.; Terzić, J. Slower cortisol response during ACTH stimulation test in autistic children. Eur. Child Adolesc. Psychiatry 2007, 17, 39–43. [Google Scholar] [CrossRef]
- Hoshino, Y.; Ohno, Y.; Murata, S.; Yokoyama, F.; Kaneko, M.; Kumashiro, H. Dexamethasone Suppression Test in Autistic Children. Psychiatry Clin. Neurosci. 1984, 38, 445–449. [Google Scholar] [CrossRef]
- Hoshino, Y.; Yokoyama, F.; Watanabe, M.; Murata, S.; Kaneko, M.; Kumashiro, H. The Diurnal Variation and Response to Dexamethasone Suppression Test of Saliva Cortisol Level in Autistic Children. Psychiatry Clin. Neurosci. 1987, 41, 227–235. [Google Scholar] [CrossRef]
- Curin, J.M.; Terzić, J.; Petković, Z.B.; Zekan, L.; Terzić, I.M.; Susnjara, I.M. Lower cortisol and higher ACTH levels in individuals with autism. J. Autism Dev. Disord. 2003, 33, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Iwata, K.; Matsuzaki, H.; Miyachi, T.; Shimmura, C.; Suda, S.; Tsuchiya, K.J.; Matsumoto, K.; Suzuki, K.; Iwata, Y.; Nakamura, K.; et al. Investigation of the serum levels of anterior pituitary hormones in male children with autism. Mol. Autism 2011, 2, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.-J.; Shou, X.-J.; Li, J.; Jia, M.-X.; Zhang, J.-S.; Guo, Y.; Wei, Q.-Y.; Zhang, X.-T.; Han, S.-P.; Zhang, R.; et al. Mothers of Autistic Children: Lower Plasma Levels of Oxytocin and Arg-Vasopressin and a Higher Level of Testosterone. PLoS ONE 2013, 8, e74849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron-Cohen, S.; Auyeung, B.; Nørgaard-Pedersen, B.; Hougaard, D.M.; Abdallah, M.W.; Melgaard, L.; Cohen, A.S.; Chakrabarti, B.; Ruta, L.; Lombardo, M.V. Elevated fetal steroidogenic activity in autism. Mol. Psychiatry 2014, 20, 369–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Baz, F.; Hamza, R.T.; Ayad, M.S.; Mahmoud, N.H. Hyperandrogenemia in male autistic children and adolescents: Relation to disease severity. Int. J. Adolesc. Med. Heal. 2014, 26, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Ruta, L.; Ingudomnukul, E.; Taylor, K.; Chakrabarti, B.; Baron-Cohen, S. Increased serum androstenedione in adults with autism spectrum conditions. Psychoneuroendocrinology 2011, 36, 1154–1163. [Google Scholar] [CrossRef]
- Ingudomnukul, E.; Baron-Cohen, S.; Wheelwright, S.J.; Knickmeyer, R. Elevated rates of testosterone-related disorders in women with autism spectrum conditions. Horm. Behav. 2007, 51, 597–604. [Google Scholar] [CrossRef]
- Knickmeyer, R.; Baron-Cohen, S.; Fane, B.A.; Wheelwright, S.J.; Mathews, G.A.; Conway, G.S.; Brook, C.G.; Hines, M. Androgens and autistic traits: A study of individuals with congenital adrenal hyperplasia. Horm. Behav. 2006, 50, 148–153. [Google Scholar] [CrossRef]
- Majewska, M.D.; Hill, M.; Urbanowicz, E.; Rok-Bujko, P.; Bieńkowski, P.; Namysłowska, I.; Mierzejewski, P. Marked elevation of adrenal steroids, especially androgens, in saliva of prepubertal autistic children. Eur. Child Adolesc. Psychiatry 2013, 23, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Takagishi, H.; Takahashi, T.; Yamagishi, T.; Shinada, M.; Inukai, K.; Tanida, S.; Mifune, N.; Horita, Y.; Hashimoto, H.; Yang, Y.; et al. Salivary testosterone levels and autism-spectrum quotient in adults. Neuro Endocrinol. Lett. 2010, 31, 837–841. [Google Scholar]
