Diabetes Leads to Alterations in Normal Metabolic Transitions of Pregnancy as Revealed by Time-Course Metabolomics



Abstract
:1. Introduction
2. Results
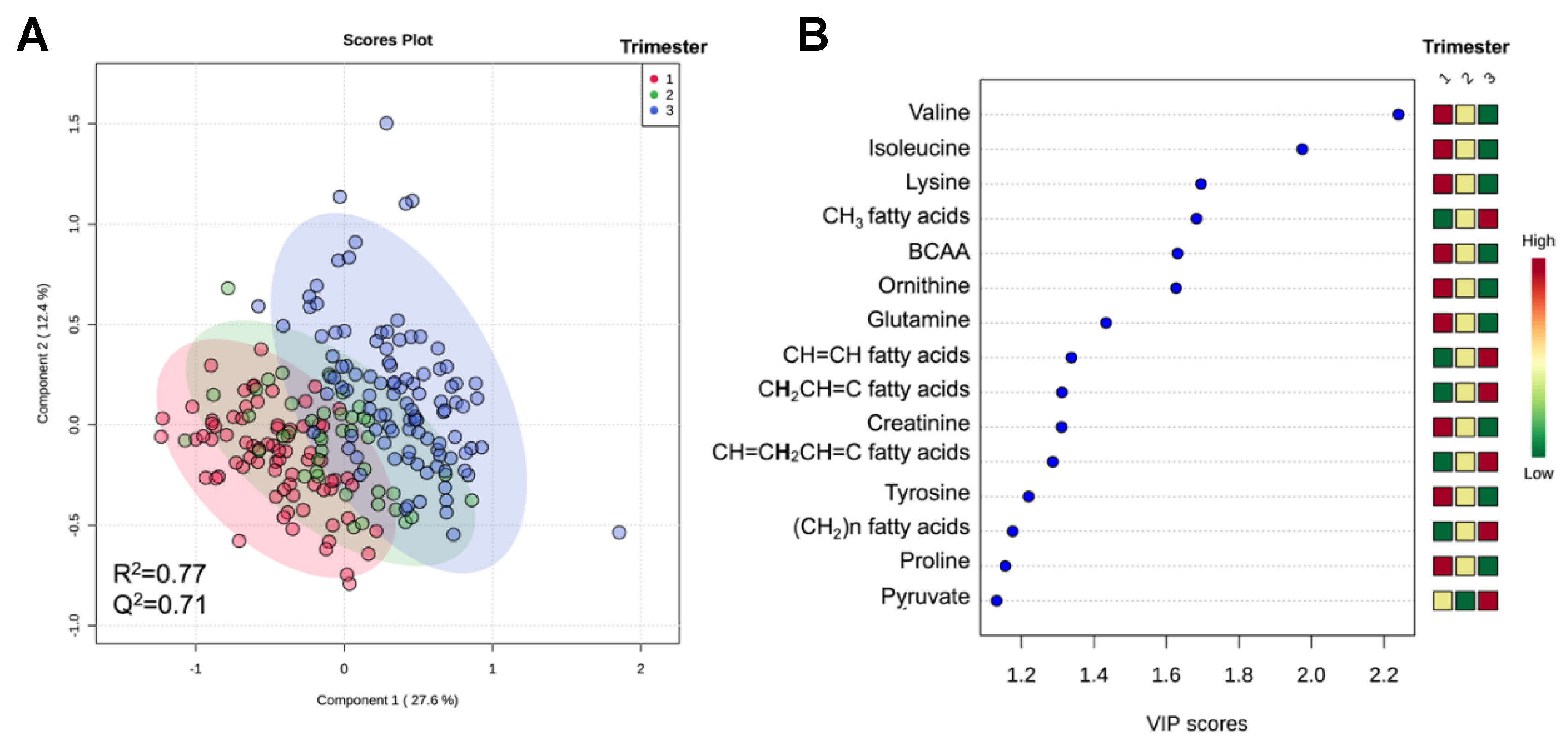
2.1. Metabolomic Analysis Reveals Maternal Metabolic Alterations in Serum across Gestation in Women with and without Diabetes
2.2. Metabolomic Analysis Reveals Maternal Metabolic Alterations across Gestation in Women with PGDM
2.3. Metabolomic Analysis Reveals Maternal Metabolic Alterations Across Gestation in Women with GDM
2.4. Metabolomic Analysis Reveals Maternal Metabolic Alterations in Women with PGDM or GDM Immediately Following Delivery
2.5. Cord Blood from Newborns of Women with PGDM or GDM Reveals Similar Metabolic Patterns
3. Discussion
3.1. Amino Acids, TCA Cycle Intermediates, Carbohydrates, and Lipids Are Altered during Pregnancy Regardless of Diabetes Diagnosis
3.2. Amino Acid, Carbohydrate, and Lipid Metabolism Are Altered in Women with PGDM during Pregnancy and the Immediate Post-Partum Period
3.3. Amino Acid, Carbohydrate, and Lipid Metabolism Changes in Women with GDM during Pregnancy
3.4. Carbohydrate and Choline Metabolism Are Altered in the Cord Blood of Newborns from Women with PGDM or GDM
4. Conclusions
5. Materials and Methods
5.1. Sample Collection
5.2. Metabolomic Analysis
5.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Emerson, K., Jr.; Saxena, B.N.; Poindexter, E.L. Caloric cost of normal pregnancy. Obstet. Gynecol. 1972, 40, 786–794. [Google Scholar]
- Brizzi, P.; Tonolo, G.; Esposito, F.; Puddu, L.; Dessole, S.; Maioli, M.; Milia, S. Lipoprotein metabolism during normal pregnancy. Am. J. Obstet. Gynecol. 1999, 181, 430–434. [Google Scholar]
- Lindsay, K.L.; Hellmuth, C.; Uhl, O.; Buss, C.; Wadhwa, P.D.; Koletzko, B.; Entringer, S. Longitudinal Metabolomic Profiling of Amino Acids and Lipids across Healthy Pregnancy. PLoS ONE 2015, 10, e0145794. [Google Scholar] [CrossRef]
- Pinto, J.; Barros, A.S.; Domingues, M.R.; Goodfellow, B.J.; Galhano, E.; Pita, C.; Almeida Mdo, C.; Carreira, I.M.; Gil, A.M. Following healthy pregnancy by NMR metabolomics of plasma and correlation to urine. J. Proteome Res. 2015, 14, 1263–1274. [Google Scholar] [CrossRef]
- Carr, B.R.; Parker, C.R.; Madden, J.D.; MacDonald, P.C.; Porter, J.C. Maternal plasma adrenocorticotropin and cortisol relationships throughout human pregnancy. Am. J. Obstet. Gynecol. 1981, 139, 416–422. [Google Scholar]
- Noctor, E.; Dunne, F.P. Type 2 diabetes after gestational diabetes: The influence of changing diagnostic criteria. World J. Diabetes 2015, 6, 234–244. [Google Scholar] [CrossRef]
- Vilar, L.; Freitas Mda, C.; Lima, L.H.; Lyra, R.; Kater, C.E. Cushing’s syndrome in pregnancy: An overview. Arq. Bras. Endocrinol. Metabol. 2007, 51, 1293–1302. [Google Scholar]
- Hoyert, D.L. Maternal mortality and related concepts. Vital Health Stat. 2007, 3, 1–13. [Google Scholar]
- Hoyert, D.L.; Xu, J. Deaths: Preliminary. Vital Health Stat. 2012, 3, 45. [Google Scholar]
- Buchanan, T. Pregnancy in Preexisting Diabetes. In Diabetes in America, 2nd ed.; Harris, M.I., Cowie, C.C., Stern, M.P., Boyko, E.J., Reiber, G.E., Bennet, P.H., Eds.; NIH Publication no. 95–1468; National Institutes of Health, National Institute of Diabetes and Digestive and Kidney Disease: Bethesda, MD, USA, 1995; pp. 719–734. [Google Scholar]
- Lawrence, J.M.; Contreras, R.; Chen, W.; Sacks, D.A. Trends in the prevalence of preexisting diabetes and gestational diabetes mellitus among a racially/ethnically diverse population of pregnant women, 1999–2005. Diabetes Care 2008, 31, 899–904. [Google Scholar] [CrossRef] [Green Version]
