Relation of Whole Blood Amino Acid and Acylcarnitine Metabolome to Age, Sex, BMI, Puberty, and Metabolic Markers in Children and Adolescents

 ,
,  , ,
, , 

Abstract
:1. Introduction
2. Results
2.1. Influence of Age and Sex on AA and AC Levels
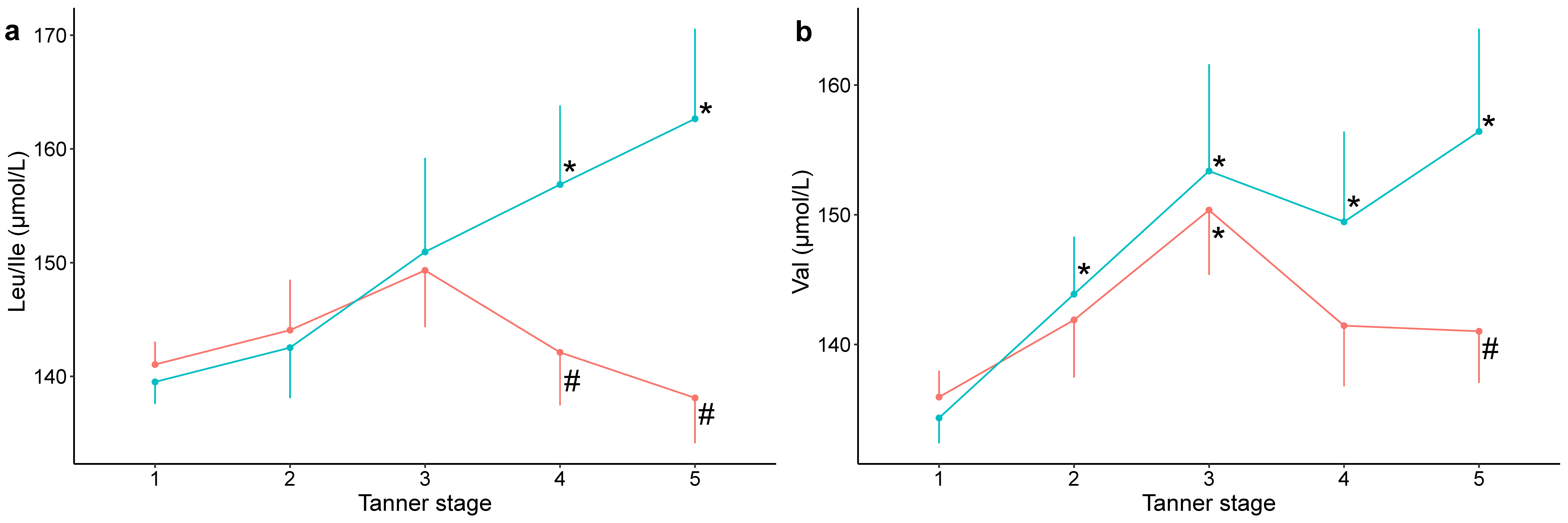
2.2. Associations of AA/AC Levels with BMI and Puberty
2.3. Correlations between AA and AC Levels and Metabolic Markers
3. Discussion
4. Materials and Methods
4.1. Study Population and Design
4.2. Sample Pretreatment and Analysis
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newgard, C.B. Metabolomics and Metabolic Diseases: Where do we stand? Cell Metab. 2017, 25, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millington, D.S. The Role of Technology in Newborn Screening. N. C. Med. J. 2019, 80, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Sweetman, L.; Millington, D.S.; Therrell, B.L.; Hannon, W.H.; Popovich, B.; Watson, M.S.; Mann, M.Y.; Lloyd-Puryear, M.A.; van Dyck, P.C. Naming and Counting Disorders (Conditions) Included in Newborn Screening Panels. Pediatrics 2006, 117, S308–S314. [Google Scholar] [CrossRef] [Green Version]
- Huffman, K.M.; Shah, S.H.; Stevens, R.D.; Bain, J.R.; Muehlbauer, M.; Slentz, C.A.; Tanner, C.J.; Kuchibhatla, M.; Houmard, J.A.; Newgard, C.B.; et al. Relationships Between Circulating Metabolic Intermediates and Insulin Action in Overweight to Obese, Inactive Men and Women. Diabetes Care 2009, 32, 1678–1683. [Google Scholar] [CrossRef] [Green Version]
- Owei, I.; Umekwe, N.; Stentz, F.; Wan, J.; Dagogo-Jack, S. Amino acid signature predictive of incident prediabetes: A case-control study nested within the longitudinal pathobiology of prediabetes in a biracial cohort. Metabolism 2019, 98, 76–83. [Google Scholar] [CrossRef]
- Shah, S.H.; Hauser, E.R.; Bain, J.R.; Muehlbauer, M.J.; Haynes, C.; Stevens, R.D.; Wenner, B.R.; Dowdy, Z.E.; Granger, C.B.; Ginsburg, G.S.; et al. High heritability of metabolomic profiles in families burdened with premature cardiovascular disease. Mol. Syst. Biol. 2009, 5, 258. [Google Scholar] [CrossRef]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite Profiles and the Risk of Developing Diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef]
- Frohnert, B.I.; Rewers, M.J. Metabolomics in childhood diabetes. Pediatr. Diabetes 2016, 17, 3–14. [Google Scholar] [CrossRef]
- Perng, W.; Tang, L.; Song, P.X.K.; Tellez-Rojo, M.M.; Cantoral, A.; Peterson, K.E. Metabolomic profiles and development of metabolic risk during the pubertal transition: A prospective study in the ELEMENT Project. Pediatr. Res. 2019, 85, 262–268. [Google Scholar] [CrossRef]
