Nematode-Trapping Fungi Produce Diverse Metabolites during Predator–Prey Interaction

 ,
,
Abstract
:
1. Introduction
2. Results
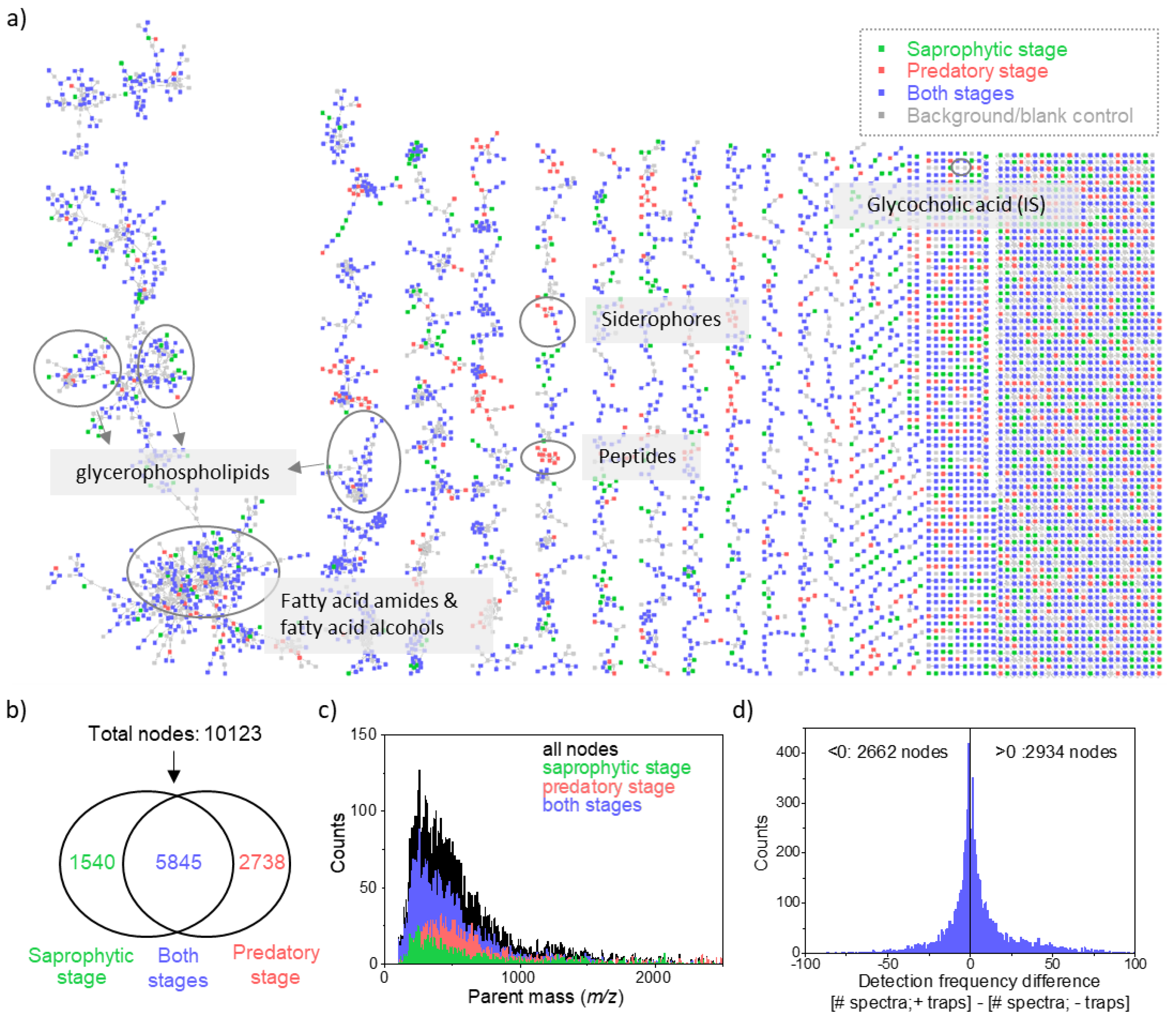
2.1. Revealing Chemical Diversity of Metabolites of Nematode-Trapping Fungi in Different Life Stages With Molecular Networking
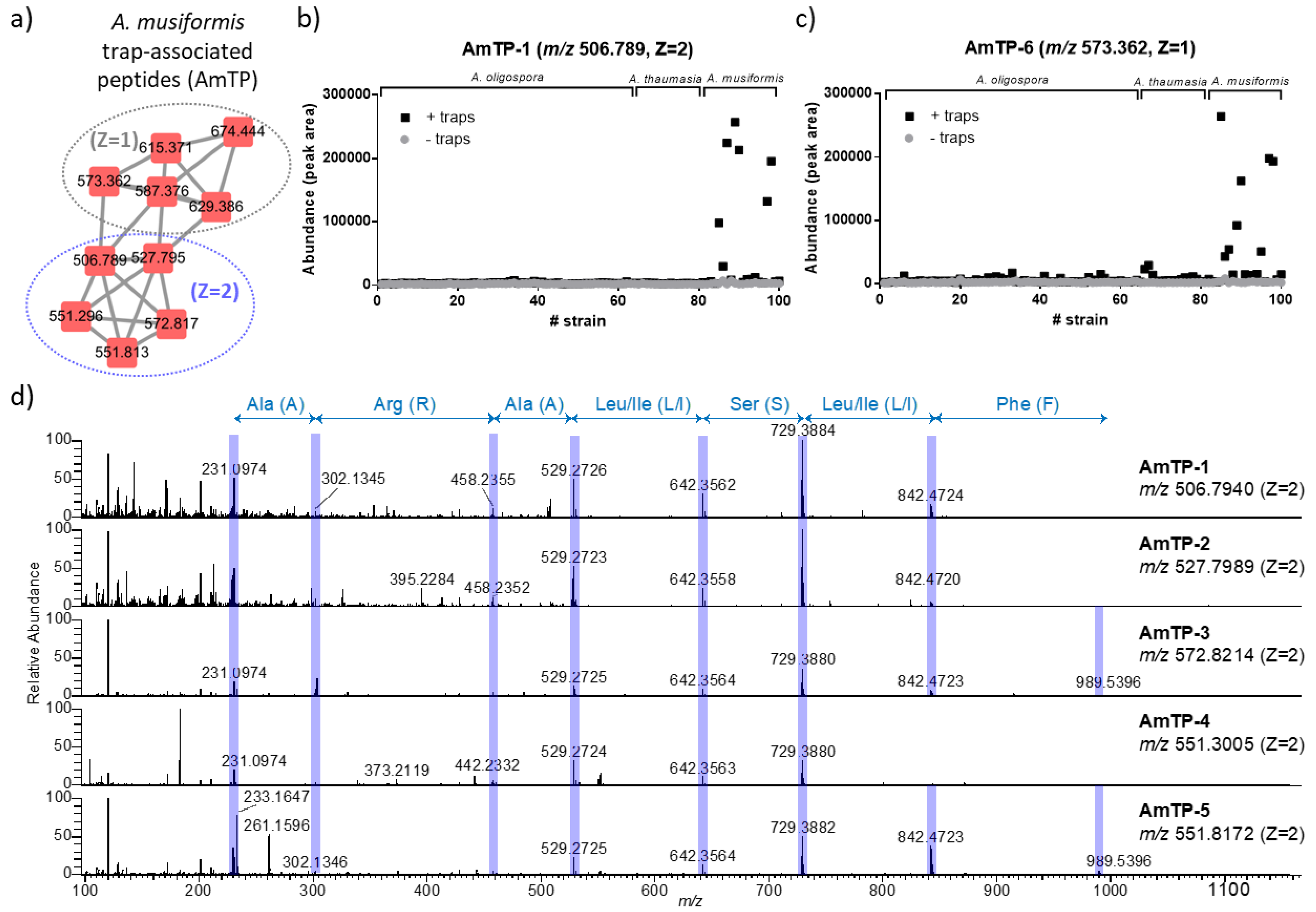
2.2. Annotation of Arthrobotrys musiformis Trap-Associated Peptides (AmTPs)
2.3. Identifying Fungal Metabolites Significantly Enriched in the Predatory Stage
2.3.1. Desferriferrichrome
2.3.2. Linoleyl Alcohol
2.3.3. Nonadecanamide
2.3.4. Citicoline
3. Discussion
4. Materials and Methods
4.1. Materials, Reagents, and Chemical Standards
4.2. Collection and Culture of Nematode-Trapping Fungi and Nematodes
4.3. Extraction of Fungal Metabolites
4.4. Untargeted LC–MS Metabolomics
4.5. Metabolomics Analysis With GNPS Molecular Networking
4.6. Quantitative Analysis and Identification of NTF Metabolites
4.7. Bioactivity Test
4.8. Data Availability
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [M+H] m/z 454.229 | [M+H] m/z 376.144 | [M+H] m/z 519.193 | [M+H] m/z 462.208 | [M+H] m/z 590.23 | [M+H] m/z 726.281 | [M+H] m/z 293.276 | |||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 110.07 | 11.61 | 57.07 | 1.62 | 72.08 | 15.06 | 60.05 | 24.86 | 72.08 | 44.2 | 65.14 | 0.97 | 56.05 | 4.93 |
| 156.08 | 9.8 | 69.07 | 6.42 | 84.04 | 24.18 | 70.07 | 12.95 | 84.04 | 21.28 | 65.14 | 3.3 | 57.07 | 3.33 |
| 166.06 | 10.09 | 72.08 | 3.18 | 88.04 | 100 | 84.04 | 15.17 | 88.04 | 83.5 | 70.07 | 0.58 | 58.07 | 4.44 |
| 196.13 | 6.19 | 81.07 | 2.41 | 91.06 | 9.05 | 86.1 | 7.72 | 102.06 | 86.46 | 86.1 | 0.29 | 71.09 | 1.91 |
| 212.16 | 5.64 | 83.09 | 2.05 | 102.06 | 73.8 | 96.04 | 10.21 | 130.05 | 19.95 | 97.56 | 0.28 | 74.06 | 6.37 |
| 231.15 | 4.45 | 86.1 | 8.99 | 118.09 | 12.04 | 97.03 | 6.18 | 158.04 | 53.11 | 565.21 | 0.31 | 75.06 | 2.32 |
| 249.16 | 100 | 95.09 | 1.89 | 130.05 | 29.73 | 98.06 | 9.01 | 185.06 | 17.05 | 579.23 | 0.23 | 88.08 | 5.04 |
| 250.15 | 4.38 | 120 | 2.56 | 158.04 | 45.8 | 116.07 | 94.24 | 188.07 | 3.73 | 606.24 | 0.74 | 89.08 | 2.6 |
| 250.16 | 10.61 | 120.1 | 7.36 | 167.06 | 11.96 | 119.09 | 20.38 | 189.12 | 11.48 | 608.25 | 0.3 | 118.09 | 1.74 |
| 255.17 | 20.37 | 144 | 5.73 | 181.06 | 7.06 | 120.09 | 12.2 | 217.08 | 14.18 | 610.21 | 0.26 | 122.08 | 3.34 |
| 276.17 | 33.82 | 157.1 | 2.12 | 183.11 | 13.45 | 126.05 | 65.61 | 227.07 | 6.37 | 623.22 | 1.07 | 123.08 | 2.47 |
| 305.15 | 12.93 | 163.1 | 2.47 | 184.07 | 13.44 | 127.04 | 6.27 | 245.08 | 100 | 624.22 | 0.38 | 124.09 | 3.11 |
| 321.23 | 11.79 | 166.1 | 1.59 | 185.06 | 20.54 | 135.08 | 5.59 | 246.08 | 3.92 | 666.26 | 4.72 | 242.25 | 6.44 |
| 322.18 | 93.05 | 177.1 | 4.68 | 217.08 | 21.39 | 138.05 | 51.83 | 255.06 | 4.11 | 667.27 | 1.69 | 243.25 | 5.65 |
| 323.18 | 10.95 | 207.1 | 4.85 | 229.12 | 22.28 | 144.07 | 69.8 | 273.07 | 53.79 | 712.28 | 0.41 | 244.25 | 23.1 |
| 393.21 | 9 | 259.1 | 1.81 | 245.08 | 68.71 | 168.07 | 17.13 | 374.12 | 6.93 | 725.27 | 0.93 | 245.26 | 2.09 |
| 410.24 | 89.14 | 375.4 | 3.72 | 273.07 | 36.49 | 186.08 | 72.9 | 384.1 | 5.27 | 726.28 | 100 | 292.27 | 100 |
| 411.24 | 11.98 | 376.1 | 100 | 299.06 | 21.93 | 204.09 | 100 | 402.11 | 62.56 | 727.28 | 22.3 | 293.28 | 20.39 |
| 454.23 | 49.59 | 377.2 | 18.14 | 344.14 | 6.37 | 305.03 | 5.16 | 403.12 | 5.49 | 735.83 | 0.24 | 293.29 | 2.07 |
| 455.23 | 7.81 | 377.3 | 5.29 | 402.11 | 8.41 | 347.14 | 5.82 | 501.18 | 5.61 | 738.61 | 0.2 | 294.28 | 5.76 |
| [M+H] m/z 317.191 | [M+H] m/z 372.311 | [M+H] m/z 688.326 | [M+H] m/z 768.539 | [M+2H] m/z 376.145 | [M+H] m/z 529.409 | [M+H] m/z 184.606 | |||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 69.07 | 6.11 | 67.05 | 23.85 | 65.22 | 13.43 | 69.07 | 13.05 | 57.07 | 2.51 | 57.07 | 10.97 | 60.08 | 5.03 |
| 70.07 | 9.3 | 69.07 | 31.65 | 70.07 | 11.57 | 71.09 | 6.61 | 69.07 | 9.72 | 67.05 | 8.06 | 86.1 | 20.62 |
| 72.08 | 100 | 71.09 | 52.72 | 86.06 | 85.99 | 81.07 | 12.78 | 72.08 | 5.09 | 69.07 | 7.42 | 95.06 | 5.27 |
| 74.06 | 6.13 | 72.08 | 23.47 | 113.07 | 20.09 | 83.09 | 16.59 | 81.07 | 3.39 | 70.07 | 10.95 | 108.08 | 13.07 |
| 86.06 | 30.19 | 81.07 | 50.84 | 114.06 | 11.62 | 86.1 | 11.62 | 83.09 | 2.97 | 71.05 | 16.6 | 110.07 | 75.47 |
| 86.1 | 27.21 | 83.09 | 50.58 | 127.09 | 22.25 | 95.09 | 14.21 | 86.1 | 9.58 | 71.09 | 8.26 | 112.04 | 6.53 |
| 87.04 | 10.43 | 86.1 | 91.67 | 128.07 | 54.43 | 97.1 | 18.65 | 95.09 | 2.55 | 72.08 | 38.37 | 117.98 | 4.69 |
| 104.07 | 39.82 | 95.09 | 48.82 | 131.08 | 68.37 | 107.09 | 8.48 | 120.08 | 6.65 | 81.07 | 22.06 | 125 | 10.85 |
| 136.08 | 10 | 97.1 | 67.3 | 145.1 | 26.85 | 109.1 | 9.18 | 157.1 | 3.1 | 84.08 | 6.72 | 131.59 | 10.63 |
