Alterations in Tissue Metabolite Profiles with Amifostine-Prophylaxed Mice Exposed to Gamma Radiation

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (a)
- to decrease the xerostomia in malignant patients receiving radiotherapy after surgery for head and neck cancer and
- (b)
2. Results
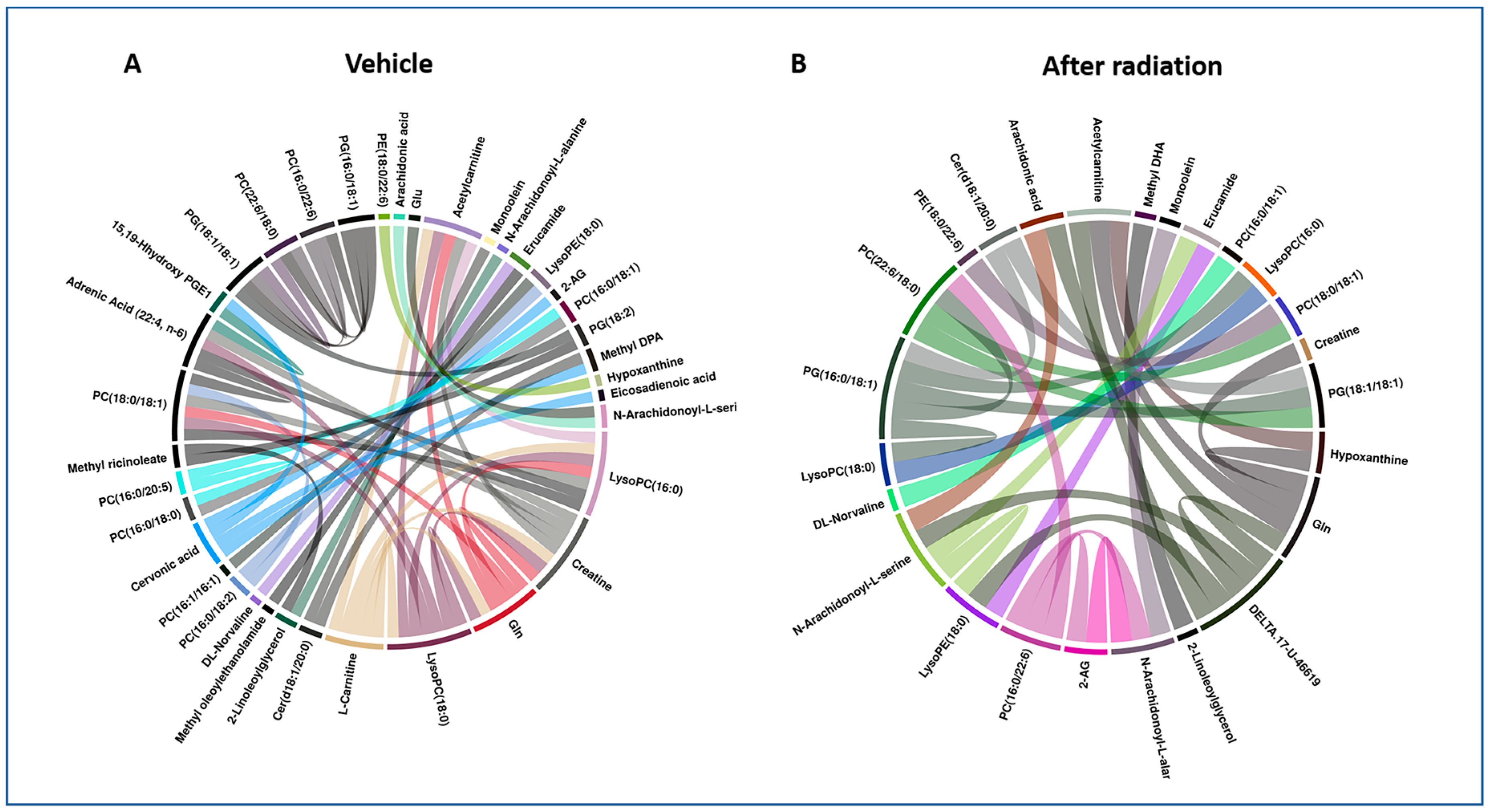
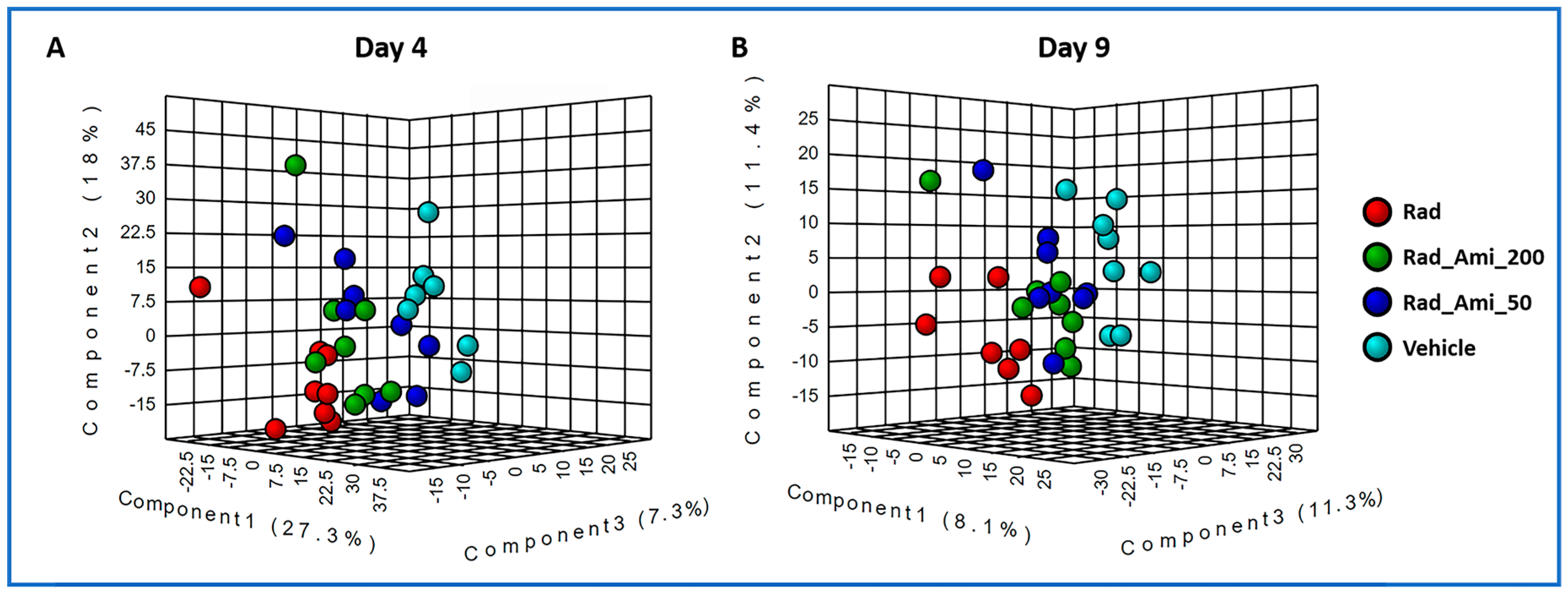
2.1. Exposure to Gamma-Radiation-Induced Robust Changes in Tissue Metabolic Profiles
2.2. Amifostine (50 and 200 mg/kg) Does Not Stimulate Major Metabolic Changes in Mice
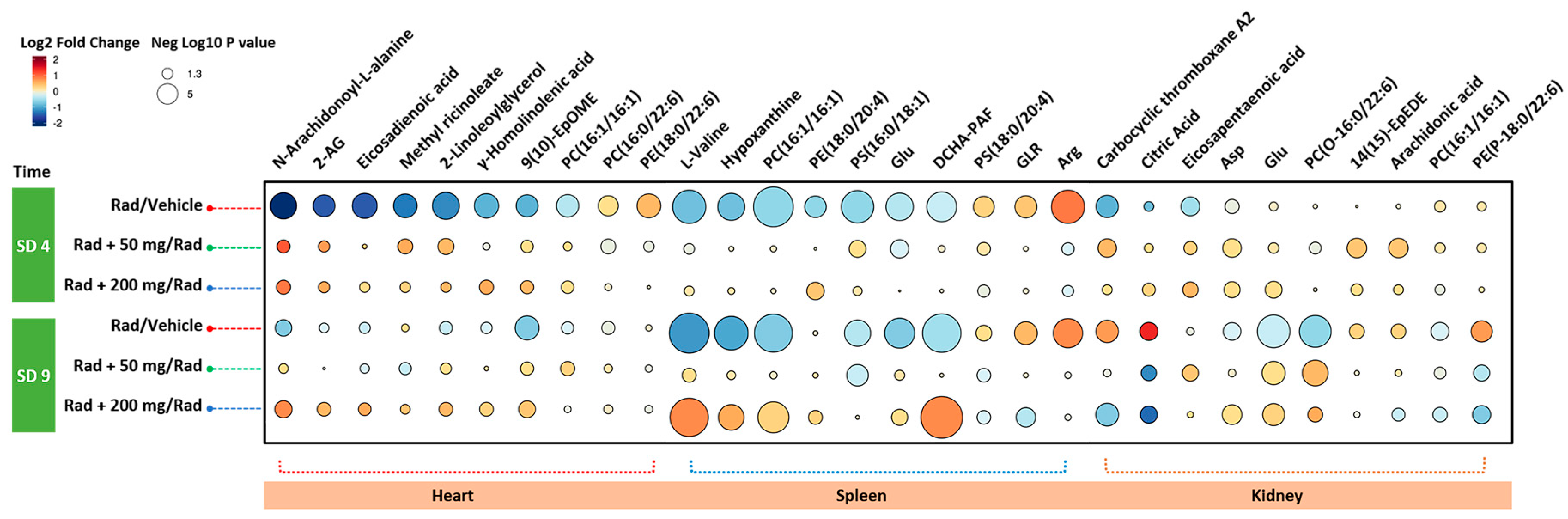
2.3. Administration of Amifostine Partially Corrects Metabolic Perturbations Caused by Ionizing Radiation in Mice Heart, Spleen, and Kidney Tissues
3. Discussion
4. Materials and Methods
4.1. Mice
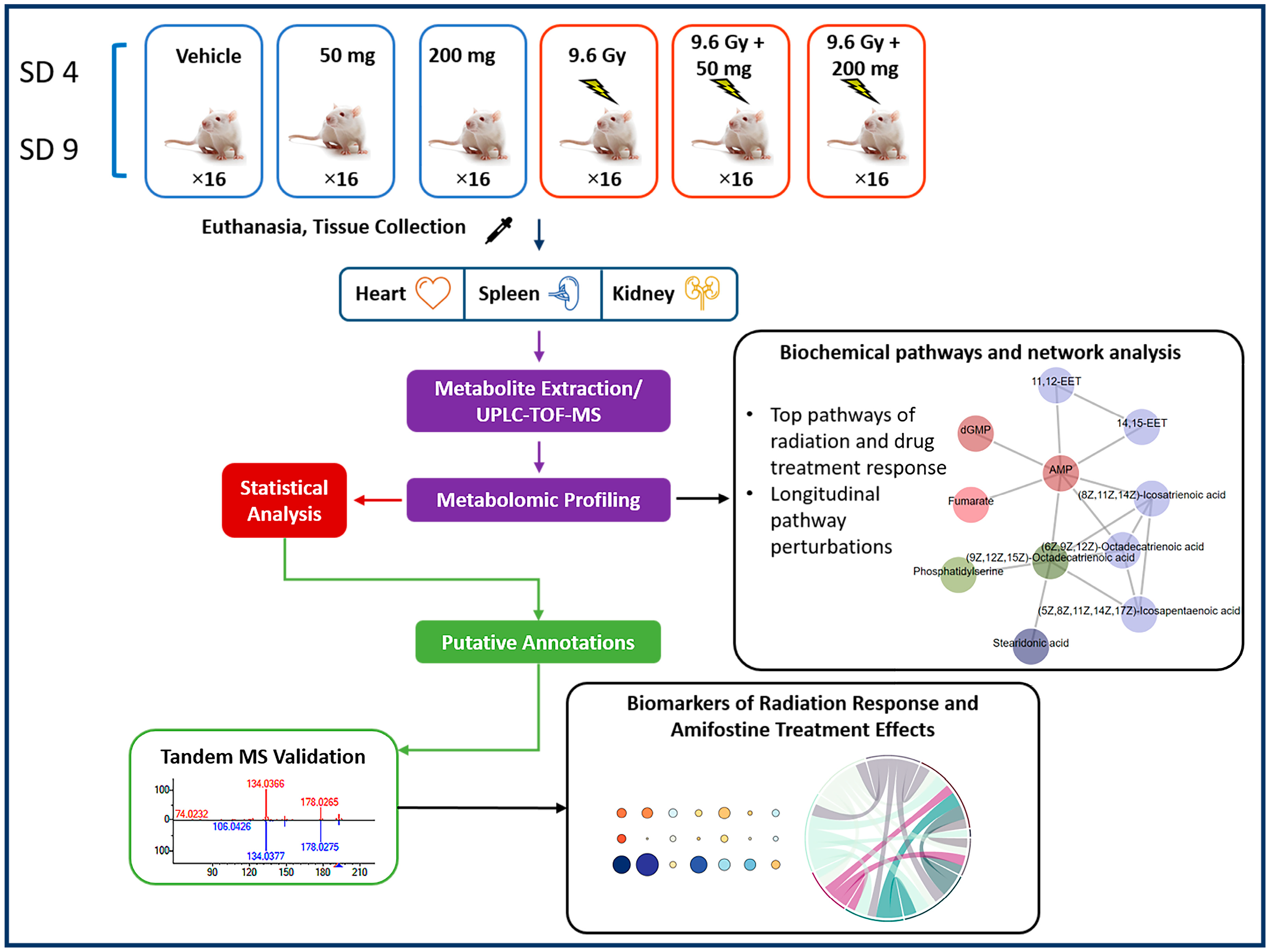
4.2. Experimental Design
4.3. Drug Administration to Mice
4.4. Radiation Exposure
4.5. Collection of Tissue Samples
4.6. Tissue Metabolomic Profile Analysis Utilizing UPLC-QTOF Mass Spectrometry
4.7. Statistical Analysis of Metabolomics Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bushberg, J.T.; Kroger, L.A.; Hartman, M.B.; Leidholdt, E.M., Jr.; Miller, K.L.; Derlet, R.; Wraa, C. Nuclear/radiological terrorism: Emergency department management of radiation casualties. J. Emerg. Med. 2007, 32, 71–85. [Google Scholar] [CrossRef] [PubMed]
- McCann, D.G.C. Radiation poisoning: Current concepts in the acute radiation syndrome. Am. J. Clin. Med. 2006, 3, 13–21. [Google Scholar]
- Singh, V.K.; Seed, T.M. The efficacy and safety of amifostine for the acute radiation syndrome. Expert Opin. Drug Saf. 2019, 18, 1077–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Seed, T.M. A review of radiation countermeasures focusing on injury-specific medicinals and regulatory approval status: Part I. Radiation sub-syndromes, animal models and FDA-approved countermeasures. Int. J. Radiat. Biol. 2017, 93, 851–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreassen, C.N.; Grau, C.; Lindegaard, J.C. Chemical radioprotection: A critical review of amifostine as a cytoprotector in radiotherapy. Semin. Radiat. Oncol. 