- Temple, R.; Murphy, H. Type 2 diabetes in pregnancy—An increasing problem. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 591–603. [Google Scholar] [CrossRef]
- Desisto, C.L.; Kim, S.Y.; Sharma, A.J. Prevalence Estimates of Gestational Diabetes Mellitus in the United States, Pregnancy Risk Assessment Monitoring System (PRAMS), 2007–2010. Prev. Chronic Dis. 2014, 11, 1–9. [Google Scholar]
- Barros, S.; Goodfellow, B.J.; Duarte, I.F.; Carreira, I.M.; Pita, C.; Gil, A.M. Following Healthy Pregnancy by Nuclear Magnetic Resonance (NMR) Metabolic Pro fi ling of Human Urine. J. Proteome Res. 2013, 12, 969–979. [Google Scholar]
- Chorell, E.; Hall, U.A.; Gustavsson, C.; Berntorp, K.; Puhkala, J.; Luoto, R.; Olsson, T.; Holmang, A. Pregnancy to postpartum transition of serum metabolites in women with gestational diabetes. Metabolism 2017, 72, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Allalou, A.; Nalla, A.; Prentice, K.J.; Liu, Y.; Zhang, M.; Dai, F.F.; Ning, X.; Osborne, L.R.; Cox, B.J.; Gunderson, E.P.; et al. A Predictive Metabolic Signature for the Transition From Gestational Diabetes Mellitus to Type 2 Diabetes. Diabetes 2016, 65, 2529–2539. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.P.; Reichetzeder, C.; Prehn, C.; von Websky, K.; Slowinski, T.; Chen, Y.P.; Yin, L.H.; Kleuser, B.; Yang, X.S.; Adamski, J.; et al. Fetal Serum Metabolites Are Independently Associated with Gestational Diabetes Mellitus. Cell. Physiol. Biochem. 2018, 45, 625–638. [Google Scholar] [CrossRef]
- Battaglia, F.C.; Meschia, G. Principle Substrates of Fetal Metabolism. Physiol. Rev. 1978, 58, 499–527. [Google Scholar]
- Lemons, J.A.; Adcock, E.W.; Jones, M.D.; Naughton, M.A.; Meschia, G.; Battaglia, F.C. Umbilical uptake of amino acids in the unstressed fetal lamb. J. Clin. Investig. 1976, 58, 1428–1434. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, P.R.; Lobo, C.; Battaglia, F.C.; Fennessey, P.V.; Wilkening, R.B.; Meschia, G. Glutamine Glutamate Exchange between Placenta and Fetal Liver. Am. J. Physiol. Endocrinol. Metab. 1995, 268, E705–E711. [Google Scholar]
- Schneider, H.; Mohlen, K.H.; Challier, J.C.; Dancis, J. Transfer of Glutamic-Acid across the Human-Placenta Perfused Invitro. Int. J. Obstet. Gynaec. 1979, 86, 299–306. [Google Scholar] [CrossRef]
- Herrera, E.; Amusquivar, E.; Lopez-Soldado, I.; Ortega, H. Maternal lipid metabolism and placental lipid transfer. Horm. Res. 2006, 65, 59–64. [Google Scholar] [CrossRef]
- Felig, P.; Lynch, V. Starvation in human pregnancy: Hypoglycemia, hypoinsulinemia, and hyperketonemia. Science 1970, 170, 990–992. [Google Scholar]
- Herrera, E.; Knopp, R.H.; Freinkel, N. Carbohydrate metabolism in pregnancy. VI. Plasma fuels, insulin, liver composition, gluconeogenesis, and nitrogen metabolism during late gestation in the fed and fasted rat. J. Clin. Investig. 1969, 48, 2260–2272. [Google Scholar] [CrossRef]
- Sweiry, J.H.; Page, K.R.; Dacke, C.G.; Abramovich, D.R.; Yudilevich, D.L. Evidence of Saturable Uptake Mechanisms at Maternal and Fetal Sides of the Perfused Human-Placenta by Rapid Paired-Tracer Dilution—Studies with Calcium and Choline. J. Dev. Physiol. 1986, 8, 435–445. [Google Scholar]
- Zeisel, S.H. Choline: Critical role during fetal development and dietary requirements in adults. Annu Rev. Nutr. 2006, 26, 229–250. [Google Scholar] [CrossRef] [Green Version]
- Ozarda Ilcol, Y.; Uncu, G.; Ulus, I.H. Free and phospholipid-bound choline concentrations in serum during pregnancy, after delivery and in newborns. Arch. Physiol. Biochem. 2002, 110, 393–399. [Google Scholar] [CrossRef]