| 173.13 | 6.72 | 98.06 | 30.11 | 173.09 | 31.49 | 111.12 | 9.99 | 163.07 | 7.07 | 86.1 | 80.38 | 132.08 | 9.37 |
| 186.08 | 28.5 | 99.08 | 48.21 | 188.1 | 27.67 | 119.09 | 100 | 177.09 | 13.41 | 95.09 | 100 | 136.08 | 13.1 |
| 200.09 | 9.53 | 107.09 | 22.87 | 214.08 | 14.98 | 120.09 | 7.67 | 207.1 | 13.88 | 120.08 | 18.97 | 152.6 | 19.85 |
| 201.12 | 9.93 | 109.1 | 25.99 | 230.11 | 16.93 | 135.08 | 25.38 | 269.14 | 2.37 | 123.08 | 38.29 | 153.1 | 100 |
| 219.13 | 10.51 | 121.1 | 24.21 | 287.13 | 18.35 | 184.07 | 86.22 | 283.15 | 2.41 | 141.09 | 26.44 | 154.11 | 8.56 |
| 232.14 | 9.22 | 125.1 | 55.11 | 302.15 | 11.01 | 185.08 | 9.87 | 375.3 | 4.42 | 159.1 | 55.03 | 156.08 | 57.32 |
| 260.14 | 7.84 | 189.11 | 67.43 | 344.16 | 29.83 | 212.2 | 6.58 | 375.33 | 2.35 | 173.12 | 23.28 | 161.6 | 12.15 |
| 316.19 | 6.39 | 238.25 | 21.94 | 345.18 | 21.88 | 319.3 | 12.97 | 376.14 | 100 | 184.07 | 7.27 | 166.1 | 11.22 |
| 316.2 | 60.68 | 277.22 | 24.3 | 516.24 | 27.3 | 336.33 | 12.52 | 376.25 | 3.34 | 283.26 | 13.97 | 184.07 | 64.13 |
| 317.13 | 9.87 | 280.26 | 66.94 | 688.33 | 100 | 354.34 | 15.36 | 377.15 | 17.36 | 313.27 | 65.21 | 184.11 | 4.52 |
| 317.2 | 20.15 | 294.24 | 100 | 689.33 | 21.33 | 355.32 | 7.48 | 377.32 | 6.1 | 341.27 | 8.24 | 308.17 | 8.36 |
| [M+H] m/z 209.128 | |
|---|---|
| m/z | Relative Intensity |
| 56.05 | 8.09 |
| 70.07 | 6.79 |
| 80.05 | 23.65 |
| 84.08 | 100 |
| 107.09 | 3.52 |
| 120.08 | 3.59 |
| 121.62 | 4.15 |
| 130.09 | 55.04 |
| 135.12 | 3.37 |
| 147.06 | 6.36 |
| 153.09 | 4.2 |
| 175.05 | 3.85 |
| 192.08 | 9.72 |
| 203.94 | 4.47 |
| 204.97 | 4.13 |
| 209.06 | 3.43 |
| 209.1 | 3.25 |
| 209.13 | 26.17 |
| 209.15 | 10.25 |
| 226.95 | 3.5 |
| [M+H] m/z 216.042 | [M+H] m/z 259.992 | [M+H] m/z 338.171 | [M+H] m/z 339.249 | [M+H] m/z 368.182 | [M+H] m/z 423.005 | [M+H] m/z 443.287 | [M+2H] m/z 344.662 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 70.07 | 7.5 | 70.07 | 16.21 | 69.07 | 2.71 | 58.07 | 0.47 | 57.07 | 25.69 | 70.03 | 100 | 70.07 | 50.76 | 57.07 | 11.17 |
| 72.08 | 16.1 | 72.08 | 31.36 | 70.07 | 9.57 | 59.07 | 0.1 | 70.07 | 81.62 | 70.07 | 29.12 | 72.08 | 100 | 70.07 | 38.89 |
| 84.04 | 5 | 84.04 | 21.93 | 72.08 | 8.7 | 60.08 | 1.15 | 72.08 | 64.89 | 72.08 | 21.37 | 74.06 | 5.76 | 71.09 | 11.23 |
| 86.1 | 2.74 | 84.08 | 33.27 | 81.07 | 2.78 | 70.07 | 0.12 | 74.06 | 57.6 | 74.02 | 17.66 | 84.04 | 8.47 | 72.08 | 33.61 |
| 116.07 | 20.21 | 86.1 | 71 | 84.04 | 3.6 | 84.08 | 0.07 | 84.04 | 27.38 | 74.06 | 5.61 | 84.08 | 11.12 | 84.04 | 11.02 |
| 129.07 | 4.51 | 115.09 | 9.04 | 84.08 | 4.54 | 86.1 | 0.08 | 84.08 | 29.26 | 84.04 | 8.23 | 86.1 | 76.85 | 86.06 | 100 |
| 130.05 | 20.18 | 116.07 | 20.55 | 86.1 | 21.37 | 95.06 | 0.08 | 86.1 | 100 | 84.08 | 13.13 | 98.98 | 6.12 | 86.1 | 43.92 |
| 134.06 | 3.41 | 126.05 | 9.66 | 110.07 | 2.4 | 104.07 | 0.14 | 110.07 | 26.36 | 86.1 | 36.16 | 100.08 | 12.28 | 87.06 | 17.66 |
| 135.07 | 3.26 | 126.09 | 7.41 | 113.07 | 6.87 | 104.11 | 100 | 112.08 | 22.14 | 88.04 | 7.25 | 110.07 | 9.15 | 100.04 | 13.69 |
| 147.08 | 5.15 | 129.1 | 10.25 | 115.09 | 6.99 | 104.14 | 0.09 | 113.07 | 27.09 | 102.06 | 4.38 | 120.08 | 10.91 | 113.07 | 21.62 |
| 153.01 | 5.74 | 132.1 | 14.99 | 120.04 | 9.3 | 105.1 | 0.17 | 120.08 | 86 | 104.11 | 4.9 | 128.11 | 81.72 | 114.06 | 30.06 |
| 155.07 | 2.54 | 135.07 | 18.18 | 120.08 | 100 | 105.11 | 3.08 | 121.03 | 38.73 | 110.07 | 7.63 | 129.1 | 9.46 | 115.05 | 19.51 |
| 157 | 17.74 | 143.12 | 8.76 | 121.08 | 5.53 | 110.07 | 0.16 | 130.09 | 78.38 | 120.08 | 14.81 | 136.08 | 8.86 | 116.07 | 16.69 |
| 170.04 | 100 | 185.13 | 7.07 | 129.1 | 2.98 | 112.9 | 0.27 | 136.08 | 57.1 | 126.09 | 5.53 | 145.13 | 9.1 | 126.09 | 11.16 |
| 171.04 | 5.8 | 186.11 | 8.94 | 143.08 | 4 | 114.89 | 0.07 | 157.1 | 22.37 | 129.1 | 8.25 | 199.18 | 20.37 | 127.09 | 13.31 |
| 181 | 9.86 | 189.13 | 7.45 | 166.09 | 7.52 | 156.04 | 0.2 | 166.09 | 45.91 | 130.05 | 5.16 | 200.09 | 9.25 | 128.07 | 21.61 |
| 199.02 | 25.56 | 197.13 | 12.81 | 191.12 | 47.59 | 156.08 | 0.13 | 175.11 | 81.37 | 135.07 | 17.89 | 216.17 | 23.44 | 131.08 | 91.89 |
| 215.14 | 2.63 | 200.95 | 22.27 | 192.12 | 3.9 | 192.02 | 0.34 | 203.1 | 60.27 | 136.08 | 8.14 | 216.21 | 32.01 | 173.09 | 16.18 |
| 216.04 | 8.76 | 213.99 | 100 | 219.11 | 15.14 | 193 | 0.18 | 221.13 | 28.81 | 156.08 | 4.42 | 217.12 | 5.35 | 188.1 | 18.46 |
| 217.1 | 5.68 | 224.95 | 12.12 | 237.12 | 11.49 | 207.01 | 0.15 | 223.11 | 79.2 | 164.05 | 16.17 | 229.15 | 11.2 | 228.13 | 15.99 |
| [M+H] m/z 250 | [M+H] m/z 262 | [M+H] m/z 271.2 | [M+H] m/z 323.2 | [M+H] m/z 354.24 | [M+H] m/z 361.217 | ||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 67.05 | 5.04 | 60.05 | 7.12 | 57.07 | 14.55 | 69.07 | 12.39 | 58.07 | 9.21 | 57.07 | 6.46 |
