Plasma 25-Hydroxyvitamin D Concentrations are Associated with Polyunsaturated Fatty Acid Metabolites in Young Children: Results from the Vitamin D Antenatal Asthma Reduction Trial

 , , , ,
, , , ,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Results from Age 1 Analysis
2.2. Results from Age 3 Analysis
2.3. Overlap between Age 1 and Age 3 Results
2.4. Sensitivity Analysis Results
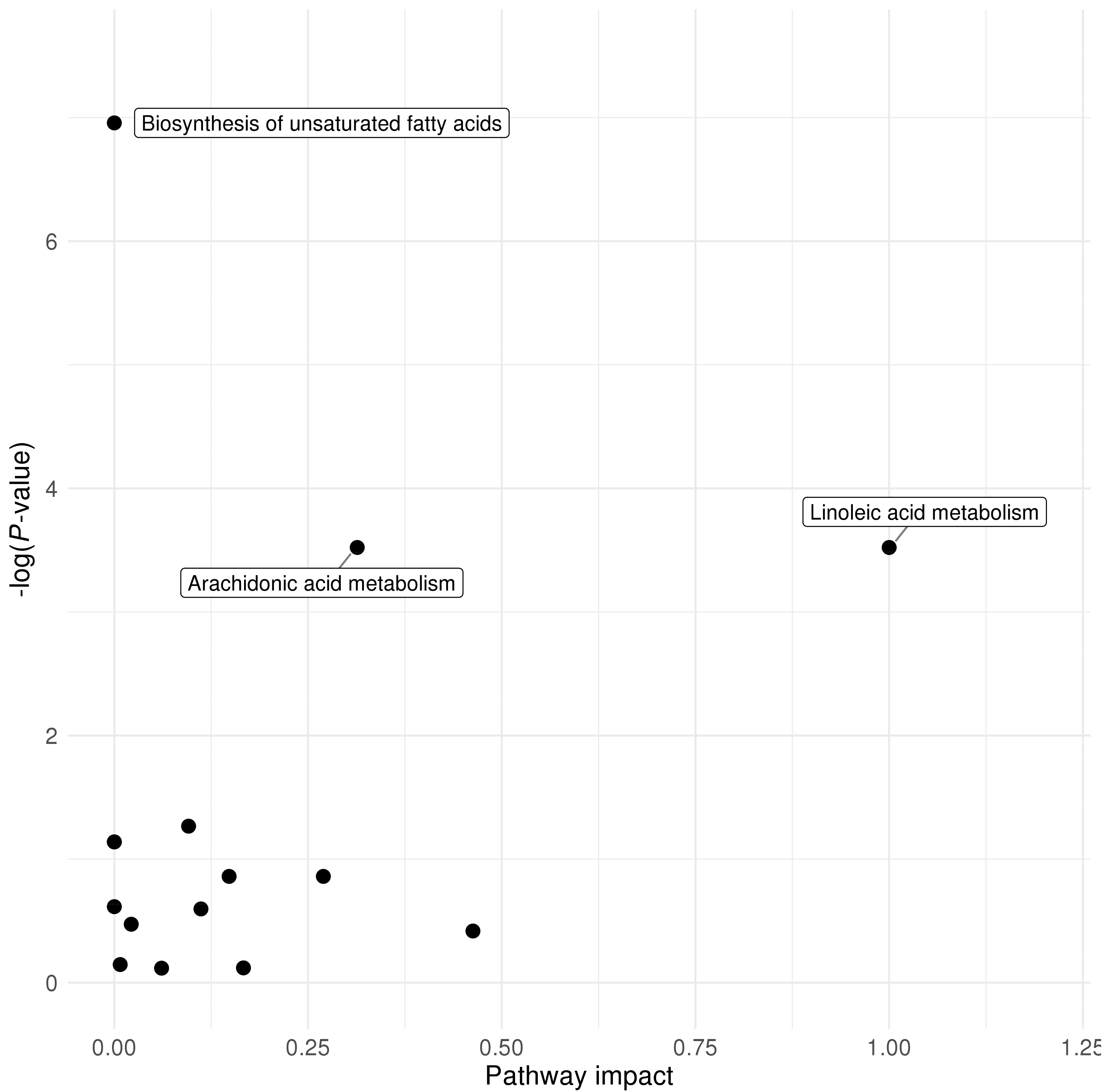
2.5. Pathway Analysis Results
2.6. Replication Analysis Results
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Vitamin D Measurements
4.3. Metabolomics Data
4.4. Statistical Analysis
4.5. Replication Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiang, X.; Kiel, D.P.; Kraft, P. The genetics of vitamin D. Bone 2018, 126, 59–77. [Google Scholar] [CrossRef] [PubMed]
- Cantorna, M.T.; Zhu, Y.; Froicu, M.; Wittke, A. Vitamin D status, 1,25-dihydroxyvitamin D3, and the immune system. Am. J. Clin. Nutr. 2004, 80, 1717S–1720S. [Google Scholar] [CrossRef] [PubMed]
- Litonjua, A.A.; Weiss, S.T. Is vitamin D deficiency to blame for the asthma epidemic? J. Allergy Clin. Immunol. 2007, 120, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Sassi, F.; Tamone, C.; D’Amelio, P. Vitamin D: Nutrient, Hormone, and Immunomodulator. Nutrients 2018, 10, 1656. [Google Scholar] [CrossRef] [Green Version]
- Hollis, B.W.; Wagner, C.L. Clinical review: The role of the parent compound vitamin D with respect to metabolism and function: Why clinical dose intervals can affect clinical outcomes. J. Clin. Endocrinol. Metab. 2013, 98, 4619–4628. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Xun, P.; Pike, K.; Wills, A.K.; Chawes, B.L.; Bisgaard, H.; Cai, W.; Wan, Y.; He, K. In utero exposure to 25-hydroxyvitamin D and risk of childhood asthma, wheeze, and respiratory tract infections: A meta-analysis of birth cohort studies. J. Allergy. Clin. Immunol. 2017, 139, 1508–1517. [Google Scholar] [CrossRef] [Green Version]
- Pilz, S.; Verheyen, N.; Grubler, M.R.; Tomaschitz, A.; Marz, W. Vitamin D and cardiovascular disease prevention. Nat. Rev. Cardiol. 2016, 13, 404–417. [Google Scholar] [CrossRef]
- Marquina, C.; Mousa, A.; Scragg, R.; de Courten, B. Vitamin D and cardiometabolic disorders: A review of current evidence, genetic determinants and pathomechanisms. Obes. Rev. 2019, 20, 262–277. [Google Scholar] [CrossRef]