- Hytten, F.; Chamberlain, G. Nutrition. In Clinical Physiology in Obstetrics; Blackwell Scientific Publications: Oxford, UK, 1980; pp. 163–192. [Google Scholar]
- Staat, B.C.; Galan, H.L.; Harwood, J.E.F.; Lee, G.; Marconi, A.M.; Paolini, C.L.; Cheung, A.; Battaglia, F.C. Transplacental Supply of Mannose and Inositol in Uncomplicated Pregnancies Using Stable Isotopes. J. Clin. Endocrinol. Metab. 2012, 97, 2497–2502. [Google Scholar] [CrossRef] [Green Version]
- Schneider, A.; Thiel, C.; Rindermann, J.; DeRossi, C.; Popovici, D.; Hoffmann, G.F.; Grone, H.J.; Korner, C. Successful prenatal mannose treatment for congenital disorder of glycosylation-Ia in mice. Nat. Med. 2012, 18, 71–73. [Google Scholar] [CrossRef]
- Edwards, S.M.; Cunningham, S.A.; Dunlop, A.L.; Corwin, E.J. The Maternal Gut Microbiome During Pregnancy. MCN Am. J. Matern Child Nurs. 2017, 42, 310–317. [Google Scholar] [CrossRef]
- Fischer, M.J. Chronic kidney disease and pregnancy: Maternal and fetal outcomes. Adv. Chronic Kidney Dis. 2007, 14, 132–145. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Tavintharan, S.; Sum, C.F.; Woon, K.; Lim, S.C.; Ong, C.N. Metabolic signature shift in type 2 diabetes mellitus revealed by mass spectrometry-based metabolomics. J. Clin. Endocrinol. Metab. 2013, 98, E1060–E1065. [Google Scholar] [CrossRef]
- Gurke, J.; Hirche, F.; Thieme, R.; Haucke, E.; Schindler, M.; Stangl, G.I.; Fischer, B.; Navarrete Santos, A. Maternal Diabetes Leads to Adaptation in Embryonic Amino Acid Metabolism during Early Pregnancy. PLoS ONE 2015, 10, e0127465. [Google Scholar] [CrossRef] [Green Version]
- Palmer, N.D.; Stevens, R.D.; Antinozzi, P.A.; Anderson, A.; Bergman, R.N.; Wagenknecht, L.E.; Newgard, C.B.; Bowden, D.W. Metabolomic Profile Associated With Insulin Resistance and Conversion to Diabetes in the Insulin Resistance Atherosclerosis Study. J. Clin. Endocrinol. Metab. 2015, 100, E463–E468. [Google Scholar] [CrossRef]
- Thalacker-Mercer, A.E.; Ingram, K.H.; Guo, F.J.; Ilkayeva, O.; Newgard, C.B.; Garvey, W.T. BMI, RQ, Diabetes, and Sex Affect the Relationships Between Amino Acids and Clamp Measures of Insulin Action in Humans. Diabetes 2014, 63, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Floegel, A.; Stefan, N.; Yu, Z.H.; Muhlenbruch, K.; Drogan, D.; Joost, H.G.; Fritsche, A.; Haring, H.U.; de Angelis, M.H.; Peters, A.; et al. Identification of Serum Metabolites Associated With Risk of Type 2 Diabetes Using a Targeted Metabolomic Approach. Diabetes 2013, 62, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Wang-Sattler, R.; Yu, Z.; Herder, C.; Messias, A.C.; Floegel, A.; He, Y.; Heim, K.; Campillos, M.; Holzapfel, C.; Thorand, B.; et al. Novel biomarkers for pre-diabetes identified by metabolomics. Mol. Syst. Biol. 2012, 8, 615. [Google Scholar] [CrossRef]
- Gannon, M.C.; Nuttall, J.A.; Nuttall, F.Q. The metabolic response to ingested glycine. Am. J. Clin. Nutr. 2002, 76, 1302–1307. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Ortiz, M.; Medina-Santillan, R.; Martinez-Abundis, E.; von Drateln, C.R. Effect of glycine on insulin secretion and action in healthy first-degree relatives of type 2 diabetes mellitus patients. Horm. Metab. Res. 2001, 33, 358–360. [Google Scholar]
- Drabkova, P.; Sanderova, J.; Kovarik, J.; kandar, R. An Assay of Selected Serum Amino Acids in Patients with Type 2 Diabetes Mellitus. Adv. Clin. Exp. Med. 2015, 24, 447–451. [Google Scholar] [CrossRef] [Green Version]