| 70.03 | 10.16 | 70.07 | 7.91 | 69.07 | 16.54 | 81.07 | 7.86 | 60.05 | 4.03 | 69.07 | 3.67 |
| 71.09 | 4.95 | 72.08 | 10.21 | 70.07 | 81.26 | 93.07 | 5.07 | 70.07 | 9.36 | 71.09 | 5.71 |
| 74.02 | 4.82 | 84.04 | 7.05 | 71.09 | 16.45 | 95.09 | 7.17 | 72.08 | 10.83 | 72.08 | 32.84 |
| 81.07 | 7.33 | 86.1 | 50.12 | 72.08 | 100 | 105.07 | 9.21 | 74.06 | 5.25 | 73.08 | 9.9 |
| 86.1 | 32.91 | 87.1 | 5.7 | 84.08 | 17.44 | 107.09 | 4.84 | 84.08 | 4.72 | 85.1 | 3.7 |
| 87.1 | 8.57 | 102.1 | 7.17 | 86.1 | 54.97 | 109.1 | 8.16 | 86.1 | 9.09 | 86.1 | 11.01 |
| 88.04 | 5.13 | 130.1 | 8.47 | 112.08 | 33.05 | 121.1 | 4.8 | 91.06 | 5.39 | 95.09 | 4.08 |
| 95.09 | 8.69 | 132.1 | 12.91 | 112.09 | 14.26 | 123.12 | 5.28 | 98.06 | 4.4 | 97.08 | 4.9 |
| 109.1 | 5.21 | 135.1 | 23.16 | 113.07 | 13.47 | 145.1 | 4.68 | 116.07 | 20.31 | 114.1 | 49.09 |
| 132.1 | 8.07 | 148.1 | 5.42 | 115.09 | 45.92 | 157.1 | 5.82 | 120.08 | 17.95 | 115.1 | 3.44 |
| 146.06 | 8.44 | 156.1 | 8.46 | 140.07 | 32.89 | 197.1 | 4.91 | 143.08 | 10.34 | 115.11 | 20.67 |
| 190.97 | 8.99 | 199 | 7.13 | 140.08 | 19.24 | 211.11 | 5.61 | 152.06 | 7 | 116.11 | 3.6 |
| 201.12 | 7.32 | 203 | 23.05 | 141.07 | 15.82 | 239.11 | 4.98 | 161.09 | 16.68 | 200.09 | 7.72 |
| 203.13 | 7.06 | 216 | 100 | 225.16 | 18.69 | 277.19 | 5.19 | 166.09 | 14.44 | 201.09 | 12.74 |
| 204 | 100 | 217 | 6.93 | 254.14 | 22.68 | 295.21 | 30.34 | 189.09 | 13.91 | 202.1 | 10.02 |
| 205 | 5.87 | 227 | 11.29 | 254.17 | 30.03 | 296.21 | 4.59 | 241.15 | 14.31 | 301.17 | 3.79 |
| 232.98 | 6.91 | 245 | 27.71 | 271.18 | 23.19 | 305.19 | 10.95 | 354.16 | 4.92 | 342.31 | 4.37 |
| 249.13 | 7.59 | 262 | 14.2 | 271.2 | 17.09 | 323.2 | 100 | 354.24 | 100 | 360.21 | 100 |
| 250 | 10.76 | 262.1 | 10.83 | 272.15 | 26.07 | 324.2 | 14.23 | 355.24 | 12.38 | 361.22 | 21.86 |
| [M+H] m/z 373.244 | [M+2H] m/z 390.099 | [M+H] m/z 443.287 | [M+H] m/z 519.265 | [M+H] m/z 550.446 | [M+H] m/z 945.327 | ||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 69.07 | 4.02 | 57.07 | 11.27 | 69.07 | 2.18 | 60.05 | 7.51 | 57.07 | 3.63 | 70.07 | 2.87 |
| 70.07 | 15.54 | 65.16 | 8.11 | 70.07 | 15.57 | 70.07 | 9.88 | 67.05 | 1.73 | 112.08 | 2.36 |
| 72.08 | 100 | 70.07 | 16.05 | 72.08 | 12.8 | 72.08 | 12.22 | 69.07 | 2.98 | 199.02 | 1.36 |
| 84.08 | 4.86 | 71.09 | 18.33 | 84.08 | 3.61 | 84.04 | 6.04 | 71.09 | 2.83 | 201.03 | 2.51 |
| 86.1 | 15.77 | 72.08 | 6.77 | 86.1 | 16.19 | 104.11 | 7.01 | 81.07 | 2.96 | 370.09 | 5.8 |
| 91.06 | 14.44 | 74.06 | 2.1 | 100.08 | 4.07 | 116.07 | 100 | 83.09 | 2.15 | 397.07 | 3.81 |
| 112.08 | 4.06 | 84.04 | 2.21 | 120.08 | 2.19 | 126.06 | 20.27 | 85.1 | 1.61 | 398.07 | 2.07 |
| 112.11 | 3.93 | 84.08 | 3.31 | 128.11 | 100 | 128.11 | 5.53 | 86.06 | 2.1 | 398.09 | 9.92 |
| 116.07 | 21.32 | 86.1 | 18.32 | 129.11 | 5.29 | 138.05 | 17.41 | 87.04 | 12.16 | 427.11 | 2.08 |
| 173.09 | 21.63 | 105.07 | 4.77 | 132.03 | 3.64 | 144.07 | 21.64 | 95.09 | 3.24 | 455.11 | 4.93 |
| 187.11 | 4.79 | 112.08 | 3.81 | 142.12 | 8 | 167.06 | 4.99 | 104.11 | 2 | 484.14 | 1.37 |
| 200.09 | 39.96 | 128.12 | 5.15 | 199.18 | 2.83 | 168.07 | 5.21 | 109.1 | 1.71 | 512.13 | 3.94 |
| 285.14 | 56.56 | 129.1 | 2.84 | 200.09 | 17.91 | 173.13 | 16.19 | 114.09 | 2.6 | 541.16 | 6.02 |
| 286.15 | 5.01 | 149.02 | 100 | 211.14 | 2.22 | 184.07 | 15.16 | 184.07 | 5.94 | 569.15 | 19.36 |
| 302.17 | 25.25 | 167.03 | 14.47 | 216.21 | 3.3 | 186.08 | 20.9 | 219.17 | 100 | 570.16 | 2.45 |
| 356.22 | 63.4 | 201.03 | 3.3 | 226.19 | 2.31 | 201.12 | 27.99 | 220.18 | 8.14 | 586.18 | 2.26 |
| 357.22 | 6.39 | 218.1 | 3.63 | 229.15 | 3.74 | 204.09 | 24.9 | 236.15 | 2.39 | 713.24 | 5.58 |
| 373.22 | 5.6 | 247.04 | 3.08 | 285.14 | 20.71 | 217.12 | 6.46 | 256.26 | 6.9 | 723.23 | 2.15 |
| 373.24 | 78.97 | 258.23 | 8.17 | 286.15 | 2.25 | 299.06 | 9.06 | 277.27 | 1.71 | 741.24 | 100 |
| 374.25 | 8.79 | 370.09 | 2.5 | 356.22 | 2.03 | 316.19 | 15.78 | 305.21 | 10.37 | 742.24 | 19.74 |
Appendix B
| m/z 413.276 [M+H] | m/z 750.558 [M+H] | m/z 314.207 [M+H] | m/z 455.189 [M+H] | m/z 484.318 [M+H] | m/z 498.328 [M+H] | m/z 528.302 [M+H] | |||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 70.07 | 100.00 | 53.00 | 0.37 | 70.07 | 66.12 | 70.07 | 80.81 | 70.07 | 100.00 | 70.07 | 100.00 | 70.07 | 100.00 |
| 72.08 | 5.78 | 57.07 | 0.91 | 71.07 | 2.18 | 72.08 | 6.66 | 72.08 | 33.04 | 72.08 | 47.48 | 72.08 | 17.11 |
| 86.10 | 34.36 | 71.09 | 12.07 | 72.08 | 25.07 | 86.10 | 23.27 | 86.10 | 75.91 | 86.10 | 30.67 | 86.10 | 36.53 |
| 141.10 | 19.95 | 72.09 | 0.31 | 86.10 | 25.57 | 112.05 | 74.89 | 120.08 | 16.10 | 116.07 | 11.04 | 132.10 | 5.93 |