- Lepage, N.; McDonald, N.; Dallaire, L.; Lambert, M. Age-specific distribution of plasma amino acid concentrations in a healthy pediatric population. Clin. Chem. 1997, 43, 2397–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teodoro-Morrison, T.; Kyriakopoulou, L.; Chen, Y.K.; Raizman, J.E.; Bevilacqua, V.; Chan, M.K.; Wan, B.; Yazdanpanah, M.; Schulze, A.; Adeli, K. Dynamic biological changes in metabolic disease biomarkers in childhood and adolescence: A CALIPER study of healthy community children. Clin. Biochem. 2015, 48, 828–836. [Google Scholar] [CrossRef]
- Uaariyapanichkul, J.; Chomtho, S.; Suphapeetiporn, K.; Shotelersuk, V.; Punnahitananda, S.; Chinjarernpan, P.; Suteerojntrakool, O. Age-Related Reference Intervals for Blood Amino Acids in Thai Pediatric Population Measured by Liquid Chromatography Tandem Mass Spectrometry. J. Nutr. Metab. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulain, T.; Baber, R.; Vogel, M.; Pietzner, D.; Kirsten, T.; Jurkutat, A.; Hiemisch, A.; Hilbert, A.; Kratzsch, J.; Thiery, J.; et al. The LIFE Child study: A population-based perinatal and pediatric cohort in Germany. Eur. J. Epidemiol. 2017, 32, 145–158. [Google Scholar] [CrossRef]
- Felig, P.; Marliss, E.; Cahill, G.F. Plasma Amino Acid Levels and Insulin Secretion in Obesity. N. Engl. J. Med. 1969, 281, 811–816. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, T.M. The Complex Role of Branched Chain Amino Acids in Diabetes and Cancer. Metabolites 2013, 3, 931–945. [Google Scholar] [CrossRef] [Green Version]
- McCormack, S.E.; Shaham, O.; McCarthy, M.A.; Deik, A.A.; Wang, T.J.; Gerszten, R.E.; Clish, C.B.; Mootha, V.K.; Grinspoon, S.K.; Fleischman, A. Circulating Branched-chain Amino Acid Concentrations Are Associated with Obesity and Future Insulin Resistance in Children and Adolescents. Pediatr. Obes. 2013, 8, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Perng, W.; Gillman, M.W.; Fleisch, A.F.; Michalek, R.D.; Watkins, S.M.; Isganaitis, E.; Patti, M.-E.; Oken, E. Metabolomic profiles and childhood obesity. Obesity (Silver Spring) 2014, 22, 2570–2578. [Google Scholar] [CrossRef]
- Adeva-Andany, M.; Souto-Adeva, G.; Ameneiros-Rodríguez, E.; Fernández-Fernández, C.; Donapetry-García, C.; Domínguez-Montero, A. Insulin resistance and glycine metabolism in humans. Amino Acids 2018, 50, 11–27. [Google Scholar] [CrossRef]