- Stancakova, A.; Civelek, M.; Saleem, N.K.; Soininen, P.; Kangas, A.J.; Cederberg, H.; Paananen, J.; Pihlajamaki, J.; Bonnycastle, L.L.; Morken, M.A.; et al. Hyperglycemia and a common variant of GCKR are associated with the levels of eight amino acids in 9,369 Finnish men. Diabetes 2012, 61, 1895–1902. [Google Scholar] [CrossRef] [Green Version]
- Kimura, K.; Nakamura, Y.; Inaba, Y.; Matsumoto, M.; Kido, Y.; Asahara, S.; Matsuda, T.; Watanabe, H.; Maeda, A.; Inagaki, F.; et al. Histidine Augments the Suppression of Hepatic Glucose Production by Central Insulin Action. Diabetes 2013, 62, 2266–2277. [Google Scholar] [CrossRef] [Green Version]
- Menge, B.A.; Schrader, H.; Ritter, P.R.; Ellrichmann, M.; Uhl, W.; Schmidt, W.E.; Meier, J.J. Selective amino acid deficiency in patients with impaired glucose tolerance and type 2 diabetes. Regul. Pept. 2010, 160, 75–80. [Google Scholar] [CrossRef]
- Akazawa, S.; Metzger, B.E.; Freinkel, N. Relationships between Glucose and Mannose during Late Gestation in Normal-Pregnancy and Pregnancy Complicated by Diabetes-Mellitus—Concurrent Concentrations in Maternal Plasma and Amniotic-Fluid. J. Clin. Endocrinol. Metab. 1986, 62, 984–989. [Google Scholar] [CrossRef]
- Pitkanen, E. Mannose, mannitol, fructose and 1,5-anhydroglucitol concentrations measured by gas chromatography/mass spectrometry in blood plasma of diabetic patients. Clin. Chim. Acta 1996, 251, 91–103. [Google Scholar]
- Aloia, J.F. Monosaccharides and polyols in diabetes mellitus and uremia. J. Lab. Clin. Med. 1973, 82, 809–817. [Google Scholar]
- Mardinoglu, A.; Stancakova, A.; Lotta, L.A.; Kuusisto, J.; Boren, J.; Bluher, M.; Wareham, N.J.; Ferrannini, E.; Groop, P.H.; Laakso, M.; et al. Plasma Mannose Levels Are Associated with Incident Type 2 Diabetes and Cardiovascular Disease. Cell Metab. 2017, 26, 281–283. [Google Scholar] [CrossRef]
- Fahien, L.A.; MacDonald, M.J. The succinate mechanism of insulin release. Diabetes 2002, 51, 2669–2676. [Google Scholar]
- Dodd, M.S.; Sousa Fialho, M.D.L.; Montes Aparicio, C.N.; Kerr, M.; Timm, K.N.; Griffin, J.L.; Luiken, J.; Glatz, J.F.C.; Tyler, D.J.; Heather, L.C. Fatty Acids Prevent Hypoxia-Inducible Factor-1alpha Signaling Through Decreased Succinate in Diabetes. JACC Basic Transl. Sci. 2018, 3, 485–498. [Google Scholar] [CrossRef]
- Liu, T.; Li, J.; Xu, F.; Wang, M.; Ding, S.; Xu, H.; Dong, F. Comprehensive analysis of serum metabolites in gestational diabetes mellitus by UPLC/Q-TOF-MS. Anal. Bioanal. Chem. 2016, 408, 1125–1135. [Google Scholar] [CrossRef]
- Pinto, J.; Almeida, L.M.; Martins, A.S.; Duarte, D.; Barros, A.S.; Galhano, E.; Pita, C.; Almeida, M.D.C.; Carreira, I.M.; Gil, A.M. Prediction of Gestational Diabetes through NMR Metabolomics of Maternal Blood. J. Proteome Res. 2015, 14, 2696–2706. [Google Scholar] [CrossRef]
- Rahimi, N.; Razi, F.; Nasli-Esfahani, E.; Qorbani, M.; Shirzad, N.; Larijani, B. Amino acid profiling in the gestational diabetes mellitus. J. Diabetes Metab. Disord. 2017, 16, 13. [Google Scholar] [CrossRef] [Green Version]
- Pappa, K.I.; Vlachos, G.; Theodora, M.; Roubelaki, M.; Angelidou, K.; Antsaklis, A. Intermediate metabolism in association with the amino acid profile during the third trimester of normal pregnancy and diet-controlled gestational diabetes. Am. J. Obstet. Gynecol. 2007, 196, e61–e65. [Google Scholar] [CrossRef]
- Nevalainen, J.; Sairanen, M.; Appelblom, H.; Gissler, M.; Timonen, S.; Ryynanen, M. First-Trimester Maternal Serum Amino Acids and Acylcarnitines Are Significant Predictors of Gestational Diabetes. Rev. Diabet. Stud. 2016, 13, 236–245. [Google Scholar] [CrossRef]
