Metabolic Profiling of Hybrids Generated from Pummelo and Citrus latipes in Relation to Their Attraction to Diaphorina citri, the Vector of Huanglongbing

 ,
,  ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
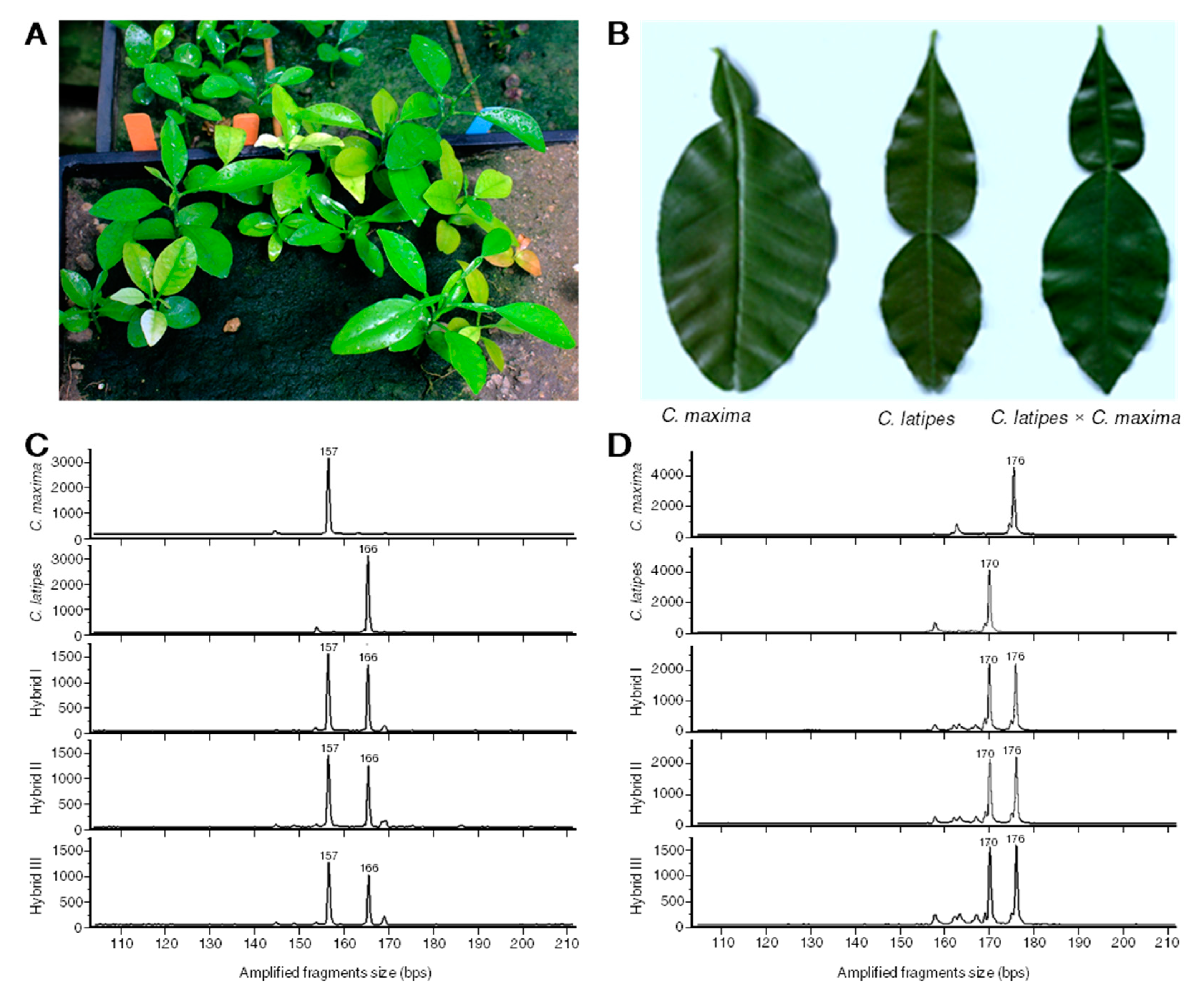
2.1. Morphology of New Hybrids
2.2. Simple Sequence Repeat
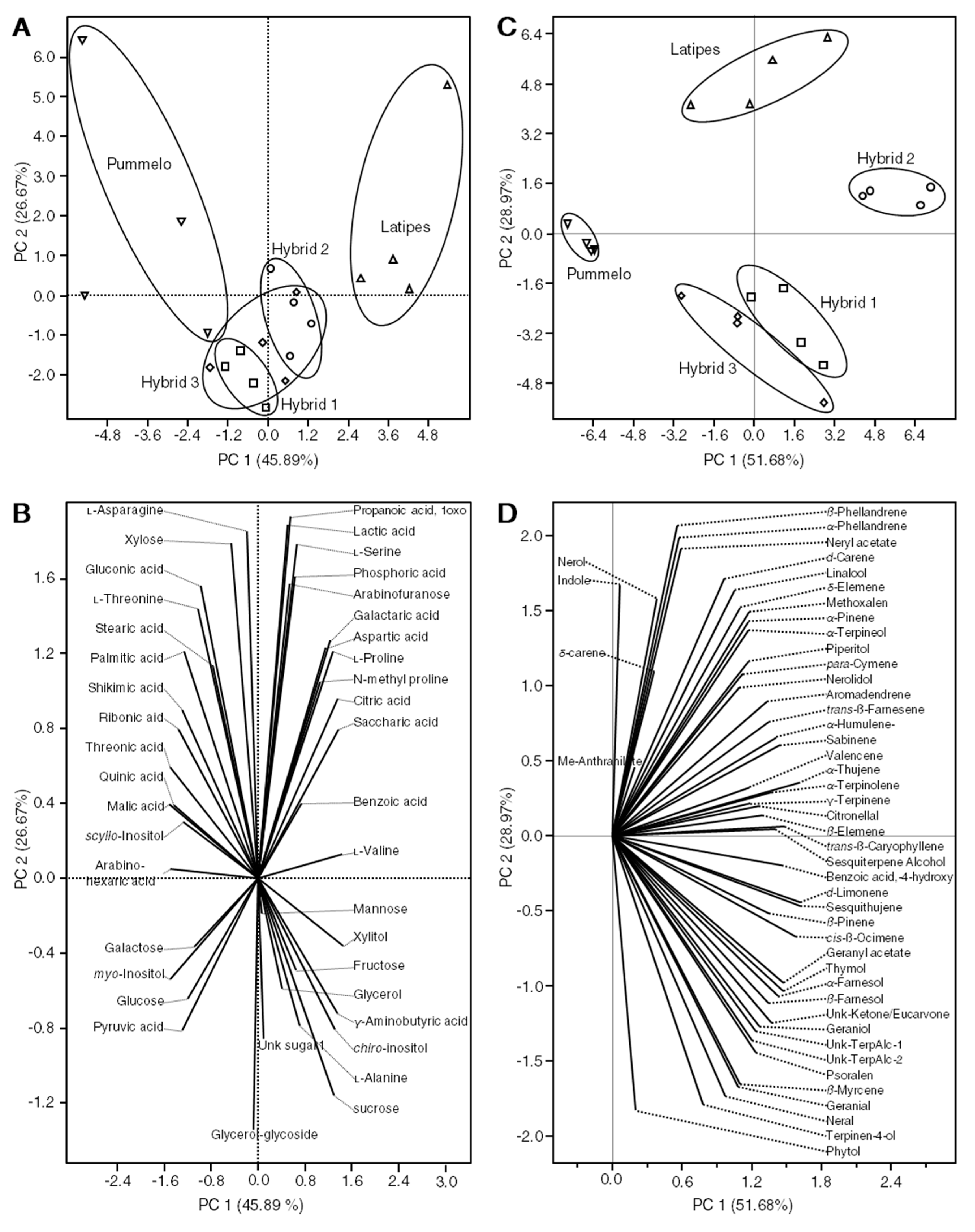
2.3. Polar Metabolites
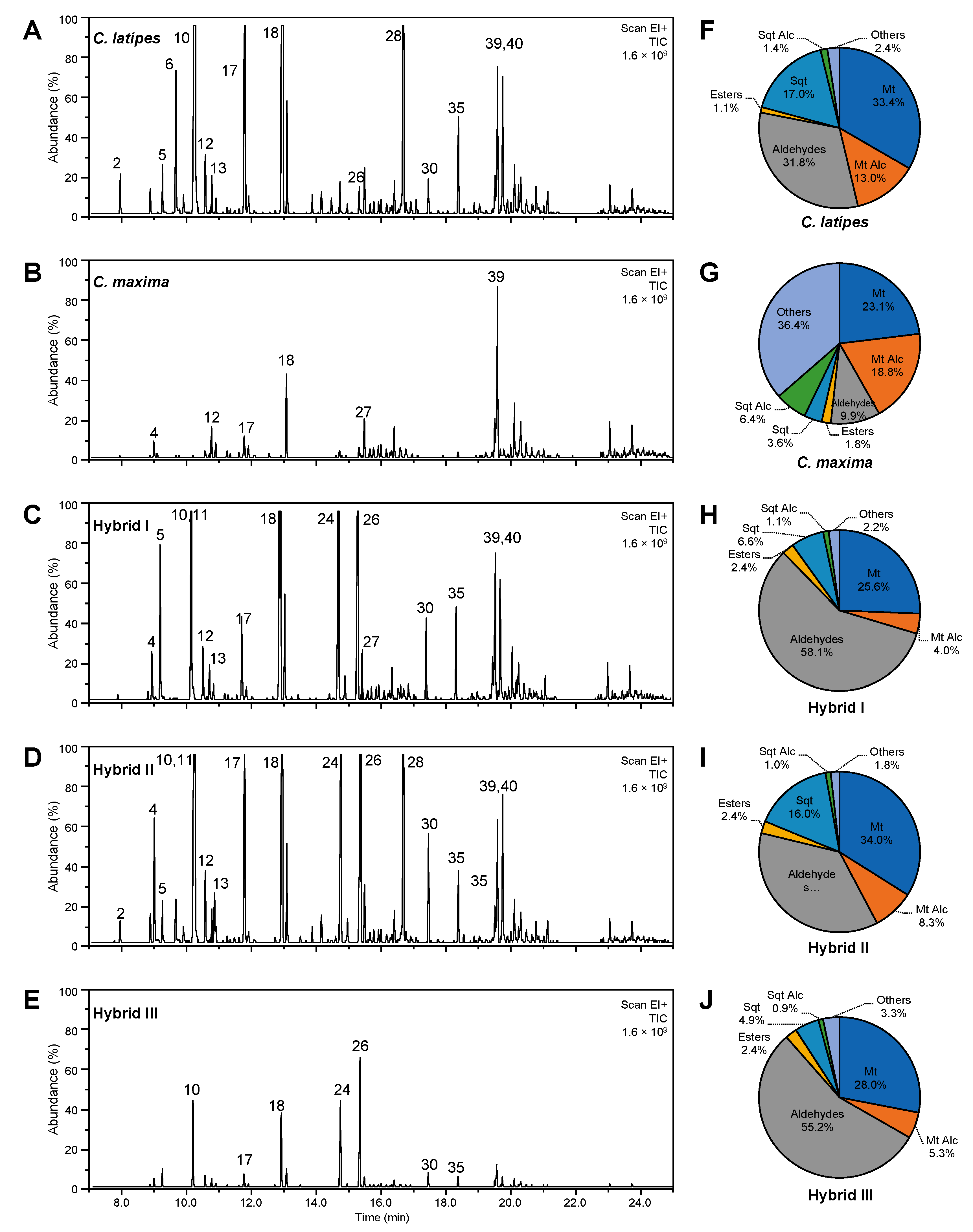
2.4. Volatile Content of Hexane Extract
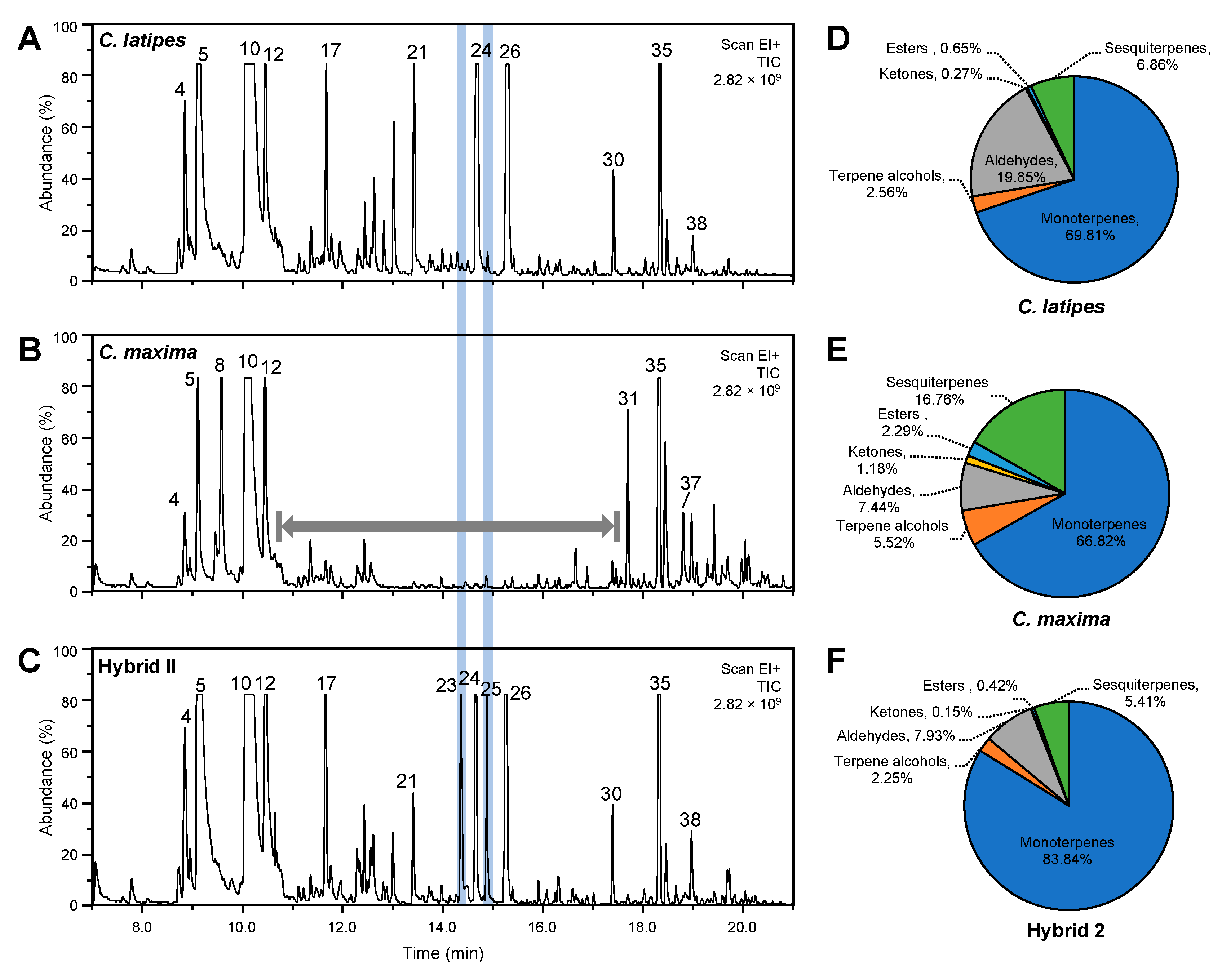
2.5. Released Volatiles Collected by Static Headspace SPME
2.6. Psyllid Attraction
3. Discussion
4. Material and Methods
4.1. Hybridization and Seedling Selection
4.2. Genomic DNA Extraction
4.3. Plant Genotyping Using SSR Markers
4.4. Extraction and Analysis of Polar Metabolites
4.5. Hexane Extraction and Analysis of Citrus Leaf Volatiles
4.6. Collection and Analysis of Released Volatiles
4.7. Identification and Quantification of Leaf VOCs and Polar Metabolites
4.8. Psyllid Attraction
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tiwari, S.; Mann, R.S.; Rogers, M.E.; Stelinski, L.L. Insecticide resistance in field populations of Asian citrus psyllid in Florida. Pest Manag. Sci. 2011, 67, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Halbert, S.E.; Manjunath, K.L. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in FLorida. Fla. Entomol. 2004, 87, 330–353. [Google Scholar] [CrossRef]
- Folimonova, S.Y.; Robertson, C.J.; Garnsey, S.M.; Gowda, S.; Dawson, W.O. Examination of the responses of different genotypes of citrus to huanglongbing (citrus greening) under different conditions. Phytopathology 2009, 99, 1346–1354. [Google Scholar] [CrossRef] [Green Version]
- Richardson, M.L.; Hall, D.G.; Westbrook, R.; Stover, E.W.; Duan, Y.P. Resistance of Poncirus and Citrus x Poncirus germplasm to the Asian citrus psyllid. J. Citrus Pathol. 2014, 1, 266. [Google Scholar] [CrossRef]