| 169.10 | 25.46 | 86.10 | 0.43 | 97.03 | 7.25 | 120.08 | 7.94 | 136.06 | 18.74 | 120.08 | 7.16 | 137.05 | 43.24 |
| 169.13 | 6.44 | 88.11 | 100.00 | 98.06 | 1.38 | 136.08 | 8.58 | 137.05 | 81.88 | 137.05 | 36.15 | 155.08 | 22.53 |
| 183.15 | 20.40 | 89.12 | 2.73 | 136.06 | 15.04 | 141.10 | 17.40 | 141.10 | 34.31 | 143.12 | 13.32 | 169.13 | 10.14 |
| 197.13 | 6.89 | 327.08 | 0.54 | 137.05 | 2.82 | 169.10 | 23.64 | 155.08 | 10.22 | 155.08 | 18.15 | 183.15 | 20.99 |
| 203.14 | 5.66 | 383.14 | 2.37 | 169.13 | 12.06 | 183.15 | 7.95 | 169.10 | 60.80 | 169.13 | 12.85 | 187.07 | 7.86 |
| 211.14 | 19.26 | 439.20 | 6.07 | 173.13 | 7.58 | 211.14 | 7.55 | 169.13 | 14.20 | 171.11 | 12.32 | 197.13 | 17.04 |
| 212.03 | 7.89 | 440.21 | 0.91 | 197.13 | 14.13 | 215.14 | 55.14 | 183.15 | 30.28 | 183.15 | 13.32 | 211.14 | 25.60 |
| 212.96 | 6.53 | 495.27 | 11.15 | 201.12 | 5.67 | 216.14 | 8.35 | 197.13 | 18.12 | 197.13 | 14.23 | 243.13 | 13.07 |
| 228.01 | 17.04 | 496.27 | 2.14 | 215.14 | 100.00 | 229.15 | 14.15 | 203.14 | 10.32 | 211.14 | 15.63 | 284.12 | 20.52 |
| 229.15 | 33.77 | 551.33 | 5.12 | 216.14 | 7.28 | 454.21 | 43.52 | 211.14 | 22.77 | 229.15 | 92.38 | 286.11 | 7.27 |
| 282.18 | 5.73 | 552.33 | 1.17 | 223.02 | 2.35 | 454.27 | 8.37 | 215.14 | 18.01 | 230.16 | 8.54 | 310.21 | 13.22 |
| 300.19 | 20.49 | 607.39 | 4.46 | 226.99 | 5.35 | 455.19 | 100.00 | 229.15 | 11.73 | 236.15 | 6.99 | 397.21 | 9.25 |
| 329.03 | 7.05 | 608.39 | 1.17 | 314.06 | 1.52 | 455.21 | 22.15 | 282.18 | 28.30 | 268.17 | 10.35 | 415.22 | 23.04 |
| 345.00 | 26.35 | 663.45 | 4.69 | 314.09 | 2.29 | 455.25 | 7.12 | 300.19 | 17.27 | 328.22 | 5.82 | 528.22 | 5.95 |
| 413.14 | 7.87 | 664.46 | 1.31 | 314.21 | 20.46 | 456.19 | 12.23 | 395.26 | 15.04 | 498.33 | 14.44 | 528.30 | 30.11 |
| 413.28 | 17.60 | 750.56 | 0.34 | 315.21 | 1.96 | 456.28 | 6.95 | 484.31 | 25.52 | 498.38 | 6.56 | 528.34 | 8.45 |
| m/z 565.167 [M+H] | m/z 145.170 [M+H] | m/z 422.065 [M+H] | m/z 356.213 [M+H] | m/z 418.167 [M+H] | m/z 647.193 [M+H] | ||||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | ||
| 70.07 | 2.05 | 58.07 | 0.30 | 58.07 | 24.79 | 58.07 | 3.18 | 60.06 | 5.77 | 58.07 | 1.45 | ||
| 72.08 | 0.99 | 60.06 | 2.36 | 70.07 | 15.51 | 69.07 | 5.12 | 70.07 | 24.03 | 70.07 | 2.65 | ||
| 81.03 | 17.41 | 60.08 | 11.09 | 72.08 | 6.34 | 70.07 | 10.99 | 72.08 | 4.31 | 72.08 | 1.08 | ||
| 82.04 | 0.50 | 69.07 | 3.49 | 84.08 | 19.46 | 72.08 | 100.00 | 84.08 | 8.70 | 77.01 | 1.67 | ||
| 84.04 | 0.73 | 70.07 | 3.43 | 86.10 | 100.00 | 73.08 | 2.72 | 86.10 | 8.23 | 84.08 | 3.75 | ||
| 84.08 | 1.20 | 72.08 | 0.51 | 218.98 | 6.38 | 84.08 | 9.70 | 95.02 | 9.48 | 86.10 | 3.24 | ||
| 86.10 | 1.62 | 84.08 | 1.17 | 284.05 | 4.88 | 86.10 | 11.31 | 110.07 | 25.38 | 95.02 | 5.00 | ||
| 110.07 | 0.72 | 86.10 | 100.00 | 318.07 | 16.13 | 110.07 | 4.47 | 112.09 | 7.50 | 117.04 | 1.06 | ||
| 129.10 | 0.94 | 87.09 | 0.23 | 322.03 | 18.65 | 112.11 | 2.63 | 129.10 | 16.63 | 129.10 | 1.85 | ||
| 136.06 | 100.00 | 87.10 | 3.84 | 326.09 | 8.05 | 129.10 | 2.40 | 168.07 | 4.11 | 160.97 | 1.11 | ||
| 137.06 | 1.01 | 112.09 | 0.29 | 370.03 | 42.32 | 132.03 | 9.35 | 175.12 | 4.10 | 168.07 | 2.30 | ||
| 137.07 | 2.86 | 116.07 | 1.71 | 371.03 | 10.69 | 144.03 | 3.91 | 199.01 | 5.99 | 199.01 | 3.82 | ||
| 161.06 | 0.94 | 121.62 | 0.44 | 376.06 | 5.80 | 156.08 | 2.99 | 253.09 | 4.93 | 244.07 | 2.94 | ||
| 216.09 | 7.03 | 121.97 | 0.27 | 393.05 | 5.34 | 200.09 | 39.67 | 268.10 | 18.53 | 268.10 | 17.27 | ||
| 234.10 | 0.96 | 130.09 | 0.26 | 394.07 | 32.52 | 211.14 | 4.11 | 286.11 | 100.00 | 286.11 | 100.00 | ||
| 286.11 | 2.66 | 130.10 | 0.62 | 395.07 | 8.85 | 285.14 | 66.27 | 287.12 | 6.04 | 287.12 | 4.33 | ||
| 332.07 | 23.31 | 144.62 | 0.69 | 412.08 | 7.77 | 286.15 | 5.89 | 401.18 | 4.03 | 288.11 | 1.10 | ||
| 333.08 | 1.16 | 145.17 | 8.02 | 422.06 | 34.08 | 328.09 | 3.14 | 418.20 | 23.30 | 324.07 | 6.70 | ||
| 412.10 | 0.97 | 146.17 | 0.46 | 423.06 | 10.78 | 356.22 | 33.52 | 419.19 | 5.39 | 424.10 | 1.04 | ||
| 430.11 | 2.99 | 158.09 | 0.53 | 440.07 | 13.11 | 357.22 | 3.70 | 419.21 | 4.38 | 624.13 | 2.78 | ||
| m/z 485.61 [M+2H] | m/z 461.605 [M+2H] | m/z 423.172 [M+H] | m/z 227.033 [M+H] | m/z 299.543 [M+2H] | m/z 557.15 [M+H] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 70.07 | 2.55 | 70.07 | 1.96 | 60.05 | 39.12 | 56.05 | 6.60 | 58.07 | 10.05 | 70.07 | 18.93 |
| 84.08 | 4.14 | 84.08 | 2.12 | 74.06 | 15.31 | 70.07 | 2.38 | 59.07 | 3.47 | 72.08 | 4.35 |
| 86.10 | 2.45 | 86.10 | 1.93 | 84.04 | 6.56 | 76.04 | 100.00 | 64.81 | 5.41 | 81.03 | 35.64 |
| 95.02 | 1.37 | 97.03 | 24.82 | 84.08 | 9.19 | 77.04 | 1.44 | 84.08 | 4.97 | 84.04 | 3.49 |