- Alves, A.; Bassot, A.; Bulteau, A.-L.; Pirola, L.; Morio, B. Glycine Metabolism and Its Alterations in Obesity and Metabolic Diseases. Nutrients 2019, 11, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegelasch, N.; Vogel, M.; Müller, E.; Tremel, N.; Jurkutat, A.; Löffler, M.; Terliesner, N.; Thiery, J.; Willenberg, A.; Kiess, W.; et al. Cystatin C serum levels in healthy children are related to age, gender, and pubertal stage. Pediatr. Nephrol. 2019, 34, 449–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Ding, Y.; Li, X.; Wu, X. Cystatin C is a disease-associated protein subject to multiple regulation. Immunol. Cell Biol. 2015, 93, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef] [Green Version]
- Lamichhane, A.P.; Crandell, J.L.; Jaacks, L.M.; Couch, S.C.; Lawrence, J.M.; Mayer-Davis, E.J. Longitudinal associations of nutritional factors with glycated hemoglobin in youth with type 1 diabetes: The SEARCH Nutrition Ancillary Study. Am. J. Clin. Nutr. 2015, 101, 1278–1285. [Google Scholar] [CrossRef] [Green Version]
- Mihalik, S.J.; Michaliszyn, S.F.; de las Heras, J.; Bacha, F.; Lee, S.; Chace, D.H.; DeJesus, V.R.; Vockley, J.; Arslanian, S.A. Metabolomic Profiling of Fatty Acid and Amino Acid Metabolism in Youth With Obesity and Type 2 Diabetes. Diabetes Care 2012, 35, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Wiklund, P.; Zhang, X.; Tan, X.; Keinänen-Kiukaanniemi, S.; Alen, M.; Cheng, S. Serum Amino Acid Profiles in Childhood Predict Triglyceride Level in Adulthood: A 7-Year Longitudinal Study in Girls. J. Clin. Endocrinol. Metab. 2016, 101, 2047–2055. [Google Scholar] [CrossRef] [Green Version]
- Brauer, R.; Leichtle, A.B.; Fiedler, G.M.; Thiery, J.; Ceglarek, U. Preanalytical standardization of amino acid and acylcarnitine metabolite profiling in human blood using tandem mass spectrometry. Metabolomics 2011, 7, 344–352. [Google Scholar] [CrossRef]
- World Medical Association Declaration of Helsinki: Ethical Principles for Medical Research Involving Human Subjects. JAMA 2013, 310, 2191–2194. [CrossRef] [Green Version]
- Quante, M.; Hesse, M.; Döhnert, M.; Fuchs, M.; Hirsch, C.; Sergeyev, E.; Casprzig, N.; Geserick, M.; Naumann, S.; Koch, C.; et al. The LIFE child study: A life course approach to disease and health. BMC Public Health 2012, 12, 1021. [Google Scholar] [CrossRef] [Green Version]
- Ceglarek, U.; Müller, P.; Stach, B.; Bührdel, P.; Thiery, J.; Kiess, W. Validation of the Phenylalanine/Tyrosine Ratio Determined by Tandem Mass Spectrometry: Sensitive Newborn Screening for Phenylketonuria. Clin. Chem. Lab. Med. 2002, 40, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Beuchel, C.; Becker, S.; Dittrich, J.; Kirsten, H.; Toenjes, A.; Stumvoll, M.; Loeffler, M.; Thiele, H.; Beutner, F.; Thiery, J.; et al. Clinical and lifestyle related factors influencing whole blood metabolite levels – A comparative analysis of three large cohorts. Mol. Metab. 