Potential Metabolite Nymphayol Isolated from Water Lily (Nymphaea stellata) Flower Inhibits MCF-7 Human Breast Cancer Cell Growth via Upregulation of Cdkn2a, pRb2, p53 and Downregulation of PCNA mRNA Expressions

 ,
,  ,
,
Abstract
:1. Introduction
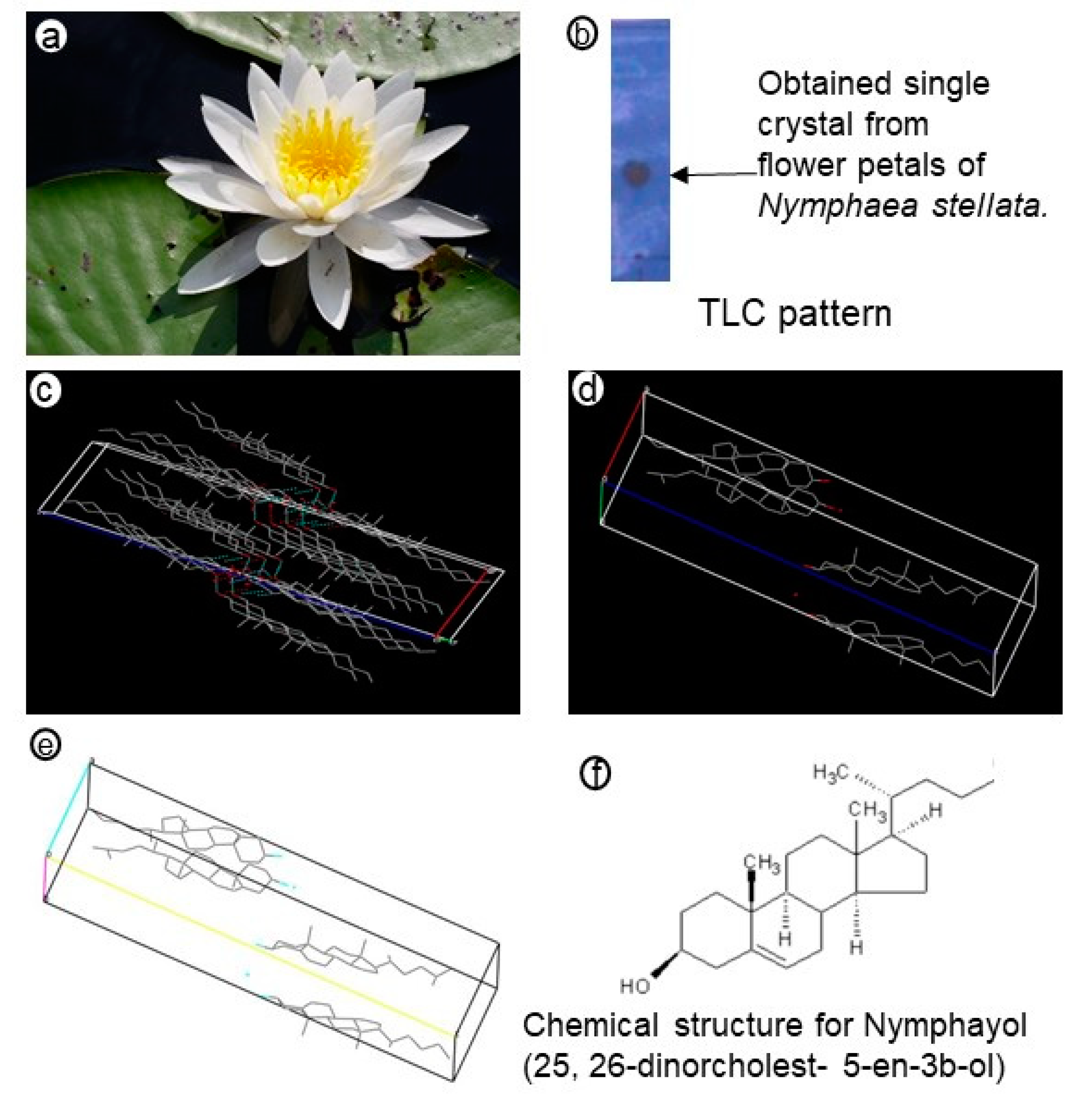
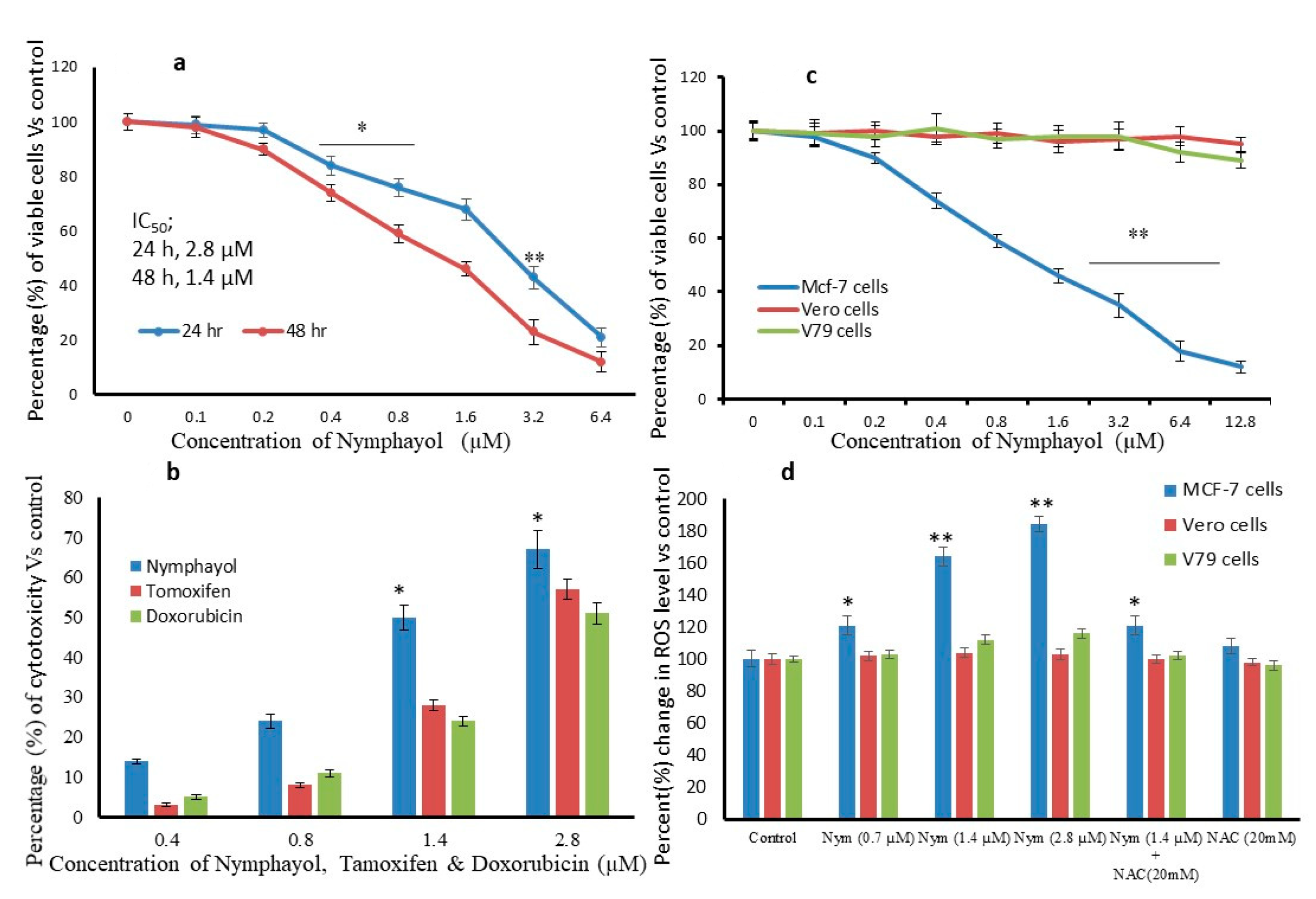
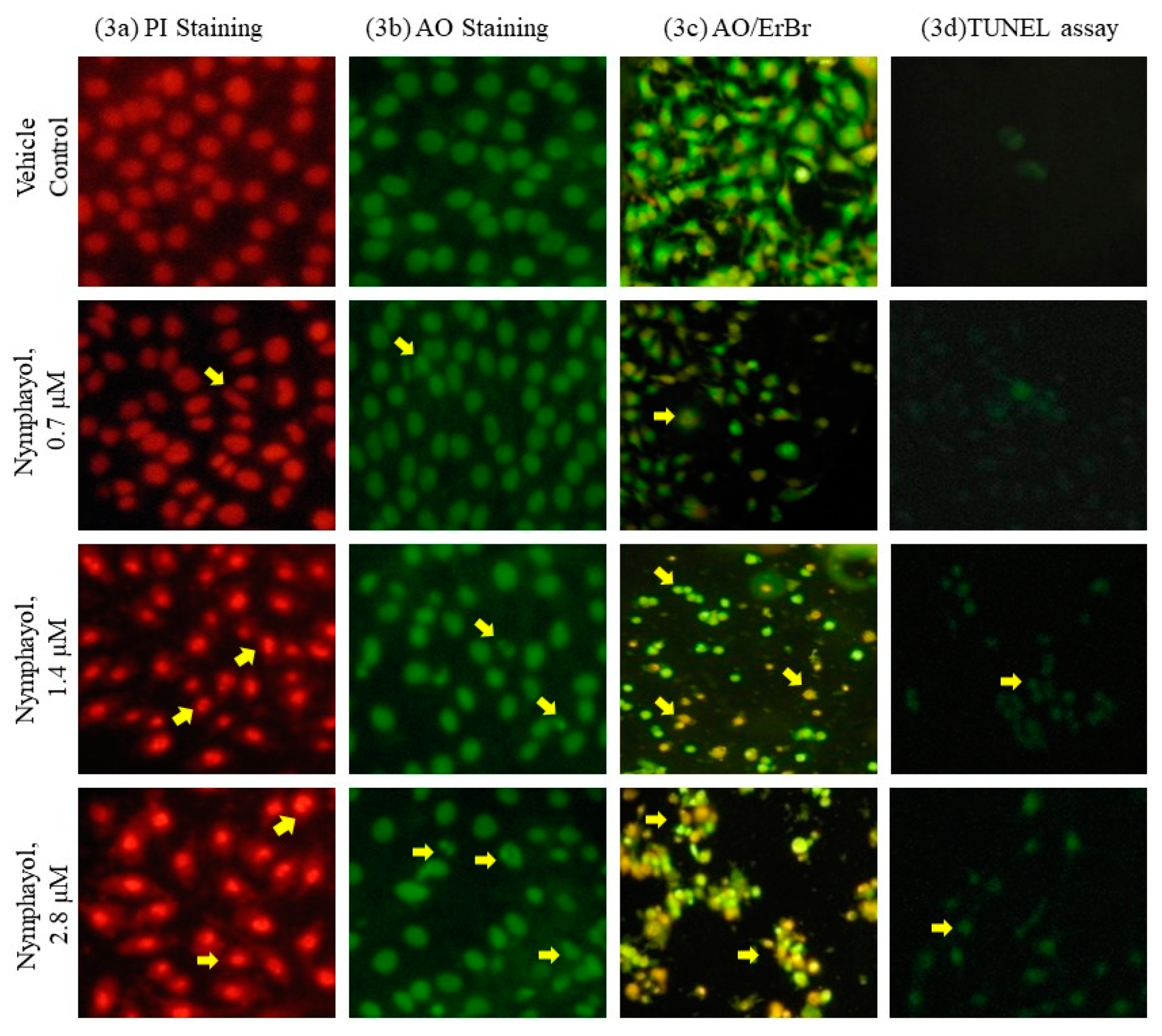
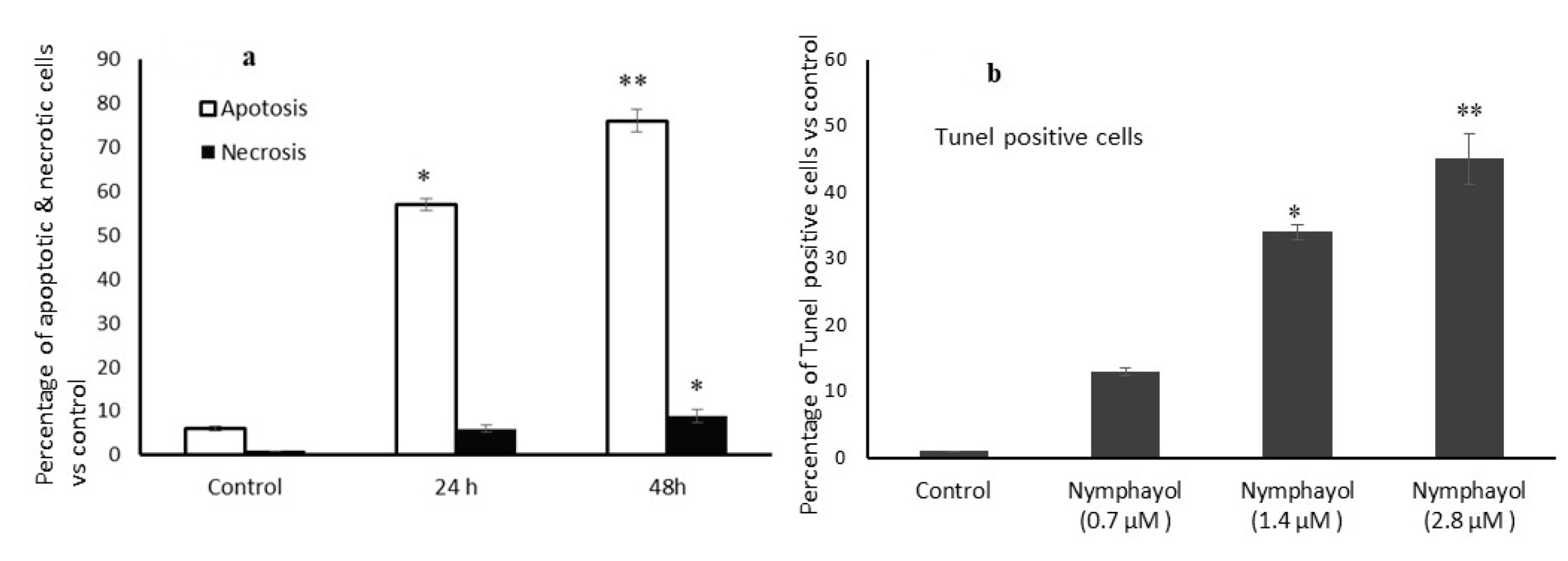
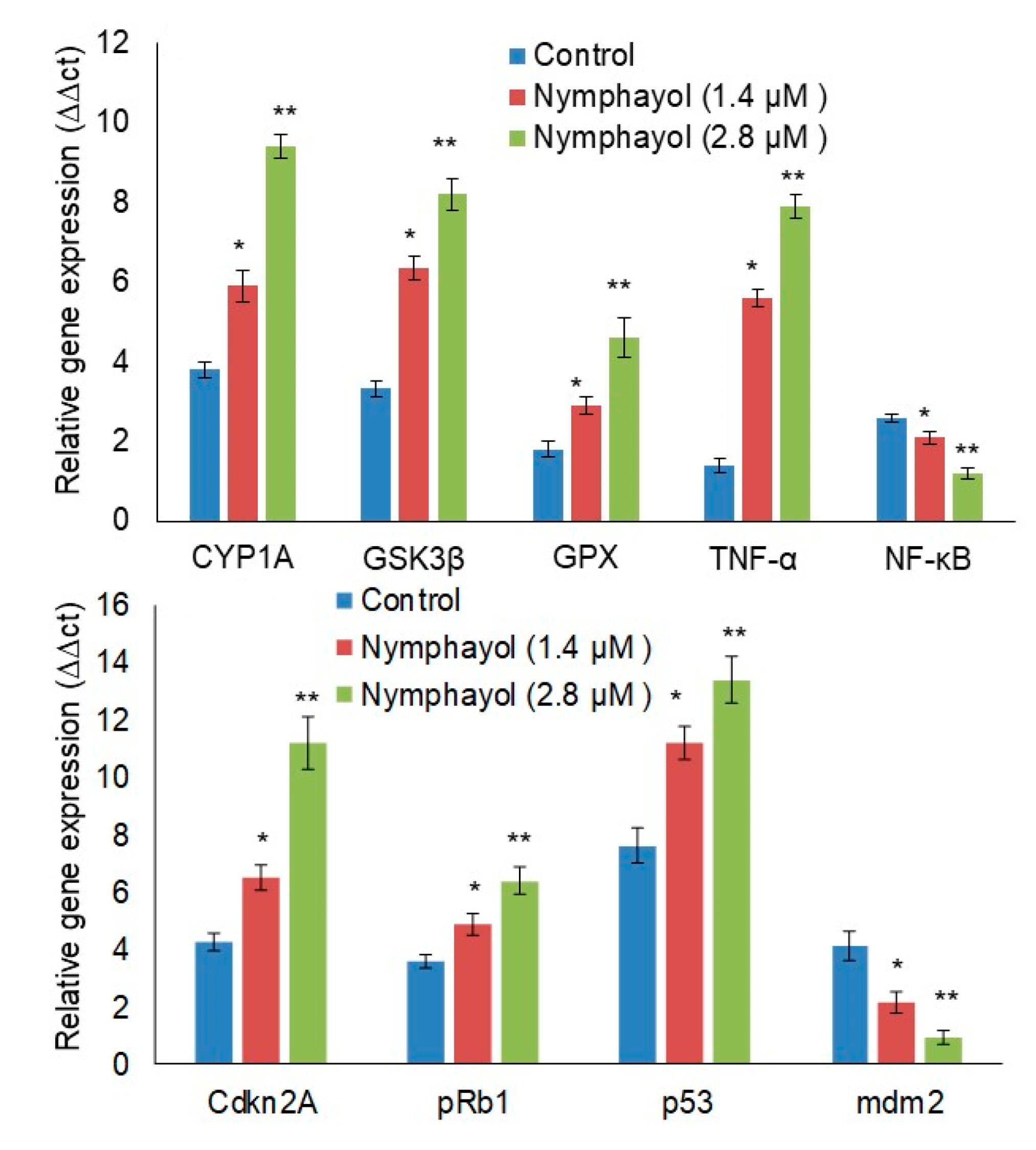
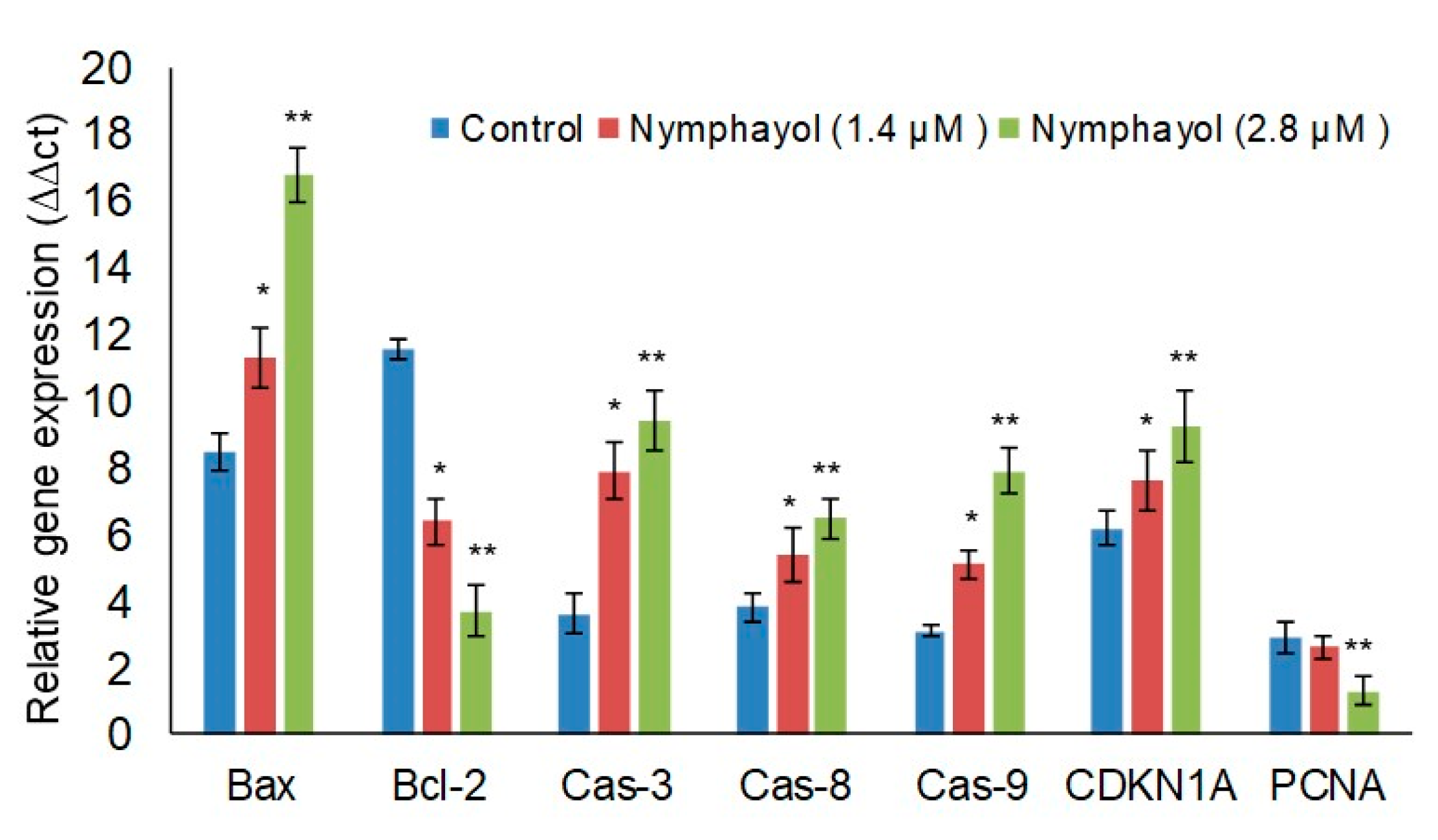
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Culture Materials
4.2. N. Stellata Flower Chloroform Extract (NSFCExt) Preparation
4.3. Isolation of Crystal by Column Chromatography
4.4. Crystal Structure Refinement Using X-Ray Diffraction Method
4.5. Cell Culture
4.6. In Vitro Cytotoxicity Assay Using 3-(4,5-Dimethylthiazol-2yl)-2,5-Diphenyl Tetrazolium Bromide (MTT)
4.7. Measurement of Intracellular Reactive Oxygen Species (ROS)
4.8. Apoptosis Related Cellular and Nuclear Morphology Analysis
4.9. Terminal Deoxynucleotidyl Transferase (TdT)-Mediated dUTP Nick End Labeling (TUNEL) Assay
4.10. Quantitative Real-Time Polymerase Chain Reaction (RT-PCR) Analysis of Tumor Suppressor, Oxidative Stress, and Apoptosis Related Genes
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Prakash, O.; Kumar, A.; Kumar, P.; Ajeet, A. Anticancer Potential of Plants and Natural Products: A Review. Am. J. Pharmacol. Sci. 2013, 1, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Indran, I.R.; Tufo, G.; Pervaiz, S.; Brenner, C. Recent advances in apoptosis, mitochondria and drug resistance in cancer cells. Biochim. et Biophys. Acta (BBA) Bioenerg. 