- Tsai, J.H.; Liu, Y.H. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. J. Econ. Entomol. 2000, 93, 1721–1725. [Google Scholar] [CrossRef] [PubMed]
- Patt, J.M.; Sétamou, M. Responses of the Asian citrus psyllid to volatiles emitted by the flushing shoots of its rutaceous host plants. Environ. Entomol. 2010, 39, 618–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sétamou, M.; Flores, D.; French, J.V.; Hall, D.G. Dispersion patterns and sampling plans for Diaphorina citri (hemiptera: Psyllidae) in citrus. J. Econ. Entomol. 2008, 101, 1478–1487. [Google Scholar] [CrossRef]
- Wenninger, E.J.; Stelinski, L.L.; Hall, D.G. Roles of Olfactory Cues, Visual cues, and mating status in orientation of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) to four different host plants. Environ. Entomol. 2009, 38, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Killiny, N.; Jones, S.E. Profiling of volatile organic compounds released from individual intact juvenile and mature citrus leaves. J. Plant Physiol. 2017, 208, 47–51. [Google Scholar] [CrossRef]
- Hijaz, F.; Killiny, N. Collection and chemical composition of phloem sap from Citrus sinensis L. Osbeck (Sweet Orange). PLoS ONE 2014, 9, e101830. [Google Scholar] [CrossRef] [Green Version]
- Hijaz, F.; Nehela, Y.; Killiny, N. Possible role of plant volatiles in tolerance against huanglongbing in citrus. Plant Signal. Behav. 2016, 11, e1138193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijaz, F.; Al-Rimawi, F.; Manthey, J.A.; Killiny, N. Phenolics, flavonoids and antioxidant capacities in Citrus species with different degree of tolerance to Huanglongbing. Plant Signal. Behav. 2020, 15, e1752447. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Valim, M.F.; Jones, S.E.; Omar, A.A.; Hijaz, F.; Gmitter, F.G.; Grosser, J.W. Metabolically speaking: Possible reasons behind the tolerance of “Sugar Belle” mandarin hybrid to huanglongbing. Plant Physiol. Biochem. 2017, 116, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killiny, N.; Jones, S.E.; Nehela, Y.; Hijaz, F.; Dutt, M.; Gmitter, F.G.; Grosser, J.W. All roads lead to Rome: Towards understanding different avenues of tolerance to huanglongbing in citrus cultivars. Plant Physiol. Biochem. 2018, 129, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Matsuda, F. Metabolomics for functional genomics, systems biology, and biotechnology. Annu. Rev. Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.; Worrich, A.; Weinhold, A.; Alka, O.; Balcke, G.; Birkemeyer, C.; Bruelheide, H.; Calf, O.W.; Dietz, S.; Dührkop, K.; et al. Current challenges in plant Eco-Metabolomics. Int. J. Mol. Sci. 2018, 19, 1385. [Google Scholar] [CrossRef] [Green Version]
- Ramadugu, C.; Keremane, M.L.; Halbert, S.E.; Duan, Y.P.; Roose, M.L.; Stover, E.; Lee, R.F. Long-term field evaluation reveals huanglongbing resistance in Citrus relatives. Plant Dis. 2016, 100, 1858–1869. [Google Scholar] [CrossRef] [Green Version]
- Gancel, A.L.; Ollitrault, P.; Froelicher, Y.; Tomi, F.; Jacquemond, C.; Luro, F.; Brillouet, J.M. Leaf volatile compounds of six citrus somatic allotetraploid hybrids originating from various combinations of lime, lemon, citron, sweet orange, and grapefruit. J. Agric. Food Chem. 2005, 53, 2224–2230. [Google Scholar] [CrossRef]
- Dicke, M. Chemical ecology of host-plant selection by herbivorous arthropods: A multitrophic perspective. Biochem. Syst. Ecol. 2000, 28, 601–617. [Google Scholar] [CrossRef]
- Coutinho-Abreu, I.V.; Forster, L.; Guda, T.; Ray, A. Odorants for surveillance and control of theAsian citrus psyllid (Diaphorina citri). PLoS ONE 2014, 9, e109236. [Google Scholar] [CrossRef]