2019, 29, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, R.; Kirsten, H.; Beutner, F.; Holdt, L.M.; Gross, A.; Teren, A.; Tönjes, A.; Becker, S.; Krohn, K.; Kovacs, P.; et al. Integration of Genome-Wide SNP Data and Gene-Expression Profiles Reveals Six Novel Loci and Regulatory Mechanisms for Amino Acids and Acylcarnitines in Whole Blood. PLoS Genet. 2015, 11, e1005510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigby, R.A.; Stasinopoulos, D.M. Generalized additive models for location, scale and shape. Appl. Statist. 2005, 54, 507–554. [Google Scholar] [CrossRef] [Green Version]
- Cole, T.J. The LMS method for constructing normalized growth standards. Eur. J. Clin. Nutr. 1990, 44, 45–60. [Google Scholar]
- Rigby, R.A.; Stasinopoulos, D.M. Using the Box-Cox t distribution in GAMLSS to model skewness and kurtosis. Stat. Model. 2006, 6, 209–229. [Google Scholar] [CrossRef]
- Vogel, M.; Kirsten, T.; Kratzsch, J.; Engel, C.; Kiess, W. A combined approach to generate laboratory reference intervals using unbalanced longitudinal data. J. Pediatr. Endocrinol. Metab. 2017, 30, 767–773. [Google Scholar] [CrossRef]
- van Buuren, S.; Fredriks, M. Worm plot: A simple diagnostic device for modelling growth reference curves. Stat. Med. 2001, 20, 1259–1277. [Google Scholar] [CrossRef]
- Kromeyer-Hauschild, K.; Wabitsch, M.; Kunze, D.; Geller, F.; Geiß, H.C.; Hesse, V.; von Hippel, A.; Jaeger, U.; Johnsen, D.; Korte, W.; et al. Perzentile für den Body-mass-Index für das Kindes- und Jugendalter unter Heranziehung verschiedener deutscher Stichproben. Monatsschr. Kinderheilkd. 2001, 149, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Vogel, M. Childsds: Data and Methods Around Reference Values in Pediatrics. R package version 0.7.2. 2019. [Google Scholar]
- Dathan-Stumpf, A.; Vogel, M.; Hiemisch, A.; Thiery, J.; Burkhardt, R.; Kratzsch, J.; Kiess, W. Pediatric reference data of serum lipids and prevalence of dyslipidemia: Results from a population-based cohort in Germany. Clin. Biochem. 2016, 49, 740–749. [Google Scholar] [CrossRef]
- Bussler, S.; Vogel, M.; Pietzner, D.; Harms, K.; Buzek, T.; Penke, M.; Händel, N.; Körner, A.; Baumann, U.; Kiess, W.; et al. New Pediatric Percentiles of Liver Enzyme Serum Levels (Alanine Aminotransferase, Aspartate Aminotransferase, c- Glutamyltransferase): Effects of Age, Sex, Body Mass Index, and Pubertal Stage. Hepatology 2018, 68, 1319–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 2 November 2019).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Laboratory Parameters | Mean (±SD) in Females | Mean (±SD) in Males |
|---|---|---|
| Glucose (mmol/L) | 4.69 (±0.43) | 4.8 (±0.43) |
| HbA1c (%) | 4.97 (±0.3) | 5.01 (±0.31) |