2011, 1807, 735–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, T.-J.; Han, L.-H.; Cong, R.-S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. et Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Chin, L.; Pomerantz, J.; A DePinho, R. The INK4a/ARF tumor suppressor: One gene—two products—two pathways. Trends Biochem. Sci. 1998, 23, 291–296. [Google Scholar] [CrossRef]
- Symonds, H.; Krall, L.; Remington, L.; Saenz-Robles, M.; Lowe, S.; Jacks, T.; Van Dyke, T. p53-Dependent apoptosis suppresses tumor growth and progression in vivo. Cell 1994, 78, 703–711. [Google Scholar] [CrossRef]
- Tas, F.; Hansel, H.; Belce, A.; Ilvan, S.; Argon, A.; Camlica, H.; Topuz, E. Oxidative Stress in Breast Cancer. Med Oncol. 2005, 22, 11–16. [Google Scholar] [CrossRef]
- Bruce, J.G. Plant Sterols as Anticancer Nutrients: Evidence for Their Role in Breast Cancer. Nutrients 2013, 5, 359–387. [Google Scholar]
- Bu, D.-X.; Erl, W.; de Martin, R.; Hansson, G.K.; Yan, Z.-Q. IKKbeta-dependent NF-kappaB pathway controls vascular inflammation and intimal hyperplasia. FASEB J. 2005, 19, 1293–1295. [Google Scholar] [CrossRef]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of Natural Products on Developing New Anti-Cancer Agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef]