- Webster, B.; Bruce, T.; Pickett, J.; Hardie, J. Volatiles functioning as host cues in a blend become nonhost cues when presented alone to the black bean aphid. Anim. Behav. 2010, 79, 451–457. [Google Scholar] [CrossRef]
- Visser, J.H. Host Odor perception in phytophagous insects. Annu. Rev. Entomol. 1986, 31, 121–144. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents-a checklist of volatile compounds isolated by head-space techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Tomaseto, A.F.; Marques, R.N.; Fereres, A.; Zanardi, O.Z.; Volpe, H.X.L.; Alquézar, B.; Peña, L.; Miranda, M.P. Orange jasmine as a trap crop to control Diaphorina citri. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Grosser, J.W.; Ćalović, M.; Serrano, P.; Pasquali, G.; Gmitter, J.; Gmitter, F.G. Verification of mandarin and pummelo somatic hybrids by expressed sequence tag-simple sequence repeat marker analysis. J. Am. Soc. Hortic. Sci. 2008, 133, 794–800. [Google Scholar] [CrossRef] [Green Version]
- Michie, M.G. Use of the Bray-Curtis similarity measure in cluster analysis of foraminiferal data. J. Int. Assoc. Math. Geol. 1982, 14, 661–667. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
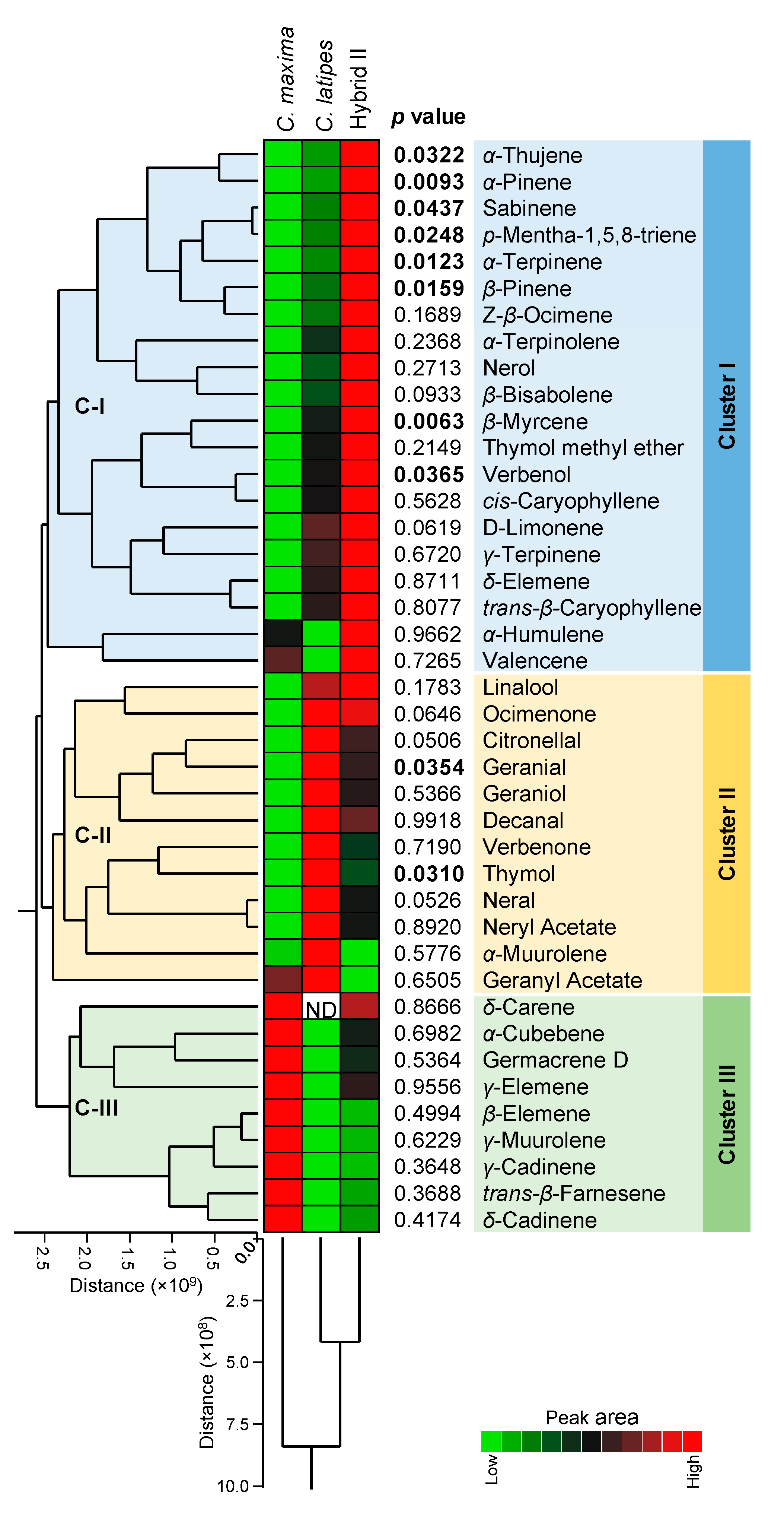{kind=link}
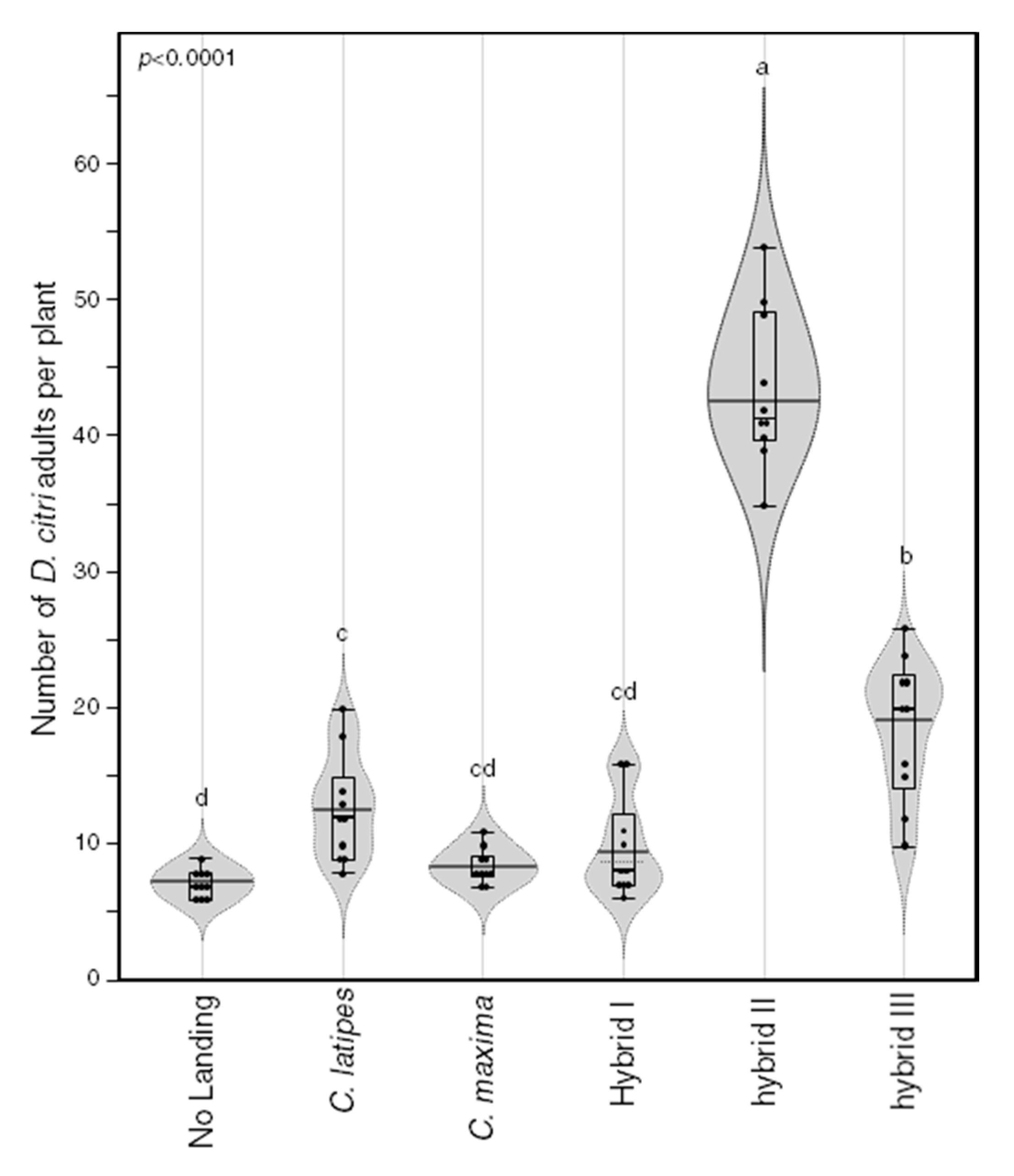{kind=link}
| EST-SSR Markers a (Base Pair) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CX6F02B | CX6F04G | CX6F06Y | CX6F10R | CX6F17B | CX6F18G | CX6F29Y | CX0035R | |||||||||
| C. maxima | 168 | 168 | 163 | 169 | 157 | 157 | 176 | 176 | 132 | 132 | 160 | 161 | 152 | 152 | 183 | 183 |
| C. latipes | 168 | 168 | 157 | 169 | 166 | 166 | 170 | 170 | 126 | 132 | 148 | 148 | 152 | 152 | 183 | 183 |
| Hybrid I | 168 | 168 | 169 | 169 | 157 | 166 | 170 | 176 | 126 | 132 | 148 | 160 | 152 | 152 | 183 | 183 |