- Feldman, D.; Krishnan, A.V.; Swami, S.; Giovannucci, E.; Feldman, B.J. The role of vitamin D in reducing cancer risk and progression. Nat. Rev. Cancer 2014, 14, 342–357. [Google Scholar] [CrossRef]
- Wolsk, H.M.; Chawes, B.L.; Litonjua, A.A.; Hollis, B.W.; Waage, J.; Stokholm, J.; Bonnelykke, K.; Bisgaard, H.; Weiss, S.T. Prenatal vitamin D supplementation reduces risk of asthma/recurrent wheeze in early childhood: A combined analysis of two randomized controlled trials. PLoS ONE 2017, 12, e0186657. [Google Scholar] [CrossRef]
- Hibbs, A.M.; Ross, K.; Kerns, L.A.; Wagner, C.; Fuloria, M.; Groh-Wargo, S.; Zimmerman, T.; Minich, N.; Tatsuoka, C. Effect of Vitamin D Supplementation on Recurrent Wheezing in Black Infants Who Were Born Preterm: The D-Wheeze Randomized Clinical Trial. JAMA 2018, 319, 2086–2094. [Google Scholar] [CrossRef]
- Jolliffe, D.A.; Greenberg, L.; Hooper, R.L.; Griffiths, C.J.; Camargo, C.A.; Kerley, C.P.; Jensen, M.E.; Mauger, D.; Stelmach, I.; Urashima, M.; et al. Vitamin D supplementation to prevent asthma exacerbations: A systematic review and meta-analysis of individual participant data. Lancet Respir. Med. 2017, 5, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, P.E.; Hawrylowicz, C.M. Vitamin D in Asthma: Mechanisms of Action and Considerations for Clinical Trials. Chest 2018, 153, 1229–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhrer, T.; Zamboni, N. High-throughput discovery metabolomics. Curr. Opin. Biotechnol. 2015, 31, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Vogt, S.; Wahl, S.; Kettunen, J.; Breitner, S.; Kastenmuller, G.; Gieger, C.; Suhre, K.; Waldenberger, M.; Kratzsch, J.; Perola, M.; et al. Characterization of the metabolic profile associated with serum 25-hydroxyvitamin D: A cross-sectional analysis in population-based data. Int. J. Epidemiol. 2016, 45, 1469–1481. [Google Scholar] [CrossRef] [PubMed]
- Nelson, S.M.; Panagiotou, O.A.; Anic, G.M.; Mondul, A.M.; Mannisto, S.; Weinstein, S.J.; Albanes, D. Metabolomics analysis of serum 25-hydroxy-vitamin D in the Alpha-Tocopherol, Beta-Carotene Cancer Prevention (ATBC) Study. Int. J. Epidemiol. 2016, 45, 1458–1468. [Google Scholar] [CrossRef] [Green Version]