2003, 13, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Capizzi, R.L. The preclinical basis for broad-spectrum selective cytoprotection of normal tissues from cytotoxic therapies by amifostine. Semin. Oncol. 1999, 26, 3–21. [Google Scholar] [CrossRef]
- Capizzi, R.L.; Oster, W. Chemoprotective and radioprotective effects of amifostine: An update of clinical trials. Int. J. Hematol. 2000, 72, 425–435. [Google Scholar]
- Capizzi, R.L. Clinical status and optimal use of amifostine. Oncology 1999, 13, 47–59; discussion 63, 67. [Google Scholar]
- Nicolatou-Galitis, O.; Sarri, T.; Bowen, J.; Di Palma, M.; Kouloulias, V.E.; Niscola, P.; Riesenbeck, D.; Stokman, M.; Tissing, W.; Yeoh, E.; et al. Systematic review of amifostine for the management of oral mucositis in cancer patients. Support. Care Cancer 2013, 21, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Zhu, S.; Li, X.; Wu, H.; Li, Y.; Hua, F. Effect of amifostine in head and neck cancer patients treated with radiotherapy: A systematic review and meta-analysis based on randomized controlled trials. PLoS ONE 2014, 9, e95968. [Google Scholar] [CrossRef]
- Seed, T.M.; Inal, C.E.; Singh, V.K. Radioprotection of hematopoietic progenitors by low dose amifostine prophylaxis. Int. J. Radiat. Biol. 2014, 90, 594–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grdina, D.J.; Kataoka, Y.; Murley, J.S. Amifostine: Mechanisms of action underlying cytoprotection and chemoprevention. Drug Metabol. Drug Interact. 2000, 16, 237–279. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.V.; Carrithers, S.L.; Parkinson, S.J.; Skurk, C.; Nuss, C.; Pooler, P.M.; Owen, C.S.; Lefer, A.M.; Waldman, S.A. Hypotensive mechanisms of amifostine. J. Clin. Pharmacol. 1996, 36, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Turrisi, A.T.; Glover, D.J.; Hurwitz, S.; Glick, J.; Norfleet, A.L.; Weiler, C.; Yuhas, J.M.; Kligerman, M.M. Final report of the phase I trial of single-dose WR-2721 [S-2-(3-aminopropylamino)ethylphosphorothioic acid]. Cancer Treat. Rep. 1986, 70, 1389–1393. [Google Scholar] [PubMed]