| 97.03 | 2.54 | 111.04 | 3.90 | 88.04 | 18.53 | 84.04 | 0.85 | 86.10 | 4.23 | 84.08 | 10.79 |
| 112.05 | 100.00 | 112.05 | 100.00 | 102.06 | 6.26 | 84.08 | 1.10 | 117.04 | 2.87 | 86.10 | 8.83 |
| 113.05 | 3.41 | 113.05 | 3.52 | 120.07 | 38.68 | 106.00 | 6.99 | 136.06 | 3.16 | 88.04 | 2.93 |
| 129.10 | 2.27 | 175.12 | 8.76 | 126.06 | 7.30 | 110.07 | 0.66 | 197.00 | 12.14 | 110.07 | 3.91 |
| 136.08 | 3.44 | 176.12 | 2.38 | 130.05 | 10.30 | 116.07 | 1.14 | 256.07 | 9.91 | 112.05 | 34.75 |
| 152.07 | 3.76 | 180.09 | 40.43 | 144.07 | 44.50 | 124.01 | 76.85 | 268.10 | 29.65 | 120.08 | 2.57 |
| 154.09 | 4.63 | 181.09 | 2.53 | 153.07 | 5.56 | 125.01 | 1.15 | 270.04 | 57.43 | 129.10 | 6.41 |
| 205.06 | 2.47 | 193.05 | 2.21 | 171.08 | 42.90 | 153.03 | 0.72 | 270.54 | 10.02 | 152.06 | 100.00 |
| 223.07 | 4.82 | 208.07 | 3.46 | 189.09 | 100.00 | 181.03 | 34.17 | 271.04 | 5.07 | 153.06 | 3.13 |
| 286.11 | 4.48 | 286.11 | 8.54 | 190.09 | 5.28 | 181.06 | 0.75 | 289.99 | 5.72 | 161.06 | 7.55 |
| 303.04 | 1.67 | 287.12 | 4.06 | 199.07 | 45.80 | 182.03 | 1.16 | 298.55 | 2.78 | 268.10 | 7.76 |
| 321.05 | 2.84 | 288.11 | 3.44 | 207.10 | 57.59 | 209.02 | 6.84 | 299.55 | 100.00 | 270.10 | 3.71 |
| 340.03 | 1.58 | 309.04 | 1.84 | 217.08 | 17.48 | 209.13 | 1.42 | 300.05 | 21.83 | 286.11 | 27.18 |
| 357.04 | 2.28 | 420.08 | 2.57 | 272.12 | 5.38 | 226.15 | 1.22 | 300.54 | 7.37 | 311.20 | 2.69 |
| 358.04 | 3.48 | 461.23 | 2.46 | 286.10 | 21.43 | 227.03 | 18.21 | 308.00 | 7.28 | 348.07 | 6.19 |
| 616.09 | 1.27 | 462.01 | 2.23 | 387.15 | 12.65 | 228.04 | 0.80 | 336.07 | 3.14 | 446.11 | 3.61 |
| m/z 228.05 [M+H] | m/z 364.186 [M+H] | m/z 425.214 [M+H] | m/z 439.291 [M+H] | m/z 470.297 [M+H] | m/z 512.344 [M+H] | m/z 524.198 [M+H] | m/z 528.339 [M+H] | ||||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 69.07 | 5.24 | 70.07 | 100.00 | 70.07 | 33.28 | 70.07 | 100.00 | 70.07 | 100.00 | 70.07 | 17.78 | 70.07 | 37.47 | 70.07 | 100.00 |
| 70.07 | 38.05 | 72.08 | 7.86 | 72.08 | 4.78 | 86.10 | 39.92 | 72.08 | 15.36 | 72.08 | 3.17 | 72.08 | 22.17 | 72.08 | 42.51 |
| 72.08 | 6.57 | 74.06 | 9.28 | 86.10 | 21.60 | 98.06 | 7.23 | 86.10 | 53.40 | 86.10 | 31.06 | 84.04 | 28.51 | 86.10 | 50.14 |
| 74.02 | 68.32 | 86.10 | 8.31 | 136.08 | 6.31 | 99.08 | 12.62 | 137.05 | 89.26 | 112.05 | 100.00 | 84.08 | 44.53 | 137.05 | 68.47 |
| 84.04 | 6.20 | 120.08 | 30.70 | 169.13 | 10.14 | 116.07 | 12.67 | 141.10 | 16.30 | 113.05 | 3.08 | 86.10 | 49.10 | 155.08 | 9.86 |
| 84.08 | 20.30 | 166.09 | 7.85 | 183.15 | 4.14 | 121.06 | 14.55 | 169.10 | 26.85 | 116.07 | 3.23 | 87.06 | 13.53 | 169.13 | 29.28 |
| 86.10 | 17.95 | 168.07 | 6.72 | 197.13 | 8.95 | 126.05 | 7.29 | 169.13 | 7.23 | 130.10 | 4.88 | 88.04 | 23.74 | 183.15 | 30.33 |
| 109.03 | 4.54 | 171.11 | 7.11 | 211.14 | 5.02 | 138.09 | 9.20 | 183.15 | 5.88 | 137.05 | 73.35 | 102.06 | 27.32 | 197.13 | 35.50 |
| 110.07 | 8.99 | 172.99 | 9.11 | 215.14 | 3.84 | 159.10 | 4.78 | 187.11 | 21.32 | 143.12 | 4.97 | 110.07 | 24.31 | 211.14 | 46.04 |
| 129.10 | 12.54 | 195.98 | 8.17 | 229.15 | 5.61 | 183.15 | 52.35 | 197.13 | 10.84 | 155.08 | 5.56 | 120.08 | 82.38 | 229.15 | 10.12 |
| 136.04 | 22.94 | 199.11 | 9.68 | 233.13 | 6.30 | 184.15 | 3.76 | 203.14 | 7.73 | 171.11 | 9.36 | 129.10 | 52.55 | 243.13 | 8.70 |
| 148.98 | 5.36 | 217.12 | 52.56 | 263.14 | 5.78 | 211.14 | 67.36 | 211.14 | 4.49 | 183.15 | 3.08 | 166.09 | 100.00 | 284.12 | 8.03 |
| 155.03 | 77.72 | 217.13 | 11.88 | 312.15 | 6.84 | 212.15 | 5.11 | 214.15 | 4.10 | 211.14 | 3.64 | 181.06 | 22.08 | 286.11 | 21.72 |
| 156.04 | 4.36 | 245.13 | 9.35 | 326.21 | 5.41 | 229.15 | 37.89 | 229.15 | 71.45 | 229.15 | 6.44 | 199.07 | 14.12 | 310.21 | 32.24 |
| 156.08 | 4.19 | 245.98 | 6.43 | 354.16 | 3.91 | 324.23 | 11.61 | 230.16 | 4.33 | 244.09 | 8.12 | 217.08 | 73.26 | 415.22 | 9.90 |
| 182.04 | 100.00 | 263.14 | 16.43 | 425.20 | 9.48 | 326.21 | 33.32 | 245.11 | 8.40 | 245.19 | 5.10 | 223.11 | 20.85 | 415.25 | 9.10 |
| 183.05 | 6.15 | 346.08 | 8.62 | 425.22 | 100.00 | 327.21 | 4.64 | 268.17 | 11.39 | 268.17 | 4.83 | 245.08 | 62.37 | 423.30 | 8.99 |
| 211.02 | 5.99 | 364.09 | 9.45 | 425.24 | 11.34 | 439.23 | 14.52 | 300.19 | 10.40 | 284.20 | 8.97 | 280.13 | 37.69 | 528.28 | 8.08 |
| 228.05 | 11.02 | 364.19 | 18.72 | 425.27 | 4.51 | 439.29 | 24.98 | 381.25 | 4.37 | 399.26 | 2.97 | 302.10 | 27.16 | 528.30 | 11.75 |
| 228.23 | 14.22 | 365.18 | 5.85 | 426.22 | 14.76 | 440.29 | 4.56 | 470.30 | 9.64 | 512.34 | 4.88 | 359.12 | 26.94 | 528.34 | 26.31 |
| m/z 532.144 [M+H] | m/z 567.35 [M+H] | m/z 599.34 [M+H] | m/z 617.182 [M+H] | m/z 618.155 [M+H] | m/z 347.653 [M+2H] | m/z 529.775 [M+2H] | |||||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | ||