- Lasky-Su, J.; Dahlin, A.; Litonjua, A.A.; Rogers, A.J.; McGeachie, M.J.; Baron, R.M.; Gazourian, L.; Barragan-Bradford, D.; Fredenburgh, L.E.; Choi, A.M.K.; et al. Metabolome alterations in severe critical illness and vitamin D status. Crit. Care 2017, 21, 193. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, J.L.; Pressman, E.K.; Cooper, E.M.; Kent, T.R.; Bar, H.Y.; O’Brien, K.O. Vitamin D Status Affects Serum Metabolomic Profiles in Pregnant Adolescents. Reprod. Sci. 2015, 22, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, K.; Stewart, C.J.; Celedon, J.C.; Mansbach, J.M.; Tierney, C.; Camargo, C.A., Jr. Serum 25-hydroxyvitamin D, metabolome, and bronchiolitis severity among infants-A multicenter cohort study. Pediatr. Allergy Immunol. 2018, 29, 441–445. [Google Scholar] [CrossRef]
- Berti, C.; Agostoni, C.; Davanzo, R.; Hypponen, E.; Isolauri, E.; Meltzer, H.M.; Steegers-Theunissen, R.P.; Cetin, I. Early-life nutritional exposures and lifelong health: Immediate and long-lasting impacts of probiotics, vitamin D, and breastfeeding. Nutr. Rev. 2017, 75, 83–97. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Series B (Method.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Statist. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated Fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Rosner, B. Percentage Points for a Generalized ESD Many-Outlier Procedure. Technometrics 1983, 25, 165–172. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- The Childhood Asthma Management Program (CAMP): Design, rationale, and methods. Childhood Asthma Management Program Research Group. Control Clin. Trials 1999, 20, 91–120. [CrossRef]
- Childhood Asthma Management Program Research, G.; Szefler, S.; Weiss, S.; Tonascia, J.; Adkinson, N.F.; Bender, B.; Cherniack, R.; Donithan, M.; Kelly, H.W.; Reisman, J.; et al. Long-term effects of budesonide or nedocromil in children with asthma. N. Engl. J. Med. 2000, 343, 1054–1063. [Google Scholar] [CrossRef]
- Nandi, A.; Wadhwani, N.; Joshi, S.R. Vitamin D deficiency influences fatty acid metabolism. Prostaglandins Leukot. Essent. Fatty. Acids. 2019, 140, 57–63. [Google Scholar] [CrossRef]
- Blighe, K.; Chawes, B.L.; Kelly, R.S.; Mirzakhani, H.; McGeachie, M.; Litonjua, A.A.; Weiss, S.T.; Lasky-Su, J.A. Vitamin D prenatal programming of childhood metabolomics profiles at age 3 y. Am. J. Clin. Nutr. 2017, 106, 1092–1099. [Google Scholar] [CrossRef] [Green Version]
- Simopoulos, A.P. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Nandi, A.A.; Wadhwani, N.S.; Joshi, S.R. Altered metabolic homeostasis between vitamin D and long chain polyunsaturated fatty acids in preeclampsia. Med. Hypotheses. 2017, 100, 31–36. [Google Scholar] [CrossRef]