- Landauer, M.R.; Davis, H.D.; Dominitz, J.A.; Weiss, J.F. Dose and time relationships of the radioprotector WR-2721 on locomotor activity in mice. Pharmacol. Biochem. Behav. 1987, 27, 573–576. [Google Scholar] [CrossRef]
- Landauer, M.R.; Davis, H.D.; Dominitz, J.A.; Weiss, J.F. Long-term effects of radioprotector WR-2721 on locomotor activity and body weight of mice following exposure to ionizing radiation. Toxicology 1988, 49, 315–323. [Google Scholar] [CrossRef]
- Landauer, M.R.; Davis, H.D.; Dominitz, J.A.; Weiss, J.F. Comparative behavioral toxicity of four sulfhydryl radioprotective compounds in mice: WR-2721, cysteamine, diethyldithiocarbamate, and N-acetylcysteine. Pharmacol. Ther. 1988, 39, 97–100. [Google Scholar] [CrossRef]
- Dorr, R.T. Radioprotectants: Pharmacology and clinical applications of amifostine. Semin. Radiat. Oncol. 1998, 8, 10–13. [Google Scholar]
- Weiss, J.F. Pharmacologic approaches to protection against radiation-induced lethality and other damage. Environ. Health Perspect. 1997, 105 (Suppl. 6), 1473–1478. [Google Scholar]
- Seed, T.M.; Fry, S.A.; Neta, R.; Weiss, J.F.; Jarrett, D.G.; Thomassen, D. Prevention and treatments: Summary statement. Mil. Med. 2002, 167, 87–93. [Google Scholar]
- Cumberland Pharmaceuticals Inc. ETHYOL-Amifostine Injection, Powder, Lyophilized, for Solution. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2017/020221s033lbl.pdf (accessed on 12 February 2020).
- Brizel, D.M.; Wasserman, T.H.; Henke, M.; Strnad, V.; Rudat, V.; Monnier, A.; Eschwege, F.; Zhang, J.; Russell, L.; Oster, W.; et al. Phase III randomized trial of amifostine as a radioprotector in head and neck cancer. J. Clin. Oncol. 2000, 18, 3339–3345. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Fatanmi, O.O.; Wise, S.Y.; Newman, V.L.; Romaine, P.L.; Seed, T.M. The potentiation of the radioprotective efficacy of two medical countermeasures, gamma-tocotrienol and amifostine, by a combination prophylactic modality. Radiat. Prot. Dosimetry 2016, 172, 302–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seed, T.; Singh, V.K.; Hanlon, B.K. Early and late changes in radiation-induced gene expression arrays following radioprotection with amifostine. J. Radiat. Cancer Res. 2019, 10, 44–57. [Google Scholar] [CrossRef]