| 58.07 | 0.74 | 70.07 | 65.23 | 70.07 | 100.00 | 58.07 | 0.76 | 70.07 | 2.45 | 58.07 | 27.46 | 84.08 | 8.41 | ||
| 70.07 | 3.38 | 86.10 | 21.66 | 86.10 | 44.91 | 70.07 | 1.26 | 72.08 | 1.04 | 70.07 | 50.18 | 86.10 | 12.23 | ||
| 72.08 | 2.46 | 116.07 | 12.41 | 137.05 | 27.41 | 72.08 | 0.74 | 84.08 | 1.76 | 72.08 | 45.56 | 110.07 | 4.69 | ||
| 81.03 | 40.01 | 141.10 | 8.06 | 155.08 | 44.56 | 77.01 | 1.49 | 86.10 | 1.65 | 84.04 | 19.65 | 120.08 | 5.07 | ||
| 82.04 | 1.13 | 152.06 | 100.00 | 183.15 | 20.06 | 81.03 | 0.76 | 97.03 | 3.07 | 84.08 | 34.24 | 126.06 | 5.00 | ||
| 84.04 | 1.40 | 169.10 | 15.10 | 187.07 | 6.82 | 84.08 | 0.98 | 125.03 | 1.82 | 86.10 | 100.00 | 129.10 | 14.21 | ||
| 84.08 | 2.28 | 183.15 | 26.43 | 197.13 | 6.14 | 86.10 | 1.94 | 129.10 | 1.48 | 88.04 | 12.81 | 141.10 | 12.70 | ||
| 86.10 | 5.32 | 197.13 | 8.24 | 203.14 | 8.74 | 95.02 | 4.73 | 139.05 | 1.99 | 110.07 | 40.86 | 147.11 | 4.39 | ||
| 95.02 | 1.21 | 211.14 | 28.46 | 211.14 | 15.70 | 97.03 | 0.49 | 155.04 | 2.09 | 129.10 | 19.66 | 169.10 | 12.91 | ||
| 102.06 | 0.74 | 214.15 | 10.25 | 256.13 | 10.45 | 112.05 | 0.76 | 167.04 | 0.97 | 155.08 | 25.91 | 175.12 | 18.64 | ||
| 110.07 | 0.92 | 229.15 | 12.39 | 268.17 | 7.70 | 117.04 | 0.88 | 180.09 | 100.00 | 181.06 | 68.76 | 226.15 | 4.24 | ||
| 112.05 | 100.00 | 240.13 | 7.80 | 284.12 | 63.19 | 129.10 | 0.74 | 181.09 | 1.01 | 242.04 | 14.60 | 246.16 | 34.96 | ||
| 113.05 | 2.51 | 308.10 | 8.00 | 300.16 | 7.19 | 136.06 | 2.11 | 181.09 | 4.18 | 256.09 | 29.04 | 268.10 | 11.23 | ||
| 120.08 | 1.25 | 324.07 | 12.19 | 369.21 | 15.05 | 168.07 | 2.57 | 191.04 | 3.66 | 285.14 | 18.03 | 286.11 | 100.00 | ||
| 127.05 | 1.19 | 326.21 | 17.99 | 397.21 | 39.58 | 199.01 | 3.99 | 209.06 | 6.76 | 286.11 | 19.53 | 287.12 | 4.57 | ||
| 129.10 | 1.67 | 566.26 | 13.11 | 482.30 | 6.13 | 244.07 | 2.55 | 213.02 | 1.75 | 322.07 | 18.01 | 301.15 | 4.33 | ||
| 207.08 | 4.40 | 567.26 | 11.19 | 486.26 | 17.44 | 268.10 | 16.28 | 227.07 | 6.58 | 324.06 | 13.05 | 445.73 | 5.51 | ||
| 268.10 | 0.69 | 567.29 | 9.16 | 510.29 | 8.89 | 269.11 | 0.61 | 245.08 | 0.90 | 326.09 | 31.72 | 529.27 | 5.27 | ||
| 286.11 | 2.08 | 567.35 | 7.19 | 599.34 | 45.88 | 286.11 | 100.00 | 286.11 | 4.04 | 356.22 | 21.41 | 529.30 | 12.87 | ||
| 323.06 | 1.57 | 568.27 | 25.51 | 600.34 | 9.91 | 287.12 | 4.65 | 439.07 | 1.40 | 373.24 | 71.91 | 531.29 | 7.68 | ||
| m/z 347.644 [M+2H] | m/z 297.120 [M+2H] | m/z 514.287 [M+H] | m/z 597.360 [M+H] | m/z 580.015 [M+H] | m/z 593.241 [M+H] | ||||||
| m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity | m/z | Relative Intensity |
| 56.05 | 13.56 | 70.03 | 8.79 | 70.07 | 100.00 | 70.07 | 100.00 | 97.03 | 15.10 | 70.07 | 10.95 |
| 60.05 | 10.27 | 74.02 | 10.53 | 72.08 | 24.34 | 86.10 | 26.87 | 136.06 | 83.03 | 84.08 | 56.74 |
| 70.07 | 26.30 | 74.06 | 12.44 | 86.10 | 33.61 | 98.06 | 8.56 | 172.95 | 14.16 | 85.03 | 7.65 |
| 72.08 | 10.67 | 84.08 | 34.93 | 127.09 | 8.60 | 126.05 | 7.99 | 178.07 | 40.52 | 86.10 | 9.17 |
| 74.02 | 19.68 | 86.10 | 5.55 | 141.10 | 8.93 | 155.08 | 20.01 | 232.08 | 22.13 | 104.11 | 10.02 |
| 74.06 | 74.58 | 88.04 | 100.00 | 155.08 | 26.24 | 183.15 | 31.82 | 234.91 | 12.70 | 129.10 | 100.00 |
| 84.04 | 18.40 | 104.11 | 28.78 | 169.10 | 12.12 | 211.14 | 27.26 | 250.09 | 19.28 | 139.09 | 7.69 |
| 84.08 | 66.67 | 116.03 | 11.45 | 169.13 | 11.17 | 242.00 | 5.29 | 250.96 | 13.55 | 184.11 | 8.55 |
| 86.10 | 34.00 | 129.10 | 33.50 | 183.15 | 11.60 | 268.17 | 12.83 | 252.92 | 77.68 | 189.09 | 7.85 |
| 88.04 | 95.68 | 134.04 | 43.91 | 197.13 | 9.91 | 324.07 | 6.35 | 268.97 | 12.61 | 209.09 | 21.33 |
| 102.06 | 10.93 | 136.06 | 26.23 | 211.14 | 8.00 | 329.03 | 8.07 | 270.93 | 22.54 | 217.08 | 10.30 |
| 116.03 | 21.93 | 136.08 | 8.40 | 228.01 | 7.00 | 337.22 | 5.56 | 290.00 | 17.53 | 226.12 | 36.56 |
| 129.10 | 79.14 | 144.07 | 6.84 | 229.15 | 12.77 | 345.00 | 8.44 | 330.93 | 27.16 | 244.13 | 20.69 |
| 130.05 | 11.67 | 177.03 | 15.72 | 231.10 | 12.30 | 365.22 | 8.32 | 348.94 | 16.85 | 272.12 | 9.60 |
| 134.04 | 100.00 | 189.09 | 6.87 | 243.98 | 7.96 | 387.22 | 15.67 | 351.95 | 26.59 | 286.11 | 8.62 |
| 217.08 | 15.77 | 209.09 | 7.25 | 254.15 | 6.61 | 478.30 | 9.06 | 386.02 | 14.80 | 308.10 | 8.33 |
| 226.12 | 13.60 | 217.08 | 5.43 | 268.17 | 8.56 | 484.28 | 33.79 | 404.03 | 18.08 | 357.10 | 7.94 |
| 230.08 | 12.52 | 226.12 | 11.46 | 360.01 | 8.39 | 485.28 | 4.74 | 465.98 | 60.22 | 359.16 | 18.95 |
| 326.09 | 11.98 | 236.99 | 7.77 | 401.20 | 10.75 | 597.36 | 25.45 | 483.99 | 100.00 | 460.20 | 7.61 |
| 347.17 | 28.71 | 296.07 | 9.84 | 514.29 | 22.25 | 598.36 | 4.95 | 507.01 | 16.47 | 593.24 | 46.09 |
References