- Das, U.N. Essential fatty acids: Biochemistry, physiology and pathology. Biotechnol. J. 2006, 1, 420–439. [Google Scholar] [CrossRef]
- Harbige, L.S. Fatty acids, the immune response, and autoimmunity: A question of n-6 essentiality and the balance between n-6 and n-3. Lipids 2003, 38, 323–341. [Google Scholar] [CrossRef]
- Kriebitzsch, C.; Verlinden, L.; Eelen, G.; van Schoor, N.M.; Swart, K.; Lips, P.; Meyer, M.B.; Pike, J.W.; Boonen, S.; Carlberg, C.; et al. 1,25-dihydroxyvitamin D3 influences cellular homocysteine levels in murine preosteoblastic MC3T3-E1 cells by direct regulation of cystathionine beta-synthase. J. Bone Miner. Res. 2011, 26, 2991–3000. [Google Scholar] [CrossRef] [Green Version]
- Signorello, M.G.; Pascale, R.; Leoncini, G. Effect of homocysteine on arachidonic acid release in human platelets. Eur. J. Clin. Invest. 2002, 32, 279–284. [Google Scholar] [CrossRef]
- Li, B.; Gao, G.; Zhang, W.; Li, B.; Yang, C.; Jiang, X.; Tian, Y.; Liang, H. Metabolomics analysis reveals an effect of homocysteine on arachidonic acid and linoleic acid metabolism pathway. Mol. Med. Rep. 2018, 17, 6261–6268. [Google Scholar] [CrossRef]
- Moreno, J.; Krishnan, A.V.; Swami, S.; Nonn, L.; Peehl, D.M.; Feldman, D. Regulation of prostaglandin metabolism by calcitriol attenuates growth stimulation in prostate cancer cells. Cancer Res. 2005, 65, 7917–7925. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, S.E. Arachidonic acid metabolites: Mediators of inflammation in asthma. Pharmacotherapy 1997, 17, 3S–12S. [Google Scholar]
- Calabrese, C.; Triggiani, M.; Marone, G.; Mazzarella, G. Arachidonic acid metabolism in inflammatory cells of patients with bronchial asthma. Allergy 2000, 55 (Suppl. 61), 27–30. [Google Scholar] [CrossRef]
- Bisgaard, H.; Stokholm, J.; Chawes, B.L.; Vissing, N.H.; Bjarnadottir, E.; Schoos, A.M.; Wolsk, H.M.; Pedersen, T.M.; Vinding, R.K.; Thorsteinsdottir, S.; et al. Fish Oil-Derived Fatty Acids in Pregnancy and Wheeze and Asthma in Offspring. N. Engl. J. Med. 2016, 375, 2530–2539. [Google Scholar] [CrossRef]
- Worobey, J. Physical activity in infancy: Developmental aspects, measurement, and importance. Am. J. Clin. Nutr. 2014, 99, 729S–733S. [Google Scholar] [CrossRef] [Green Version]
- McKennan, C.; Ober, C.; Nicolae, D. Estimation and inference in metabolomics with non-random missing data and latent factors. ArXiv 2019, arXiv:1909.02644. Available online: https://arxiv.org/abs/1909.02644 (accessed on 1 April 2020).