- Cheema, A.K.; Li, Y.; Girgis, M.; Jayatilake, M.; Simas, M.; Wise, S.Y.; Olabisi, A.O.; Seed, T.M.; Singh, V.K. Metabolomic studies in tissues of mice treated with amifostine and exposed to gamma-radiation. Sci. Rep. 2019, 9, 15701. [Google Scholar] [CrossRef] [PubMed]
- Baradaran-Ghahfarokhi, M. Radiation-induced kidney injury. J. Renal Inj. Prev. 2012, 1, 49–50. [Google Scholar] [CrossRef]
- Cheema, A.K.; Mehta, K.Y.; Fatanmi, O.O.; Wise, S.Y.; Hinzman, C.P.; Wolff, J.; Singh, V.K. A Metabolomic and lipidomic serum signature from nonhuman primates administered with a promising radiation countermeasure, gamma-tocotrienol. Int. J. Mol. Sci. 2017, 19, 79. [Google Scholar] [CrossRef] [Green Version]
- Culy, C.R.; Spencer, C.M. Amifostine: An update on its clinical status as a cytoprotectant in patients with cancer receiving chemotherapy or radiotherapy and its potential therapeutic application in myelodysplastic syndrome. Drugs 2001, 61, 641–684. [Google Scholar] [CrossRef]
- Valeyrie-Allanore, L.; Poulalhon, N.; Fagot, J.P.; Sekula, P.; Davidovici, B.; Sidoroff, A.; Mockenhaupt, M. Stevens-Johnson syndrome and toxic epidermal necrolysis induced by amifostine during head and neck radiotherapy. Radiother. Oncol. 2008, 87, 300–303. [Google Scholar] [CrossRef]
- Ali, B.H.; Al Moundhri, M.S. Agents ameliorating or augmenting the nephrotoxicity of cisplatin and other platinum compounds: A review of some recent research. Food Chem. Toxicol. 2006, 44, 1173–1183. [Google Scholar] [CrossRef]
- Rades, D.; Fehlauer, F.; Bajrovic, A.; Mahlmann, B.; Richter, E.; Alberti, W. Serious adverse effects of amifostine during radiotherapy in head and neck cancer patients. Radiother. Oncol. 2004, 70, 261–264. [Google Scholar] [CrossRef]