- Yan-Do, R.; MacDonald, P.E. Impaired “Glycine”-mia in Type 2 Diabetes and Potential Mechanisms Contributing to Glucose Homeostasis. Endocrinology 2017, 158, 1064–1073. [Google Scholar] [CrossRef]
- Washburn, S.E.; Caudill, M.A.; Malysheva, O.; MacFarlane, A.J.; Behan, N.A.; Harnett, B.; MacMillan, L.; Pongnopparat, T.; Brosnan, J.T.; Brosnan, M.E. Formate metabolism in fetal and neonatal sheep. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E921–E927. [Google Scholar] [CrossRef]
- Maitre, L.; Fthenou, E.; Athersuch, T.; Coen, M.; Toledano, M.B.; Holmes, E.; Kogevinas, M.; Chatzi, L.; Keun, H.C. Urinary metabolic profiles in early pregnancy are associated with preterm birth and fetal growth restriction in the Rhea mother-child cohort study. BMC Med. 2014, 12, 110. [Google Scholar] [CrossRef] [Green Version]
- Boden, G.; Laakso, M. Lipids and glucose in type 2 diabetes: What is the cause and effect? Diabetes Care 2004, 27, 2253–2259. [Google Scholar]
- Verges, B. Lipid disorders in type 1 diabetes. Diabetes Metab. 2009, 35, 353–360. [Google Scholar] [CrossRef]
- Torri, G.M.; Torri, J.; Gulian, J.M.; Vion-Dury, J.; Viout, P.; Cozzone, P.J. Magnetic resonance spectroscopy of serum and acute-phase proteins revisited: A multiparametric statistical analysis of metabolite variations in inflammatory, infectious and miscellaneous diseases. Clin. Chim. Acta 1999, 279, 77–96. [Google Scholar]
- Barbour, L.A.; McCurdy, C.E.; Hernandez, T.L.; Kirwan, J.P.; Catalano, P.M.; Friedman, J.E. Cellular mechanisms for insulin resistance in normal pregnancy and gestational diabetes. Diabetes Care 2007, 30 (Suppl. 2), S112–S119. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, J. Diabetes and pregnancy; blood sugar of newborn infants during fasting and glucose administration. Nord. Med. 1952, 47, 1049. [Google Scholar]
- Desoye, G.; Nolan, C.J. The fetal glucose steal: An underappreciated phenomenon in diabetic pregnancy. Diabetologia 2016, 59, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.; Teng, C.; Timmerman, M.; Meschia, G.; Battaglia, F.C. Production and utilization of amino acids by ovine placenta in vivo. Am. J. Physiol. 1998, 274, E13–E22. [Google Scholar]
- Holm, M.B.; Bastani, N.E.; Holme, A.M.; Zucknick, M.; Jansson, T.; Refsum, H.; Morkrid, L.; Blomhoff, R.; Henriksen, T.; Michelsen, T.M. Uptake and release of amino acids in the fetal-placental unit in human pregnancies. PLoS ONE 2017, 12, e0185760. [Google Scholar] [CrossRef]
- Shaw, G.M.; Finnell, R.H.; Blom, H.J.; Carmichael, S.L.; Vollset, S.E.; Yang, W.; Ueland, P.M. Choline and risk of neural tube defects in a folate-fortified population. Epidemiology 2009, 20, 714–719. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, A.; Piquard, F.; Dellenbach, P.; Haberey, P. Placenta-fetal “alanine-lactate cycle” in the human during late gestation. Placenta 1993, 14, 103–114. [Google Scholar]
- Tuuli, M.G.; Stout, M.J.; Shanks, A.; Odibo, A.O.; Macones, G.A.; Cahill, A.G. Umbilical Cord Arterial Lactate Compared With pH for Predicting Neonatal Morbidity at Term. Obstet. Gynecol. 2014, 124, 756–761. [Google Scholar] [CrossRef] [Green Version]
- Dona, A.C.; Jimenez, B.; Schafer, H.; Humpfer, E.; Spraul, M.; Lewis, M.R.; Pearce, J.T.M.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Precision High-Throughput Proton NMR Spectroscopy of Human Urine, Serum, and Plasma for Large-Scale Metabolic Phenotyping. Anal. Chem. 2014, 86, 9887–9894. [Google Scholar] [CrossRef]