- Ignacimuthu, S.; Subash-Babu, P. A process for preparation of a novel compound, Nymphayol [17-(hexan-2-yl)-10,13-dimehylhexadechydro-1H-cyclopenta[a]phenantheren-3-ol] from Nymphaea stellata willd. Flower with antidiabetic property. Indian Patent Journal No. 1567/CHE/A, 2007. [Google Scholar]
- Raja, M.M.M.; Sethiya, N.; Mishra, S.H. A comprehensive review on Nymphaea stellata: A traditionally used bitter. J. Adv. Pharm. Technol. Res. 2010, 1, 311–319. [Google Scholar] [CrossRef] [Green Version]
- Subash-Babu, P.; Ignacimuthu, S.; Alshatwi, A. Nymphayol increases glucose-stimulated insulin secretion by RIN-5F cells and GLUT4-mediated insulin sensitization in type 2 diabetic rat liver. Chem.-Biol. Interact. 2015, 226, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Antonisamy, P.; Subash-Babu, P.; Alshatwi, A.; Aravinthan, A.; Ignacimuthu, S.; Choi, K.C.; Kim, J.-H. Gastroprotective effect of nymphayol isolated from Nymphaea stellata (Willd.) flowers: Contribution of antioxidant, anti-inflammatory and anti-apoptotic activities. Chem. Interact. 2014, 224, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Min, E.-Y.; Kim, I.-H.; Lee, J.; Kim, E.-Y.; Choi, Y.-H.; Nam, T.-J. The effects of fucodian on senescence are controlled by the p16INK4a-pRb and p14Arf-p53 pathways in hepatocellular carcinoma and hepatic cell lines. Int. J. Oncol. 2014, 45, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvakumari, E.S.; Shantha, S.; Prabhu, P.T.; Sreenathkumar, T.C. Antiproliferative activity of Ethanolic flower extract from Nymphaea pubescens willd against human cervical and breast carcinoma in vitro. Int. Res. J. Pharm. 2012, 3, 124–125. [Google Scholar]