| Hybrid II | 168 | 168 | 169 | 169 | 157 | 166 | 170 | 176 | 126 | 132 | 148 | 161 | 152 | 152 | 183 | 183 |
| Hybrid III | 168 | 168 | 169 | 169 | 157 | 166 | 170 | 176 | 126 | 132 | 148 | 160 | 152 | 152 | 183 | 183 |
| Peak No. | VOC y | RT | Citrus maxima | Citrus latipes | Hybrid I | Hybrid II | Hybrid III | p-Value x |
|---|---|---|---|---|---|---|---|---|
| 1 | α-Thujene v | 7.78 | ND | ND | 0.01 ± 0.01 b | 0.22 ± 0.20 a | 0.01 ± 0.01 b | 0.0091 |
| 2 | α-Pinene v | 7.95 | 0.02 ± 0.01 c | 1.92 ± 0.65 ab | 0.47 ± 0.33 bc | 2.65 ± 1.30 a | 0.25 ± 0.22 c | 0.0002 |
| 3 | Sabinene v | 8.90 | 0.07 ± 0.04 b | 1.15 ± 0.45 b | 0.80 ± 0.50 b | 3.05 ± 1.05 a | 0.69 ± 0.31 b | <0.0001 |
| 4 | β-Pinene v | 9.00 | 0.51 ± 0.25 b | 0.05 ± 0.06 b | 4.53 ± 2.38 b | 11.84 ± 4.50 a | 4.08 ± 1.29 b | <0.0001 |
| 5 | β-Myrcene u | 9.25 | 0.04 ± 0.02 c | 8.81 ± 3.30 c | 46.38 ± 17.31 a | 13.70 ± 2.53 bc | 31.80 ± 9.98 ab | <0.0001 |
| 6 | α-Phellandrene s | 9.66 | 0.04 ± 0.01 c | 4.26 ± 1.30 a | 0.14 ± 0.10 c | 1.98 ± 0.24 b | 0.13 ± 0.09 c | <0.0001 |
| 7 | γ-Carene u | 9.71 | ND | 0.15 ± 0.15 ns | 0.04 ± 0.03 ns | 0.03 ± 0.01 ns | 0.09 ± 0.09 ns | 0.1076 |
| 8 | δ-Carene u | 9.90 | 0.09 ± 0.16 b | 1.04 ± 0.39 a | 0.02 ± 0.01 b | 1.25 ± 0.26 a | 0.02 ± 0.02 b | <0.0001 |
| 9 | ρ-Cymene w,t | 10.05 | ND | 0.03 ± 0.02 ns | ND | 0.09 ± 0.09 ns | ND | 0.3062 |
| 10 | d-Limonene s | 10.20 | 0.14 ± 0.07 c | 19.30 ± 10.79 b | 30.70 ± 6.61 ab | 35.59 ± 6.22 a | 24.21 ± 4.76 ab | <0.0001 |
| 11 | β-Phellandrene w,s | 10.30 | ND | 17.34 ± 4.67 a | ND | 8.14 ± 2.09 b | ND | 0.0214 |
| 12 | Z-β-Ocimene s | 10.60 | 0.12 ± 0.05 c | 1.79 ± 0.65 b | 3.11 ± 0.70 ab | 3.73 ± 0.90 a | 2.88 ± 0.55 ab | <0.0001 |
| 13 | γ-Terpinene s | 10.70 | 0.37 ± 0.08 b | 0.53 ± 0.10 b | 0.69 ± 0.18 b | 3.58 ± 1.13 a | 0.78 ± 0.10 b | <0.0001 |
| 14 | Unknown terpene alcohol 1 r | 11.25 | 0.20 ± 0.17 b | 0.54 ± 0.31 ab | 1.07 ± 0.60 a | 0.96 ± 0.23 ab | 1.16 ± 0.40 a | 0.0117 |