- Le Guennec, A.; Tayyari, F.; Edison, A.S. Alternatives to Nuclear Overhauser Enhancement Spectroscopy Presat and Carr-Purcell-Meiboom-Gill Presat for NMR-Based Metabolomics. Anal. Chem. 2017, 89, 8582–8588. [Google Scholar] [CrossRef] [Green Version]
- Bingol, K.; Li, D.W.; Zhang, B.; Bruschweiler, R. Comprehensive Metabolite Identification Strategy Using Multiple Two-Dimensional NMR Spectra of a Complex Mixture Implemented in the COLMARm Web Server. Anal. Chem. 2016, 88, 12411–12418. [Google Scholar] [CrossRef] [Green Version]
- Walejko, J.M.; Chelliah, A.; Keller-Wood, M.; Gregg, A.; Edison, A.S. Global Metabolomics of the Placenta Reveals Distinct Metabolic Profiles between Maternal and Fetal Placental Tissues Following Delivery in Non-Labored Women. Metabolites 2018, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar]
- Walejko, J.M.; Koelmel, J.P.; Garrett, T.J.; Edison, A.S.; Keller-Wood, M. Multiomics approach reveals metabolic changes in the heart at birth. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1212–E1223. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | p-Value | FDR | Change during Pregnancy (±) |
|---|---|---|---|
| HbA1c 1 | 4.22 × 10−3 | 5.33 × 10−3 | + |
| Amino Acids | |||
| Alanine | 7.34 × 10−4 | 1.01 × 10−3 | − |
| Arginine | 1.72 × 10−9 | 4.29 × 10−9 | − |
| BCAA 2 | 4.44 × 10−15 | 1.68 × 10−14 | − |
| Dimethylglycine | 1.55 × 10−4 | 2.29 × 10−4 | − |
| Glutamate | 1.41 × 10−5 | 2.26 × 10−5 | + |
| Glutamine | 3.42 × 10−18 | 1.51 × 10−17 | − |
| Glycine | 7.43 × 10−4 | 1.01 × 10−3 | − |
| Histidine | 8.38 × 10−5 | 1.27 × 10−4 | − |
| Isoleucine | 2.17 × 10−21 | 1.44 × 10−20 | − |
| Leucine | 5.21 × 10−5 | 8.12 × 10−5 | − |
| Lysine | 1.86 × 10−19 | 9.86 × 10−19 | − |
| Ornithine | 6.60 × 10−15 | 2.33 × 10−14 | − |
| Phenylalanine | 4.12 × 10−8 | 7.53 × 10−8 | + |
| Proline | 4.71 × 10−11 | 1.39 × 10−10 | − |
| Serine | 1.37 × 10−5 | 2.26 × 10−5 | − |
| Threonine | 1.46 × 10−3 | 1.89 × 10−3 | + |
| Tyrosine | 4.11 × 10−8 | 7.53 × 10−8 | − |
| Valine | 8.08 × 10−25 | 1.07 × 10−23 | − |
| TCA Cycle | |||
| Citrate | 3.34 × 10−9 | 7.38 × 10−9 | + |
| Lactate | 9.04 × 10−9 | 1.84 × 10−8 | + |
| Pyruvate | 8.08 × 10−11 | 2.25 × 10−10 | + |
| Carbohydrates | |||
| Mannose | 1.46 × 10−3 | 1.89 × 10−3 | + |
| UDP-Glucose 3 | 1.52 × 10−8 | 2.98 × 10−8 | + |
| Ketones | |||
| Acetoacetate | 5.61 × 10−4 | 8.03 × 10−4 | + |
| Acetone | 1.87 × 10−9 | 4.31 × 10−9 | + |
| 3-Hydroxybutyrate | 3.50 × 10−6 | 5.98 × 10−6 | + |
| Lipids | |||
| CH3 fatty acids | 9.37 × 10−34 | 4.97 × 10−32 | + |
| (-CH2)n fatty acids | 1.27 × 10−24 | 1.35 × 10−23 | + |
| Beta-CH2 fatty acids | 3.92 × 10−23 | 3.46 × 10−22 | + |
| CH2CH=C fatty acids | 1.30 × 10−26 | 2.30 × 10−25 | + |
| Alpha-CH2 fatty acids | 7.18 × 10−19 | 3.46 × 10−18 | + |
| C=CCH2C=C fatty acids | 8.87 × 10−23 | 6.72 × 10−22 | + |
| Glyceryl C1,3H | 4.08 × 10−12 | 1.35 × 10−11 | + |
| Glyceryl C2H | 4.75 × 10−20 | 2.80 × 10−19 | + |
| CH=CH fatty acids | 8.32 × 10−28 | 2.20 × 10−26 | + |
| Other | |||
| Betaine | 1.78 × 10−9 | 4.29 × 10−9 | − |
| Glycolipids (GPC/PC) 3,4 | 1.96 × 10−10 | 5.19 × 10−10 | − |
| N-acetyl-glycoproteins | 2.28 × 10−6 | 4.03 × 10−6 | + |
| Isobutyrate | 1.11 × 10−11 | 3.46 × 10−11 | + |
| Myoinositol | 6.25 × 10−3 | 7.70 × 10−3 | − |
| Creatinine | 3.21 × 10−15 | 1.31 × 10−14 | − |
| Dimethylsulfone | 8.56 × 10−9 | 1.81 × 10−8 | − |
| Metabolite | p-Value | FDR |
|---|---|---|
| PGDM 1 vs. Controls | ||
| Increased PGDM | ||
| HbA1c 2 | 4.81 × 10−21 | 2.55 × 10−19 |
| Glucose | 1.64 × 10−3 | 0.04 |
| Isoleucine | 7.47 × 10−3 | 0.08 |