- Bradstreet, J.J.; Smith, S.; Granpeesheh, R.; El-Dahr, J.M.; Rossignol, D. Spironolactone might be a desirable immunologic and hormonal intervention in autism spectrum disorders. Med. Hypotheses 2007, 68, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Kolkhof, P.; Bärfacker, L. 30 YEARS OF THE MINERALOCORTICOID RECEPTOR: Mineralocorticoid receptor antagonists: 60 years of research and development. J. Endocrinol. 2017, 234, T125–T140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beggiato, A.; Peyre, H.; Maruani, A.; Scheid, I.; Rastam, M.; Amsellem, F.; Gillberg, C.I.; Leboyer, M.; Bourgeron, T.; Gillberg, C.; et al. Gender differences in autism spectrum disorders: Divergence among specific core symptoms. Autism Res. 2016, 10, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Hindström, S. The knowledge about autism in girls must increase. Lakartidningen 2017, 114. [Google Scholar]
- Popper, K.R. Logik der Forschung. Tübingen: Mohr Siebeck; Original Work Published: Tuebingen, Germany, 1934. [Google Scholar]
- Aufdenblatten, M.; Baumann, M.; Raio, L.; Dick, B.; Frey, B.M.; Schneider, H.; Surbek, D.; Hocher, B.; Mohaupt, M. Prematurity Is Related to High Placental Cortisol in Preeclampsia. Pediatr. Res. 2009, 65, 198–202. [Google Scholar] [CrossRef] [Green Version]
- Henschkowski, J.; Stuck, A.E.; Frey, B.M.; Gillmann, G.; Dick, B.; Frey, F.J.; Mohaupt, M.G. Age-dependent decrease in 11β-hydroxysteroid dehydrogenase type 2 (11β-HSD2) activity in hypertensive patients. Am. J. Hypertens. 2008, 21, 644–649. [Google Scholar] [CrossRef] [Green Version]
- Krone, N.; Hughes, B.A.; Lavery, G.G.; Stewart, P.M.; Arlt, W.; Shackleton, C.H. Gas chromatography/mass spectrometry (GC/MS) remains a pre-eminent discovery toOl in clinical steroid investigations even in the era of fast liquid chromatography tandem mass spectrometry (LC/MS/MS). J Steroid Biochem. Mol. Biol. 2010, 121, 496–504. [Google Scholar] [CrossRef]
- Brämswig, J.; Dübbers, A. Disorders of Pubertal Development. Dtsch. Aerzteblatt Online 2009, 106, 295–304. [Google Scholar] [CrossRef]
- Brix, N.; Ernst, A.; Lauridsen, L.L.B.; Parner, E.; Støvring, H.; Olsen, J.; Henriksen, T.B.; Ramlau-Hansen, C.H. Timing of puberty in boys and girls: A population-based study. Paediatr. Périnat. Epidemiol. 2018, 33, 70–78. [Google Scholar] [CrossRef]
- Tordjman, S.; Anderson, G.M.; McBride, P.A.; Hertzig, M.E.; Snow, M.E.; Hall, L.M.; Ferrari, P.; Cohen, D.J. Plasma androgens in autism. J. Autism Dev. Disord. 1995, 25, 295–304. [Google Scholar] [CrossRef]
- Lachenbruch, P.A.; Cohen, J. Statistical Power Analysis for the Behavioral Sciences (2nd ed.). J. Am. Stat. Assoc. 1989, 84, 1096. [Google Scholar] [CrossRef]
- Uanhoro, J.O. Effect size calculators. 2017. Available online: https://effect-size-calculator.herokuapp.com/ (accessed on 2 March 2020).