| 15 | Unknown terpene alcohol 2 r | 11.35 | ND | 0.11 ± 0.03 ab | 0.17 ± 0.05 a | 0.16 ± 0.02 ab | 0.18 ± 0.05 a | 0.0132 |
| 16 | α-Terpinolene s | 11.50 | 0.01 ± 0.01 b | 0.06 ± 0.06 b | 0.05 ± 0.07 b | 0.27 ± 0.05 a | 0.06 ± 0.04 b | <0.0001 |
| 17 | Linalool r | 11.80 | 0.55 ± 0.12 b | 14.02 ± 3.53 a | 4.34 ± 0.50 b | 10.95 ± 1.81 a | 4.30 ± 0.69 b | <0.0001 |
| 18 | Citronellal q | 12.90 | ND | 42.73 ± 8.32 ab | 53.40 ± 12.39 a | 28.14 ± 6.97 bc | 19.35 ± 0.87 c | <0.0001 |
| 19 | Indole w,r | 13.41 | ND | 0.11 ± 0.16 ns | 0.02 ± 0.03 ns | ND | ND | 0.3786 |
| 20 | Terpinen-4-ol r | 13.51 | ND | 0.02 ± 0.02 b | 0.30 ± 0.21 a | 0.22 ± 0.07 ab | 0.33 ± 0.11 a | 0.0431 |
| 21 | α-Terpineol r | 13.86 | ND | 0.53 ± 0.21 b | 0.09 ± 0.02 c | 0.81 ± 0.10 a | ND | <0.0001 |
| 22 | Piperitol w,r | 14.16 | ND | 0.56 ± 0.28 b | ND | 1.31 ± 0.51 a | ND | 0.0350 |
| 23 | Nerol r | 14.45 | ND | 0.55 ± 0.16 a | 0.15 ± 0.17 b | 0.12 ± 0.01 b | 0.14 ± 0.06 b | <0.0001 |
| 24 | Neral p | 14.75 | 0.14 ± 0.05 b | 1.17 ± 1.73 b | 64.99 ± 29.56 a | 56.58 ± 15.11 a | 67.42 ± 6.82 a | <0.0001 |
| 25 | Geraniol r | 14.95 | ND | 0.27 ± 0.09 b | 1.25 ± 0.30 a | 1.17 ± 0.12 a | 0.95 ± 0.19 a | <0.0001 |
| 26 | Geranial p | 15.36 | 0.60 ± 0.55 b | 2.25 ± 1.62 b | 94.98 ± 44.93 a | 83.56 ± 22.25 a | 95.57 ± 41.18 a | 0.0002 |
| 27 | Thymol r | 15.75 | 0.28 ± 0.07 c | 0.40 ± 0.07 bc | 0.49 ± 0.08 ab | 0.50 ± 0.03 ab | 0.53 ± 0.09 a | 0.0007 |
| 28 | δ-Elemene o | 16.77 | ND | 10.45 ± 3.08 b | 0.37 ± 0.14 c | 15.09 ± 2.46 a | 0.20 ± 0.09 c | <0.0001 |
| 29 | Neryl acetate p | 17.10 | 0.23 ± 0.09 b | 1.54 ± 0.40 a | 0.57 ± 0.20 b | 0.64 ± 0.17 b | 0.28 ± 0.03 b | <0.0001 |
| 30 | Geranyl acetate p | 17.45 | 0.01 ± 0.01 d | 2.90 ± 0.84 cd | 12.48 ± 1.66 ab | 14.62 ± 4.61 a | 8.97 ± 1.67 c | <0.0001 |