- Phillips, G.L., 2nd; Bernstein, S.H.; Liesveld, J.L.; Abboud, C.N.; Becker, M.W.; Constine, L.S.; Ifthikharuddin, J.J.; Loughner, J.E.; Milner, L.A.; Vesole, D.H.; et al. A Phase I trial: Dose escalation of melphalan in the “BEAM” regimen using amifostine cytoprotection. Biol. Blood Marrow Transplant. 2011, 17, 1033–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duparc, A.; Delaporte, E.; Coche, B.; Piette, F.; Mortier, L. [Amifostine-induced Stevens-Johnson syndrome during radiotherapy]. Ann. Dermatol. Venereol. 2008, 135, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Pandit, J.J.; Allen, C.; Little, E.; Formenti, F.; Harris, A.L.; Robbins, P.A. Does amifostine reduce metabolic rate? Effect of the drug on gas exchange and acute ventilatory hypoxic response in humans. Pharmaceuticals 2015, 8, 186–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koukourakis, M.I.; Giatromanolaki, A.; Zois, C.E.; Kalamida, D.; Pouliliou, S.; Karagounis, I.V.; Yeh, T.L.; Abboud, M.I.; Claridge, T.D.; Schofield, C.J.; et al. Normal tissue radioprotection by amifostine via Warburg-type effects. Sci. Rep. 2016, 6, 30986. [Google Scholar] [CrossRef] [Green Version]
- Saada, H.N.; Said, U.Z.; Mahdy, E.M.; Elmezayen, H.E.; Shedid, S.M. Fish oil omega-3 fatty acids reduce the severity of radiation-induced oxidative stress in the rat brain. Int. J. Radiat. Biol. 2014, 90, 1179–1183. [Google Scholar] [CrossRef]
- Pearce, L.L.; Zheng, X.; Martinez-Bosch, S.; Kerr, P.P.; Khlangwiset, P.; Epperly, M.W.; Fink, M.P.; Greenberger, J.S.; Peterson, J. l-arginine is a radioprotector for hematopoietic progenitor cells. Radiat. Res. 2012, 177, 792–803. [Google Scholar] [CrossRef] [Green Version]
- Shukla, J.; Chatterjee, S.; Thakur, V.S.; Premachandran, S.; Checker, R.; Poduval, T.B. l-Arginine reverses radiation-induced immune dysfunction: The need for optimum treatment window. Radiat. Res. 2009, 171, 180–187. [Google Scholar] [CrossRef]
- Yu, A.C.; Chan, P.H.; Fishman, R.A. Effects of arachidonic acid on glutamate and gamma-aminobutyric acid uptake in primary cultures of rat cerebral cortical astrocytes and neurons. J. Neurochem. 1986, 47, 1181–1189. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ebina, S.; Nakanishi, H.; Nakahata, N. Thromboxane A2 receptor-mediated signal transduction in rabbit aortic smooth muscle cells. Gen. Pharmacol. 1995, 26, 1489–1498. [Google Scholar] [CrossRef]