- Barron, G.L. The nematode-destroying fungi. In The Nematode-Destroying Fungi; Canadian Biological Publications Ltd.: Guelph, ON, Canada, 1977. [Google Scholar]
- Nordbring-Hertz, B.; Jansson, H.B.; Tunlid, A. Nematophagous fungi. In Els; John Wiley & Sons, Ltd.: Chichester, UK, 2011. [Google Scholar] [CrossRef]
- Chitwood, D.J. Nematicides. In Encyclopedia of Agrochemicals; Plimmer, J.R., Gammon, D.W., Ragsdale, N.A., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2003. [Google Scholar] [CrossRef]
- Nicol, J.M.; Turner, S.J.; Coyne, D.L.; Nijs, L.D.; Hockland, S.; Maafi, Z.T. Current nematode threats to world agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J., Gheysen, G., Fenoll, C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 21–43. [Google Scholar] [CrossRef]
- Hsueh, Y.-P.; Mahanti, P.; Schroeder, F.C.; Sternberg, P.W. Nematode-trapping fungi eavesdrop on nematode pheromones. Curr. Biol. 2013, 23, 83–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, X.-M.; Zhang, K.-Q. Arthrobotrys oligospora: A model organism for understanding the interaction between fungi and nematodes. Mycology 2011, 2, 59–78. [Google Scholar] [CrossRef]
- Yang, J.; Wang, L.; Ji, X.; Feng, Y.; Li, X.; Zou, C.; Xu, J.; Ren, Y.; Mi, Q.; Wu, J.; et al. Genomic and proteomic analyses of the fungus Arthrobotrys oligospora provide insights into nematode-trap formation. PLoS Pathog. 2011, 7, e1002179. [Google Scholar] [CrossRef] [Green Version]
- Youssar, L.; Wernet, V.; Hensel, N.; Yu, X.; Hildebrand, H.-G.; Schreckenberger, B.; Kriegler, M.; Hetzer, B.; Frankino, P.; Dillin, A.; et al. Intercellular communication is required for trap formation in the nematode-trapping fungus Duddingtonia flagrans. PLoS Genet. 2019, 15, e1008029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Liu, Z.; Liu, L.; Li, J.; Gao, H.; Yang, J.; Zhang, K.-Q. The nitrate assimilation pathway is involved in the trap formation of Arthrobotrys oligospora, a nematode-trapping fungus. Fungal Genet. Biol. 2016, 92, 33–39. [Google Scholar] [CrossRef]
- Hsueh, Y.-P.; Gronquist, M.R.; Schwarz, E.M.; Nath, R.D.; Lee, C.-H.; Gharib, S.; Schroeder, F.C.; Sternberg, P.W. Nematophagous fungus Arthrobotrys oligospora mimics olfactory cues of sex and food to lure its nematode prey. eLife 2017, 6, e20023. [Google Scholar] [CrossRef]
- Wang, B.-L.; Chen, Y.-H.; He, J.-N.; Xue, H.-X.; Yan, N.; Zeng, Z.-J.; Bennett, J.W.; Zhang, K.-Q.; Niu, X.-M. Integrated metabolomics and morphogenesis reveal volatile signaling of the nematode-trapping fungus Arthrobotrys oligospora. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-T.; Vidal-Diez de Ulzurrun, G.; Gonçalves, A.P.; Lin, H.-C.; Chang, C.-W.; Huang, T.-Y.; Chen, S.-A.; Lai, C.-K.; Isheng, J.T.; Schroeder, C.F.; et al. Natural diversity in the predatory behavior facilitates the establishment of a new robust model strain for nematode-trapping fungi. Proc. Natl. Acad. Sci. USA 2020, in press. [Google Scholar]
- Vidal-Diez de Ulzurrun, G.; Hsueh, Y.-P. Predator-prey interactions of nematode-trapping fungi and nematodes: Both sides of the coin. Appl. Microbiol. Biotechnol. 2018, 102, 3939–3949. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Kuo, T.-H.; Huang, H.-C.; Hsu, C.-C. Mass spectrometry imaging guided molecular networking to expedite discovery and structural analysis of agarwood natural products. Anal. Chim. Acta 2019, 1080, 95–103. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Melnik, A.V.; Koyama, N.; Lu, X.; Schorn, M.; Fang, J.; Aguinaldo, K.; Lincecum, T.L., Jr.; Ghequire, M.G.; Carrion, V.J.; et al. Indexing the Pseudomonas specialized metabolome enabled the discovery of poaeamide b and the bananamides. Nat. Microbiol. 2016, 2, 16197. [Google Scholar] [CrossRef] [Green Version]
- Emery, T. Role of ferrichrome as a ferric ionophore in Ustilago sphaerogen. Biochemistry 1971, 10, 1483–1488. [Google Scholar] [CrossRef] [PubMed]
- Haselwandter, K.; Häninger, G.; Ganzera, M. Hydroxamate siderophores of the ectomycorrhizal fungi Suillus granulatus and S. luteus. BioMetals 2011, 24, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Mascuch, S.J.; Moree, W.J.; Hsu, C.-C.; Turner, G.G.; Cheng, T.L.; Blehert, D.S.; Kilpatrick, A.M.; Frick, W.F.; Meehan, M.J.; Dorrestein, P.C.; et al. Direct detection of fungal siderophores on bats with white-nose syndrome via fluorescence microscopy-guided ambient ionization mass spectrometry. PLoS ONE 2015, 10, e0119668. [Google Scholar] [CrossRef] [PubMed]
- Hannauer, M.; Barda, Y.; Mislin, G.L.A.; Shanzer, A.; Schalk, I.J. The ferrichrome uptake pathway in Pseudomonas aeruginosa involves an iron release mechanism with acylation of the siderophore and recycling of the modified desferrichrome. J. Bacteriol. 2010, 192, 1212. [Google Scholar] [CrossRef] [Green Version]