- Goff, M.L.; Ferrec, E.L.; Mayer, C.; Mimouni, V.; Lagadic-Gossmann, D.; Schoefs, B.; Ulmann, L. Microalgal carotenoids and phytosterols regulate biochemical mechanisms involved in human health and disease prevention. Biochimie 2019, 167, 106–118. [Google Scholar] [CrossRef]
- Cadenas, E.; Davies, K.J. Mitochondrial free radical generation, oxidative stress, and aging. Free. Radic. Boil. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Subash-Babu, P.; Alshammari, G.M.; Ignacimuthu, S.; Alshatwi, A. Epoxy clerodane diterpene inhibits MCF-7 human breast cancer cell growth by regulating the expression of the functional apoptotic genes Cdkn2A, Rb1, mdm2 and p53. Biomed. Pharmacother. 2017, 87, 388–396. [Google Scholar] [CrossRef]
- Yadav, D.K.; Rai, R.; Kumar, N.; Surjeet, S.; Sanjeev, M.; Praveen, S.; Priyanka, S.; Horacio, P.; Ricardo, L.M.; Eun, H.C.; et al. New arylated benzo[h]quinolines induce anti-cancer activity by oxidative stress-mediated DNA damage. Sci. Rep. 2016, 6, 38128. [Google Scholar] [CrossRef]
- Nag, S.; Qin, J.-J.; Srivenugopal, K.S.; Wang, M.; Zhang, R. The MDM2-p53 pathway revisited. J. Biomed. Res. 2013, 27, 254–271. [Google Scholar]
- Lawrence, H.R.; Li, Z.; Yip, M.R.; Sung, S.-S.; Lawrence, N.J.; McLaughlin, M.L.; McManus, G.; Zaworotko, M.J.; Sebti, S.M.; Chen, J.; et al. Identification of a disruptor of the MDM2-p53 protein–protein interaction facilitated by high-throughput in silico docking. Bioorganic Med. Chem. Lett. 2009, 19, 3756–3759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shangary, S.; Wang, S. Small-molecule inhibitors of the MDM2-p53 protein-protein interaction to reactivate p53 function: A novel approach for cancer therapy. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 223–241. [Google Scholar] [CrossRef] [Green Version]
- Akl, H.; Vervloessem, T.; Kiviluoto, S.; Bittremieux, M.; Parys, J.B.; De Smedt, H.; Bultynck, G. A dual role for the anti-apoptotic Bcl-2 protein in cancer: Mitochondria versus endoplasmic reticulum. Biochim. et Biophys. Acta (BBA) Bioenerg. 2014, 1843, 2240–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.C.; Clark, G.M.; Allred, D.C.; Mohsin, S.; Chamness, G.; Elledge, R.M. Survival of patients with metastatic breast carcinoma. Cancer 2003, 97, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.; McDonnell, J.M.; Korsmeyer, S.J. BCL-2 family members and the mitochondria in apoptosis. Genome Res. 1999, 13, 1899–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, P. Mitochondria, Bioenergetics and Apoptosis in Cancer. Trends Cancer 2017, 3, 857–870. [Google Scholar] [CrossRef]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Boil. 2008, 9, 231–241. [Google Scholar] [CrossRef]
- Wang, X.; Ohnishi, T. p53-dependent signal transduction induced by stress. J. Radiat. Res. 1997, 38, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Dulic, V.; Stein, G.H.; Far, D.F.; Reed, S.I. Nuclear accumulation of p21Cip1 at the onset of mitosis: A role at the G2/M-phase transition. Mol. Cell