- Pallant, J.F. SPSS Survival Manual: A Step by Step Guide to Data Analysis Using SPSS for Windows; Allen & Unwin.: Crows Nest, Australia, 2005. [Google Scholar]
- Ishimoto, H.; Ginzinger, D.G.; Jaffe, R.B. Adrenocorticotropin Preferentially Up-Regulates Angiopoietin 2 in the Human Fetal Adrenal Gland: Implications for Coordinated Adrenal Organ Growth and Angiogenesis. J. Clin. Endocrinol. Metab. 2006, 91, 1909–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimoto, H.; Minegishi, K.; Higuchi, T.; Furuya, M.; Asai, S.; Kim, S.H.; Tanaka, M.; Yoshimura, Y.; Jaffe, R. The periphery of the human fetal adrenal gland is a site of angiogenesis: Zonal differential expression and regulation of angiogenic factors. J. Clin. Endocrinol. Metab. 2008, 93, 2402–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitoshi, I.; Robert, B. Development and Function of the Human Fetal Adrenal Cortex: A Key Component in the Feto-Placental Unit. Endocr. Rev. 2011, 32, 317–355. [Google Scholar]
- Hu, V.; Sarachana, T.; Sherrard, R.M.; Kocher, K. Investigation of sex differences in the expression of RORA and its transcriptional targets in the brain as a potential contributor to the sex bias in autism. Mol. Autism 2015, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Sarachana, T.; Xu, M.; Wu, R.-C.; Hu, V. Sex Hormones in Autism: Androgens and Estrogens Differentially and Reciprocally Regulate RORA, a Novel Candidate Gene for Autism. PLoS ONE 2011, 6, e17116. [Google Scholar] [CrossRef]
- Sarachana, T.; Hu, V. Differential recruitment of coregulators to the RORA promoter adds another layer of complexity to gene (dys) regulation by sex hormones in autism. Mol. Autism 2013, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, B.; Dudbridge, F.; Kent, L.; Wheelwright, S.J.; Hill-Cawthorne, G.A.; Allison, C.; Banerjee-Basu, S.; Baron-Cohen, S. Genes related to sex steroids, neural growth, and social-emotional behavior are associated with autistic traits, empathy, and Asperger syndrome. Autism Res. 2009, 2, 157–177. [Google Scholar] [CrossRef]
- The Autism Genome Project. Consortium. Mapping autism risk loci using genetic linkage and chromosomal rearrangements. Nat. Genet. 2007, 39, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Jacquemont, M.-L.; Sanlaville, D.; Redon, R.; Raoul, O.; Cormier-Daire, V.; Lyonnet, S.; Amiel, J.; Le Merrer, M.; Heron, D.; De Blois, M.-C.; et al. Array-based comparative genomic hybridisation identifies high frequency of cryptic chromosomal rearrangements in patients with syndromic autism spectrum disorders. J. Med. Genet. 2006, 43, 843–849. [Google Scholar] [CrossRef]