| BCAA 3 | 0.04 | 0.15 |
| Mannose | 0.04 | 0.15 |
| Decreased PGDM | ||
| Glycerol | 4.62 × 10−3 | 0.08 |
| Creatinine | 6.24 × 10−3 | 0.08 |
| Threonine | 0.01 | 0.10 |
| Succinate | 0.01 | 0.11 |
| Glutamine | 0.02 | 0.12 |
| Glycine | 0.03 | 0.15 |
| Myoinositol | 0.03 | 0.15 |
| Histidine | 0.03 | 0.15 |
| Citrate | 0.03 | 0.15 |
| Dimethylglycine | 0.04 | 0.15 |
| GDM 4 vs. Controls | ||
| Increased GDM | ||
| Glyceryl C2H | 0.01 | 0.16 |
| Glyceryl C1,3H | 0.02 | 0.16 |
| Alpha-CH2 fatty acids | 0.03 | 0.16 |
| N-acetyl-glycoproteins | 0.03 | 0.16 |
| CH2CH=C fatty acids | 0.04 | 0.16 |
| (-CH2)n fatty acids | 0.04 | 0.16 |
| CH=CH fatty acids | 0.04 | 0.16 |
| Decreased GDM | ||
| Succinate | 0.01 | 0.16 |
| Formate | 0.02 | 0.16 |
| Serine | 0.02 | 0.16 |
| Threonine | 0.03 | 0.16 |
| Isobutyrate | 0.03 | 0.16 |
| Acetate | 0.03 | 0.16 |
| Glutamine | 0.05 | 0.16 |
| Glycine | 0.05 | 0.16 |
| Creatinine | 0.05 | 0.16 |
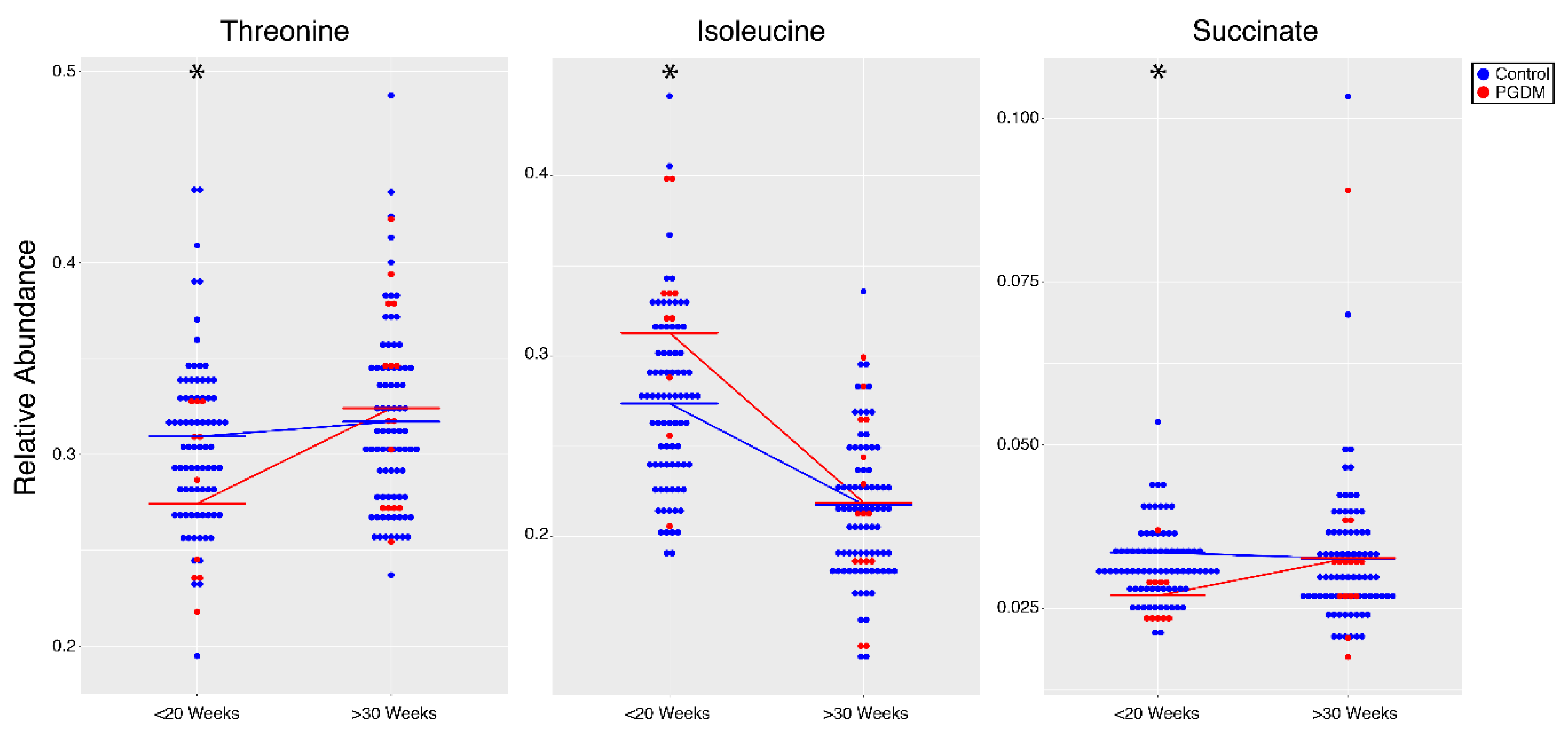
| Metabolite | p-Value | <20 Weeks | >30 Weeks |
|---|---|---|---|
| PGDM 1 vs. Controls | |||
| HbA1c 2 | 1.23 × 10−4 | PGDM > Ctrl | PGDM > Ctrl |
| Threonine | 0.01 | Ctrl > PGDM | - |
| Succinate | 0.02 | Ctrl > PGDM | - |
| Isoleucine | 0.03 | PGDM > Ctrl | - |
| Acetate | 0.03 | - | PGDM > Ctrl |
| Acetone | 0.04 | - | PGDM > Ctrl |
| GDM 3 vs. Controls | <18 Weeks | >30 Weeks | |
| Acetoacetate | 2.32 × 10−3 | - | GDM > Ctrl |
| Succinate | 6.44 × 10−3 | Ctrl > GDM | - |
| Acetate | 0.01 | - | - |
| Formate | 0.02 | Ctrl > GDM | - |
| 3-Hydroxybutyrate | 0.02 | - | GDM > Ctrl |
| Glucose | 0.03 | - | GDM > Ctrl |
| Glutamine | 0.04 | - | - |
| Pathway | Class | Significant Metabolites | p-Value | FDR | |
|---|---|---|---|---|---|
| PGDM 1 vs. Controls | Increased | Decreased | |||
| Galactose metabolism | Carbohydrate Metabolism | Glucose, Mannose | Glycerol, Myo-inositol | 8.77 × 10−5 | 3.68 × 10−3 |
| Glyoxylate and dicarboxylate metabolism | Carbohydrate Metabolism | Acetate | Citrate, Glycine, Glutamine | 1.75 × 10−4 | 4.89 × 10−3 |
| Alanine, aspartate and glutamate metabolism | Amino Acid Metabolism | - | Glutamine, Citrate, Succinate | 2.08 × 10−3 | 0.04 |
| Glycine, serine and threonine metabolism | Amino Acid Metabolism | - | Dimethylglycine, Glycine, Threonine | 3.36 × 10−3 | 0.05 |
| Citrate cycle (TCA cycle) | Carbohydrate Metabolism | - | Succinate, Citrate | 0.02 | 0.18 |
| GDM 2 vs. Controls | Increased | Decreased | |||
| Glyoxylate and dicarboxylate metabolism | Carbohydrate Metabolism | - | Serine, Glycine, Acetate, Formate, Glutamine | 1.94 × 10−6 | 1.63 × 10−4 |
| Butanoate metabolism | Carbohydrate Metabolism | 3-Hydroxybutyrate, Acetoacetate | Succinate | 1.53 × 10−4 | 6.44 × 10−3 |
| Synthesis and degradation of ketone bodies | Lipid Metabolism | 3-Hydroxybutyrate, Acetoacetate | - | 5.43 × 10−4 | 0.01 |
| Glycine, serine and threonine metabolism | Amino Acid Metabolism | - | Glycine, Serine, Threonine | 1.70 × 10−3 | 0.03 |
| Alanine, aspartate and glutamate metabolism | Amino Acid Metabolism | - | Glutamine, Succinate | 0.02 | 0.22 |
| Metabolite | p-Value | FDR | Trend |
|---|---|---|---|
| PGDM 1 vs. Controls | |||
| HbA1c 2 | 4.28 × 10−12 | 2.27 × 10−10 | PGDM > ctrl |
| Mannose | 3.87 × 10−4 | 0.01 | PGDM > ctrl |
| Glucose | 1.54 × 10−3 | 0.03 | PGDM > ctrl |
| Amino acids | |||
| Histidine | 4.50 × 10−3 | 0.06 | Ctrl > PGDM |
| Tyrosine | 0.01 | 0.10 | Ctrl > PGDM |
| Asparagine | 0.03 | 0.12 | Ctrl > PGDM |
| Glutamine | 0.03 | 0.12 | Ctrl > PGDM |
| Ornithine | 0.04 | 0.12 | Ctrl > PGDM |