Biol. 1998, 18, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichloro fluorescein assay using microplate reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Leite, M.; Quinta-Costa, M.; Leite, P.S.; Guimaraes, J.E. Critical Evaluation of Techniques to Detect and Measure Cell Death – Study in a Model of UV Radiation of the Leukaemic Cell Line HL60. Anal. Cell. Pathol. 1999, 19, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, D.B. Multiple Range Tests for Correlated and Heteroscedastic Means. Biometrics 1957, 13, 164–176. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Empirical formula | C50 H86 O3 |
| Formula weight | 735.19 |
| Temperature | 293(2) K |
| Wavelength | 0.71073 A |
| Crystal system, space group | Monoclinic, P21 |
| Unit cell dimensions | a = 9.618(5) A alpha = 90.000(5) deg. b = 7.518(5) A beta = 94.483(5) deg. c = 37.491(5) A gamma = 90.000(5) deg. |
| Volume | 2703(2) A3 |
| Z, Calculated density | 2, 0.903 Mg/m3 |
| Absorption coefficient | 0.054 mm−1 |
| F(000) | 820 |
| Crystal size | 0.3 × 0.1 × 0.1 mm |
| Theta range for data collection | 2.12 to 20.45 deg. |
| Limiting indices | −9 ≤ h ≤ 9, −7 ≤ k ≤ 7, −36 ≤ l ≤ 36 |
| Reflections collected/unique | 16593/5337 [R(int) = 0.0406] |
| Completeness to theta = 20.45 | 99.2% |
| Absorption correction | Semi-empirical from equivalents |
| Max. and min. transmission | 0.9560 and 0.9140 |
| Refinement method | Full-matrix least-squares on F2 |
| Data/restraints/parameters | 5337/11/484 |
| Goodness-of-fit on F^2 | 1.105 |