| Triglycerides (mmol/L) | 0.95 (±0.64) | 0.86 (±0.63) |
| Total cholesterol (mmol/L) | 4.29 (±0.75) | 4.13 (±0.73) |
| LDL-C (mmol/L) | 2.47 (±0.65) | 2.32 (±0.62) |
| HDL-C (mmol/L) | 1.54 (±0.37) | 1.57 (±0.42) |
| Cystatin C (mg/L) | 0.86 (±0.12) | 0.90 (±0.13) |
| Creatinine (µmol/L) | 49.88 (±13.86) | 50.98 (±16.12) |
| ALT (µkat/L) | 0.3 (±0.16) | 0.33 (±0.12) |
| AST(µkat/L) | 0.52 (±0.17) | 0.56 (±0.14) |
| TSH (mU/L) | 2.62 (±1.22) | 2.74 (±1.27) |
| FT3 (pmol/L) | 6.43 (±0.87) | 6.68 (±0.78) |
| FT4 (pmol/L) | 16.0 (±2.01) | 15.91 (±1.97) |
| BMI-SDS | ||||
|---|---|---|---|---|
| Underweight | Normal Weight | Overweight | Obese | |
| Females, n (%) | 166 (8.6) | 1557 (80.7) | 136 (7.1) | 70 (3.6) |
| Males, n (%) | 187 (9.3) | 1659 (82.4) | 112 (5.6) | 55 (2.7) |
| Puberty Status (Tanner Stage) | |||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Females, n (%) | 985 (56.3) | 193 (11.0) | 152 (8.7) | 175 (10.0) | 245 (14.0) |
| Females mean age (±SD) in years | 4.9 (3.3) | 10.8 (1.2) | 12.5 (1.3) | 14.2 (1.6) | 15.7 (1.5) |
| Males, n (%) | 1058 (73.1) | 194 (13.4) | 55 (3.8) | 79 (5.5) | 61 (4.2) |
| Males mean age (±SD) in years | 5.3 (3.7) | 11.4 (1.2) | 13.1 (1.0) | 14.2 (1.3) | 15.7 (1.4) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirschel, J.; Vogel, M.; Baber, R.; Garten, A.; Beuchel, C.; Dietz, Y.; Dittrich, J.; Körner, A.; Kiess, W.; Ceglarek, U. Relation of Whole Blood Amino Acid and Acylcarnitine Metabolome to Age, Sex, BMI, Puberty, and Metabolic Markers in Children and Adolescents. Metabolites 2020, 10, 149. https://doi.org/10.3390/metabo10040149
Hirschel J, Vogel M, Baber R, Garten A, Beuchel C, Dietz Y, Dittrich J, Körner A, Kiess W, Ceglarek U. Relation of Whole Blood Amino Acid and Acylcarnitine Metabolome to Age, Sex, BMI, Puberty, and Metabolic Markers in Children and Adolescents. Metabolites. 2020; 10(4):149. https://doi.org/10.3390/metabo10040149
Chicago/Turabian StyleHirschel, Josephin, Mandy Vogel, Ronny Baber, Antje Garten, Carl Beuchel, Yvonne Dietz, Julia Dittrich, Antje Körner, Wieland Kiess, and Uta Ceglarek. 2020. "Relation of Whole Blood Amino Acid and Acylcarnitine Metabolome to Age, Sex, BMI, Puberty, and Metabolic Markers in Children and Adolescents" Metabolites 10, no. 4: 149. https://doi.org/10.3390/metabo10040149
APA StyleHirschel, J., Vogel, M., Baber, R., Garten, A., Beuchel, C., Dietz, Y., Dittrich, J., Körner, A., Kiess, W., & Ceglarek, U. (2020). Relation of Whole Blood Amino Acid and Acylcarnitine Metabolome to Age, Sex, BMI, Puberty, and Metabolic Markers in Children and Adolescents. Metabolites, 10(4), 149. https://doi.org/10.3390/metabo10040149