| Methionine | 0.05 | 0.12 | Ctrl > PGDM |
| Ketones | |||
| Acetone | 0.01 | 0.11 | PGDM > ctrl |
| Acetoacetate | 0.04 | 0.12 | PGDM > ctrl |
| 3-Hydroxybutyrate | 0.05 | 0.12 | PGDM > ctrl |
| Lipids | |||
| (-CH2)n fatty acids | 0.02 | 0.12 | PGDM > ctrl |
| Alpha-CH2 fatty acids | 0.03 | 0.12 | PGDM > ctrl |
| Glyceryl C1,3H | 0.03 | 0.12 | PGDM > ctrl |
| Beta-CH2 fatty acids | 0.04 | 0.12 | PGDM > ctrl |
| Glyceryl C2H | 0.04 | 0.12 | PGDM > ctrl |
| CH2CH=C fatty acids | 0.04 | 0.12 | PGDM > ctrl |
| N-acetyl-glycoproteins | 0.04 | 0.12 | PGDM > ctrl |
| Other | |||
| Betaine | 0.05 | 0.12 | PGDM > ctrl |
| Creatinine | 9.95 × 10−3 | 0.10 | Ctrl > PGDM |
| GDM 3 vs. Controls | |||
| HbA1c 2 | 3.03 × 10−3 | 0.16 | GDM > Ctrl |
| Mannose | 0.02 | 0.61 | GDM > Ctrl |
| Pathway | Class | Significant Metabolites | p-Value | FDR | |
|---|---|---|---|---|---|
| PGDM 1 vs. Controls | Increased | Decreased | |||
| Synthesis and degradation of ketone bodies | Lipid Metabolism | 3-Hydroxybutyrate, Acetoacetate | - | 4.53 × 10−4 | 0.02 |
| Arginine biosynthesis | Amino Acid Metabolism | - | Ornithine, Glutamine | 3.98 × 10−3 | 0.10 |
| Butanoate metabolism | Carbohydrate Metabolism | 3-Hydroxybutyrate, Acetoacetate | - | 4.57 × 10−3 | 0.10 |
| Galactose metabolism | Carbohydrate Metabolism | Glucose, Mannose | - | 0.01 | 0.19 |
| Alanine, aspartate and glutamate metabolism | Amino Acid Metabolism | - | Asparagine, Glutamine | 0.02 | 0.19 |
| Phenylalanine, tyrosine and tryptophan biosynthesis | Amino Acid Metabolism | - | Tyrosine | 0.03 | 0.30 |
| Tyrosine metabolism | Amino Acid Metabolism | Acetoacetate | Tyrosine | 0.03 | 0.32 |
| Metabolite | p-Value | FDR | Mean Control (SE) | Mean Diabetes (SE) | FC (Diabetes/Control) |
|---|---|---|---|---|---|
| PGDM 1 (n = 6) vs. Controls (n = 22) | |||||
| Mannose | 0.02 | 0.53 | 0.06 (0.003) | 0.08 (0.009) | 0.34 |
| Glucose | 0.02 | 0.53 | 1.86 (0.083) | 2.33 (0.214) | 0.33 |
| Dimethylglycine | 0.05 | 0.59 | 0.02 (0.001) | 0.02 (0.003) | 0.31 |
| GDM 2 (n = 7) vs. Controls (n = 22) | |||||
| Mannose | 3.92 × 10−4 | 0.02 | 0.06 (0.003) | 0.09 (0.005) | 0.44 |
| Dimethylglycine | 0.02 | 0.40 | 0.02 (0.001) | 0.02 (0.003) | 0.39 |
| Glycine | 0.03 | 0.40 | 0.87 (0.034) | 0.72 (0.036) | −0.27 |
| Betaine | 0.03 | 0.40 | 0.26 (0.005) | 0.28 (0.013) | 0.14 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walejko, J.M.; Chelliah, A.; Keller-Wood, M.; Wasserfall, C.; Atkinson, M.; Gregg, A.; Edison, A.S. Diabetes Leads to Alterations in Normal Metabolic Transitions of Pregnancy as Revealed by Time-Course Metabolomics. Metabolites 2020, 10, 350. https://doi.org/10.3390/metabo10090350
Walejko JM, Chelliah A, Keller-Wood M, Wasserfall C, Atkinson M, Gregg A, Edison AS. Diabetes Leads to Alterations in Normal Metabolic Transitions of Pregnancy as Revealed by Time-Course Metabolomics. Metabolites. 2020; 10(9):350. https://doi.org/10.3390/metabo10090350
Chicago/Turabian StyleWalejko, Jacquelyn M., Anushka Chelliah, Maureen Keller-Wood, Clive Wasserfall, Mark Atkinson, Anthony Gregg, and Arthur S. Edison. 2020. "Diabetes Leads to Alterations in Normal Metabolic Transitions of Pregnancy as Revealed by Time-Course Metabolomics" Metabolites 10, no. 9: 350. https://doi.org/10.3390/metabo10090350
APA StyleWalejko, J. M., Chelliah, A., Keller-Wood, M., Wasserfall, C., Atkinson, M., Gregg, A., & Edison, A. S. (2020). Diabetes Leads to Alterations in Normal Metabolic Transitions of Pregnancy as Revealed by Time-Course Metabolomics. Metabolites, 10(9), 350. https://doi.org/10.3390/metabo10090350