- Litonjua, A.A.; Lange, N.E.; Carey, V.J.; Brown, S.; Laranjo, N.; Harshfield, B.J.; O’Connor, G.T.; Sandel, M.; Strunk, R.C.; Bacharier, L.B.; et al. The Vitamin D Antenatal Asthma Reduction Trial (VDAART): Rationale, design, and methods of a randomized, controlled trial of vitamin D supplementation in pregnancy for the primary prevention of asthma and allergies in children. Contemp. Clin. Trials. 2014, 38, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Litonjua, A.A.; Carey, V.J.; Laranjo, N.; Harshfield, B.J.; McElrath, T.F.; O’Connor, G.T.; Sandel, M.; Iverson, R.E.; Lee-Paritz, A.; Strunk, R.C.; et al. Effect of Prenatal Supplementation With Vitamin D on Asthma or Recurrent Wheezing in Offspring by Age 3 Years: The VDAART Randomized Clinical Trial. JAMA 2016, 315, 362–370. [Google Scholar] [CrossRef]
- Ersfeld, D.L.; Rao, D.S.; Body, J.J.; Sackrison, J.L., Jr.; Miller, A.B.; Parikh, N.; Eskridge, T.L.; Polinske, A.; Olson, G.T.; MacFarlane, G.D. Analytical and clinical validation of the 25 OH vitamin D assay for the LIAISON automated analyzer. Clin. Biochem. 2004, 37, 867–874. [Google Scholar] [CrossRef]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef]
- Kelly, R.S.; McGeachie, M.J.; Lee-Sarwar, K.A.; Kachroo, P.; Chu, S.H.; Virkud, Y.V.; Huang, M.; Litonjua, A.A.; Weiss, S.T.; Lasky-Su, J. Partial Least Squares Discriminant Analysis and Bayesian Networks for Metabolomic Prediction of Childhood Asthma. Metabolites 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.; Marsal, S.; Julia, A. Analytical methods in untargeted metabolomics: State of the art in 2015. Front Bioeng. Biotechnol. 2015, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Demirkan, A.; van Duijn, C.M.; Ugocsai, P.; Isaacs, A.; Pramstaller, P.P.; Liebisch, G.; Wilson, J.F.; Johansson, A.; Rudan, I.; Aulchenko, Y.S.; et al. Genome-wide association study identifies novel loci associated with circulating phospho- and sphingolipid concentrations. PLoS Genet 2012, 8, e1002490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brehm, J.M.; Schuemann, B.; Fuhlbrigge, A.L.; Hollis, B.W.; Strunk, R.C.; Zeiger, R.S.; Weiss, S.T.; Litonjua, A.A.; Childhood Asthma Management Program Research, G. Serum vitamin D levels and severe asthma exacerbations in the Childhood Asthma Management Program study. J. Allergy Clin. Immunol. 2010, 126, 52–58.e55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, R.S.; Chawes, B.L.; Blighe, K.; Virkud, Y.V.; Croteau-Chonka, D.C.; McGeachie, M.J.; Clish, C.B.; Bullock, K.; Celedon, J.C.; Weiss, S.T.; et al. An Integrative Transcriptomic and Metabolomic Study of Lung Function in Children With Asthma. Chest 2018, 154, 335–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paynter, N.P.; Balasubramanian, R.; Giulianini, F.; Wang, D.D.; Tinker, L.F.; Gopal, S.; Deik, A.A.; Bullock, K.; Pierce, K.A.; Scott, J.; et al. Metabolic Predictors of Incident Coronary Heart Disease in Women. Circulation 2018, 137, 841–853. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| All Subjects (n = 450) | 25(OH)D ≤ 30 ng/mL (n = 254) | 25(OH)D > 30 ng/mL (n = 196) | P-Value 1 | |