- Cook, E.H., Jr.; Scherer, S.W. Copy-number variations associated with neuropsychiatric conditions. Nature 2008, 455, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.; Ronemus, M.; Yamrom, B.; Lee, Y.-H.; Leotta, A.; Kendall, J.; Marks, S.; Lakshmi, B.; Pai, D.; Ye, K.; et al. Rare De Novo and Transmitted Copy-Number Variation in Autistic Spectrum Disorders. Neuron 2011, 70, 886–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anney, R.; Klei, L.; Pinto, D.; Regan, R.; Conroy, J.; Magalhães, T.R.; Correia, C.; Abrahams, B.S.; Sykes, N.; Pagnamenta, A.T.; et al. A genome-wide scan for common alleles affecting risk for autism. Hum. Mol. Genet. 2010, 19, 4072–4082. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.A.; Casals, F.; Gauthier, J.; Hamdan, F.F.; Keebler, J.; Boyko, A.R.; Bustamante, C.D.; Piton, A.; Spiegelman, D.; Henrion, E.; et al. A Population Genetic Approach to Mapping Neurological Disorder Genes Using Deep Resequencing. PLoS Genet. 2011, 7, e1001318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iossifov, I.; Ronemus, M.; Levy, D.; Wang, Z.; Hakker, I.; Rosenbaum, J.; Yamrom, B.; Lee, Y.-H.; Narzisi, G.; Leotta, A.; et al. De Novo Gene Disruptions in Children on the Autistic Spectrum. Neuron 2012, 74, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neale, B.M.; Kou, Y.; Liu, L.; Ma’Ayan, A.; Samocha, K.; Sabo, A.; Lin, C.-F.; Stevens, C.; Wang, L.-S.; Makarov, V.; et al. Patterns and rates of exonic de novo mutations in autism spectrum disorders. Nature 2012, 485, 242–245. [Google Scholar] [CrossRef]
- O’Roak, B.; Vives, L.; Girirajan, S.; Karakoc, E.; Krumm, N.; Coe, B.P.; Levy, R.; Ko, A.; Lee, C.; Smith, J.D.; et al. Sporadic autism exomes reveal a highly interconnected protein network of de novo mutations. Nature 2012, 485, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Sanders, S.J.; Murtha, M.T.; Gupta, A.R.; Murdoch, J.D.; Raubeson, M.J.; Willsey, A.J.; Ercan-Sencicek, A.G.; DiLullo, N.M.; Parikshak, N.N.; Stein, J.; et al. De novo mutations revealed by whole-exome sequencing are strongly associated with autism. Nature 2012, 485, 237–241. [Google Scholar] [CrossRef]
- Menashe, I.; Grange, P.; Larsen, E.C.; Banerjee-Basu, S.; Mitra, P.P. Co-expression Profiling of Autism Genes in the Mouse Brain. PLoS Comput. Boil. 2013, 9, 1003128. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.N.; Kollu, R.; Banerjee-Basu, S. AutDB: A gene reference resource for autism research. Nucleic Acids Res. 2008, 37, D832–D836. [Google Scholar] [CrossRef] [Green Version]
- Vorstman, J.A.S.; Parr, J.R.; Moreno-De-Luca, D.; Anney, R.; Nurnberger, J.I., Jr.; Hallmayer, J.F. Autism genetics: Opportunities and challenges for clinical translation. Nat. Rev. Genet. 2017, 18, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Garber, K. Neuroscience - Autism’s cause may reside in abnormalities at the synapse. Science 2007, 317, 190–191. [Google Scholar] [CrossRef] [PubMed]
- Bourgeron, T. A synaptic trek to autism. Curr. Opin. Neurobiol. 2009, 19, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Durand, C.M.; Betancur, C.; Boeckers, T.M.; Bockmann, J.; Chaste, P.; Fauchereau, F.; Nygren, G.; Rastam, M.; Gillberg, I.C.; Anckarsäter, H.; et al. Mutations in the gene encoding the synaptic scaffolding protein SHANK3 are associated with autism spectrum disorders. Nat. Genet. 2006, 39, 25–27. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Urinary Steroid Hormone Metabolites | Autistic Girls | Control Girls | 95% CI | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (μg/mmol Creatinine) | Median | Mean | SEM | p-Value | Median | Mean | SEM | Effect Size | Lower Limit | Upper Limit |