| Final R indices [I > 2sigma(I)] | R1 = 0.0842, wR2 = 0.2505 |
| R indices (all data) | R1 = 0.0991, wR2 = 0.2687 |
| Absolute structure parameter | −2(4) |
| Extinction coefficient | 0.0013(17) |
| Largest diff. peak and hole | 0.404 and −0.218 e.A−3 |
| Primer | Forward Sequence (5’ to 3’) | Reverse Sequence (5’ to 3’) |
|---|---|---|
| CYP1A1 | GCTGACTTCATCCCTATTCTTCG | TTTTGTAGTGCTCCTTGACCATCT |
| GSK3β | GGAACTCCAACAAGGGAGCA | TTCGGGGTCGGAAGACCTT |
| GPX | GTGCTCGGCTTCCCGTGCAAC | CTCGAAGAGCATGAAGTTGGGC |
| TNF-α | CTCTTCTGCCTGCTGCACTTTG | ATGGGCTACAGGCTTGTCACTC |
| NF-κB | GCGCTTCTCTGCCTTCCTTA | TCTTCAGGTTTGATGCCCCC |
| CDKN2A | CCTTCCAATGACTCCCTCC | TCAGAAACCCTAGTTCAAAGGA |
| pRb1 | CTCGTGCTGATGCTACTGAGGA | GGTCGGCGCAGTTGGGCTCC |
| p53 | CCTCAGCATCTTATCCGAGTGG | TGGATGGTGGTACAGTCAGAGC |
| mdm2 | CCCAAGACAAAGAAGAGAGTGTGG | CTGGGCAGGGCTTATTCCTTTTCT |
| Bax | TCAGGATGCGTCCACCAAGAAG | TGTGTCCACGGCGGCAATCATC |
| Bcl-2 | GTGGATGACTGAGTACCT | CCAGGAGAAATCAAACAGAG |
| Caspase-3 | ACATGGAAGCGAATCAATGGACTC | AAGGACTCAAATTCTGTTGCCACC |
| Caspase-8 | CCGCAAAGGAAGCAAGAA | GGTAGGTAATCAGCAAATCCAGT |
| Caspase- 9 | GCTCTTCCTTTGTTCATC | CTCTTCCTCCACTGTTCA |
| CDKN1A | AGGTGGACCTGGAGACTCTCAG | TCCTCTTGGAGAAGATCAGCCG |
| PCNA | CAAGTAATGTCGATAAAGAGGAGG | GTGTCACCGTTGAAGAGAGTGG |