- Ding, X.; Murray, P.A. Cellular mechanisms of thromboxane A2-mediated contraction in pulmonary veins. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 289, L825–L833. [Google Scholar] [CrossRef]
- National Research Council of the National Academy of Sciences. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Nagy, V. Accuracy considerations in EPR dosimetry. Appl. Radiat. Isot. 2000, 52, 1039–1050. [Google Scholar] [CrossRef]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Prince, J.T.; Marcotte, E.M. Chromatographic alignment of ESI-LC-MS proteomics data sets by ordered bijective interpolated warping. Anal. Chem. 2006, 78, 6140–6152. [Google Scholar] [CrossRef] [PubMed]
- Libiseller, G.; Dvorzak, M.; Kleb, U.; Gander, E.; Eisenberg, T.; Madeo, F.; Neumann, S.; Trausinger, G.; Sinner, F.; Pieber, T.; et al. IPO: A tool for automated optimization of XCMS parameters. BMC Bioinform. 2015, 16, 118. [Google Scholar] [CrossRef] [Green Version]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef]
- Gagnebin, Y.; Tonoli, D.; Lescuyer, P.; Ponte, B.; de Seigneux, S.; Martin, P.Y.; Schappler, J.; Boccard, J.; Rudaz, S. Metabolomic analysis of urine samples by UHPLC-QTOF-MS: Impact of normalization strategies. Ana.l Chim. Acta 2017, 955, 27–35. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vazquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Sud, M.; Fahy, E.; Cotter, D.; Brown, A.; Dennis, E.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; et al. LMSD: LIPID MAPS structure database. Nucleic Acids Res. 2007, 35, D527–D532. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Park, Y.; Duraisingham, S.; Strobel, F.H.; Khan, N.; Soltow, Q.A.; Jones, D.P.; Pulendran, B. Predicting network activity from high throughput metabolomics. PLoS Comput. Biol. 2013, 