| Androgen-Metabolites | ||||||||||
| Androsterone | 28.5 | 71.7 | 21.4 | 0.15 | 35.2 | 42.1 | 9.1 | 0.46 | −0.25 | 1.16 |
| Etiocholanolone | 24.6 | 57.8 | 16.6 | 0.17 | 26.4 | 38.8 | 14.3 | 0.32 | −0.39 | 1.00 |
| Androstenediol | 1.7 | 2.2 | 0.4 | 0.02 | 1.2 | 1.3 | 0.2 | 0.71 | 0.01 | 1.44 |
| 11-Oxo-Etiocholanolon | 30.4 | 35.1 | 6.7 | 0.11 | 19.2 | 20.9 | 3.0 | 0.67 | −0.04 | 1.39 |
| 11b-Hydroxy-Androsteron | 29.5 | 36.6 | 5.5 | 0.08 | 24.4 | 22.1 | 3.2 | 0.77 | 0.09 | 1.53 |
| 11b-Hydroxy-Etiocholanolon | 28.9 | 26.0 | 5.2 | 0.06 | 12.5 | 13.1 | 2.0 | 0.78 | 0.06 | 1.50 |
| Dehydroepiandrosteron (Wilcoxon) | 2.8 | 15.2 | 8.4 | 0.17 | 3.7 | 4.3 | 1.0 | 0.47 | ||
| 5-Androstene-3b,17b-diol | 3.4 | 6.8 | 2.6 | 0.10 | 1.8 | 2.2 | 0.4 | 0.62 | −0.09 | 1.33 |
| 16a-Hydroxy-DHEA | 8.2 | 57.6 | 36.9 | 0.20 | 6.0 | 8.0 | 1.5 | 0.48 | −0.33 | 1.07 |
| 5-Androstene-3b,16a,17b-triol | 8.1 | 36.8 | 16.8 | 0.12 | 7.2 | 8.6 | 1.9 | 0.59 | −0.11 | 1.31 |
| 5-Pregnene-3b, 16a,17b-triol | 4.4 | 11.6 | 4.4 | 0.70 | 6.8 | 9.7 | 1.7 | 0.14 | −0.50 | 0.89 |
| Testosterone | 0.8 | 0.8 | 0.1 | 0.02 | 0.5 | 0.5 | 0.1 | 0.83 | 0.26 | 1.73 |
| 5a-Dihydrotestosteron | 1.2 | 1.2 | 0.2 | 0.94 | 1.2 | 1.2 | 0.1 | 0.03 | −0.51 | 0.87 |
| Oestrogen-Metabolites | ||||||||||
| Estriol | 0.1 | 0.6 | 0.4 | 0.11 | 0.1 | 0.4 | 0.1 | 0.19 | −0.60 | 0.78 |
| 17b-Estradiol (Wilcoxon) | 0.046 | 0.1 | 0.0 | 0.10 | 0.1 | 0.2 | 0.1 | −0.49 | ||
| Progesterone-Metabolites | ||||||||||
| 17-Hydroxypregnanolon (Wilcoxon) | 2.7 | 5.5 | 1.4 | 0.33 | 3.3 | 3.5 | 0.6 | 0.47 | ||
| Pregnanediol (Wilcoxon) | 11.6 | 18.9 | 4.6 | 0.50 | 10.8 | 12.7 | 2.4 | 0.44 | ||
| Pregnanetriol | 18.4 | 33.0 | 9.2 | 0.17 | 19.3 | 20.5 | 3.5 | 0.46 | −0.23 | 1.18 |
| 11-Oxo-Pregnanetriol (Wilcoxon) | 1.0 | 1.2 | 0.2 | 0.99 | 1.1 | 1.4 | 0.4 | −0.20 | ||
| Aldosterone-Metabolites | ||||||||||
| Tetrahydroaldosterone | 1.9 | 2.2 | 0.5 | 0.05 | 4.1 | 1.1 | 0.1 | 0.72 | 0.04 | 1.48 |
| Corticosterone-Metabolites | ||||||||||
| TetrahydroDOC | 0.5 | 0.6 | 0.1 | 0.71 | 1.0 | 0.5 | 0.1 | 0.14 | −0.50 | 0.89 |
| Tetrahydrodehydrocorticosterone | 9.5 | 10.2 | 1.6 | 0.71 | 0.4 | 9.4 | 1.2 | 0.14 | −0.59 | 0.80 |
| Tetrahydrocorticosterone (Wilcoxon) | 9.3 | 9.2 | 1.0 | 0.4965 | 9.7 | 9.6 | 2.1 | −0.07 | ||
| 5a-Tetrahydrocorticosteron | 18.6 | 18.9 | 2.8 | 0.43 | 8.2 | 24.8 | 6.7 | −0.30 | −1.00 | 0.40 |
| 18-Hydroxy-Tetrahydrocompound A (Wilcoxon) | 1.1 | 1.9 | 0.6 | 0.04 | 24.8 | 7.1 | 2.1 | −0.81 | ||
| Cortisone | 9.0 | 10.2 | 0.9 | 0.70 | 3.9 | 10.9 | 1.5 | −0.15 | −0.89 | 0.50 |
| Cortisone-Metabolites | ||||||||||
| Tetrahydrocortisone | 176.6 | 203.4 | 22.6 | 0.75 | 10.3 | 194.1 | 25.0 | 0.10 | −0.60 | 0.79 |
| a-Cortolon | 65.2 | 70.2 | 7.7 | 0.23 | 194.1 | 56.4 | 7.0 | 0.48 | −0.22 | 1.18 |
| b-Cortolon | 169.0 | 220.4 | 69.1 | 0.02 | 62.2 | 43.3 | 5.5 | 0.85 | 0.17 | 1.63 |