| GAPDH | GTCTCCTCTGACTTCAACAGCG | ACCACCCTGTTGCTGTAGCCAA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Harbi, L.N.; Subash-Babu, P.; Binobead, M.A.; Alhussain, M.H.; AlSedairy, S.A.; Aloud, A.A.; Alshatwi, A.A. Potential Metabolite Nymphayol Isolated from Water Lily (Nymphaea stellata) Flower Inhibits MCF-7 Human Breast Cancer Cell Growth via Upregulation of Cdkn2a, pRb2, p53 and Downregulation of PCNA mRNA Expressions. Metabolites 2020, 10, 280. https://doi.org/10.3390/metabo10070280
Al-Harbi LN, Subash-Babu P, Binobead MA, Alhussain MH, AlSedairy SA, Aloud AA, Alshatwi AA. Potential Metabolite Nymphayol Isolated from Water Lily (Nymphaea stellata) Flower Inhibits MCF-7 Human Breast Cancer Cell Growth via Upregulation of Cdkn2a, pRb2, p53 and Downregulation of PCNA mRNA Expressions. Metabolites. 2020; 10(7):280. https://doi.org/10.3390/metabo10070280
Chicago/Turabian StyleAl-Harbi, Laila Naif, Pandurangan Subash-Babu, Manal Abdulaziz Binobead, Maha Hussain Alhussain, Sahar Abdulaziz AlSedairy, Amal A Aloud, and Ali A Alshatwi. 2020. "Potential Metabolite Nymphayol Isolated from Water Lily (Nymphaea stellata) Flower Inhibits MCF-7 Human Breast Cancer Cell Growth via Upregulation of Cdkn2a, pRb2, p53 and Downregulation of PCNA mRNA Expressions" Metabolites 10, no. 7: 280. https://doi.org/10.3390/metabo10070280
APA StyleAl-Harbi, L. N., Subash-Babu, P., Binobead, M. A., Alhussain, M. H., AlSedairy, S. A., Aloud, A. A., & Alshatwi, A. A. (2020). Potential Metabolite Nymphayol Isolated from Water Lily (Nymphaea stellata) Flower Inhibits MCF-7 Human Breast Cancer Cell Growth via Upregulation of Cdkn2a, pRb2, p53 and Downregulation of PCNA mRNA Expressions. Metabolites, 10(7), 280. https://doi.org/10.3390/metabo10070280