9, e1003123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardinassi, L.G.; Arevalo-Herrera, M.; Herrera, S.; Cordy, R.J.; Tran, V.; Smith, M.R.; Johnson, M.S.; Chacko, B.; Liu, K.H.; Darley-Usmar, V.M.; et al. Integrative metabolomics and transcriptomics signatures of clinical tolerance to Plasmodium vivax reveal activation of innate cell immunity and T cell signaling. Redox Biol. 2018, 17, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, C.; Bommakanti, G.; Gardinassi, L.; Loebbermann, J.; Johnson, M.J.; Hakimpour, P.; Hagan, T.; Benitez, L.; Todor, A.; Machiah, D.; et al. mTOR regulates metabolic adaptation of APCs in the lung and controls the outcome of allergic inflammation. Science 2017, 357, 1014–1021. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Sullivan, N.L.; Rouphael, N.; Yu, T.; Banton, S.; Maddur, M.S.; McCausland, M.; Chiu, C.; Canniff, J.; Dubey, S.; et al. Metabolic phenotypes of response to vaccination in humans. Cell 2017, 169, 862–877. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheema, A.K.; Li, Y.; Girgis, M.; Jayatilake, M.; Fatanmi, O.O.; Wise, S.Y.; Seed, T.M.; Singh, V.K. Alterations in Tissue Metabolite Profiles with Amifostine-Prophylaxed Mice Exposed to Gamma Radiation. Metabolites 2020, 10, 211. https://doi.org/10.3390/metabo10050211
Cheema AK, Li Y, Girgis M, Jayatilake M, Fatanmi OO, Wise SY, Seed TM, Singh VK. Alterations in Tissue Metabolite Profiles with Amifostine-Prophylaxed Mice Exposed to Gamma Radiation. Metabolites. 2020; 10(5):211. https://doi.org/10.3390/metabo10050211
Chicago/Turabian StyleCheema, Amrita K., Yaoxiang Li, Michael Girgis, Meth Jayatilake, Oluseyi O. Fatanmi, Stephen Y. Wise, Thomas M. Seed, and Vijay K. Singh. 2020. "Alterations in Tissue Metabolite Profiles with Amifostine-Prophylaxed Mice Exposed to Gamma Radiation" Metabolites 10, no. 5: 211. https://doi.org/10.3390/metabo10050211
APA StyleCheema, A. K., Li, Y., Girgis, M., Jayatilake, M., Fatanmi, O. O., Wise, S. Y., Seed, T. M., & Singh, V. K. (2020). Alterations in Tissue Metabolite Profiles with Amifostine-Prophylaxed Mice Exposed to Gamma Radiation. Metabolites, 10(5), 211. https://doi.org/10.3390/metabo10050211