|---|---|---|---|---|
| Age 1 25(OH)D ng/mL, mean (SD) | 29.6 (10.3) | 23.2 (5.2) | 38.0 (9.3) | <0.001 |
| Age 1 BMI kg/m2, mean (SD) | 17.4 (2.2) | 17.4 (1.9) | 17.4 (2.5) | 0.796 |
| Sex, n (%) | 0.594 | |||
| Female | 205 (45.6) | 119 (46.9) | 86 (43.9) | |
| Male | 245 (54.4) | 135 (53.1) | 110 (56.1) | |
| Race, n (%) | 0.043 | |||
| African American | 221 (49.1) | 126 (49.6) | 95 (48.5) | |
| Other | 86 (19.1) | 39 (15.4) | 47 (24.0) | |
| White | 143 (31.8) | 89 (35.0) | 54 (27.6) | |
| Ethnicity, n (%) | 0.019 | |||
| Hispanic or Latino | 160 (35.6) | 78 (30.7) | 82 (41.8) | |
| Not Hispanic or Latino | 290 (64.4) | 176 (69.3) | 114 (58.2) | |
| Study site, n (%) | 0.003 | |||
| Boston | 140 (31.1) | 71 (28.0) | 69 (35.2) | |
| San Diego | 149 (33.1) | 75 (29.5) | 74 (37.8) | |
| St. Louis | 161 (35.8) | 108 (42.5) | 53 (27.0) | |
| Season of blood collection, n (%) | 0.062 | |||
| Spring | 124 (27.6) | 59 (23.2) | 65 (33.2) | |
| Summer | 108 (24.0) | 63 (24.8) | 45 (23.0) | |
| Fall | 106 (23.6) | 69 (27.2) | 37 (18.9) | |
| Winter | 112 (24.9) | 63 (24.8) | 49 (25.0) | |
| Asthma/wheeze by age 3, n (%) | 0.669 | |||
| No | 309 (68.7) | 177 (69.7) | 132 (67.3) | |
| Yes | 141 (31.3) | 77 (30.3) | 64 (32.7) | |
| Treatment (in pregnancy), n (%) | 0.885 | |||
| 4400 IU/day vitamin D | 229 (50.9) | 126 (49.6) | 95 (48.5) | |
| 400 IU/day vitamin D | 221 (49.1) | 128 (50.4) | 101 (51.5) |
| All Subjects (n = 407) | 25(OH)D ≤ 30 ng/mL (n = 359) | 25(OH)D > 30 ng/mL (n = 48) | P-Value 1 | |
|---|---|---|---|---|
| Age 3 25(OH)D ng/mL, mean (SD) | 20.8 (8.4) | 18.7 (6.3) | 36.3 (6.1) | <0.001 |
| Age 3 BMI kg/m2, mean (SD) | 16.7 (1.9) | 16.7 (1.9) | 16.5 (1.5) | 0.390 |
| Sex, n (%) | 0.243 | |||
| Female | 189 (46.4) | 171 (47.6) | 18 (37.5) | |
| Male | 218 (53.6) | 188 (52.4) | 30 (62.5) | |
| Race, n (%) | 0.002 | |||
| African American | 197 (48.4) | 185 (51.5) | 12 (25.0) | |
| Other | 77 (18.9) | 62 (17.3) | 15 (31.2) | |
| White | 133 (32.7) | 112 (31.2) | 21 (43.8) | |
| Ethnicity, n (%) | 0.097 | |||
| Hispanic or Latino | 131 (32.2) | 110 (30.6) | 21 (43.8) | |
| Not Hispanic or Latino | 276 (67.8) | 249 (69.4) | 27 (56.2) | |
| Study site, n (%) | 0.020 | |||
| Boston | 86 (21.1) | 80 (22.3) | 6 (12.5) | |
| San Diego | 140 (34.4) | 115 (32.0) | 25 (52.1) | |
| St. Louis | 181 (44.5) | 164 (45.7) | 17 (35.4) | |