| 20a-Dihydrocortison | 1.1 | 1.2 | 0.1 | 0.87 | 47.2 | 1.2 | 0.2 | 0.07 | −0.69 | 0.69 |
| 20b-Dihydrocortison | 3.4 | 3.7 | 0.5 | 0.49 | 1.2 | 3.2 | 0.5 | 0.29 | −0.44 | 0.95 |
| Cortisol | 5.9 | 6.3 | 0.6 | 0.02 | 3.0 | 4.1 | 0.5 | 0.41 | 0.26 | 1.73 |
| Cortisol-Metabolites | ||||||||||
| Tetrahydrocortisol | 61.9 | 68.4 | 8.5 | 0.62 | 4.7 | 61.5 | 9.0 | 0.21 | −0.49 | 0.90 |
| 5a-Tetrahydrocortisol | 56.7 | 77.0 | 16.1 | 0.54 | 52.4 | 96.7 | 23.7 | -0.25 | −0.94 | 0.45 |
| a-Cortol | 10.8 | 13.5 | 1.8 | 0.65 | 89.4 | 12.1 | 1.9 | 0.20 | −0.55 | 0.84 |
| b-Cortol | 12.8 | 15.2 | 2.3 | 0.08 | 12.0 | 26.0 | 4.4 | -0.75 | −1.52 | -0.08 |
| 20a-Dihydrocortisol (Wilcoxon) | 2.3 | 3.2 | 0.6 | 0.40 | 3.2 | 3.6 | 0.7 | -0.15 | ||
| 6b-Hydroxycortisol | 9.3 | 11.1 | 1.5 | 0.55 | 9.8 | 9.8 | 1.4 | 0.23 | −0.47 | 0.92 |
| 18-Hydroxycortisol | 27.0 | 35.4 | 4.8 | 0.02 | 19.9 | 21.3 | 3.1 | 0.83 | −0.47 | 0.92 |
| Total | ||||||||||
| Total Androgens | 150.0 | 359.4 | 105.1 | 0.08 | 122.0 | 172.8 | 29.8 | 0.60 | −0.11 | 1.31 |
| Total of Cortisol and Cortisone | 610.9 | 740.9 | 101.3 | 0.13 | 605.2 | 552.1 | 75.9 | 0.53 | −0.17 | 1.24 |
| Total - All Measured Metabolites | 982.2 | 1206.3 | 170.2 | 0.08 | 835.2 | 814.4 | 96.3 | 0.70 | 0.03 | 1.47 |
| Autistic Girls | Control Girls | ||||
|---|---|---|---|---|---|
| Mean | SEM | p-Value | Mean | SEM | |
| 18OHTHA/THALDO | 1.175 | 0.399 | 0.014 | 1.595 | 0.419 |
| 21-Hydroxylase Deficit | |||||
| 17HP/(THE + THF + 5aTHF) | 0.017 | 0.003 | 0.026 | 0.010 | 0.002 |
| PT/(THE + THF + 5aTHF) | 0.098 | 0.016 | 0.064 | 0.073 | 0.013 |
| 100 × PT’ONE/(THE + THF + 5aTHF) | 0.358 | 0.043 | 0.644 | 0.404 | 0.099 |
| 17-Hydroxylase Deficit | |||||
| (THA + THB + 5aTHB)/(THE + THF + 5aTHF) | 0.256 | 0.012 | 0.407 | 0.287 | 0.021 |
| 100 × THDOC/(THE + THF + 5aTHF) | 0.257 | 0.069 | 0.157 | 0.145 | 0.018 |
| 11-Hydroxylase Deficit | |||||
| 100 × THS/(THE + THF + 5aTHF) | 1.682 | 0.152 | 0.330 | 1.864 | 0.151 |
| 100 × THDOC/(THE + THF + 5aTHF) | 0.257 | 0.069 | 0.157 | 0.145 | 0.018 |
| 11-Beta Hydroxylase | |||||
| F/E | 0.608 | 0.035 | 0.002 | 0.445 | 0.026 |
| (THF + 5aTHF)/THE | 0.642 | 0.058 | 0.290 | 0.044 | 0.004 |
| (F + E)/THE + THF + 5aTHF | 0.057 | 0.005 | 0.788 | 0.044 | 0.004 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gasser, B.A.; Kurz, J.; Dick, B.; Mohaupt, M.G. Are Steroid Hormones Dysregulated in Autistic Girls? Diseases 2020, 8, 6. https://doi.org/10.3390/diseases8010006
Gasser BA, Kurz J, Dick B, Mohaupt MG. Are Steroid Hormones Dysregulated in Autistic Girls? Diseases. 2020; 8(1):6. https://doi.org/10.3390/diseases8010006
Chicago/Turabian StyleGasser, Benedikt Andreas, Johann Kurz, Bernhard Dick, and Markus Georg Mohaupt. 2020. "Are Steroid Hormones Dysregulated in Autistic Girls?" Diseases 8, no. 1: 6. https://doi.org/10.3390/diseases8010006
APA StyleGasser, B. A., Kurz, J., Dick, B., & Mohaupt, M. G. (2020). Are Steroid Hormones Dysregulated in Autistic Girls? Diseases, 8(1), 6. https://doi.org/10.3390/diseases8010006