| 31 | β-Elemene o | 17.70 | 0.02 ± 0.01 c | 0.08 ± 0.04 b | 0.10 ± 0.01 ab | 0.15 ± 0.02 a | 0.07 ± 0.04 c | <0.0001 |
| 32 | Sesquithujene o | 17.90 | 0.03 ± 0.01 ns | 0.04 ± 0.00 ns | 0.05 ± 0.01 ns | 0.05 ± 0.02 ns | 0.04 ± 0.02 ns | 0.1753 |
| 33 | Anthranilate methyl ester w,p | 18.05 | ND | 0.31 ± 0.07 ns | ND | ND | 0.34 ± 0.22 ns | 0.5573 |
| 34 | Unknown Ketone/Eucarvone p | 18.20 | 0.02 ± 0.00 b | 0.02 ± 0.01 b | 0.15 ± 0.07 ab | 0.24 ± 0.05 a | 0.20 ± 0.18 ab | 0.0071 |
| 35 | trans-β-Caryophyllene o | 18.33 | 0.09 ± 0.04 c | 3.14 ± 0.55 a | 3.54 ± 0.61 a | 2.59 ± 0.59 ab | 2.12 ± 0.26 b | <0.0001 |
| 36 | Aromadendrene o | 18.50 | ND | 0.12 ± 0.04 b | 0.05 ± 0.01 bc | 0.32 ± 0.08 a | 0.02 ± 0.02 c | <0.0001 |
| 37 | trans-β-Farnesene o | 18.88 | ND | 0.25 ± 0.06 a | 0.22 ± 0.04 a | 0.22 ± 0.02 a | 0.11 ± 0.06 b | <0.0001 |
| 38 | α-Humulene o | 19.03 | ND | 0.34 ± 0.09 a | 0.32 ± 0.19 a | 0.32 ± 0.06 a | 0.16 ± 0.09 b | 0.0013 |
| 39 | Valencene o | 19.75 | ND | 3.92 ± 0.77 bc | 4.60 ± 1.13 ab | 6.21 ± 1.67 a | 2.21 ± 0.37 c | <0.0001 |
| 40 | Benzoic acid,4-hydroxy p | 20.11 | 5.49 ± 2.17 b | 7.95 ± 0.69 ab | 8.05 ± 0.62 a | 8.23 ± 0.52 a | 8.23 ± 1.02 a | 0.0201 |
| 41 | Nerolidol n | 20.25 | ND | 0.63 ± 0.08 a | 0.44 ± 0.10 b | 0.38 ± 0.09 bc | 0.24 ± 0.07 c | <0.0001 |
| 42 | Sesquiterpene Alcohol n | 21.12 | 0.03 ± 0.01 c | 0.68 ± 0.13 ab | 0.90 ± 0.18 a | 1.02 ± 0.15 a | 0.47 ± 0.24 b | <0.0001 |
| 43 | α-Farnesol n | 22.75 | 0.11 ± 0.03 ns | 0.12 ± 0.02 ns | 0.15 ± 0.03 ns | 0.15 ± 0.04 ns | 0.14 ± 0.07 ns | 0.5531 |
| 44 | β-Farnesol n | 22.85 | 0.12 ± 0.03 ns | 0.13 ± 0.03 ns | 0.18 ± 0.04 ns | 0.18 ± 0.05 ns | 0.16 ± 0.11 ns | 0.5697 |
| 45 | Psoralen n | 25.00 | 0.01 ± 0.00 b | 0.03 ± 0.03 b | 0.16 ± 0.02 a | 0.13 ± 0.09 ab | 0.13 ± 0.08 ab | 0.0060 |
| 46 | Methoxalen n | 28.20 | ND | 0.39 ± 0.23 b | 0.07 ± 0.03 c | 0.44 ± 0.13 a | 0.11 ± 0.12 bc | 0.0005 |