| Season of blood collection, n (%) | 0.687 | |||
| Spring | 86 (21.1) | 75 (20.9) | 11 (22.9) | |
| Summer | 114 (28.0) | 98 (27.3) | 16 (33.3) | |
| Fall | 140 (34.4) | 127 (35.4) | 13 (27.1) | |
| Winter | 67 (16.5) | 59 (16.4) | 8 (16.7) | |
| Asthma/wheeze by age 3, n (%) | 1.000 | |||
| No | 301 (74.0) | 266 (74.1) | 35 (72.9) | |
| Yes | 106 (26.0) | 93 (25.9) | 13 (27.1) | |
| Treatment (in pregnancy), n (%) | 1.000 | |||
| 4400 IU/day vitamin D | 208 (51.1) | 183 (51.0) | 25 (52.1) | |
| 400 IU/day vitamin D | 199 (48.9) | 176 (49.0) | 23 (47.9) |
| Metabolite | Pathway | Age 1 Result 2 | Age 3 Result 3 | ||||
|---|---|---|---|---|---|---|---|
| P-Value | 95% CI | P-Value | 95% CI | ||||
| docosapentaenoate (n-6 DPA; 22:5) | Long Chain PUFA | −2.27 | 9.74 × 10−6 | (−3.27, −1.27) | −1.40 | 6.47 × 10−4 | (−2.20, −0.60) |
| glycine | Glycine, Serine and Threonine Metabolism | −2.06 | 3.03 × 10−5 | (−3.02, −1.10) | −1.33 | 8.89 × 10−4 | (−2.11, −0.55) |
| 1-palmitoyl-GPE (16:0) | Lysophospholipid | −2.00 | 4.22 × 10−5 | (−2.95, −1.05) | −1.42 | 4.29 × 10−4 | (−2.20, −0.63) |
| serine | Glycine, Serine and Threonine Metabolism | −1.96 | 7.21 × 10−5 | (−2.92, −1.00) | −1.32 | 9.43 × 10−4 | (−2.11, −0.54) |
| N-acetyltaurine | Methionine, Cysteine, SAM and Taurine Metabolism | −1.95 | 7.75 × 10−5 | (−2.90, −0.99) | −1.36 | 6.96 × 10−4 | (−2.15, −0.58) |
| N-palmitoylglycine | Fatty Acid Metabolism (Acyl Glycine) | −1.91 | 1.23 × 10−4 | (−2.88, −0.94) | −1.57 | 9.27 × 10−5 | (−2.36, −0.79) |
| sphingomyelin (d18:2/16:0, d18:1/16:1) | Sphingomyelins | −1.80 | 3.06 × 10−4 | (−2.77, −0.83) | −1.52 | 1.66 × 10−4 | (−2.31, −0.74) |
| arachidonate (20:4 n-6) | Long Chain PUFA | −1.78 | 3.10 × 10−4 | (−2.74, −0.82) | −1.40 | 6.20 × 10−4 | (−2.19, −0.60) |
| palmitoyl-linoleoyl-glycerol (16:0/18:2) | Diacylglycerol | −1.68 | 5.83 × 10−4 | (−2.64, −.073) | −1.38 | 5.89 × 10−4 | (−2.16, −0.60) |
| linoleate (18:2 n-6) | Long Chain PUFA | −1.65 | 7.23 × 10−4 | (−2.60, −0.70) | −1.65 | 4.70 × 10−5 | (−2.44, −0.86) |
| hydroxyproline | Urea cycle; Arginine and Proline Metabolism | −1.75 | 7.38 × 10−4 | (−2.77, −0.74) | −1.67 | 5.06 × 10−5 | (−2.47, −0.87) |
| 1-stearoyl-GPE (18:0) | Lysophospholipid | −1.65 | 7.44 × 10−4 | (−2.61, −0.70) | −1.57 | 6.57 × 10−5 | (−2.34, −0.81) |
| Metabolite | Pathway | Age 1 Result 2 | Age 3 Result 3 | ||||
|---|---|---|---|---|---|---|---|
| P-Value | 95% CI | P-Value | 95% CI | ||||
| N-acetyltaurine | Methionine, Cysteine, SAM and Taurine Metabolism | −1.90 | 7.01 × 10−6 | (−2.72, −1.08) | −1.37 | 4.05 × 10−4 | (−2.13, −0.62) |
| docosapentaenoate (n-6 DPA; 22:5) | Long Chain PUFA | −1.91 | 1.29 × 10−5 | (−2.77, −1.06) | −1.40 | 4.28 × 10−4 | (−2.17, −0.62) |