- Kradel, J.; Pommer, E.H.; Effland, H. Further results with tridemorph (N-Tridecyl-2, 6-dimethyl-morpholin) against cereal mildew (Erysiphe graminis). Meded. Rijksfac. Landbouwwet. Gent. 1969, 34, 772–778. [Google Scholar]
- McMaster, C.R.; Bell, R.M. Phosphatidylcholine biosynthesis via the CDP-choline pathway in saccharomyces cerevisiae. Multiple mechanisms of regulation. J. Biol. Chem. 1994, 269, 14776–14783. [Google Scholar]
- Di Marzo, V. ‘Endocannabinoids’ and other fatty acid derivatives with cannabimimetic properties: Biochemistry and possible physiopathological relevance. Biochim. Biophys. Acta-Lipids Lipid Metab. 1998, 1392, 153–175. [Google Scholar] [CrossRef]
- McKinney, M.K.; Cravatt, B.F. Structure and function of fatty acid amide hydrolase. Annu. Rev. Biochem. 2005, 74, 411–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, W.B. Fatty aldehyde and fatty alcohol metabolism: Review and importance for epidermal structure and function. Biochim. Biophys. Actamol. Cell. Biol. Lipids 2014, 1841, 377–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paltauf, F. Ether lipids in biomembranes. Chem. Phys. Lipids 1994, 74, 101–139. [Google Scholar] [CrossRef]
- Su, K.L.; Schmid, H.H.O. Metabolism of long-chain polyunsaturated alcohols in myelinating brain. J. Lipid Res. 1972, 13, 452–457. [Google Scholar] [PubMed]
- Stadler, M.; Anke, H.; Sterner, O. Linoleic acid—The nematicidal principle of several nematophagous fungi and its production in trap-forming submerged cultures. Arch. Microbiol. 1993, 160, 401–405. [Google Scholar] [CrossRef]
- Khan, M.R.; Mohidin, F.A.; Khan, U.; Ahamad, F. Native Pseudomonas spp. Suppressed the root-knot nematode in in vitro and in vivo, and promoted the nodulation and grain yield in the field grown mungbean. Biol. Control. 2016, 101, 159–168. [Google Scholar] [CrossRef]
- Kirienko, N.V.; Kirienko, D.R.; Larkins-Ford, J.; Wählby, C.; Ruvkun, G.; Ausubel, F.M. Pseudomonas aeruginosa disrupts Caenorhabditis elegans iron homeostasis, causing a hypoxic response and death. Cell Host Microbe 2013, 13, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Proença, D.N.; Heine, T.; Senges, C.H.R.; Bandow, J.E.; Morais, P.V.; Tischler, D. Bacterial metabolites produced under iron limitation kill pinewood nematode and attract Caenorhabditis elegans. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Autenrieth, I.; Hantke, K.; Heesemann, J. Immunosuppression of the host and delivery of iron to the pathogen: A possible dual role of siderophores in the pathogenesis of microbial infections? Med. Microbiol. Immunol. 1991, 180, 135–141. [Google Scholar] [CrossRef]
- Van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; et al. The Natural Products Atlas: An open access knowledge base for microbial natural products discovery. ACS Cent. Sci. 2019, 5, 1824–1833. [Google Scholar] [CrossRef] [Green Version]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Jarmusch, A.K.; Vargas, F.; Aksenov, A.A.; Gauglitz, J.M.; Weldon, K.; Petras, D.; da Silva, R.; Quinn, R.; Melnik, A.V.; et al. Mass spectrometry searches using MASST. Nat. Biotechnol. 2020, 38, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Gray, N.F. Ecology of nematophagous fungi: Methods of collection, isolation and maintenance of predatory and endoparasitic fungi. Mycopathologia 1984, 86, 143–153. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haug, K.; Cochrane, K.; Nainala, V.C.; Williams, M.; Chang, J.; Jayaseelan, K.V.; O’Donovan, C. Metabolights: A resource evolving in response to the needs of its scientific community. Nucleic Acids Res. 2020, 48, D440–D444. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, T.-H.; Yang, C.-T.; Chang, H.-Y.; Hsueh, Y.-P.; Hsu, C.-C. Nematode-Trapping Fungi Produce Diverse Metabolites during Predator–Prey Interaction. Metabolites 2020, 10, 117. https://doi.org/10.3390/metabo10030117
Kuo T-H, Yang C-T, Chang H-Y, Hsueh Y-P, Hsu C-C. Nematode-Trapping Fungi Produce Diverse Metabolites during Predator–Prey Interaction. Metabolites. 2020; 10(3):117. https://doi.org/10.3390/metabo10030117
Chicago/Turabian StyleKuo, Ting-Hao, Ching-Ting Yang, Hsin-Yuan Chang, Yen-Ping Hsueh, and Cheng-Chih Hsu. 2020. "Nematode-Trapping Fungi Produce Diverse Metabolites during Predator–Prey Interaction" Metabolites 10, no. 3: 117. https://doi.org/10.3390/metabo10030117
APA StyleKuo, T. -H., Yang, C. -T., Chang, H. -Y., Hsueh, Y. -P., & Hsu, C. -C. (2020). Nematode-Trapping Fungi Produce Diverse Metabolites during Predator–Prey Interaction. Metabolites, 10(3), 117. https://doi.org/10.3390/metabo10030117