| 47 | Phytol n | 28.95 | 0.04 ± 0.04 ns | 0.04 ± 0.01 ns | 0.76 ± 1.37 ns | 0.07 ± 0.04 ns | 1.14 ± 1.54 ns | 0.3385 |
| Total VOCs | 9.37 ± 4.10 c | 152.78 ± 46.31 b | 341.35 ± 123.53 a | 321.67 ± 78.64 a | 280.29 ± 74.23 a | <0.0001 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Killiny, N.; Jones, S.E.; Hijaz, F.; Kishk, A.; Santos-Ortega, Y.; Nehela, Y.; Omar, A.A.; Yu, Q.; Gmitter, F.G., Jr.; Grosser, J.W.; et al. Metabolic Profiling of Hybrids Generated from Pummelo and Citrus latipes in Relation to Their Attraction to Diaphorina citri, the Vector of Huanglongbing. Metabolites 2020, 10, 477. https://doi.org/10.3390/metabo10120477
Killiny N, Jones SE, Hijaz F, Kishk A, Santos-Ortega Y, Nehela Y, Omar AA, Yu Q, Gmitter FG Jr., Grosser JW, et al. Metabolic Profiling of Hybrids Generated from Pummelo and Citrus latipes in Relation to Their Attraction to Diaphorina citri, the Vector of Huanglongbing. Metabolites. 2020; 10(12):477. https://doi.org/10.3390/metabo10120477
Chicago/Turabian StyleKilliny, Nabil, Shelley E. Jones, Faraj Hijaz, Abdelaziz Kishk, Yulica Santos-Ortega, Yasser Nehela, Ahmad A. Omar, Qibin Yu, Fred G. Gmitter, Jr., Jude W. Grosser, and et al. 2020. "Metabolic Profiling of Hybrids Generated from Pummelo and Citrus latipes in Relation to Their Attraction to Diaphorina citri, the Vector of Huanglongbing" Metabolites 10, no. 12: 477. https://doi.org/10.3390/metabo10120477
APA StyleKilliny, N., Jones, S. E., Hijaz, F., Kishk, A., Santos-Ortega, Y., Nehela, Y., Omar, A. A., Yu, Q., Gmitter, F. G., Jr., Grosser, J. W., & Dutt, M. (2020). Metabolic Profiling of Hybrids Generated from Pummelo and Citrus latipes in Relation to Their Attraction to Diaphorina citri, the Vector of Huanglongbing. Metabolites, 10(12), 477. https://doi.org/10.3390/metabo10120477