| dihomo-linolenate (20:3 n-3 or n-6) | Long Chain PUFA | −1.88 | 1.60 × 10−5 | (−2.72, −1.03) | −1.33 | 8.10 × 10−4 | (−2.11, −0.56) |
| glycine | Glycine, Serine and Threonine Metabolism | −1.64 | 1.05 × 10−4 | (−2.46, −0.82) | −1.38 | 3.67 × 10−4 | (−2.13, −0.62) |
| arachidonate (20:4 n-6) | Long Chain PUFA | −1.62 | 1.13 × 10−4 | (−2.44, −0.80) | −1.34 | 6.58 × 10−4 | (−2.12, −0.57) |
| linoleate (18:2 n-6) | Long Chain PUFA | −1.56 | 1.79 × 10−4 | (−2.37, −0.75) | −1.63 | 3.14 × 10−5 | (−2.39, −0.87) |
| serine | Glycine, Serine and Threonine Metabolism | −1.59 | 1.87 × 10−4 | (−2.41, −0.76) | −1.30 | 7.74 × 10−4 | (−2.05, −0.55) |
| N-palmitoylglycine | Fatty Acid Metabolism (Acyl Glycine) | −1.50 | 4.28 × 10−4 | (−2.34, −0.67) | −1.65 | 2.21 × 10−5 | (−2.40, −0.89) |
| 1-palmitoyl-GPE (16:0) | Lysophospholipid | −1.47 | 5.11 × 10−4 | (−2.29, −0.64) | −1.53 | 8.81 × 10−5 | (−2.28, −0.77) |
| linoleoyl ethanolamide | Endocannabinoid | −1.43 | 6.57 × 10−4 | (−2.25, −0.61) | −1.29 | 9.53 × 10−4 | (−2.06, −0.53) |
| valylglycine | Dipeptide | −1.44 | 6.59 × 10−4 | (−2.26, −0.61) | −1.81 | 4.43 × 10−6 | (−2.57, −1.04) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, M.; Kelly, R.S.; Kachroo, P.; Chu, S.H.; Lee-Sarwar, K.; Chawes, B.L.; Bisgaard, H.; Litonjua, A.A.; Weiss, S.T.; Lasky-Su, J. Plasma 25-Hydroxyvitamin D Concentrations are Associated with Polyunsaturated Fatty Acid Metabolites in Young Children: Results from the Vitamin D Antenatal Asthma Reduction Trial. Metabolites 2020, 10, 151. https://doi.org/10.3390/metabo10040151
Huang M, Kelly RS, Kachroo P, Chu SH, Lee-Sarwar K, Chawes BL, Bisgaard H, Litonjua AA, Weiss ST, Lasky-Su J. Plasma 25-Hydroxyvitamin D Concentrations are Associated with Polyunsaturated Fatty Acid Metabolites in Young Children: Results from the Vitamin D Antenatal Asthma Reduction Trial. Metabolites. 2020; 10(4):151. https://doi.org/10.3390/metabo10040151
Chicago/Turabian StyleHuang, Mengna, Rachel S. Kelly, Priyadarshini Kachroo, Su H. Chu, Kathleen Lee-Sarwar, Bo L. Chawes, Hans Bisgaard, Augusto A. Litonjua, Scott T. Weiss, and Jessica Lasky-Su. 2020. "Plasma 25-Hydroxyvitamin D Concentrations are Associated with Polyunsaturated Fatty Acid Metabolites in Young Children: Results from the Vitamin D Antenatal Asthma Reduction Trial" Metabolites 10, no. 4: 151. https://doi.org/10.3390/metabo10040151
APA StyleHuang, M., Kelly, R. S., Kachroo, P., Chu, S. H., Lee-Sarwar, K., Chawes, B. L., Bisgaard, H., Litonjua, A. A., Weiss, S. T., & Lasky-Su, J. (2020). Plasma 25-Hydroxyvitamin D Concentrations are Associated with Polyunsaturated Fatty Acid Metabolites in Young Children: Results from the Vitamin D Antenatal Asthma Reduction Trial. Metabolites, 10(4), 151. https://doi.org/10.3390/metabo10040151